Circulating Extracellular Vesicles: The Missing Link between Physical Exercise and Depression Management?

 ,
,  and
and
Abstract
:1. Introduction
2. Search and Selection Criteria to Avoid Confounding Factors

3. Evidence Supporting the Triad Physical Exercise-Extracellular Vesicles-Depression
3.1. Aerobic Exercise and Depression


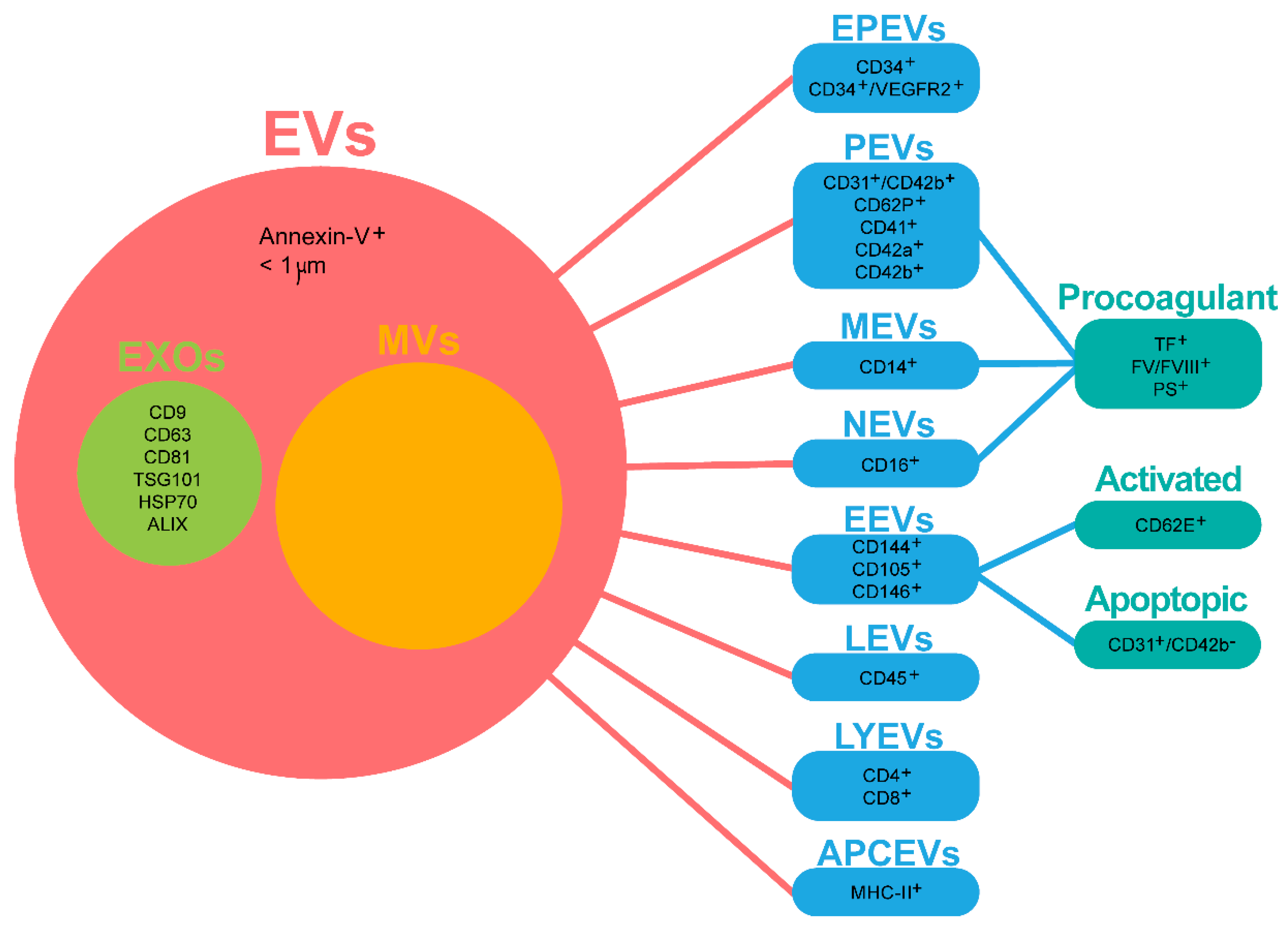{kind=link}
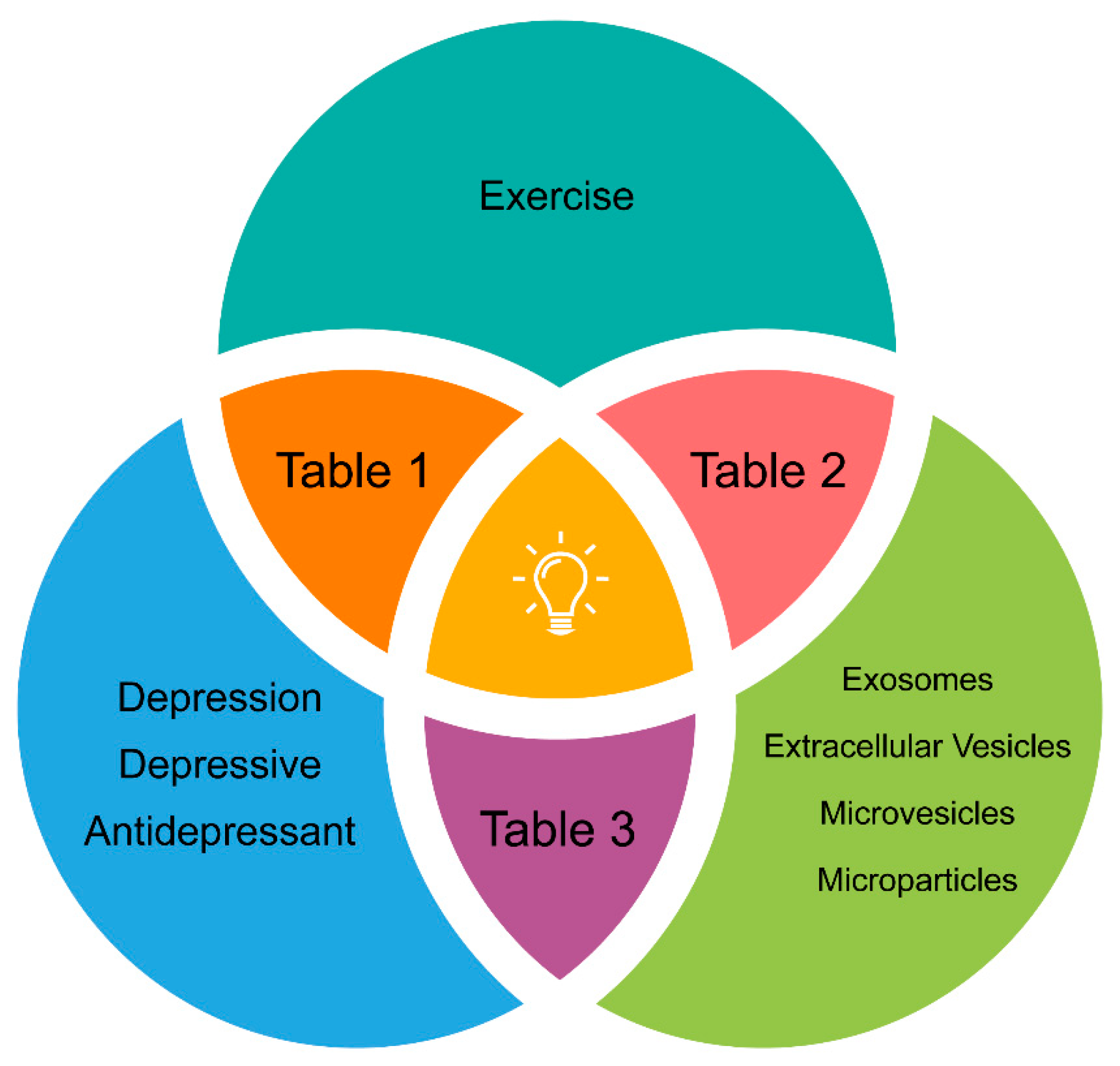{kind=link}
{kind=link}
| Study Year | Sample | Physical Exercise | Severity of Depression | Major Findings | |
|---|---|---|---|---|---|
| Chronic Aerobic Physical Exercise | |||||
| 2019[9] | Sedentary subjects with MDD (22 ± 2 years; n = 11) | Frequency and duration: 3 days/week; 12 weeks (chronic)Intensity: MI incrementalType: cycling, running, elliptical use, rowing or a combinationTreatment groups: 1 (PE) | HAMD17 | ↓ depression scores after PE; time spent in target HRR zone correlated with ↓ anxiety symptoms | |
| 2017[78] | Sedentary university students with non-psychotic MDD (mostly female; 21 ± 2 years; n = 15) | Frequency and duration: 3 days/week; 8 weeks (chronic)Intensity: LI and MI Type: running or cyclingTreatment groups: 2 (LI PE; MI PE) | BDI-II score | ↓ depressive symptoms after MI PE (not LI PE) | |
| 2016[76] | Males with depression (24.5 years; n = 9–11) | Frequency and duration: 3 days/week; 3 weeks (chronic)Intensity: LI, MI and HIType: running or cyclingTreatment groups: 3 (LI PE; MI PE; HI PE) | HAMD and MADRS | ↓ depressive symptoms after both MI and HI PE (not LI PE) | |
| 2015[79] | Adolescents with depression (CDI-2 score > 14, 15 ± 1 years; n = 43–44) | Frequency and duration: 2 days/week; 6 weeks (chronic)Intensity: MI intervalType: circuit training (abdominal, back, ball arm-based exercises, bouncing, static and dynamic balance, squat and cycling)Treatment groups: 2 (PE + TAU; TAU) | CDI-2 score | ↓ depressive symptoms at 6-months follow-up; unchanged depressive symptoms immediately after treatment | |
| 2015[77] | Sedentary subjects with mild to moderate depression (PHQ-9 score > 9, mostly female; 43 ± 12 years; n = 310) | Frequency and duration: 3 days/week; 12 weeks (chronic)Intensity: LI, MI and HIType: yoga class (LI), aerobics (MI and HI)Treatment groups: 3 (PE (LI, MI and HI); ICBT and TAU) | PHQ-9 score and MADRS | ↓ depressive symptoms for PE and ICBT (compared to TAU) | |
| ↓ depression severity for LI PE (compared to TAU and MI PE) and for HI PE (compared to MI PE)4 subgroups (TAU (n = 310), LI PE (n = 106), MI PE (n = 105) and HI PE (n = 99)) [80] | Secondary Analyses (2017) | ||||
| ↓ IL-6 was paralleled by ↓ depression severity after PE1 subgroup (PE (n = 116)) [81] | |||||
| ↑ treatment response to PE for Met allele carriers without exposure to childhood adversity but currently on antidepressants (compared to Val homozygotes); ↑ serum BDNF for Met carriers; BDNF Val66Met or childhood adversity did not predict treatment response5 subgroups (2—BDNF variation Val66Met (Val/Val, n = 377 and Met carriers, n = 170) and 3—childhood adversity (no, mild-to-moderate and high)) [83] | |||||
| Unchanged kynurenine and kynurenic acid plasma levels1 subgroup (PE (n = 117)) [82] | |||||
3.2. Aerobic Exercise and Extracellular Vesicles
| Study Year | Sample | Physical Exercise | Extracellular Vesicles | Major Findings |
|---|---|---|---|---|
| Acute Aerobic Physical Exercise | ||||
| 2019 [92] | Fit females (pre-, peri- and post-menopausal; 45 ± 1 years; n = 11–13) | Frequency and duration: acute Intensity: MI Type: running | Blood collection: pre- and 30 min after PE Isolation and detection: cell-free plasma; FC (calibration beads) Characterization: FC | ↓ EEVs (↓ activated and ↓ apoptotic) after PE; ↑ percentage of activated EEVs (but not apoptotic) after PE; no effect of menopausal status on EEVs |
| 2019 [99] | Male subjects (23 ± 3 years; n = 15) | Frequency and duration: acute (participants completed one exercise and one control trial separated by at least 5 d in a randomized crossover design (www.randomization.com). Intensity: HI Type: running | Blood collection: pre-, 0 and 90 min after PE Isolation and detection: cell-free plasma; ultracentrifugation, FC Characterization: FC | Unchanged EV concentration or diameter (NTA); ↑ EV counts immediately after PE (FC); ↓ procoagulant PEVs/NEVs at 0/90 min after PE |
| 2019 [53] | Male athletes (28 ± 4 years; n = 21) | Frequency and duration: acute Intensity: incremental to exhaustion Type: cycling | Blood collection: pre-, during and 2 min after PE Isolation and detection: cell-free plasma; size-exclusion chromatography, EXOs immuno-bead isolation (CD9+/CD63+/CD81+) Characterization: NTA, FC, proteomics | ↑ EVs during PE (highest levels at peak PE); ↑ EXO markers, ↑ EEVs, ↑ PEVs, ↑ MEVs, LYEVs and ↑ APCEVs after PE; markers of muscle (SGCA) not detected |
| 2018 [49] | Male subjects (27 ± 1 years; n = 11) | Frequency and duration: acute Intensity: incremental to exhaustion Type: cycling | Blood collection: pre-, 0 min and 4 h after PE Isolation and detection: cell-free plasma; ultracentrifugation Characterization: NTA, proteomics | ↑ EVs after PE, returning to basal in 4 h; ↑ 322 proteins after PE (biogenesis and function of EXOs and small EVs and several biological processes, most notably the glycolytic pathway) |
