
Protein Engineering Approaches to Enhance Fungal Laccase Production in S. cerevisiae

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
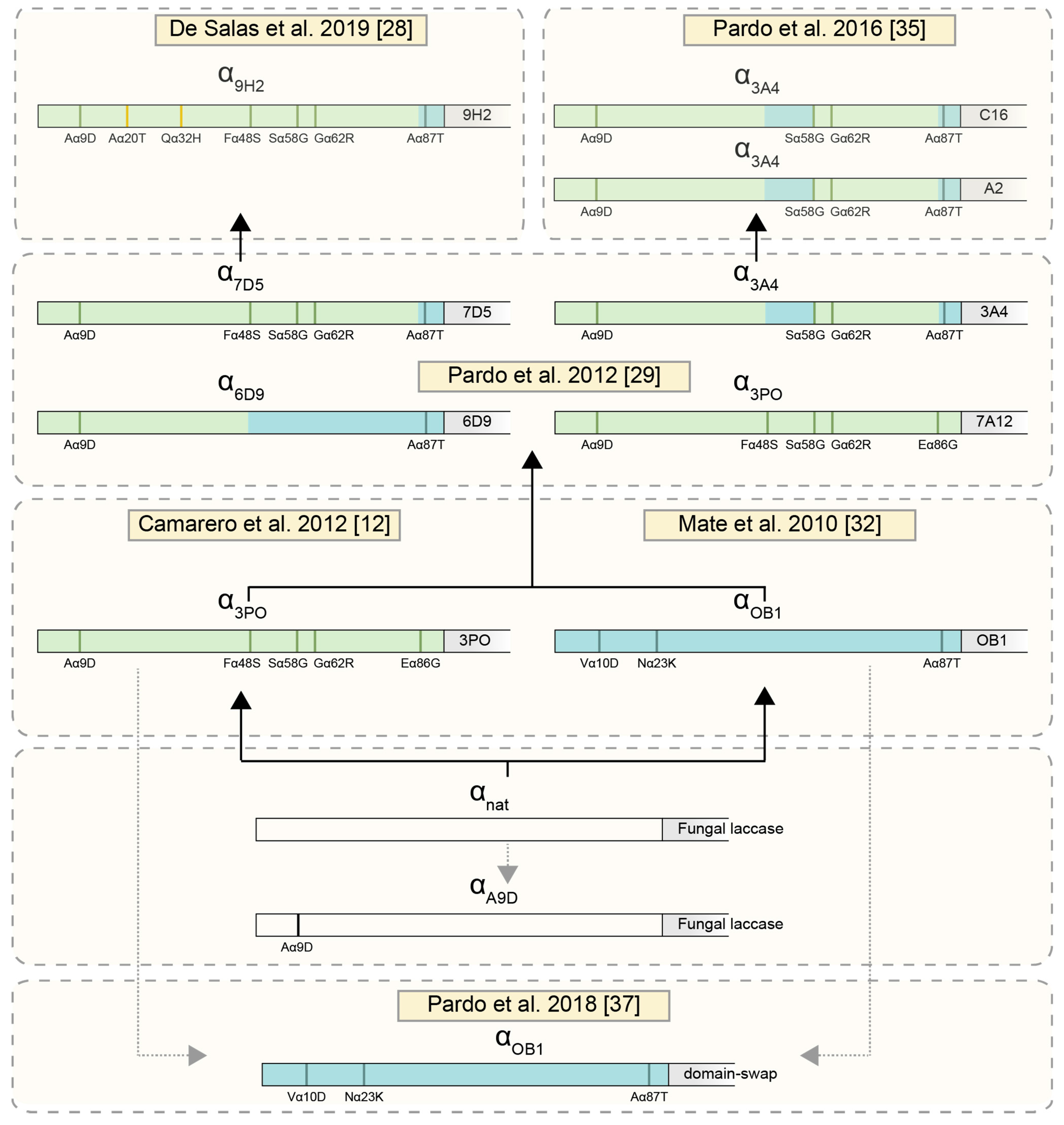
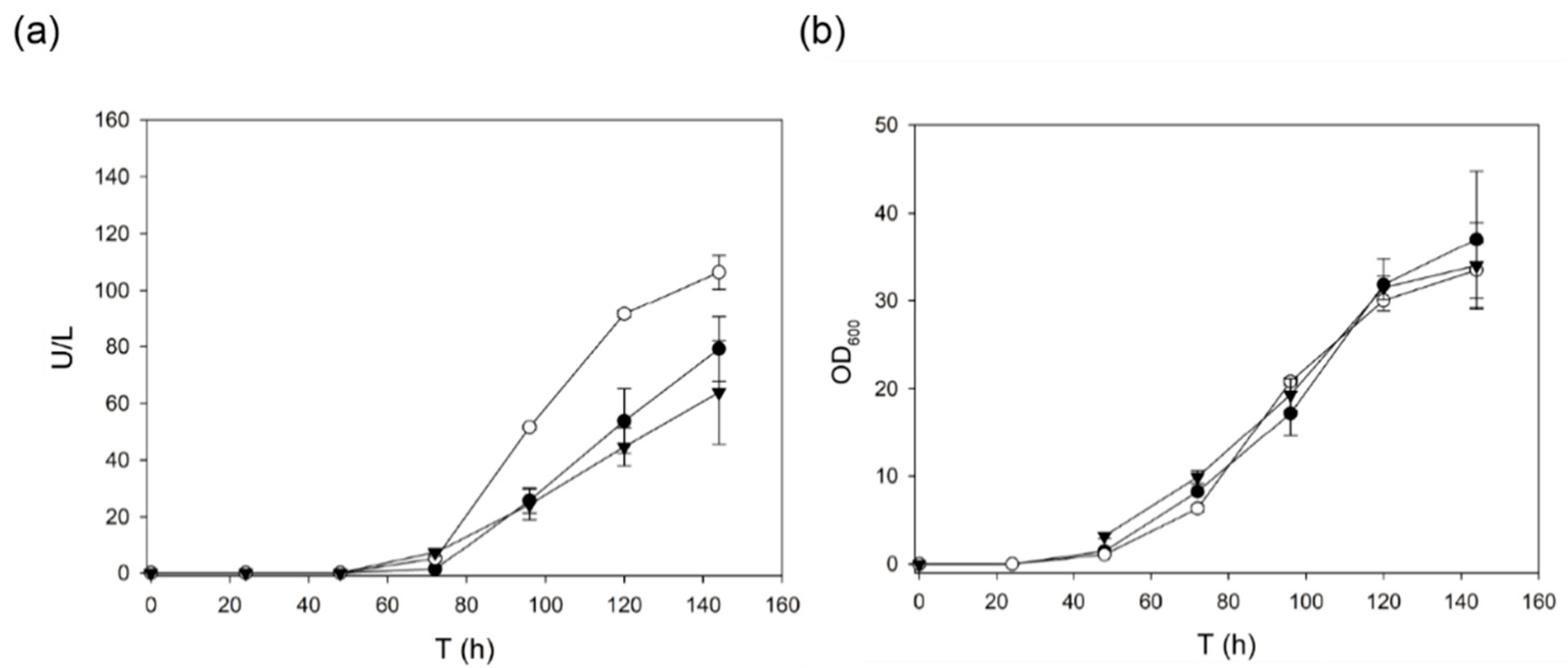
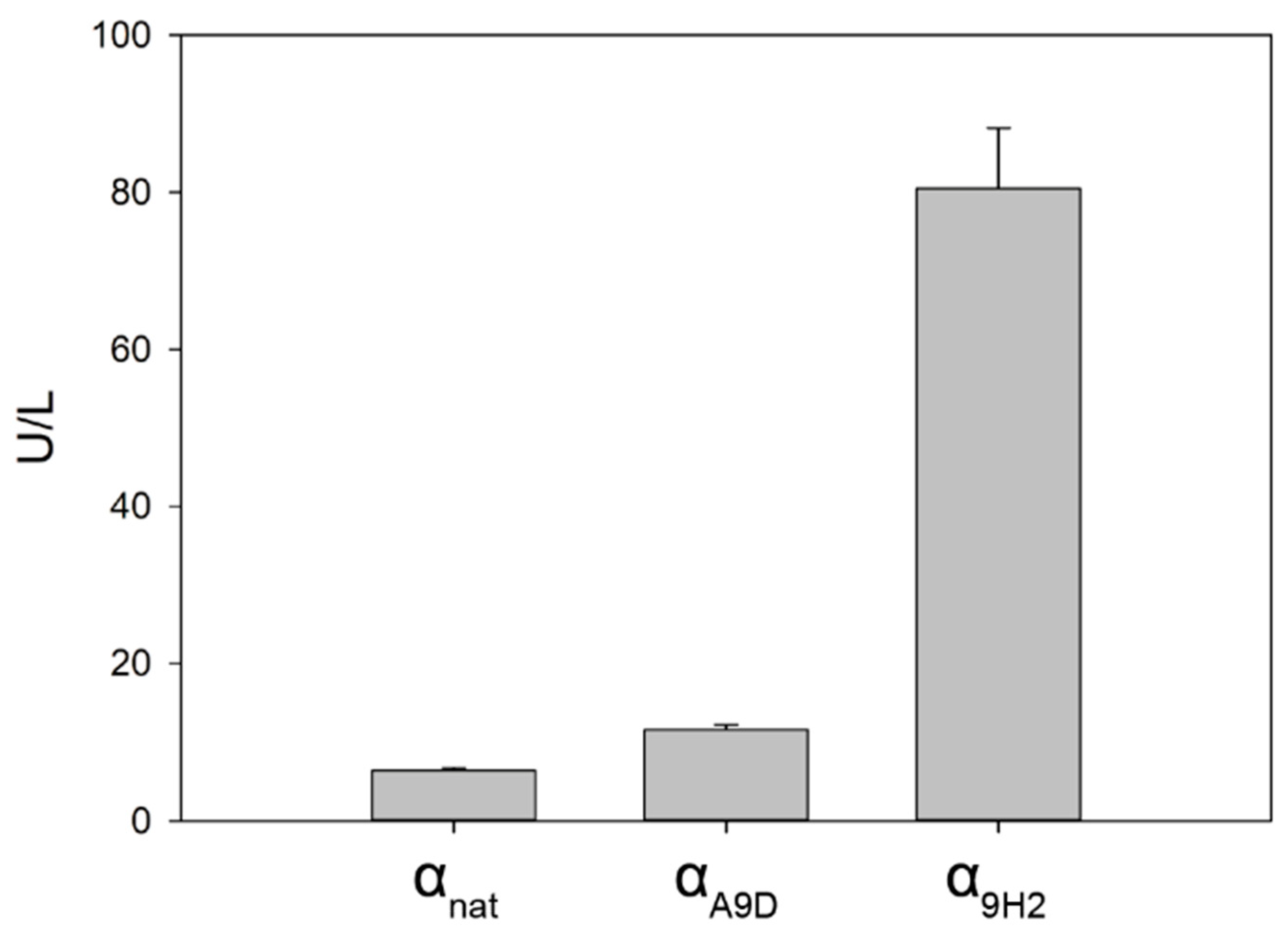
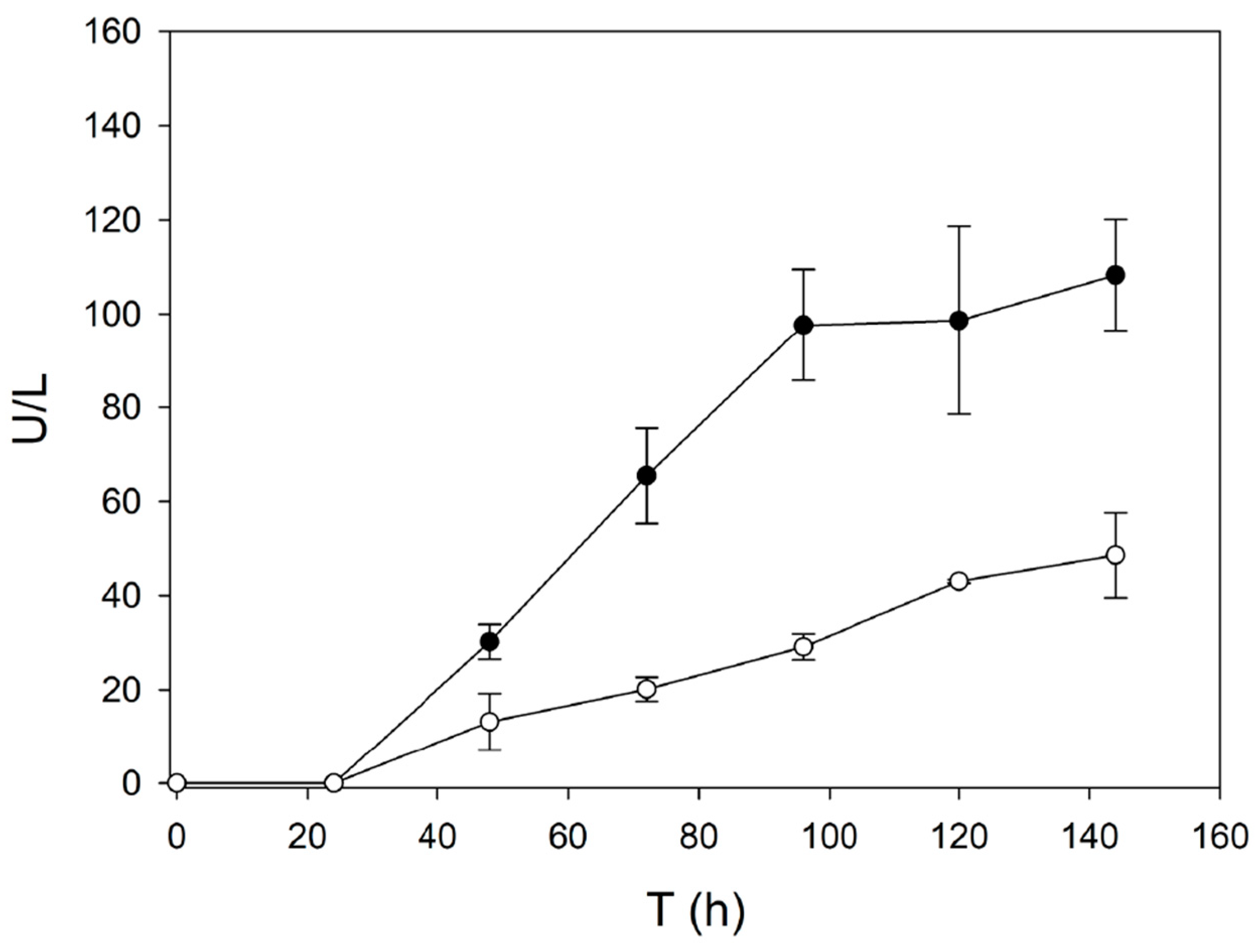
2.1. Use of Evolved α-Factor Preproleader Sequences to Raise Laccase Secretion
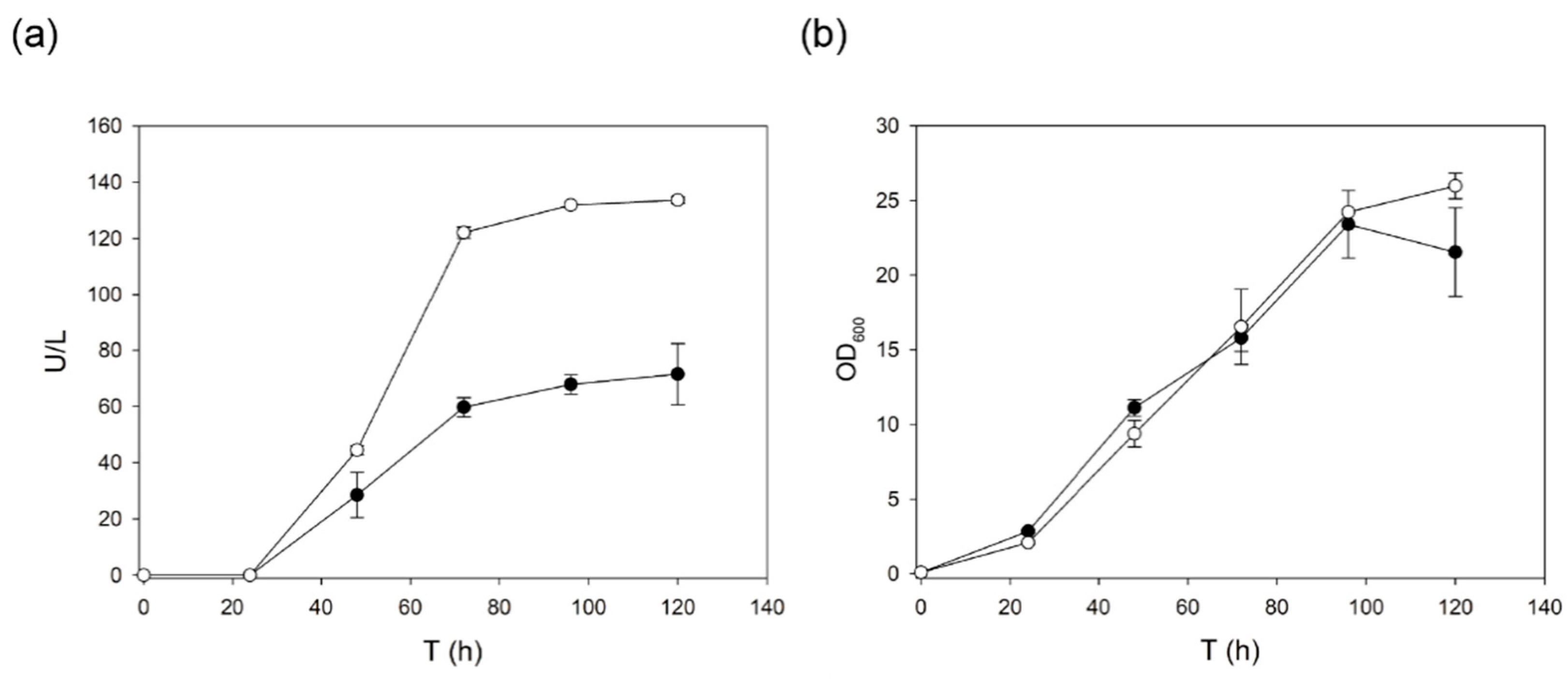
2.2. Engineering of New N-Glycosylation Sites in the Enzyme
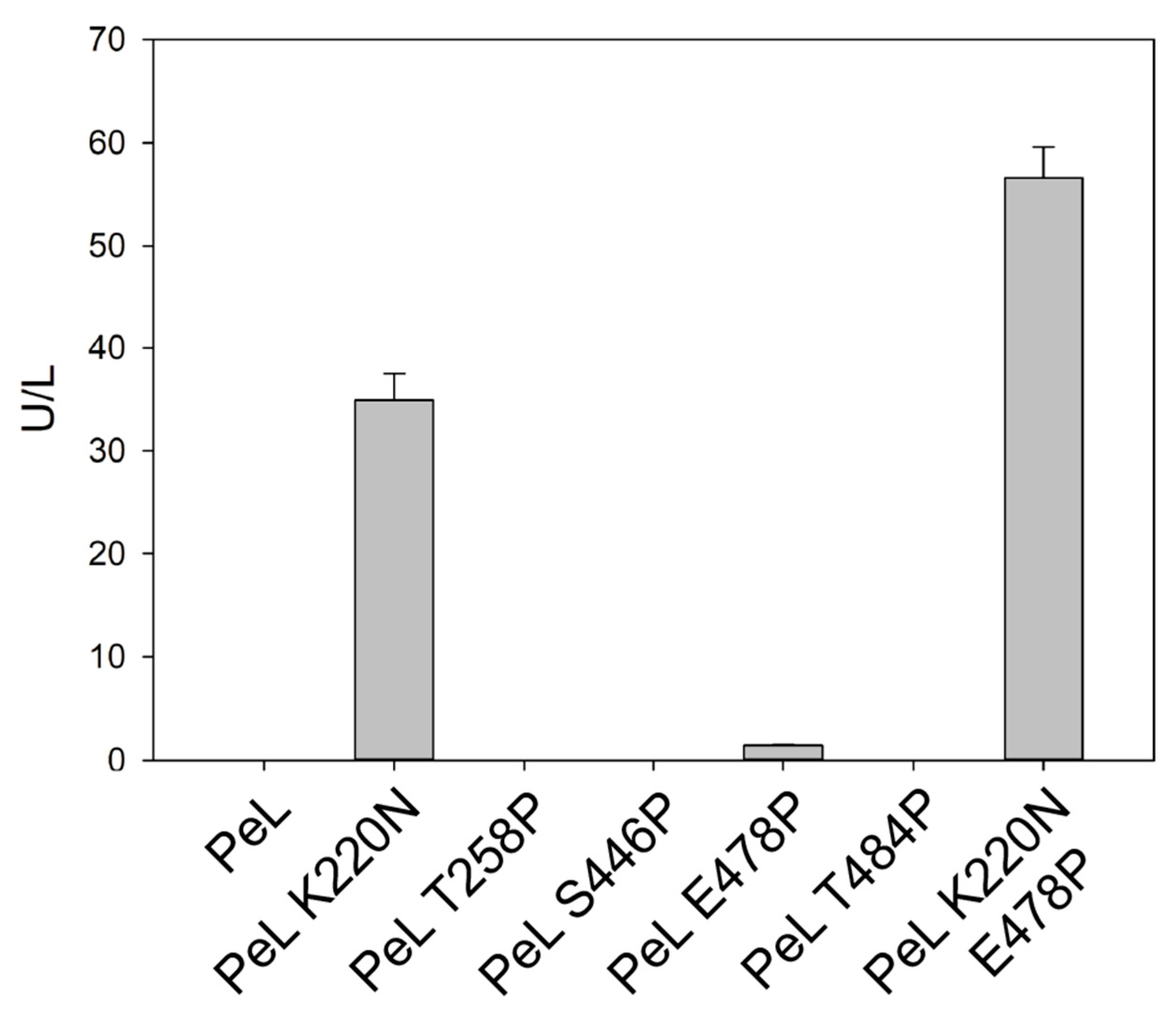
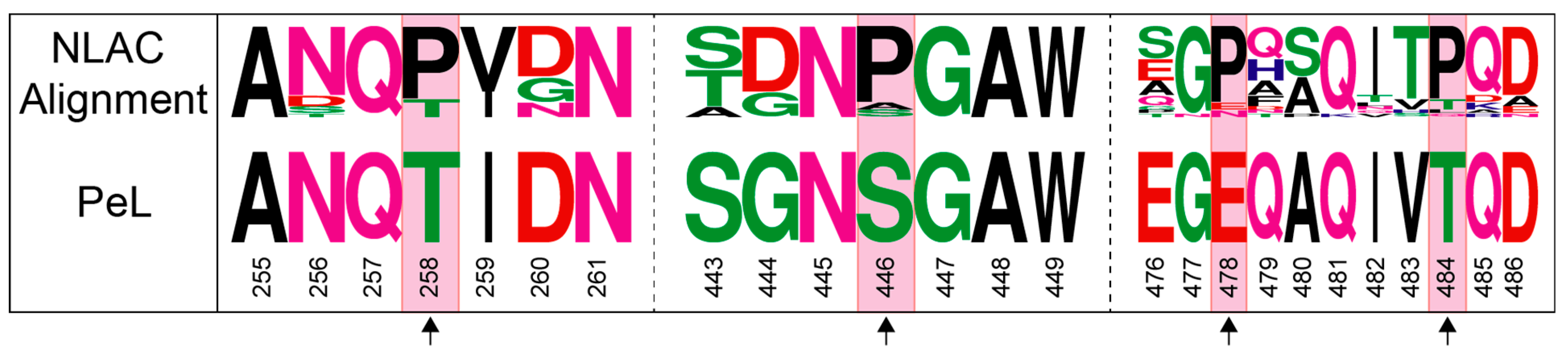
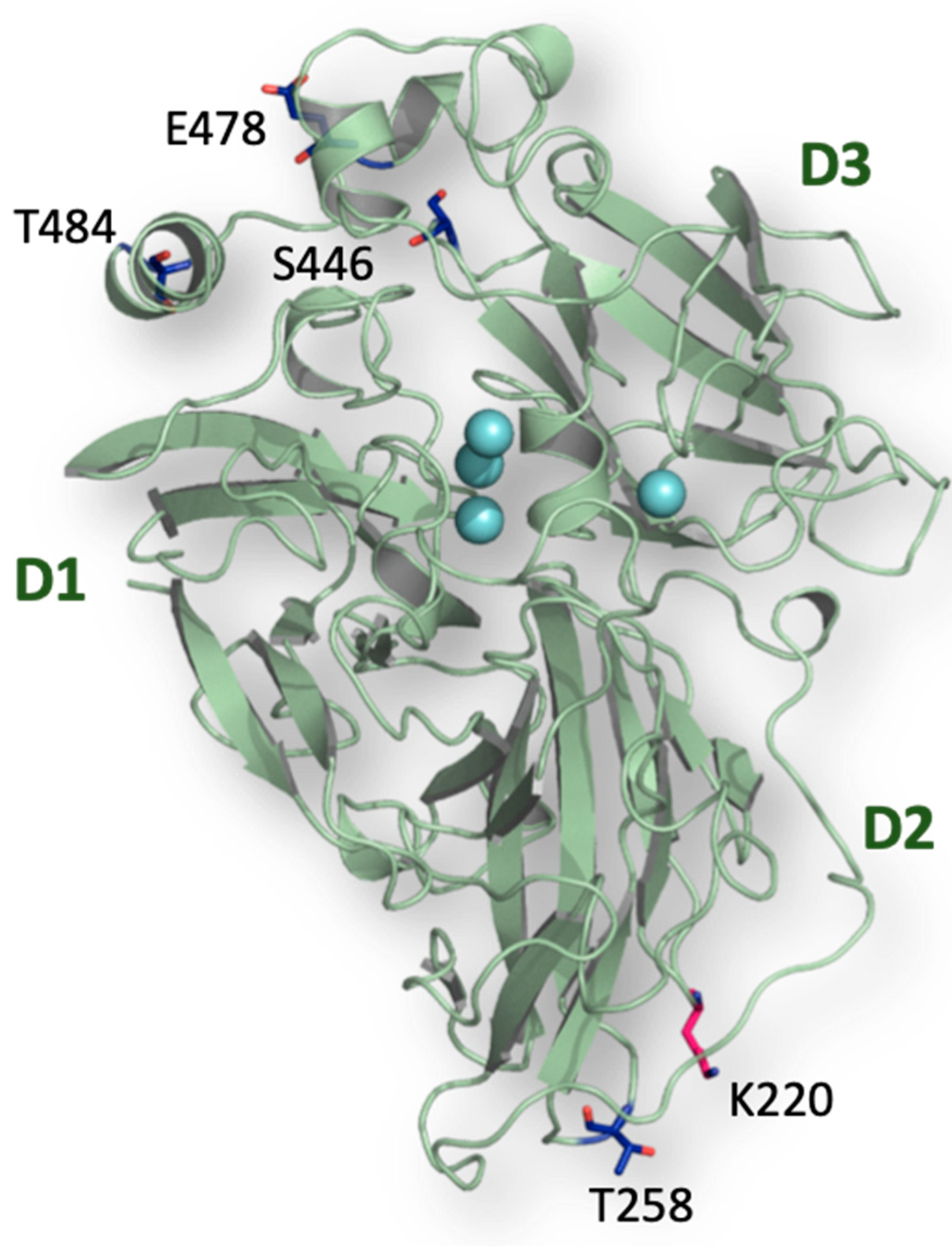
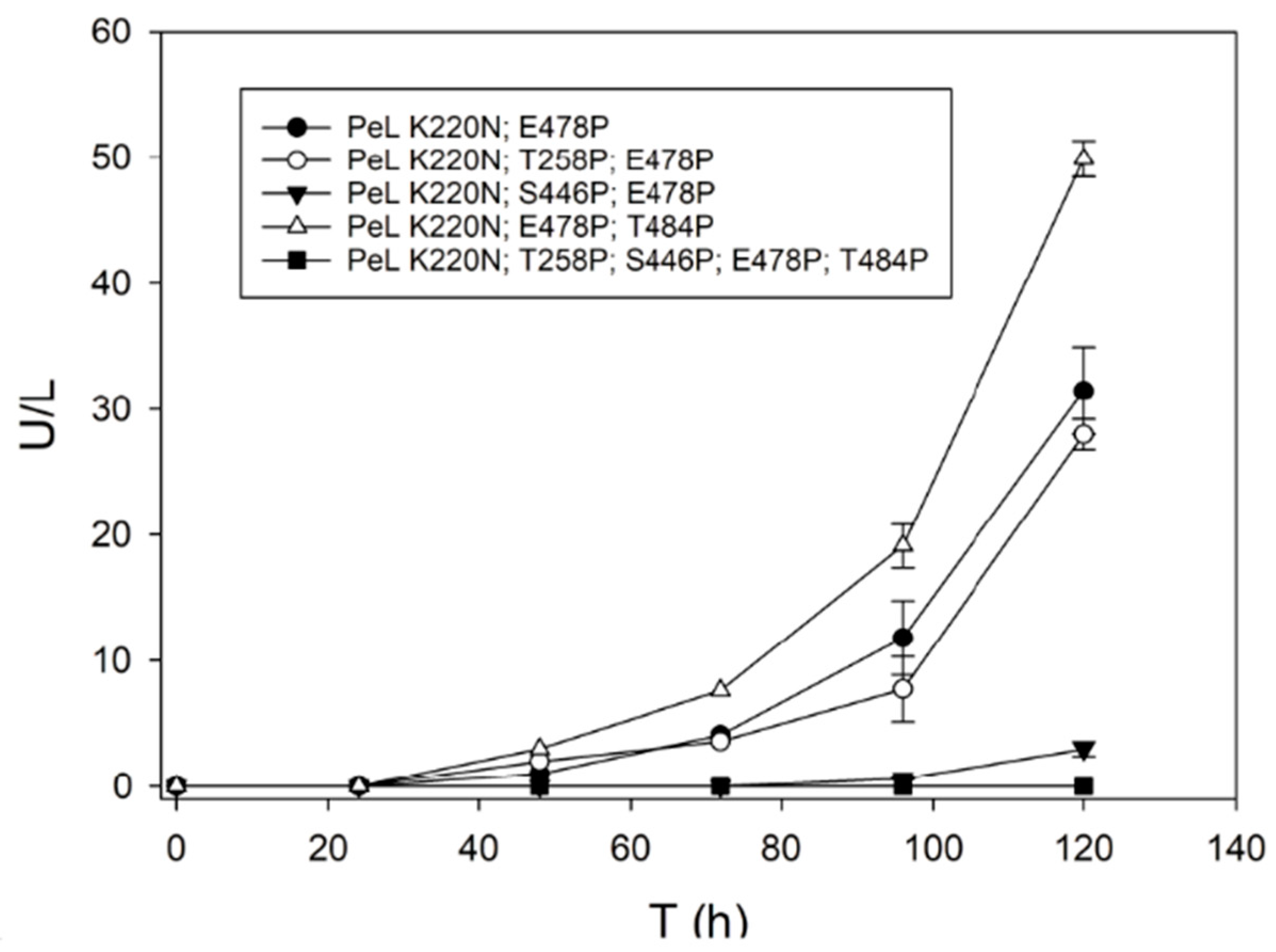
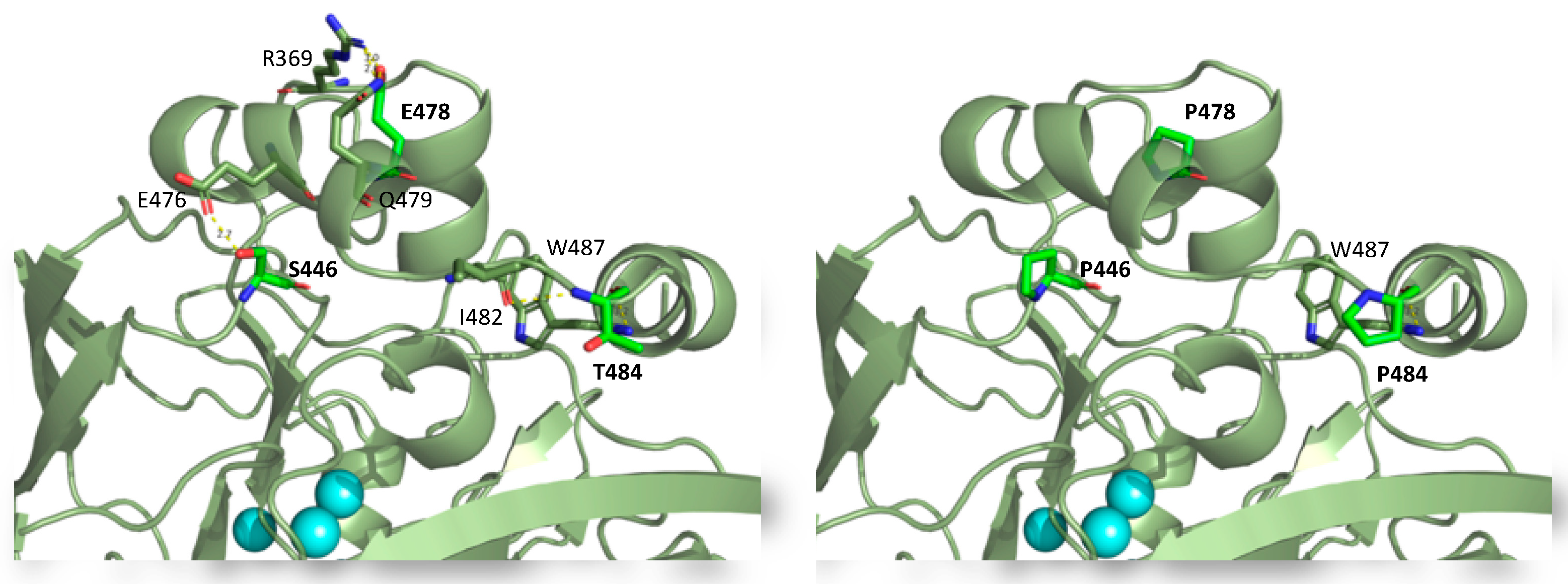
2.3. Consensus Enzyme Design
2.4. Mutations Accumulated in the Protein CDS during Directed Evolution
3. Materials and Methods
3.1. Reagents and Strains
3.2. Culture and Media
3.3. Predictions and Modelling
3.4. Laccase Variants and Libraries Constructions in S. cerevisiae
3.4.1. Signal Peptides
3.4.2. N-Glycosylation and Consensus Prolines
3.5. Laccase Production
3.5.1. Flask Fermentation
3.5.2. Microwell Production
3.6. Laccase Characterization
3.6.1. pH Activity Profile
3.6.2. Thermostability Assay
3.6.3. pH Stability Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.M.; Solomon, E.I. Electron Transfer and Reaction Mechanism of Laccases. Cell. Mol. Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, M.F.; Monza, E.; Jørgensen, L.J.; Ernst, H.A.; Piontek, K.; Bjerrum, M.J.; Martinez, Á.T.; Camarero, S.; Guallar, V. Simulating Substrate Recognition and Oxidation in Laccases: From Description to Design. J. Chem. Theory Comput. 2017, 13, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- Hakulinen, N.; Rouvinen, J. Three-Dimensional Structures of Laccases. Cell. Mol. Life Sci. 2015, 72, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Piontek, K.; Antorini, M.; Choinowski, T. Crystal Structure of a Laccase from the Fungus Trametes Versicolor at 1.90-Å Resolution Containing a Full Complement of Coppers. J. Biol. Chem. 2002, 277, 37663–37669. [Google Scholar] [CrossRef] [Green Version]
