Cellular Redox State Acts as Switch to Determine the Direction of NNT-Catalyzed Reaction in Cystic Fibrosis Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
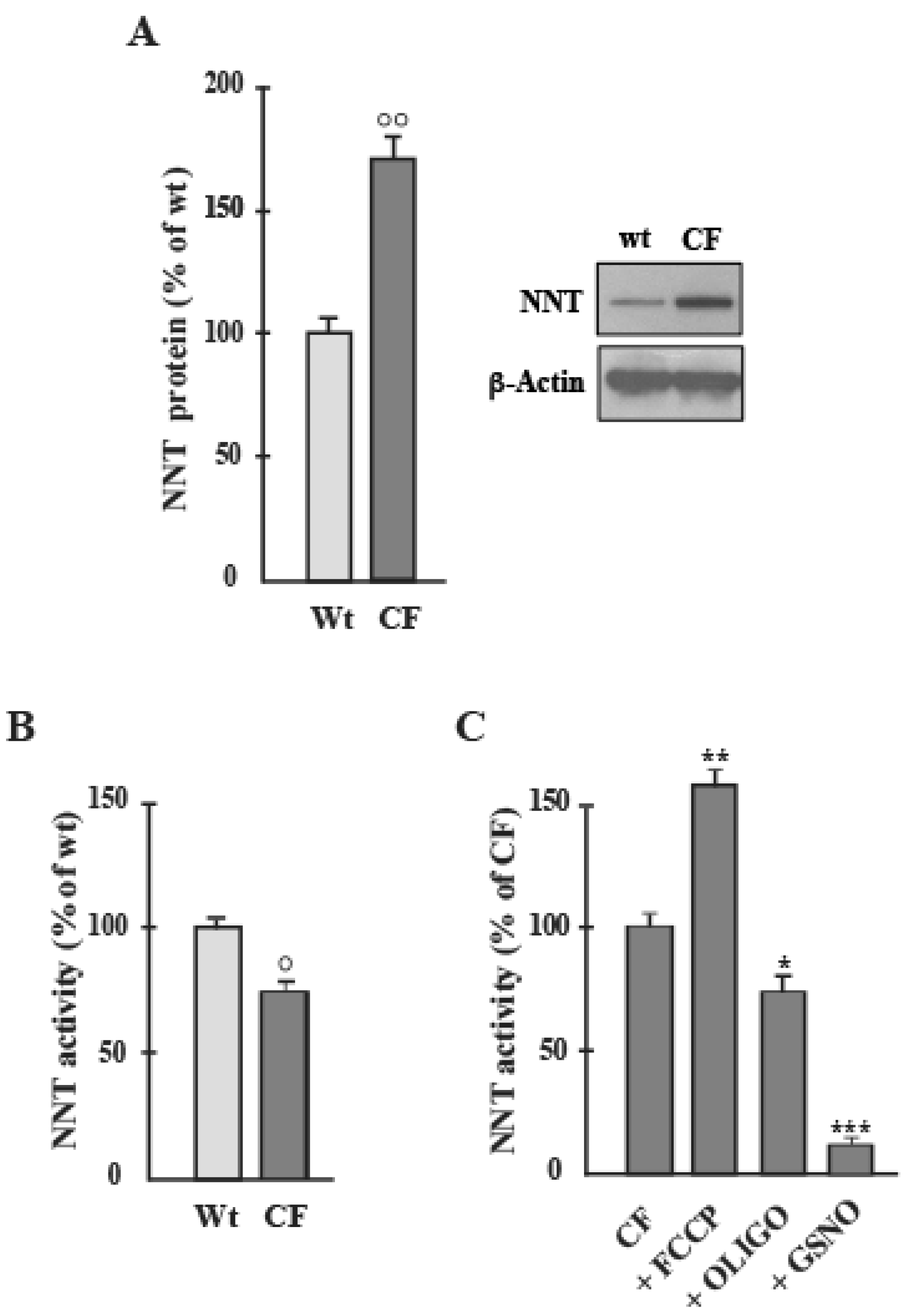
2.1. NNT Expression and Activity in CF Airway Cells
- (i)
- first, lower level of NADPH and higher level of NADH were confirmed in CF than in Wt cells (not shown), as in [25];
- (ii)
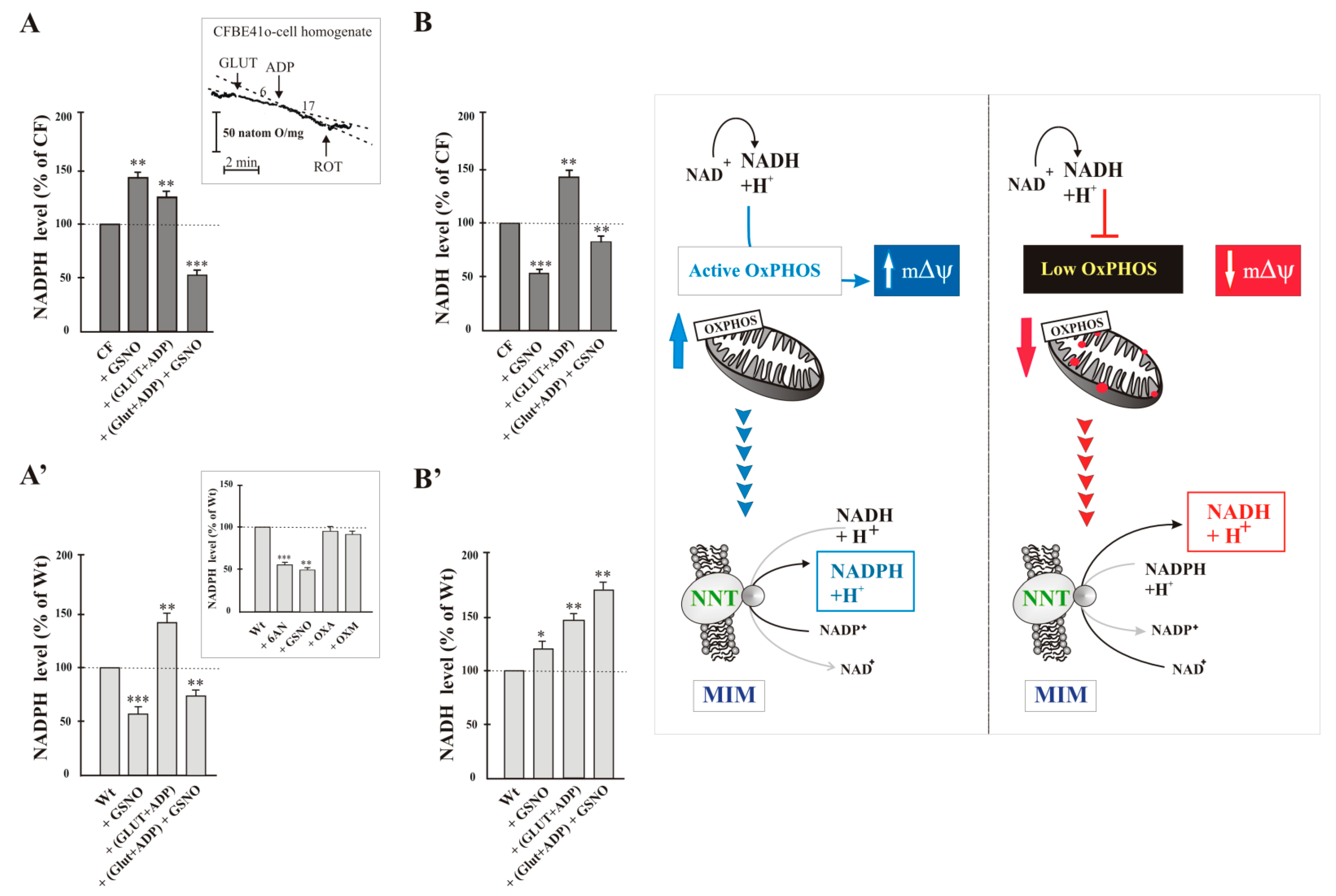
- the cellular enzymes most involved in the production of NADPH are the cytosolic enzyme G6PDH and the mitochondrial NNT; neither Malic enzyme, nor isocitrate dehydrogenase contributed substantially to the production of NADPH in the cell. To test this, the inhibitors of NADP-dependent enzymes were added separately to cells and after 10 min NADPH level was assayed and compared to that of control cells treated with vehicle only. NADPH level did not change significantly in Wt cells treated with oxalate (OXA, 1 mM), a specific inhibitor of malic enzyme [26], or oxalomalate (OXM, 3 mM), a competitive inhibitor of NADP+-dependent isocitrate dehydrogenase [27], but it was markedly reduced by 6-aminonicotinamide (6AN, 5 mM, by approximately 50%), compound that inhibit G6PDH [24], and by GSNO (1 mM, by approximately 45%) (see inset to Figure 2A’).
