
Intestinal SIRT1 Deficiency-Related Intestinal Inflammation and Dysbiosis Aggravate TNFα-Mediated Renal Dysfunction in Cirrhotic Ascitic Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
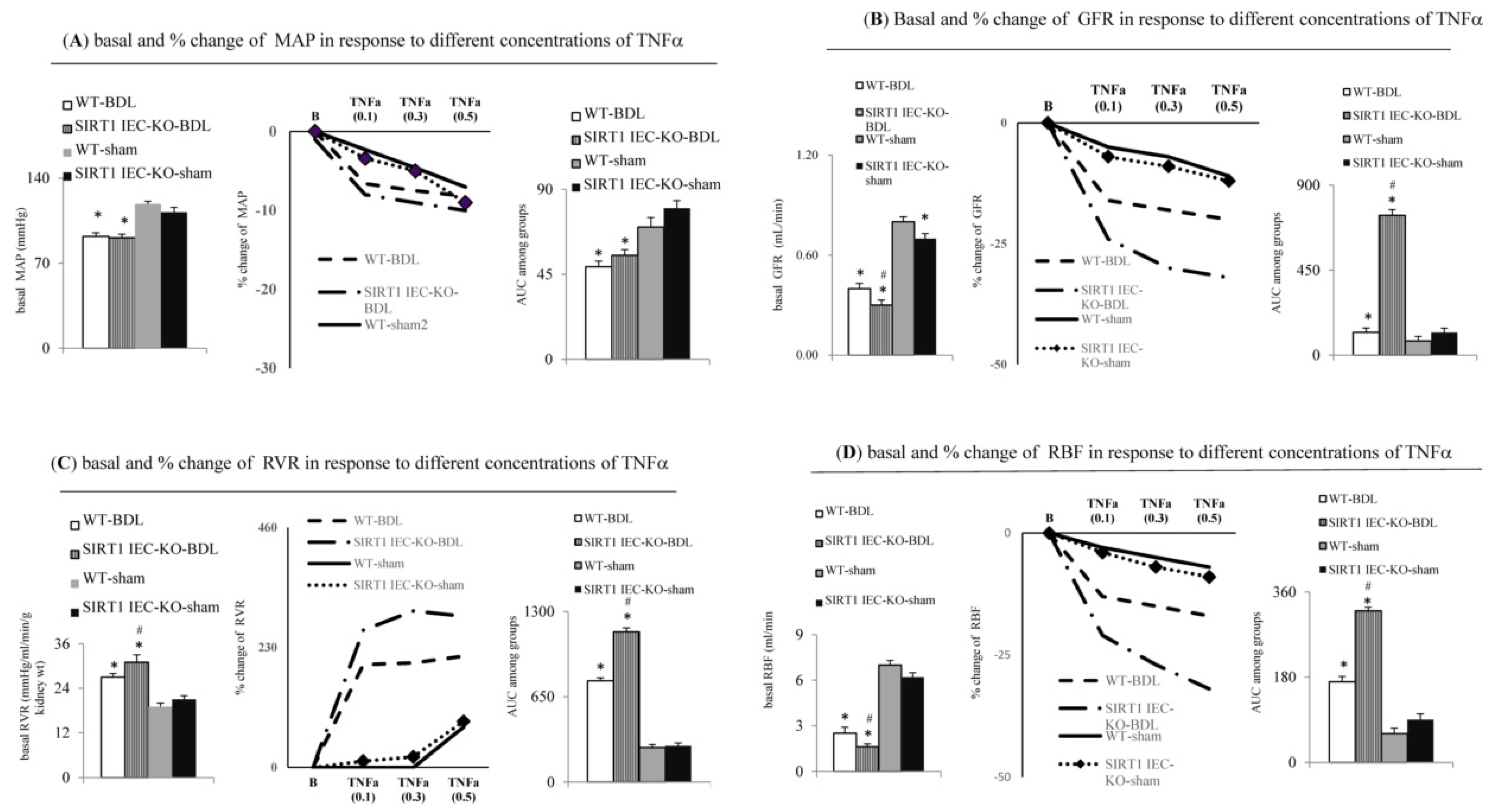
2.1. Intestinal SIRT1 Deficiency Aggravates Severity of Renal Dysfunction in SIRT1IEC-KO-BDL-Cirrhotic Mice with Ascites
2.2. Genetic Deletion of Intestinal SIRT1 Exacerbates TNFα-Mediated Intestinal Inflammation and Barrier Dysfunction in SIRT1IEC KO—BDL Mice
2.3. Intestinal SIRT1 Deficiency Aggravated Renal Dysfunction through the Upregulation of TNFα-Activated Signals in SIRT1IEC KO-BDL Mice
2.4. Direct Evidence for the Aggravation of TNFa-Induced Renal Dysfunction in SIRT1IEC KO BDL-Cirrhotic Mice due to the Deficiency of Intestinal SIRT1
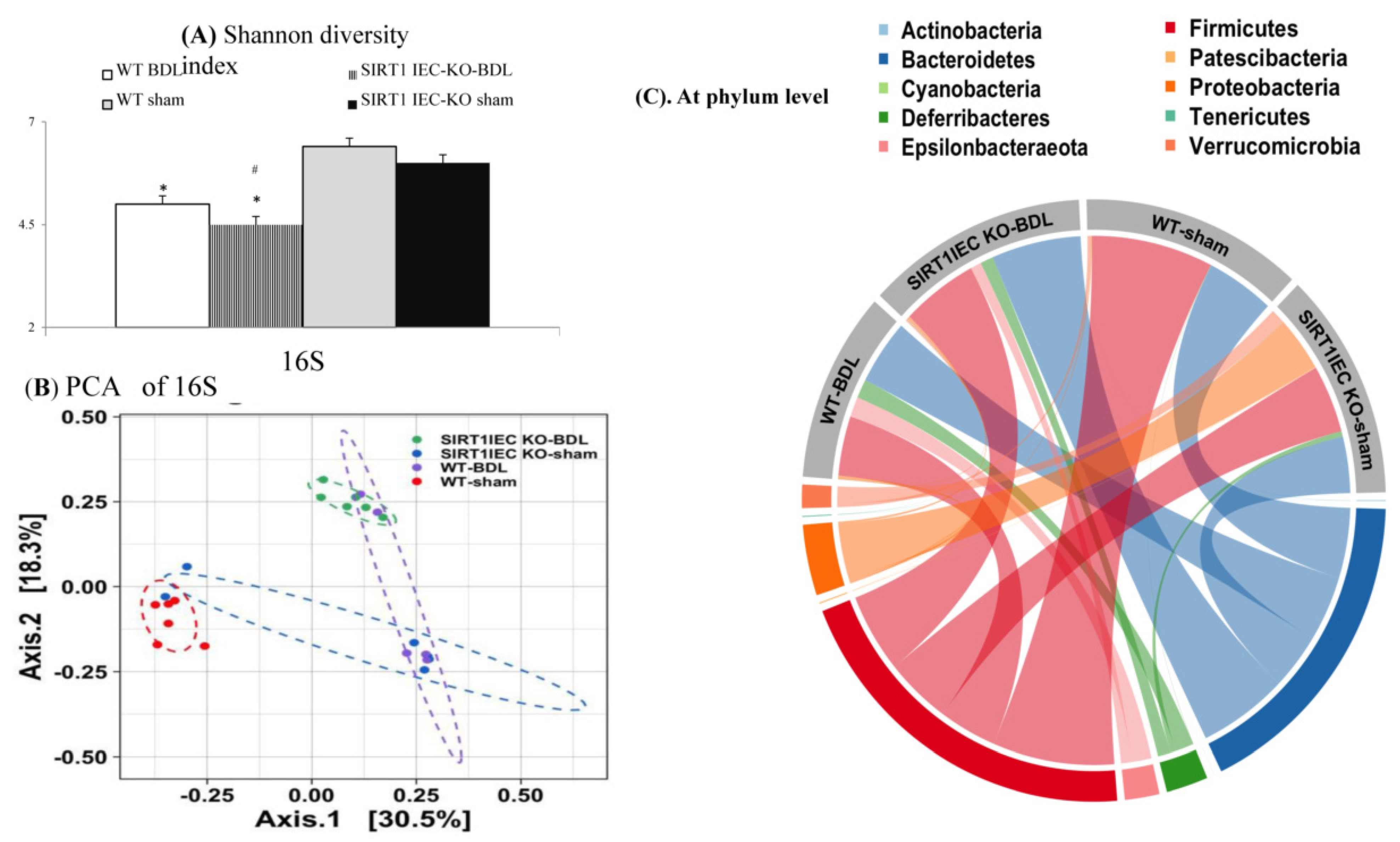
2.5. Genetic Deletion of Intestinal SIRT1 Aggravates the Intestinal Bacterial Dysbiosis in SIRT1IEC KO-Mice
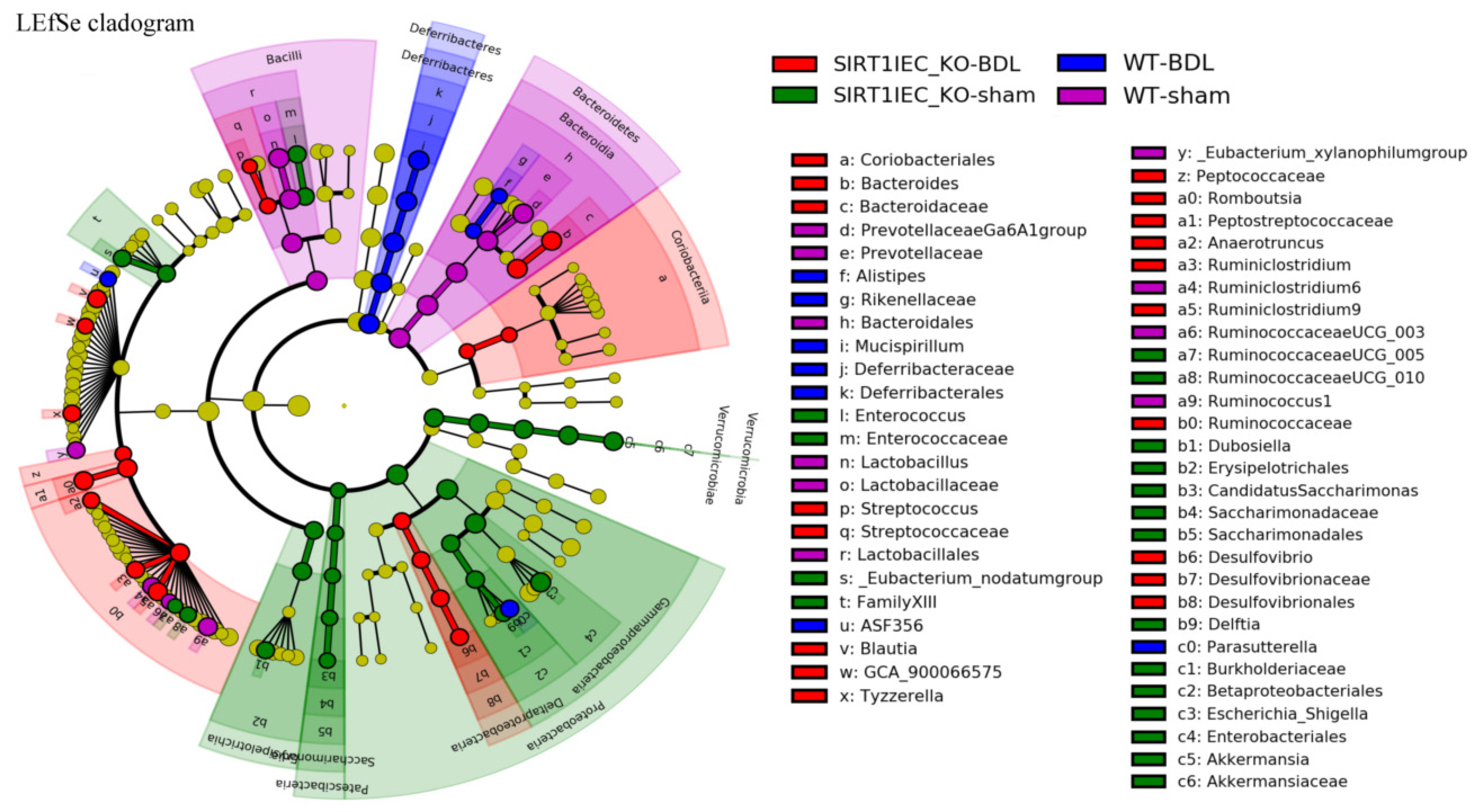