- Glazunova, O.A.; Polyakov, K.M.; Fedorova, T.V.; Dorovatovskii, P.V.; Koroleva, O.V. Elucidation of the Crystal Structure of Coriolopsis Caperata Laccase: Restoration of the Structure and Activity of the Native Enzyme from the T2-Depleted Form by Copper Ions. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Maestre-Reyna, M.; Liu, W.C.; Jeng, W.Y.; Lee, C.C.; Hsu, C.A.; Wen, T.N.; Wang, A.H.J.; Shyur, L.F. Structural and Functional Roles of Glycosylation in Fungal Laccase from Lentinus Sp. PLoS ONE 2015, 10, 0120601. [Google Scholar] [CrossRef]
- Riva, S. Laccases: Blue Enzymes for Green Chemistry. Trends Biotechnol. 2006, 24, 219–226. [Google Scholar] [CrossRef]
- Cassland, P.; Jönsson, L.J. Characterization of a Gene Encoding Trametes Versicolor Laccase A and Improved Heterologous Expression in Saccharomyces Cerevisiae by Decreased Cultivation Temperature. Appl. Microbiol. Biotechnol. 1999, 52, 393–400. [Google Scholar] [CrossRef]
- Kojima, Y.; Tsukuda, Y.; Kawai, Y.; Tsukamoto, A.; Sugiura, J.; Sakaino, M.; Kita, Y. Cloning, Sequence Analysis, and Expression of Ligninolytic Phenoloxidase Genes of the White-Rot Basidiomycete Coriolus Hirsutus. J. Biol. Chem. 1990, 265, 15224–15230. [Google Scholar] [CrossRef]
- Larsson, S.; Cassland, P.; Jönsson, L.J. Development of a Saccharomyces Cerevisiae Strain with Enhanced Resistance to Phenolic Fermentation Inhibitors in Lignocellulose Hydrolysates by Heterologous Expression of Laccase. Appl. Environ. Microbiol. 2001, 67, 1163–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarero, S.; Pardo, I.; Cañas, A.I.; Molina, P.; Record, E.; Martínez, A.T.; Martínez, M.J.; Alcalde, M. Engineering Platforms for Directed Evolution of Laccase from Pycnoporus Cinnabarinus. Appl. Environ. Microbiol. 2012, 78, 1370–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraco, V.; Ercole, C.; Festa, G.; Giardina, P.; Piscitelli, A.; Sannia, G. Heterologous Expression of Heterodimeric Laccase from Pleurotus Ostreatus in Kluyveromyces Lactis. Appl. Microbiol. Biotechnol. 2008, 77, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Jolivalt, C.; Madzak, C.; Brault, A.; Caminade, E.; Malosse, C.; Mougin, C. Expression of Laccase IIIb from the White-Rot Fungus Trametes Versicolor in the Yeast Yarrowia Lipolytica for Environmental Applications. Appl. Microbiol. Biotechnol. 2005, 66, 450–456. [Google Scholar] [CrossRef]