2.2. Interdependence of NADPH-Linked Enzymes, i.e., G6PDH and NNT
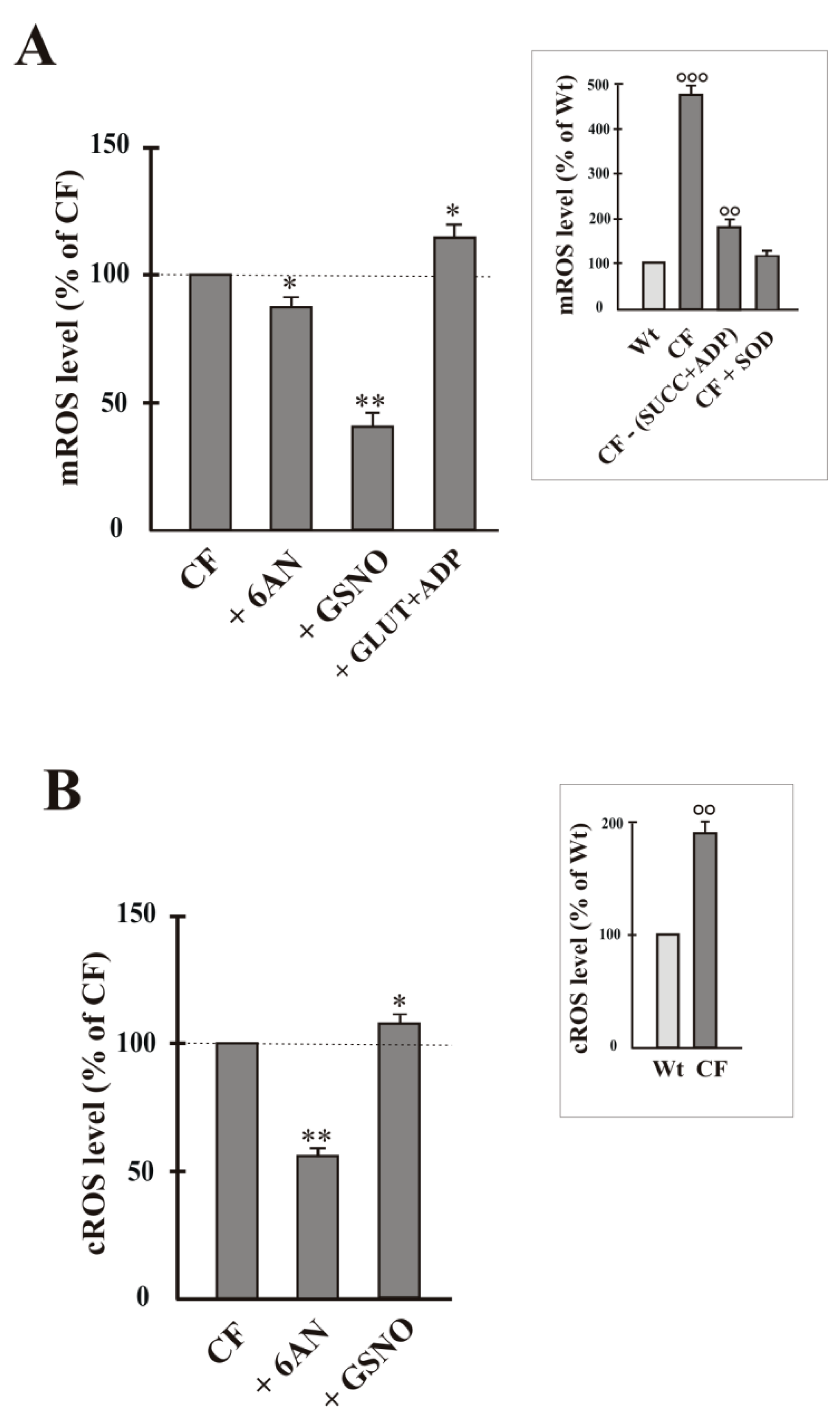
2.3. NNT Activity Regulates Mitochondria-Dependent ROS Production
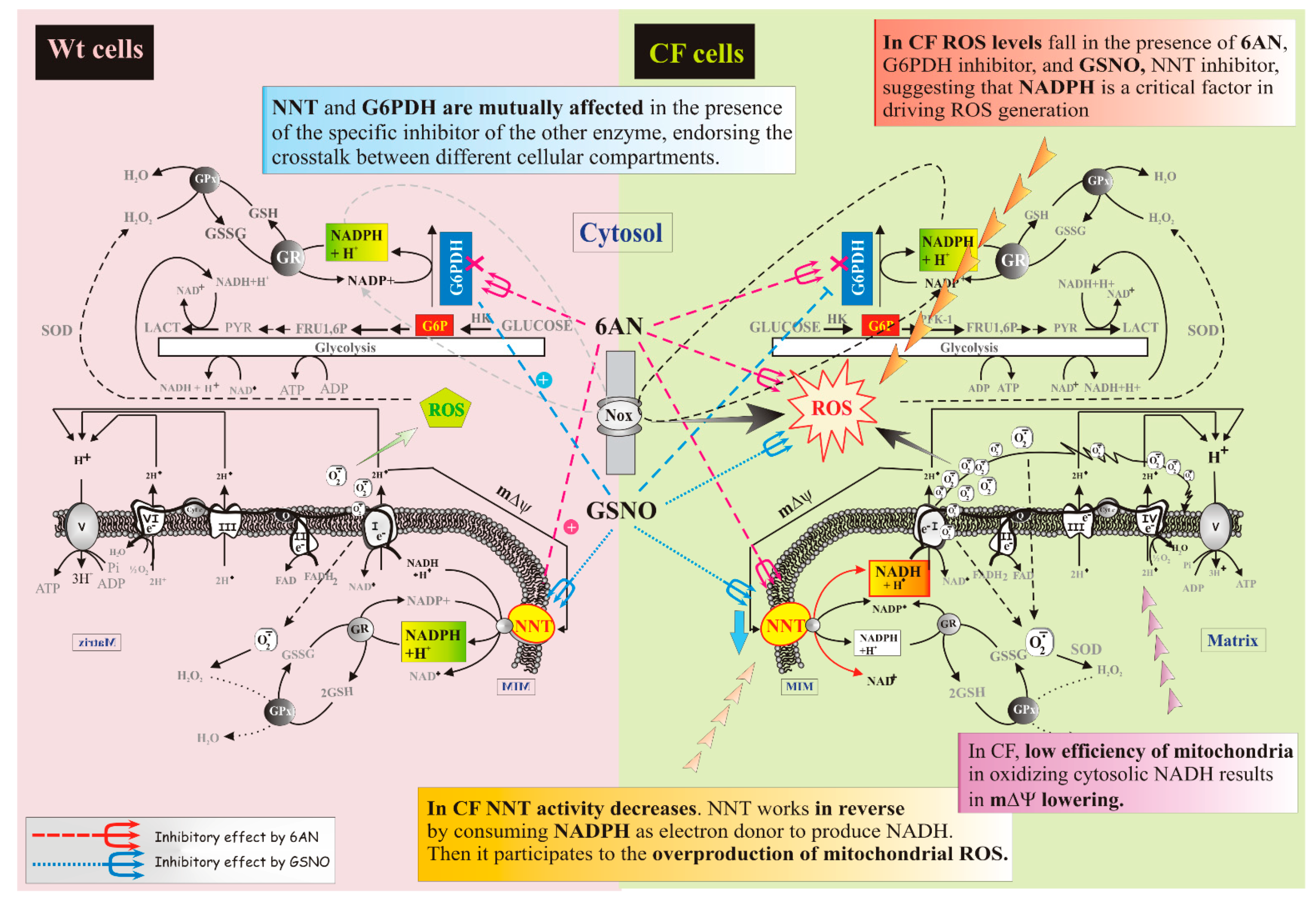
3. Discussion
3.1. How NNT Works in CF Cells
3.2. Crosstalk between the Cytosolic and Mitochondrial NADPH Pools
3.3. How ROS Levels Respond to NNT Loss
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Cell Suspension and Homogenate Preparation
4.4. Protein Extraction and Western Blotting
4.5. Enzyme Assays
4.6. Polarographic Measurements
4.7. Assessment of Cellular and Mitochondrial ROS Production
4.8. NADPH and NADH Measurements
4.9. Statistical Analysis and Computing
5. Conclusions
- (i)
- the cellular redox state, together with the low mΔΨ, pushes to drive NNT reverse reaction, at the expense of its antioxidant potential (Figure 2A,B);
- (ii)
- working in reverse and under poor mitochondrial functionality conditions, the reduced activity of NNT prevents the NADH, produced by the reaction, from causing an explosion of ROS by the damaged respiratory chain (Figure 2A and Figure 4A) [22,24], as instead observed in the presence of (GLUT + ADP) (Figure 4A);
- (iii)
- NNT activity reduction in the presence of 6AN, a G6PDH inhibitor, (Figure 3) endorses the crosstalk between different cellular compartments;
- (iv)
- the greatly reduced level of mitochondrial ROS in NNT-loss cells suggests that the reverse NNT activity contributes substantially to their production (Figure 4A).
- (v)
- The data are schematically reproduced in the picture (Figure 5). In particular, the mutual affection of NNT and G6PDH in the presence of the specific inhibitor of the other enzyme is clearly visible in the Figure: it represents a special condition supporting crosstalk between different cellular compartments.
Author Contributions
Funding
Conflicts of Interest
References