| 2018 [95] | Males and females (24 ± 5 years; n = 20) | Frequency and duration: acute Intensity: MI continuous or HI interval Type: running | Blood collection: pre-, during and after (0, 30, 60, 90 and 120 min) PE Isolation and detection: cell-free plasma; FC (calibration beads) Characterization: FC | Analysis of EVs at all seven times revealed no significant interaction; ↓ activated EEVs immediately after MI continuous PE (not HI interval); sex-differentiated analysis showed ↓ activated EEVs after MI PE only in women; unchanged EPEVs; menstrual cycle phase did not affect EVs |
| 2018 [94] | Marathon runners (49 ± 6 years; n = 99) | Frequency and duration: acute Intensity: marathon running Type: running | Blood collection: pre-, 0 min and 2 d after PE Isolation and detection: cell-free plasma; FC Characterization: FC | ↑ EEVs (↑ activated and ↑ apoptotic) and ↑ PEVs after marathon, returning to basal within 2 days; ↓ MEVs and ↓ LEVs after marathon, remaining ↓; number of participants’ marathon runs in total (but not running time) correlated with ↑ EEVs (activated and apoptotic) |
| 2018 [93] | Male subjects (25 ± 4 years; n = 10) | Frequency and duration: acute Intensity: HI Type: cycling | Blood collection: pre, 5 min and 4 h after PE Isolation and detection: cell-free plasma; size exclusion columns Characterization: WB, qRT-PCR | EXO markers detected (CD63, HSP70); ↑ 12/29 target miRNA after PE (miR-1-3p, -16-5p and 222-3p coincident with muscle and plasma; miR-23a-3p, 208a-3p and -150-5p coincident with muscle; miR- 486-5p, 378a-5p, 126-3p coincident with plasma; and miR- 23b-3p, 451a and 186-5p only in EXOs), associated with myogenic differentiation, fibre identification, blood vessel formation and insulin responsiveness |
| 2016 [55] | Active subjects (25 ± 1 years; n = 18) | Frequency and duration: acute Intensity: HI incremental Type: cycling | Blood collection: pre- and 0 min after PE Isolation and detection: cell-free plasma; FC (<1 μm) Characterization: FC | ↑ activated EEVs in male and ↑ EPEVs in female after PE (unchanged for men and women combined); ↓ EPEVs baseline in female |
| 2015 [91] | Sedentary and fit males (26 ± 5 years; n = 18) | Frequency and duration: acute Intensity: HI Type: running | Collection: pre- and 60 min after PE Isolation and detection: cell-free plasma; ultracentrifugation; density gradient separation Characterization: EXOs isolation kit, FC, WB, qRT-PCR | 5% of EVs were SGCA+ (<1 µm); 60–65% of SGCA+ EVs were CD81+; ↑ miR-206/miR16 in SGCA+ EVs; ↑ miR-206 in TSG101/SGCA+ EVs; ↑ miR-181a-5p after PE; correlation between VO2 max and EVs miR-1, miR-133b, miR-206, miR-499 and miR-181a |
| Chronic Aerobic Physical Exercise | ||||
| 2017 [103] | Sedentary African Americans (53 ± 1 years; n = 10–23) | Frequency and duration: 3 days/week; 6 months (chronic) Intensity: MI incremental Type: walking/jogging, stair stepping, cycling, rowing, arm and elliptical cross-training | Collection: pre- and after PE Protocol Isolation and detection: cell-free plasma; filtration, FC (<1 μm) Characterization: FC | ↓ activated EEVs and unchanged total EEVs after PE |
| 2015 [101] | Sedentary males (23 ± 1 years; n = 20) | Frequency and duration: 5 days/week, 5 weeks (chronic) Intensity: MI and HI interval Type: cycling | Blood collection: pre- and 0 min after HET (100 W under 12% O2 for 30 min); HET was performed 2 d before and 2 d after PE Isolation and detection: cell-free plasma; filtration/FC (annexin-V+, <1 µm) Characterization: FC | MI and LI PE attenuated HET-induced ↑ NEVs |
| 2015 [54] | Sedentary males (21 ± 0.4 years; n = 40) | Frequency and duration: 5 days/week, 5 weeks (chronic) Intensity: MI Type: NE (21% O2) or HE (15% O2) cycling | Blood collection: pre- and 0 min after HET (100 W under 12% O2 for 30 min); HET was performed 2 d before and 2 d after PE Isolation and detection: cell-free plasma; FC (FSC, calibration beads, annexin-V+) Characterization: FC | ↑ MEVs and ↑ procoagulant MEVs after acute HET, supressed (↓) after both NE and HE 5-wk PE protocol (more pronounced for HE) |
3.3. Depression and Extracellular Vesicles
| Study Year | Sample | Severity of Depression | Extracellular Vesicles | Major Findings |
|---|---|---|---|---|
| 2020 [111] | Subjects with MDD (drug-free; 28 ± 1.7 years; n = 33) or Schizophrenia (drug-free; 29 ± 1; n = 36) and HC (29 ± 1; n = 46) | HAMD and MADRS | Blood collection: overnight fasting Isolation and detection: serum; SEC + centrifugation/concentration Characterization: qRT-PCR, miRNA-seq, bioinformatics | 38 blood exosomal miRNAs differently expressed in MDD and HC, 24 were upregulated and 14 downregulated (from 351 miRNAs for differential analysis); axon guidance and development, dendrite, Wnt signaling pathway, neuron-to-neuron synapse and PI3K-Akt signaling pathway were closely related to MDD; ↑ hsa-miR-139-5p top differentially expressed between MDD and HC (specific for MDD, not Schizophrenia); hsa-miR-139-5p and a cluster of 15 miRNAs or 10 miRNAs with excellent performance to differentiate between MDD and schizophrenia (MDD-specific) |