- Gelo-Pujic, M.; Kim, H.-H.; Butlin, N.G.; Palmore, G.T.R. Electrochemical Studies of a Truncated Laccase Produced in Pichia Pastoris. Appl. Environ. Microbiol. 1999, 65, 5515–5521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jönsson, L.J.; Saloheimo, M.; Penttilä, M. Laccase from the White-Rot Fungus Trametes Versicolor: CDNA Cloning of Lcc1 and Expression in Pichia Pastoris. Curr. Genet. 1997, 32, 425–430. [Google Scholar] [CrossRef]
- Otterbein, L.; Record, E.; Longhi, S.; Asther, M.; Moukha, S. Molecular Cloning of the CDNA Encoding Laccase from Pycnoporus Cinnabarinus I-937 and Expression in Pichia Pastoris. Eur. J. Biochem. 2000, 267, 1619–1625. [Google Scholar] [CrossRef]
- Saloheimo, M.; Niku-Paavola, M.L. Heterologous Production of a Ligninolytic Enzyme: Expression of the Phlebia Radiata Laccase Gene in Trichoderma Reesei. BioTechnology 1991, 9, 987–990. [Google Scholar] [CrossRef]
- De Salas, F.; Cañadas, R.; Santiago, G.; Virseda-Jerez, A.; Vind, J.; Gentili, P.; Martínez, A.T.; Guallar, V.; Muñoz, I.G.; Camarero, S. Structural and Biochemical Insights into an Engineered High-Redox Potential Laccase Overproduced in Aspergillus. Int. J. Biol. Macromol. 2019, 141, 855–866. [Google Scholar] [CrossRef]
- Record, E.; Punt, P.J.; Chamkha, M.; Labat, M.; Van Den Hondel, C.A.M.J.J.; Asther, M. Expression of the Pycnoporus Cinnabarinus Laccase Gene in Aspergillus Niger and Characterization of the Recombinant Enzyme. Eur. J. Biochem. 2002, 269, 602–609. [Google Scholar] [CrossRef]
- Gonzalez-Perez, D.; Garcia-Ruiz, E.; Alcalde, M. Saccharomyces Cerevisiae in Directed Evolution: An Efficient Tool to Improve Enzymes. Bioeng. Bugs 2012, 3, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çelik, E.; Çalik, P. Production of Recombinant Proteins by Yeast Cells. Biotechnol. Adv. 2012, 30, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Vieira Gomes, A.; Souza Carmo, T.; Silva Carvalho, L.; Mendonça Bahia, F.; Parachin, N. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshitake, A.; Katayama, Y.; Nakamura, M.; Iimura, Y.; Kawai, S.; Morohoshi, N. N-Linked Carbohydrate Chains Protect Laccase III from Proteolysis in Coriolus Versicolor. Microbiology 1993, 139, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.J.; Kepp, K.P. Stability Mechanisms of a Thermophilic Laccase Probed by Molecular Dynamics. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Conde, R.; Cueva, R.; Pablo, G.; Polaina, J.; Larriba, G. A Search for Hyperglycosylation Signals in Yeast Glycoproteins. J. Biol. Chem. 2004, 279, 43789–43798. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Chen, E.Y.; Lugovoy, J.M.; Chang, C.N.; Hitzeman, R.A.; Seeburg, P.H. Saccharomyces Cerevisiae Contains Two Discrete Genes Coding for the α-Factor Pheromone. Nucleic Acids Res. 1983, 11, 4049–4063. [Google Scholar] [CrossRef]
- De Salas, F.; Aza, P.; Gilabert, J.F.; Santiago, G.; Kilic, S.; Sener, M.E.; Vind, J.; Guallar, V.; Martínez, A.T.; Camarero, S. Engineering of a Fungal Laccase to Develop a Robust, Versatile and Highly-Expressed Biocatalyst for Sustainable Chemistry. Green Chem. 2019, 21, 5374–5385. [Google Scholar] [CrossRef] [Green Version]
- Pardo, I.; Vicente, A.I.; Mate, D.M.; Alcalde, M.; Camarero, S. Development of Chimeric Laccases by Directed Evolution. Biotechnol. Bioeng. 2012, 109, 2978–2986. [Google Scholar] [CrossRef]
- Rakestraw, J.A.; Sazinsky, S.L.; Piatesi, A.; Antipov, E.; Wittrup, K.D. Directed Evolution of a Secretory Leader for the Improved Expression of Heterologous Proteins and Full-Length Antibodies in Saccharomyces Cerevisiae. Biotechnol. Bioeng. 2009, 103, 1192–1201. [Google Scholar] [CrossRef] [Green Version]
- Lin-Cereghino, G.P.; Stark, C.M.; Kim, D.; Chang, J.; Shaheen, N.; Poerwanto, H.; Agari, K.; Moua, P.; Low, L.K.; Tran, N.; et al. The Effect of α-Mating Factor Secretion Signal Mutations on Recombinant Protein Expression in Pichia Pastoris. Gene 2013, 519, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maté, D.; García-Burgos, C.; García-Ruiz, E.; Ballesteros, A.O.; Camarero, S.; Alcalde, M. Laboratory Evolution of High-Redox Potential Laccases. Chem. Biol. 2010, 17, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Bulter, T.; Alcalde, M.; Sieber, V.; Meinhold, P.; Schlachtbauer, C.; Arnold, F.H. Functional Expression of a FungallLaccase in Saccharomyces Cerevisiae by Directed Evolution. Appl. Environ. Microbiol. 