- Blackera, T.S.; Duchena, M.R. Investigating mitochondrial redox state using NADH and NADPH autofluorescence. Free Radic Biol. Med. 2016, 100, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, W.; Wang, R.-S.; Handy, D.E.; Loscalzo, J. NAD(H) and NADP(H) Redox Couples and Cellular Energy Metabolism. Antioxid. Redox Signal. 2018, 28, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.P.; Calvo, S.E.; Mootha, V.K. Spatiotemporal compartmentalization of hepatic NADH and NADPH metabolism. J. Biol. Chem. 2018, 293, 7508–7516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handy, D.E.; Loscalzo, J. Redox Regulation of Mitochondrial Function. Antioxid. Redox Signal. 2012, 16, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- Fessel, J.P.; Oldham, W.M. Pyridine Dinucleotides from Molecules to Man. Antioxid. Redox Signal. 2017, 28, 180–212. [Google Scholar] [CrossRef] [PubMed]
- Sheeran, F.L.; Rydström, J.; Shakhparonov, M.I.; Pestov, N.B.; Pepe, S. Diminished NADPH transhydrogenase activity and mitochondrial redox regulation in human failing myocardium. Biochim. Biophys. Acta 2010, 1797, 1138–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, T.J.S.; Kuczynski, C.; Sezgin, E.; Zhu, C.-T.; Kumagai, S.; Eanes, W.F. Quantifying Interactions Within the NADP(H) Enzyme Network in Drosophila melanogaster. Genetics 2009, 182, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Du, W.; Wu, M. Regulation of the pentose phosphate pathway in cancer. Protein Cell 2014, 5, 592–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhang, Z.; Hoshino, A.; Zheng, H.D.; Morley, M.; Arany, Z.; Rabinowitz, J.D. NADPH production by the oxidative pentose-phosphate pathway supports folate metabolism. Nat. Metab. 2019, 1, 404–415. [Google Scholar] [CrossRef]
- Pollak, N.; Niere, M.; Ziegler, M. NAD kinase levels control the NADPH concentration in human cells. J. Biol. Chem. 2007, 282, 33562–33571. [Google Scholar] [CrossRef] [Green Version]
- Nikiforov, A.; Dölle, C.; Niere, M.; Ziegler, M. Pathways and subcellular compartmentation of NAD biosynthesis in human cells: From entry of extracellular precursors to mitochondrial NAD generation. J. Biol. Chem. 2011, 286, 21767–21778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.A.; Parker, S.J.; Fiske, B.P.; McCloskey, D.; Gui, D.Y.; Green, K.R.; Vokes, N.I.; Feist, A.M.; Vander Heiden, M.G.; Metallo, C.M. Tracing Compartmentalized NADPH Metabolism in the Cytosol and Mitochondria of Mammalian Cells. Mol. Cell 2014, 55, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampjut, D.; Sazanov, L.A. Structure and mechanism of mitochondrial proton-translocating transhydrogenase. Nature. 