| 2020 [113] | Subjects with MDD in a current major depressive episode (43 ± 2 years; n = 64) and age and sex-matched HC (38 ± 2 years; n = 29) | HDRS-21 | Blood collection: 6 h fasting, 6 h no PE Isolation and detection: plasma; precipitation + ultracentrifugation; WB expression array (CD63, CD81, ALIX, FLOT1, ICAM1, EpCam, ANXA5, and TSG101) Characterization: ExoCET (AChE activity), FC, ELISA | Unchanged EXOs between MDD and HC; ↑ L1CAM+ EXOs in MDD; ↑ IRS-1 in L1CAM+ EXOs in MDD (no sex or psychotropic medication differences); no between-group difference in pSer-IRS-1 in L1CAM+ EXOs (only sex differences in MDD group); IRS-1 in L1CAM+ EXOs correlated with systemic IR for HC but not MDD; no relationship between IRS-1 in L1CAM+ EXOs and depression severity in MDD; the highest IRS-1 in L1CAM+ EXOs correlated with depressed mood, feelings of guilt, suicidality and anhedonia (MDD) |
| 2020 [112] | Subjects with MDD (34 ± 11; n = 30) or BD subjects (46 ± 10; n = 42) and HC (43 ± 6; n = 36) | HAMD and YMRS | Blood collection: no specifications Isolation and detection: serum; centrifugation + filtration (0.22 μm) Characterization: PCR, bioinformatics | EVs Prevotella 2 and Ruminococcaceae UCG-002 genera significantly more prevalent in MDD than BD or HC; apoptosis function differed between groups |
| 2018 [114] | MDD Subjects in acute episode of MDD (drug-free, 31 ± 7; n = 34) and sex and age-matched HC (n = 34) | BDI-II | Blood collection: no specifications Isolation and detection: plasma; sandwich ELISA detection (CD81) Characterization: sandwich ELISA | Neuron-related blood biomarkers moderately to strongly positively correlated with CD81; ↑ IL-34/CD81 in MDD (compared to control); SYP/CD81 and TNFR1/CD81 positively correlated with depression scores |
4. Cautionary Note on Modulating Factors
5. Future Directions: PE–EVs–Depression
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Depression and Other Common Mental Disorders: Global Health Estimates; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Stanton, R.; To, Q.G.; Khalesi, S.; Williams, S.L.; Alley, S.J.; Thwaite, T.L.; Fenning, A.S.; Vandelanotte, C. Depression, Anxiety and Stress during COVID-19: Associations with Changes in Physical Activity, Sleep, Tobacco and Alcohol Use in Australian Adults. Int. J. Environ. Res. Public Health 2020, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Biswas, P.; Ghosh, R.; Chatterjee, S.; Dubey, M.J.; Chatterjee, S.; Lahiri, D.; Lavie, C.J. Psychosocial impact of COVID-19. Diabetes Metab. Syndr. 2020, 14, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Salari, N.; Hosseinian-Far, A.; Jalali, R.; Vaisi-Raygani, A.; Rasoulpoor, S.; Mohammadi, M.; Rasoulpoor, S.; Khaledi-Paveh, B. Prevalence of stress, anxiety, depression among the general population during the COVID-19 pandemic: A systematic review and meta-analysis. Glob. Health 2020, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Otte, C.; Gold, S.M.; Penninx, B.W.; Pariante, C.M.; Etkin, A.; Fava, M.; Mohr, D.C.; Schatzberg, A.F. Major depressive disorder. Nat. Rev. Dis. Primers 2016, 2, 16065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.F.; Castanheira, L.; Sebastião, A.M.; Telles-Correia, D. Depression assessment in clinical trials and pre-clinical tests: A critical review. Curr. Top. Med. Chem. 2018, 18, 1677–1703. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D. Voluntary exercise and depression-like behavior in rodents: Are we running in the right direction? J. Mol. Endocrinol. 2018, 60, R77–R95. [Google Scholar] [CrossRef] [PubMed]
- Penn, E.; Tracy, D.K. The drugs don’t work? antidepressants and the current and future pharmacological management of depression. Ther. Adv. Psychopharmacol. 2012, 2, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, N.; Courtright, A.K.; De Somma, E.; MacQueen, G.M.; MacMaster, F.P. Aerobic exercise in depressed youth: A feasibility and clinical outcomes pilot. Early Interv. Psychiatry 2019, 13, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Morres, I.D.; Hatzigeorgiadis, A.; Stathi, A.; Comoutos, N.; Arpin-Cribbie, C.; Krommidas, C.; Theodorakis, Y. Aerobic exercise for adult patients with major depressive disorder in mental health services: A systematic review and meta-analysis. Depress. Anxiety 2019, 36, 39–53. [Google Scholar] [CrossRef] [Green Version]
- NCCMH. Depression: The Treatment and Management of Depression in Adults; Updated Edition; British Psychological Society: Leicester, UK, 2010. [Google Scholar]
- Belvederi Murri, M.; Ekkekakis, P.; Magagnoli, M.; Zampogna, D.; Cattedra, S.; Capobianco, L.; Serafini, G.; Calcagno, P.; Zanetidou, S.; Amore, M. Physical Exercise in Major Depression: Reducing the Mortality Gap While Improving Clinical Outcomes. Front. Psychiatry 2018, 9, 762. [Google Scholar] [CrossRef] [Green Version]
- Warburton, D.E.; Nicol, C.W.; Bredin, S.S. Health benefits of physical activity: The evidence. CMAJ 2006, 174, 801–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandolesi, L.; Polverino, A.; Montuori, S.; Foti, F.; Ferraioli, G.; Sorrentino, P.; Sorrentino, G. Effects of Physical Exercise on Cognitive Functioning and Wellbeing: Biological and Psychological Benefits. Front. Psychol. 2018, 9, 509. [Google Scholar] [CrossRef] [PubMed]