2003, 69, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Barrero, J.J.; Casler, J.C.; Valero, F.; Ferrer, P.; Glick, B.S. An Improved Secretion Signal Enhances the Secretion of Model Proteins from Pichia Pastoris. Microb. Cell Fact. 2018, 17, 161. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Santiago, G.; Gentili, P.; Lucas, F.; Monza, E.; Medrano, F.J.; Galli, C.; Martínez, A.T.; Guallar, V.; Camarero, S. Re-Designing the Substrate Binding Pocket of Laccase for Enhanced Oxidation of Sinapic Acid. Catal. Sci. Technol. 2016, 6, 3900–3910. [Google Scholar] [CrossRef] [Green Version]
- Mateljak, I.; Tron, T.; Alcalde, M. Evolved α-Factor Prepro-Leaders for Directed Laccase Evolution in Saccharomyces Cerevisiae. Microb. Biotechnol. 2017, 10, 1830–1836. [Google Scholar] [CrossRef]
- Pardo, I.; Rodríguez-Escribano, D.; Aza, P.; de Salas, F.; Martínez, A.T.; Camarero, S. A Highly Stable Laccase Obtained by Swapping the Second Cupredoxin Domain. Sci. Rep. 2018, 8, 15669. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Dueñas, F.J.; Barrasa, J.M.; Sánchez-García, M.; Camarero, S.; Miyauchi, S.; Serrano, A.; Linde, D.; Babiker, R.; Drula, E.; Ayuso-Fernández, I.; et al. Genomic Analysis Enlightens Agaricales Lifestyle Evolution and Increasing Peroxidase Diversity. Mol. Biol. Evol. 2020, 19, msaa301. [Google Scholar] [CrossRef]
- Vite-Vallejo, O.; Palomares, L.A.; Dantán-González, E.; Ayala-Castro, H.G.; Martínez-Anaya, C.; Valderrama, B.; Folch-Mallol, J. The Role of N-Glycosylation on the Enzymatic Activity of a Pycnoporus Sanguineus Laccase. Enzyme Microb. Technol. 2009, 45, 233–239. [Google Scholar] [CrossRef]
- Glazunova, O.A.; Moiseenko, K.V.; Kamenihina, I.A.; Isaykina, T.U.; Yaropolov, A.I.; Fedorova, T.V. Laccases with Variable Properties from Different Strains of Steccherinum Ochraceum: Does Glycosylation Matter? Int. J. Mol. Sci. 2019, 20, 2008. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Wu, Y.; Zhang, Y.; Fang, W.; Xiao, Y.; Fang, Z. Role of N-Glycosylation on the Specific Activity of a Coprinopsis Cinerea Laccase Lcc9 Expressed in Pichia Pastoris. J. Biosci. Bioeng. 2019, 128, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Bieler, N.; Kenzom, T.; Chhabra, M.; Ansorge-Schumacher, M.; Mishra, S. Cloning, Sequence Analysis, Expression of Cyathus Bulleri Laccase in Pichia Pastoris and Characterization of Recombinant Laccase. BMC Biotechnol. 