2.6. Shifting to Proinflammatory Bacteria that have Biological Relevance for Intestinal Epithelial Barrier Dysfunction in SIRT1IEC-KO-BDL Mice
2.7. Intestinal Bacterial Dysbiosis was Associated with the Upregulation of Renal Inflammatory and Barrier-Disrupted Markers in SIRT1IEC-KO Mice
2.8. Direct Link between Intestinal Bacterial Dysbiosis and Severe Renal Dysfunction in SIRT1IEC-KO-BDL Mice
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Groupings
5.3. Common Measurements between Two Sets of Mice
5.4. Experiments in the First Set of four Groups of Mice
5.4.1. Systemic and Renal Hemodynamic Measurements
5.4.2. Measurements of Intestinal Permeability and Intestinal Inflammation
5.4.3. Measurements of Tissue Profiles
5.4.4. 16S rRNA Gene Sequencing Analysis for Intestinal Bacterial Microbiome
5.4.5. Direct Correlation between Intestinal Dysbiosis and Renal Dysfunction of Cirrhotic Mice
5.5. Experiments in the Second set of Mice
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bassegoda, O.; Huelin, P.; Ariza, X.; Solé, C.; Juanola, A.; Gratacós-Ginès, J.; Carol, M.; Graupera, I.; Pose, E.; Napoleone, L.; et al. Development of chronic kidney disease after acute kidney injury in patients with cirrhosis is common and impairs clinical outcomes. J. Hepatol. 2020, 72, 1132–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeli, P.; Garcia-Tsao, G.; Nadim, M.K.; Parikh, C.R. News in pathophysiology, definition and classification of hepatorenal syndrome: A step beyond the International Club of Ascites (ICA) consensus document. J. Hepatol. 2019, 71, 811–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sole, C.; Sola, E.; Morales-Ruiz, M.; Huelin, P.; Carol, M.; Moreira, R.; Graupera, I.; Pose, E.; Napoleone, L.; De Prada, G.; et al. Characterization of systemic inflammatory response in hepatorenal syndrome in cirrhosis. A major role for il-6, TNF-alpha, and VCAM. J. Hepatol. 2018, 68, S698. [Google Scholar] [CrossRef]