- Guthold, R.; Stevens, G.A.; Riley, L.M.; Bull, F.C. Worldwide trends in insufficient physical activity from 2001 to 2016: A pooled analysis of 358 population-based surveys with 1· 9 million participants. Lancet Glob. Health 2018, 6, e1077–e1086. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Recommendations on Physical Activity for Health; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Gelenberg, A.J.; Freeman, M.P.; Markowitz, J.C.; Rosenbaum, J.F.; Thase, M.E.; Trivedi, M.H.; Van Rhoads, R.S.; Reus, V.I.; DePaulo, J.R., Jr.; Fawcett, J.A. Practice guideline for the treatment of patients with major depressive disorder third edition. Am. J. Psychiatry 2010, 167, 1. [Google Scholar]
- Patel, H.; Alkhawam, H.; Madanieh, R.; Shah, N.; Kosmas, C.E.; Vittorio, T.J. Aerobic vs anaerobic exercise training effects on the cardiovascular system. World J. Cardiol. 2017, 9, 134–138. [Google Scholar] [CrossRef]
- Kvam, S.; Kleppe, C.L.; Nordhus, I.H.; Hovland, A. Exercise as a treatment for depression: A meta-analysis. J. Affect. Disord. 2016, 202, 67–86. [Google Scholar] [CrossRef]
- Grazina, R.; Massano, J. Physical exercise and Parkinson’s disease: Influence on symptoms, disease course and prevention. Rev. Neurosci. 2013, 24, 139–152. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, T.; Chu, J.M.; Chen, Y.; Dunnett, S.; Ho, Y.S.; Wong, G.T.; Chang, R.C. The beneficial effects of physical exercise in the brain and related pathophysiological mechanisms in neurodegenerative diseases. Lab. Investig. 2019, 99, 943–957. [Google Scholar] [CrossRef]
- Radak, Z.; Hart, N.; Sarga, L.; Koltai, E.; Atalay, M.; Ohno, H.; Boldogh, I. Exercise plays a preventive role against Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2010, 20, 777–783. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, P.; Stritt, C.; Bloch, W.; Schmidt, F.P.; Hubner, S.T.; Binnebossel, S.; Schenk, A.; Oberste, M. The effects of different aerobic exercise intensities on serum serotonin concentrations and their association with Stroop task performance: A randomized controlled trial. Eur. J. Appl. Physiol. 2016, 116, 2025–2034. [Google Scholar] [CrossRef]
- Kerling, A.; Kuck, M.; Tegtbur, U.; Grams, L.; Weber-Spickschen, S.; Hanke, A.; Stubbs, B.; Kahl, K.G. Exercise increases serum brain-derived neurotrophic factor in patients with major depressive disorder. J. Affect. Disord. 2017, 215, 152–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, A.M.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, T.C.; Marques-Aleixo, I.; Beleza, J.; Oliveira, P.J.; Ascensao, A.; Magalhaes, J. Physical Exercise and Brain Mitochondrial Fitness: The Possible Role Against Alzheimer’s Disease. Brain Pathol. 2016, 26, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Safdar, A.; Tarnopolsky, M.A. Exosomes as Mediators of the Systemic Adaptations to Endurance Exercise. Cold Spring Harb. Perspect. Med. 2018, 8, a029827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, J.M.; Kroemer, G.; Zitvogel, L. Extracellular vesicles: Masters of intercellular communication and potential clinical interventions. J. Clin. Investig. 2016, 126, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Tkach, M.; Thery, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [Green Version]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neuro. Oncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small Gtpases 2017, 8, 220–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugholm, L.H.; Revenfeld, A.L.; Sondergaard, E.K.; Jorgensen, M.M. Antibody-Based Assays for Phenotyping of Extracellular Vesicles. Biomed Res. Int. 2015, 2015, 524817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, N.; Cvjetkovic, A.; Jang, S.C.; Crescitelli, R.; Hosseinpour Feizi, M.A.; Nieuwland, R.; Lotvall, J.; Lasser, C. Detailed analysis of the plasma extracellular vesicle proteome after separation from lipoproteins. Cell. Mol. Life Sci. Cmls 2018, 75, 2873–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Jiang, D.; Li, W.; Xiang, X.; Zhao, J.; Yu, B.; Wang, C.; He, Z.; Zhu, L.; Yang, Y. Evaluation of serum extracellular vesicles as noninvasive diagnostic markers of glioma. Theranostics 2019, 9, 5347–5358. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cauvi, D.M.; Bernardino, E.M.A.; Lara, B.; Lizardo, R.E.; Hawisher, D.; Bickler, S.; De Maio, A. Isolation and characterization of human urine extracellular vesicles. Cell Stress Chaperones 2018, 23, 943–953. [Google Scholar] [CrossRef]
- Iwai, K.; Yamamoto, S.; Yoshida, M.; Shiba, K. Isolation of Extracellular Vesicles in Saliva Using Density Gradient Ultracentrifugation. Methods Mol. Biol. 2017, 1660, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Foers, A.D.; Chatfield, S.; Dagley, L.F.; Scicluna, B.J.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Enrichment of extracellular vesicles from human synovial fluid using size exclusion chromatography. J. Extracell. Vesicles 2018, 7, 1490145. [Google Scholar] [CrossRef]