2012, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Madzak, C.; Otterbein, L.; Chamkha, M.; Moukha, S.; Asther, M.; Gaillardin, C.; Beckerich, J.M. Heterologous Production of a Laccase from the Basidiomycete Pycnoporus Cinnabarinus in the Dimorphic Yeast Yarrowia Lipolytica. FEMS Yeast Res. 2005, 5, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Yu, X. Enhanced Expression of Heterologous Proteins in Yeast Cells via the Modification of N-Glycosylation Sites. Bioengineered 2015, 6, 115–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagt, C.M.J.; Kleiken, B.; Verwaal, R.; De Jong, M.D.M.; Muller, W.H.; SMITS, A.; Visser, C.; Boonstra, J.; Verkleij, A.J.; Verrips, C.T. Introduction of an N-Glycosylation Site Increases Secretion of Heterologous Proteins in Yeasts. Appl. Environ. Microbiol. 2000, 66, 4940–4944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porebski, B.T.; Buckle, A.M. Consensus Protein Design. Protein Eng. Des. Sel. 2016, 29, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic Enzymes: Sources, Uses, and Molecular Mechanisms for Thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Bloom, J.D.; Labthavikul, S.T.; Otey, C.R.; Arnold, F.H. Protein Stability Promotes Evolvability. Proc. Natl. Acad. Sci. USA 2006, 103, 5869–5874. [Google Scholar] [CrossRef] [Green Version]
- Jäckel, C.; Bloom, J.D.; Kast, P.; Arnold, F.H.; Hilvert, D. Consensus Protein Design without Phylogenetic Bias. J. Mol. Biol. 2010, 399, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Matthews, B.W.; Nicholson, H.; Becktel, W.J. Enhanced Protein Thermostability from Site-Directed Mutations That Decrease the Entropy of Unfolding. Proc. Natl. Acad. Sci. USA 1987, 84, 6663–6667. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Fernandez, B.J.; Risso, V.A.; Sanchez-Ruiz, J.M.; Alcalde, M. Consensus Design of an Evolved High-Redox Potential Laccase. Front. Bioeng. Biotechnol. 2020, 8, 354. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Reilly, P.J.; Ford, C. Effect of Introducing Proline Residues on the Stability of Aspergillus Awamori. Protein Eng. 1997, 10, 1199–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, K.; Higashi, R.; Okada, J.; Mukaiyama, A.; Tadokoro, T.; Koga, Y.; Kanaya, S. Proline Effect on the Thermostability and Slow Unfolding of a Hyperthermophilic Protein. J. Biochem. 