2019, 573, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, J.A.; Figueira, T.R.; Ravagnani, F.G.; Oliveira, H.C.F.; Vercesi, A.E.; Castilho, R.F. A spontaneous mutation in the nicotinamide nucleotide transhydrogenase gene of C57BL/6J mice results in mitochondrial redox abnormalities. Free Radic. Biol. Med. 2013, 63, 446–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mailloux, R.J. Mitochondrial Antioxidants and the Maintenance of Cellular Hydrogen Peroxide Levels. Oxidative Med. Cell Longev. 2018, 2018, 7857251. [Google Scholar] [CrossRef] [PubMed]
- Rydström, J. Mitochondrial NADPH, transhydrogenase and disease. Biochim. Biophys. Acta 2006, 1757, 721–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickel, A.G.; von Hardenberg, A.; Hohl, M.; Löffler, J.R.; Kohlhaas, M.; Becker, J.; Reil, J.C.; Kazakov, A.; Bonnekoh, J.; Stadelmaier, M.; et al. Reversal of mitochondrial transhydrogenase causes oxidative stress in heart failure. Cell Metab. 2015, 22, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Ronchi, J.A.; Francisco, A.; Passos, L.A.; Figueira, T.R.; Castilho, R.F. The contribution of nicotinamide nucleotide transhydrogenase to peroxide detoxification is dependent on the respiratory state and counterbalanced by other sources of NADPH in liver mitochondria. J. Biol. Chem. 2016, 291, 20173–20187. [Google Scholar] [CrossRef] [Green Version]
- Stein, L.R.; Imai, S.-I. The dynamic regulation of NAD metabolism in mitochondria. Trends Endocrinol. Metab. 2012, 23, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. Redox Modulation by Reversal of the Mitochondrial Nicotinamide Nucleotide Transhydrogenase. Cell Metab. 2015, 22, 363–365. [Google Scholar] [CrossRef] [Green Version]