- Chiasserini, D.; van Weering, J.R.; Piersma, S.R.; Pham, T.V.; Malekzadeh, A.; Teunissen, C.E.; de Wit, H.; Jimenez, C.R. Proteomic analysis of cerebrospinal fluid extracellular vesicles: A comprehensive dataset. J. Proteom. 2014, 106, 191–204. [Google Scholar] [CrossRef]
- Ebert, B.; Rai, A.J. Isolation and Characterization of Amniotic Fluid-Derived Extracellular Vesicles for Biomarker Discovery. Methods Mol. Biol. 2019, 1885, 287–294. [Google Scholar] [CrossRef]
- Zonneveld, M.I.; Brisson, A.R.; van Herwijnen, M.J.; Tan, S.; van de Lest, C.H.; Redegeld, F.A.; Garssen, J.; Wauben, M.H.; Nolte-’t Hoen, E.N. Recovery of extracellular vesicles from human breast milk is influenced by sample collection and vesicle isolation procedures. J. Extracell. Vesicles 2014, 3, 24215. [Google Scholar] [CrossRef] [Green Version]
- Tavakolizadeh, J.; Roshanaei, K.; Salmaninejad, A.; Yari, R.; Nahand, J.S.; Sarkarizi, H.K.; Mousavi, S.M.; Salarinia, R.; Rahmati, M.; Mousavi, S.F.; et al. MicroRNAs and exosomes in depression: Potential diagnostic biomarkers. J. Cell. Biochem. 2018, 119, 3783–3797. [Google Scholar] [CrossRef] [PubMed]
- Gruzdev, S.K.; Yakovlev, A.A.; Druzhkova, T.A.; Guekht, A.B.; Gulyaeva, N.V. The Missing Link: How Exosomes and miRNAs can Help in Bridging Psychiatry and Molecular Biology in the Context of Depression, Bipolar Disorder and Schizophrenia. Cell. Mol. Neurobiol. 2019, 39, 729–750. [Google Scholar] [CrossRef]
- Ilgin, C.; Topuzoglu, A. Extracellular Vesicles in Psychiatry Research in the Context of RDoC Criteria. Psychiatry Investig. 2018, 15, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Strawbridge, R.; Young, A.H.; Cleare, A.J. Biomarkers for depression: Recent insights, current challenges and future prospects. Neuropsychiatr. Dis. Treat. 2017, 13, 1245–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unutzer, J.; Park, M. Strategies to improve the management of depression in primary care. Prim. Care 2012, 39, 415–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.E.; Egan, C.L.; Cron, L.; Watt, K.I.; Kuchel, R.P.; et al. Extracellular Vesicles Provide a Means for Tissue Crosstalk during Exercise. Cell Metab. 2018, 27, 237–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Merwe, Y.; Steketee, M.B. Extracellular Vesicles: Biomarkers, Therapeutics, and Vehicles in the Visual System. Curr. Ophthalmol. Rep. 2017, 5, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, E.N.; Mourot, L.; Rakobowchuk, M. Exercise-Derived Microvesicles: A Review of the Literature. Sports Med. 2018, 48, 2025–2039. [Google Scholar] [CrossRef] [Green Version]
- Highton, P.J.; Martin, N.; Smith, A.C.; Burton, J.O.; Bishop, N.C. Microparticles and Exercise in Clinical Populations. Exerc. Immunol. Rev. 2018, 24, 46–58. [Google Scholar]
- Brahmer, A.; Neuberger, E.; Esch-Heisser, L.; Haller, N.; Jorgensen, M.M.; Baek, R.; Mobius, W.; Simon, P.; Kramer-Albers, E.M. Platelets, endothelial cells and leukocytes contribute to the exercise-triggered release of extracellular vesicles into the circulation. J. Extracell. Vesicles 2019, 8, 1615820. [Google Scholar] [CrossRef]
- Wang, J.S.; Chang, Y.L.; Chen, Y.C.; Tsai, H.H.; Fu, T.C. Effects of normoxic and hypoxic exercise regimens on monocyte-mediated thrombin generation in sedentary men. Clin. Sci. 2015, 129, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Lansford, K.A.; Shill, D.D.; Dicks, A.B.; Marshburn, M.P.; Southern, W.M.; Jenkins, N.T. Effect of acute exercise on circulating angiogenic cell and microparticle populations. Exp. Physiol. 2016, 101, 155–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. Biomed Res. Int. 2018, 2018, 8545347. [Google Scholar] [CrossRef]
- Hartjes, T.A.; Mytnyk, S.; Jenster, G.W.; van Steijn, V.; van Royen, M.E. Extracellular Vesicle Quantification and Characterization: Common Methods and Emerging Approaches. Bioengineering 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Thery, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Soekmadji, C.; Hill, A.F.; Wauben, M.H.; Buzas, E.I.; Di Vizio, D.; Falcon-Perez, J.M.; Gardiner, C.; Hochberg, F.; Kurochkin, I.V.; et al. Updating the MISEV minimal requirements for extracellular vesicle studies: Building bridges to reproducibility. J. Extracell. Vesicles 2017, 6, 1396823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.; Kang, B.; Kim, O.Y.; Choi, D.S.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An integrated database of high-throughput data for systemic analyses of extracellular vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef] [PubMed]
- Gezsi, A.; Kovacs, A.; Visnovitz, T.; Buzas, E.I. Systems biology approaches to investigating the roles of extracellular vesicles in human diseases. Exp. Mol. Med. 2019, 51, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trovato, E.; Di Felice, V.; Barone, R. Extracellular Vesicles: Delivery Vehicles of Myokines. Front. Physiol. 2019, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Cabral, D.F.; Rice, J.; Morris, T.P.; Rundek, T.; Pascual-Leone, A.; Gomes-Osman, J. Exercise for Brain Health: An Investigation into the Underlying Mechanisms Guided by Dose. Neurother. J. Am. Soc. Exp. Neurother. 2019, 16, 580–599. [Google Scholar] [CrossRef]