2009, 145, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Yun, R.H.; Anderson, A.; Hermans, J. Proline in Α-helix: Stability and Conformation Studied by Dynamics Simulation. Proteins Struct. Funct. Bioinform. 1991, 10, 219–228. [Google Scholar] [CrossRef]
- Yang, J.M.; Yoneda, K.; Morita, E.; Imamura, S.; Nam, K.; Lee, E.S.; Steinert, P.M. An Alanine to Proline Mutation in the 1A Rod Domain of the Keratin 10 Chain in Epidermolytic Hyperkeratosis. J. Investig. Dermatol. 1997, 109, 692–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, E.C.; Wilman, H.R.; Kelm, S.; Shi, J.; Deane, C.M. Examining the Conservation of Kinks in Alpha Helices. PLoS ONE 2016, 11, 0157553. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, S.H.; Hirano, T. HEAT Repeats—Versatile Arrays of Amphiphilic Helices Working in Crowded Environments? J. Cell Sci. 2016, 129, 3963–3970. [Google Scholar] [CrossRef] [Green Version]
- Kumeta, M.; Konishi, H.A.; Zhang, W.; Sakagami, S.; Yoshimura, S.H. Prolines in the α-Helix Confer the Structural Flexibility and Functional Integrity of Importin-β. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Ge, M.; Pan, X.M. The Contribution of Proline Residues to Protein Stability Is Associated with Isomerization Equilibrium in Both Unfolded and Folded States. Extremophiles 2009, 13, 481–489. [Google Scholar] [CrossRef]
- Das, M.; Basu, G. Glycine Rescue of β-Sheets from Cis-Proline. J. Am. Chem. Soc. 2012, 134, 16536–16539. [Google Scholar] [CrossRef]
- Kuncha, S.K.; Mazeed, M.; Singh, R.; Kattula, B.; Routh, S.B.; Sankaranarayanan, R. A Chiral Selectivity Relaxed Paralog of DTD for Proofreading TRNA Mischarging in Animalia. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusano, A.M.; Mekmouche, Y.; Meglecz, E.; Tron, T. Plasticity of Laccase Generated by Homeologous Recombination in Yeast. FEBS J. 2009, 276, 5471–5480. [Google Scholar] [CrossRef] [PubMed]