- Valdivieso, A.G.; Santa-Coloma, T.A. CFTR activity and mitochondrial function. Redox Biol. 2013, 1, 190–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atlante, A.; Favia, M.; Bobba, A.; Guerra, L.; Casavola, V.; Reshkin, S.J. Characterization of mitochondrial function in cells with impaired cystic fibrosis transmembrane conductance regulator (CFTR) function. J. Bioenerg. Biomembr. 2016, 48, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Favia, M.; de Bari, L.; Bobba, A.; Atlante, A. An Intriguing Involvement of Mitochondria in Cystic Fibrosis. J. Clin. Med. 2019, 8, 1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favia, M.; de Bari, L.; Lassandro, R.; Atlante, A. Modulation of glucose-related metabolic pathways controls glucose level in airway surface liquid and fight oxidative stress in cystic fibrosis cells. J. Bioenerg. Biomembr. 2019, 51, 203–218. [Google Scholar] [CrossRef] [PubMed]
- de Bari, L.; Favia, M.; Bobba, A.; Lassandro, R.; Guerra, L.; Atlante, A. Aberrant GSH reductase and NOX activities concur with defective CFTR to pro-oxidative imbalance in cystic fibrosis airways. J. Bioenerg. Biomembr. 2018, 50, 117–1129. [Google Scholar] [CrossRef] [PubMed]
- Pon, J.; Napoli, E.; Luckhart, S.; Giulivi, C. Mitochondrial NAD+-dependent malic enzyme from Anopheles stephensi: A possible novel target for malaria mosquito control. Malar. J. 2011, 10, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-H.; Park, J.-W. Oxalomalate, a competitive inhibitor of NADP+-dependent isocitrate dehydrogenase, enhances lipid peroxidation-mediated oxidative damage in U937 cells. Arch. Biochem. Biophys. 2003, 416, 31–37. [Google Scholar] [CrossRef]
- Li, S.; Zhuang, Z.; Wu, T.; Lin, J.-C.; Liu, Z.-X.; Zhou, L.-F.; Dai, T.; Lu, L.; Ju, H.-Q. Nicotinamide nucleotide transhydrogenase-mediated redox homeostasis promotes tumor growth and metastasis in gastric cancer. Redox Biol. 2018, 18, 246–255. [Google Scholar] [CrossRef]
- Meimaridou, E.; Kowalczyk, J.; Guasti, L.; Hughes, C.R.; Wagner, F.; Frommolt, P.; Nürnberg, P.; Mann, N.P.; Banerjee, R.; Saka, H.N.; et al. Mutations in NNT encoding nicotinamide nucleotide transhydrogenase cause familial glucocorticoid deficiency. Nat. Genet. 2012, 44, 740–742. [Google Scholar] [CrossRef] [Green Version]