- Ignacio, Z.M.; da Silva, R.S.; Plissari, M.E.; Quevedo, J.; Reus, G.Z. Physical Exercise and Neuroinflammation in Major Depressive Disorder. Mol. Neurobiol. 2019, 56, 8323–8335. [Google Scholar] [CrossRef] [PubMed]
- Mee-Inta, O.; Zhao, Z.W.; Kuo, Y.M. Physical Exercise Inhibits Inflammation and Microglial Activation. Cells 2019, 8, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2019, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.F.; Paes, F.; Arias-Carrion, O.; Ferreira Rocha, N.B.; de Sa Filho, A.S.; Machado, S. Neural Mechanisms of Exercise: Anti-Depression, Neurogenesis, and Serotonin Signaling. CNS Neurol. Disord. Drug Targets 2015, 14, 1307–1311. [Google Scholar] [CrossRef]
- Lee, M.C.; Byun, K.; Kim, J.S.; Lee, H.; Kim, K. Trends in exercise neuroscience: Raising demand for brain fitness. J. Exerc. Rehabil. 2019, 15, 176–179. [Google Scholar] [CrossRef]
- Pareja-Galeano, H.; Mayero, S.; Perales, M.; Garatachea, N.; Santos-Lozano, A.; Fiuza-Luces, C.; Emanuele, E.; Galvez, B.G.; Sanchis-Gomar, F.; Lucia, A. Biological Rationale for Regular Physical Exercise as an Effective Intervention for the Prevention and Treatment of Depressive Disorders. Curr. Pharm. Des. 2016, 22, 3764–3775. [Google Scholar] [CrossRef]
- Gujral, S.; Aizenstein, H.; Reynolds, C.F., 3rd; Butters, M.A.; Erickson, K.I. Exercise effects on depression: Possible neural mechanisms. Gen. Hosp. Psychiatry 2017, 49, 2–10. [Google Scholar] [CrossRef]
- Loprinzi, P.D. The effects of physical exercise on parahippocampal function. Physiol. Int. 2019, 106, 114–127. [Google Scholar] [CrossRef]
- Li, A.; Yau, S.Y.; Machado, S.; Wang, P.; Yuan, T.F.; So, K.F. Enhancement of Hippocampal Plasticity by Physical Exercise as a Polypill for Stress and Depression: A Review. Cns Neurol. Disord. Drug Targets 2019, 18, 294–306. [Google Scholar] [CrossRef]
- Bolijn, S.; Lucassen, P.J. How the Body Talks to the Brain; Peripheral Mediators of Physical Activity-Induced Proliferation in the Adult Hippocampus. Brain Plast. 2015, 1, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Han, C.; Wang, J.; Zhou, J.; Liang, C.; Ranganna, K.; Song, Y.H. Exosomes Mediate the Beneficial Effects of Exercise. Adv. Exp. Med. Biol. 2017, 1000, 333–353. [Google Scholar] [CrossRef] [PubMed]
- Al Shweiki, M.R.; Oeckl, P.; Steinacker, P.; Hengerer, B.; Schonfeldt-Lecuona, C.; Otto, M. Major depressive disorder: Insight into candidate cerebrospinal fluid protein biomarkers from proteomics studies. Expert Rev. Proteom. 2017, 14, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Balchin, R.; Linde, J.; Blackhurst, D.; Rauch, H.L.; Schonbachler, G. Sweating away depression? The impact of intensive exercise on depression. J. Affect. Disord. 2016, 200, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, M.; Kraepelien, M.; Ojehagen, A.; Lindefors, N.; Zeebari, Z.; Kaldo, V.; Forsell, Y. Physical exercise and internet-based cognitive-behavioural therapy in the treatment of depression: Randomised controlled trial. Br. J. Psychiatry 2015, 207, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, R.L.; Brush, C.J.; Ehmann, P.J.; Alderman, B.L. A randomized trial of aerobic exercise on cognitive control in major depression. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2017, 128, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Carter, T.; Guo, B.; Turner, D.; Morres, I.; Khalil, E.; Brighton, E.; Armstrong, M.; Callaghan, P. Preferred intensity exercise for adolescents receiving treatment for depression: A pragmatic randomised controlled trial. BMC Psychiatry 2015, 15, 247. [Google Scholar] [CrossRef] [Green Version]
- Helgadottir, B.; Forsell, Y.; Hallgren, M.; Moller, J.; Ekblom, O. Long-term effects of exercise at different intensity levels on depression: A randomized controlled trial. Prev. Med. 2017, 105, 37–46. [Google Scholar] [CrossRef]
- Lavebratt, C.; Herring, M.P.; Liu, J.J.; Wei, Y.B.; Bossoli, D.; Hallgren, M.; Forsell, Y. Interleukin-6 and depressive symptom severity in response to physical exercise. Psychiatry Res. 2017, 252, 270–276. [Google Scholar] [CrossRef]
- Millischer, V.; Erhardt, S.; Ekblom, O.; Forsell, Y.; Lavebratt, C. Twelve-week physical exercise does not have a long-lasting effect on kynurenines in plasma of depressed patients. Neuropsychiatr. Dis. Treat. 2017, 13, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Millischer, V.; Zeebari, Z.; Forsell, Y.; Lavebratt, C. BDNF Val66Met and childhood adversity on response to physical exercise and internet-based cognitive behavioural therapy in depressed Swedish adults. J. Psychiatr. Res. 2017, 93, 50–58. [Google Scholar] [CrossRef]
- De Oliveira Bristot, V.J.; de Bem Alves, A.C.; Cardoso, L.R.; da Luz Scheffer, D.; Aguiar, A.S., Jr. The Role of PGC-1alpha/UCP2 Signaling in the Beneficial Effects of Physical Exercise on the Brain. Front. Neurosci. 2019, 13, 292. [Google Scholar] [CrossRef] [Green Version]
- De Assis, G.G.; Gasanov, E.V.; de Sousa, M.B.C.; Kozacz, A.; Murawska-Cialowicz, E. Brain derived neutrophic factor, a link of aerobic metabolism to neuroplasticity. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2018, 69, 351–358. [Google Scholar] [CrossRef]