- Vicente, A.I.; Viña-Gonzalez, J.; Santos-Moriano, P.; Marquez-Alvarez, C.; Ballesteros, A.O.; Alcalde, M. Evolved Alkaline Fungal Laccase Secreted by Saccharomyces Cerevisiae as Useful Tool for the Synthesis of C–N Heteropolymeric Dye. J. Mol. Catal. B Enzym. 2016, 134, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Romanos, M.A.; Scorer, C.A.; Clare, J.J. Foreign Gene Expression in Yeast: A Review. Yeast 1992, 8, 423–488. [Google Scholar] [CrossRef] [PubMed]
- Tuller, T.; Waldman, Y.Y.; Kupiec, M.; Ruppin, E. Translation Efficiency Is Determined by Both Codon Bias and Folding Energy. Proc. Natl. Acad. Sci. USA 2010, 107, 3645–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Puigbo, P.; Guzman, E.; Romeu, A.; Garcia-Vallve, S. OPTIMIZER: A Web Server for Optimizing the Codon Usage of DNA Sequences. Nucleic Acids Res. 2007, 35, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, M.; Zumarraga, M.; Polaina, J.; Ballesteros, A.; Plou, F. Combinatorial Saturation Mutagenesis by In Vivo Overlap Extension for the Engineering of Fungal Laccases. Comb. Chem. High Throughput Screen. 2006, 9, 719–727. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PeL Variant | T50 (°C) | pH 3 | pH 5 | pH 7 | pH 9 |
|---|---|---|---|---|---|
| K220N | 46 ± 0.5 | - | - | - | - |
| K220N, E478P | 49 ± 0.1 | 23 ± 3 | 93 ± 8 | 88 ± 5 | 85 ± 1 |
| K220N, E478P, T484P | 47 ±0.1 | 15 ± 1 | 93 ± 2 | 100 ± 1 | 84 ± 2 |
| K220N, S446P, E478P | 43 ± 0.2 | 32 ± 1 | 95 ± 2 | 92 ± 1 | 83 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aza, P.; de Salas, F.; Molpeceres, G.; Rodríguez-Escribano, D.; de la Fuente, I.; Camarero, S. Protein Engineering Approaches to Enhance Fungal Laccase Production in S. cerevisiae. Int. J. Mol. Sci. 2021, 22, 1157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031157
Aza P, de Salas F, Molpeceres G, Rodríguez-Escribano D, de la Fuente I, Camarero S. Protein Engineering Approaches to Enhance Fungal Laccase Production in S. cerevisiae. International Journal of Molecular Sciences. 2021; 22(3):1157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031157
Chicago/Turabian StyleAza, Pablo, Felipe de Salas, Gonzalo Molpeceres, David Rodríguez-Escribano, Iñigo de la Fuente, and Susana Camarero. 2021. "Protein Engineering Approaches to Enhance Fungal Laccase Production in S. cerevisiae" International Journal of Molecular Sciences 22, no. 3: 1157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031157