- Aon, M.; Cortassa, S.; Maack, C.; O’Rourke, B. Sequential opening of mitochondrial ion channels as a function of glutathione redox thiol status. J. Biol. Chem. 2007, 282, 21889–21900. [Google Scholar] [CrossRef] [Green Version]
- Mullen, A.R.; Hu, Z.; Shi, X.; Jiang, L.; Boroughs, L.K.; Kovacs, Z.; Boriack, R.; Rakheja, D.; Sullivan, L.B.; Linehan, W.M.; et al. Oxidation of alpha-ketoglutarate is required for reductive carboxylation in cancer cells with mitochondrial defects. Cell Rep. 2014, 7, 1679–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbattiscianni, A.C.; Favia, M.; Mancini, M.T.; Cardone, R.A.; Guerra, L.; Monterisi, S.; Castellani, S.; Laselva, O.; Di Sole, F.; Conese, M.; et al. Correctors of mutant CFTR enhance subcortical cAMP-PKA signaling through modulating ezrin phosphorylation and cytoskeleton organization. J. Cell Sci. 2016, 129, 1128–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, K.M.; Janes, M.S.; Pehar, M.; Monette, J.S.; Ross, M.F.; Hagen, T.M.; Murphy, M.P.; Beckman, J.S. Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc. Natl. Acad. Sci. USA 2006, 103, 15038–15043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibon, Y.; Larher, F. Cycling assay for nicotinamide adenine dinucleotides: NaCl precipitation and ethanol solubilisation of the reduced tetrazolium. Anal. Biochem. 1997, 251, 153–157. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favia, M.; Atlante, A. Cellular Redox State Acts as Switch to Determine the Direction of NNT-Catalyzed Reaction in Cystic Fibrosis Cells. Int. J. Mol. Sci. 2021, 22, 967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020967
Favia M, Atlante A. Cellular Redox State Acts as Switch to Determine the Direction of NNT-Catalyzed Reaction in Cystic Fibrosis Cells. International Journal of Molecular Sciences. 2021; 22(2):967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020967
Chicago/Turabian StyleFavia, Maria, and Anna Atlante. 2021. "Cellular Redox State Acts as Switch to Determine the Direction of NNT-Catalyzed Reaction in Cystic Fibrosis Cells" International Journal of Molecular Sciences 22, no. 2: 967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020967