- Verhagen, M.; van der Meij, A.; van Deurzen, P.A.; Janzing, J.G.; Arias-Vasquez, A.; Buitelaar, J.K.; Franke, B. Meta-analysis of the BDNF Val66Met polymorphism in major depressive disorder: Effects of gender and ethnicity. Mol. Psychiatry 2010, 15, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, S.; Nesse, R.M.; Stoltenberg, S.F.; Li, S.; Gleiberman, L.; Chakravarti, A.; Weder, A.B.; Burmeister, M. A BDNF coding variant is associated with the NEO personality inventory domain neuroticism, a risk factor for depression. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2003, 28, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Mata, J.; Thompson, R.J.; Gotlib, I.H. BDNF genotype moderates the relation between physical activity and depressive symptoms. Health Psychol. Off. J. Div. Health Psychol. Am. Psychol. Assoc. 2010, 29, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Eichner, N.Z.M.; Erdbrugger, U.; Malin, S.K. Extracellular Vesicles: A Novel Target for Exercise-Mediated Reductions in Type 2 Diabetes and Cardiovascular Disease Risk. J. Diabetes Res. 2018, 2018, 7807245. [Google Scholar] [CrossRef] [Green Version]
- Guescini, M.; Canonico, B.; Lucertini, F.; Maggio, S.; Annibalini, G.; Barbieri, E.; Luchetti, F.; Papa, S.; Stocchi, V. Muscle Releases Alpha-Sarcoglycan Positive Extracellular Vesicles Carrying miRNAs in the Bloodstream. PLoS ONE 2015, 10, e0125094. [Google Scholar] [CrossRef] [Green Version]
- Serviente, C.; Burnside, A.; Witkowski, S. Moderate-intensity exercise reduces activated and apoptotic endothelial microparticles in healthy midlife women. J. Appl. Physiol. 2019, 126, 102–110. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, R.F.; Woodhead, J.S.T.; Zeng, N.; Blenkiron, C.; Merry, T.L.; Cameron-Smith, D.; Mitchell, C.J. Circulatory exosomal miRNA following intense exercise is unrelated to muscle and plasma miRNA abundances. Am. J. Physiology. Endocrinol. Metab. 2018, 315, E723–E733. [Google Scholar] [CrossRef]
- Schwarz, V.; Dusing, P.; Liman, T.; Werner, C.; Herm, J.; Bachelier, K.; Krull, M.; Brechtel, L.; Jungehulsing, G.J.; Haverkamp, W.; et al. Marathon running increases circulating endothelial- and thrombocyte-derived microparticles. Eur. J. Prev. Cardiol. 2018, 25, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Shill, D.D.; Lansford, K.A.; Hempel, H.K.; Call, J.A.; Murrow, J.R.; Jenkins, N.T. Effect of exercise intensity on circulating microparticles in men and women. Exp. Physiol. 2018, 103, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Brooks, G.A. Respiratory gas-exchange ratios during graded exercise in fed and fasted trained and untrained men. J. Appl. Physiol. 1999, 86, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeis, C.; Helmig, S.; Tug, S.; Simon, P.; Kramer-Albers, E.M. Physical exercise induces rapid release of small extracellular vesicles into the circulation. J. Extracell. Vesicles 2015, 4, 28239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, E.N.; Gonzalez-Alonso, J.; Parris, C.; Rakobowchuk, M. Exercise intensity modulates the appearance of circulating microvesicles with proangiogenic potential upon endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1297–H1310. [Google Scholar] [CrossRef] [PubMed]
- Highton, P.J.; Goltz, F.R.; Martin, N.; Stensel, D.J.; Thackray, A.E.; Bishop, N.C. Microparticle Responses to Aerobic Exercise and Meal Consumption in Healthy Men. Med. Sci. Sports Exerc. 2019, 51, 1935–1943. [Google Scholar] [CrossRef]
- Leroyer, A.S.; Anfosso, F.; Lacroix, R.; Sabatier, F.; Simoncini, S.; Njock, S.M.; Jourde, N.; Brunet, P.; Camoin-Jau, L.; Sampol, J.; et al. Endothelial-derived microparticles: Biological conveyors at the crossroad of inflammation, thrombosis and angiogenesis. Thromb. Haemost. 2010, 104, 456–463. [Google Scholar] [CrossRef]
- Chen, Y.C.; Ho, C.W.; Tsai, H.H.; Wang, J.S. Interval and continuous exercise regimens suppress neutrophil-derived microparticle formation and neutrophil-promoted thrombin generation under hypoxic stress. Clin. Sci. 2015, 128, 425–436. [Google Scholar] [CrossRef]
- Rautou, P.E.; Vion, A.C.; Amabile, N.; Chironi, G.; Simon, A.; Tedgui, A.; Boulanger, C.M. Microparticles, vascular function, and atherothrombosis. Circ. Res. 2011, 109, 593–606. [Google Scholar] [CrossRef]
- Kretzschmar, J.; Babbitt, D.M.; Diaz, K.M.; Feairheller, D.L.; Sturgeon, K.M.; Perkins-Ball, A.M.; Williamson, S.T.; Ling, C.; Grimm, H.; Brown, M.D. Differential Response to Exercise in African Americans with High Levels of Inflammation. Ethn. Dis. 2017, 27, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Baggish, A.L.; Hale, A.; Weiner, R.B.; Lewis, G.D.; Systrom, D.; Wang, F.; Wang, T.J.; Chan, S.Y. Dynamic regulation of circulating microRNA during acute exhaustive exercise and sustained aerobic exercise training. J. Physiol. 2011, 589, 3983–3994. [Google Scholar] [CrossRef] [PubMed]
- Bye, A.; Rosjo, H.; Aspenes, S.T.; Condorelli, G.; Omland, T.; Wisloff, U. Circulating microRNAs and aerobic fitness—The HUNT-Study. PLoS ONE 2013, 8, e57496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, S.; Israel, S.; Nagy, C.; Turecki, G. The emerging role of exosomes in mental disorders. Transl. Psychiatry 2019, 9, 122. [Google Scholar] [CrossRef]
- Matsumoto, J.; Stewart, T.; Banks, W.A.; Zhang, J. The Transport Mechanism of Extracellular Vesicles at the Blood-Brain Barrier. Curr. Pharm. Des. 2017, 23, 6206–6214. [Google Scholar] [CrossRef]
- Andras, I.E.; Toborek, M. Extracellular vesicles of the blood-brain barrier. Tissue Barriers 2016, 4, e1131804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroto-Escuela, D.O.; Agnati, L.F.; Bechter, K.; Jansson, A.; Tarakanov, A.O.; Fuxe, K. The role of transmitter diffusion and flow versus extracellular vesicles in volume transmission in the brain neural-glial networks. Philos. Trans. R. Soc. Lond. Ser. Biol. Sci. 2015, 370, 20140183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapogiannis, D.; Mustapic, M.; Shardell, M.D.; Berkowitz, S.T.; Diehl, T.C.; Spangler, R.D.; Tran, J.; Lazaropoulos, M.P.; Chawla, S.; Gulyani, S.; et al. Association of Extracellular Vesicle Biomarkers With Alzheimer Disease in the Baltimore Longitudinal Study of Aging. JAMA Neurol. 2019, 76, 1340–1351. [Google Scholar] [CrossRef]
- Wei, Z.X.; Xie, G.J.; Mao, X.; Zou, X.P.; Liao, Y.J.; Liu, Q.S.; Wang, H.; Cheng, Y. Exosomes from patients with major depression cause depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2020, 45, 1050–1058. [Google Scholar] [CrossRef]
- Rhee, S.J.; Kim, H.; Lee, Y.; Lee, H.J.; Park, C.H.K.; Yang, J.; Kim, Y.K.; Kym, S.; Ahn, Y.M. Comparison of serum microbiome composition in bipolar and major depressive disorders. J. Psychiatr. Res. 2020, 123, 31–38. [Google Scholar] [CrossRef]
- Nasca, C.; Dobbin, J.; Bigio, B.; Watson, K.; de Angelis, P.; Kautz, M.; Cochran, A.; Mathe, A.A.; Kocsis, J.H.; Lee, F.S.; et al. Insulin receptor substrate in brain-enriched exosomes in subjects with major depression: On the path of creation of biosignatures of central insulin resistance. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- Kuwano, N.; Kato, T.A.; Mitsuhashi, M.; Sato-Kasai, M.; Shimokawa, N.; Hayakawa, K.; Ohgidani, M.; Sagata, N.; Kubo, H.; Sakurai, T.; et al. Neuron-related blood inflammatory markers as an objective evaluation tool for major depressive disorder: An exploratory pilot case-control study. J. Affect. Disord. 2018, 240, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Reviews. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Denman, S.E.; Morrison, M.; Yu, Z.; Dore, J.; Leclerc, M.; McSweeney, C.S. Dysbiosis of fecal microbiota in Crohn’s disease patients as revealed by a custom phylogenetic microarray. Inflamm. Bowel Dis. 2010, 16, 2034–2042. [Google Scholar] [CrossRef]
- Moretti, M.; Fraga, D.B.; Rodrigues, A.L.S. Ascorbic Acid to Manage Psychiatric Disorders. CNS Drugs 2017, 31, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciobanu, L.G.; Sachdev, P.S.; Trollor, J.N.; Reppermund, S.; Thalamuthu, A.; Mather, K.A.; Cohen-Woods, S.; Stacey, D.; Toben, C.; Schubert, K.O.; et al. Co-expression network analysis of peripheral blood transcriptome identifies dysregulated protein processing in endoplasmic reticulum and immune response in recurrent MDD in older adults. J. Psychiatr. Res. 2018, 107, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Molina, C.; Sandoval, M.; Henzi, R.; Ramirez, J.P.; Varas-Godoy, M.; Luarte, A.; Lafourcade, C.A.; Lopez-Verrilli, A.; Smalla, K.H.; Kaehne, T.; et al. Small Extracellular Vesicles in Rat Serum Contain Astrocyte-Derived Protein Biomarkers of Repetitive Stress. Int. J. Neuropsychopharmacol. 2019, 22, 232–246. [Google Scholar] [CrossRef]
- Neven, K.Y.; Nawrot, T.S.; Bollati, V. Extracellular Vesicles: How the External and Internal Environment Can Shape Cell-To-Cell Communication. Curr. Environ. Health Rep. 2017, 4, 30–37. [Google Scholar] [CrossRef]
- Harrison, M.; Murphy, R.P.; O’Connor, P.L.; O’Gorman, D.J.; McCaffrey, N.; Cummins, P.M.; Moyna, N.M. The endothelial microparticle response to a high fat meal is not attenuated by prior exercise. Eur. J. Appl. Physiol. 2009, 106, 555–562. [Google Scholar] [CrossRef]
- Wilhelm, E.N.; Gonzalez-Alonso, J.; Chiesa, S.T.; Trangmar, S.J.; Kalsi, K.K.; Rakobowchuk, M. Whole-body heat stress and exercise stimulate the appearance of platelet microvesicles in plasma with limited influence of vascular shear stress. Physiol. Rep. 2017, 5, e13496. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, E.; Reis, J.; Rodrigues, M.; Ribeiro, C.F.; Pereira, F.C. Circulating Extracellular Vesicles: The Missing Link between Physical Exercise and Depression Management? Int. J. Mol. Sci. 2021, 22, 542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020542
Soares E, Reis J, Rodrigues M, Ribeiro CF, Pereira FC. Circulating Extracellular Vesicles: The Missing Link between Physical Exercise and Depression Management? International Journal of Molecular Sciences. 2021; 22(2):542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020542
Chicago/Turabian StyleSoares, Edna, Julie Reis, Mariana Rodrigues, Carlos Fontes Ribeiro, and Frederico C. Pereira. 2021. "Circulating Extracellular Vesicles: The Missing Link between Physical Exercise and Depression Management?" International Journal of Molecular Sciences 22, no. 2: 542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020542