- Zipser, R.D.; Radvan, G.H.; Kronborg, I.J.; Duke, R.; Little, T.E. Urinary thromboxane B2 and prostaglandin E2 in the hepatorenal syndrome: Evidence for increased vasoconstrictor and decreased vasodilator factors. Gastroenterology 1983, 84, 697–703. [Google Scholar] [CrossRef]
- Shah, N.; Dhar, D.; Mohammed, F.E.Z.; Habtesion, A.; Davies, N.A.; Jover-Cobos, M.; Macnaughtan, J.; Sharma, V.; Olde Damink, S.W.M.; Mookerjee, R.P.; et al. Prevention of acute kidney injury in a rodent model of cirrhosis following selective gut decontamination is associated with reduced renal TLR4 expression. J. Hepatol. 2012, 56, 1047–1053. [Google Scholar] [CrossRef]
- Rodriguez, E.; Pereira, G.H.; Solà, E.; Elia, C.; Barreto, R.; Pose, E.; Colmenero, J.; Fernandez, J.; Navasa, M.; Arroyo, V.; et al. Treatment of type 2 hepatorenal syndrome in patients awaiting transplantation: Effects on kidney function and transplantation outcomes. Liver Transpl. 2015, 21, 1347–1354. [Google Scholar] [CrossRef]
- Best, L.M.; Freeman, S.; Sutton, A.J.; Hawkins, N.; Tsochatzis, E.; Gurusamy, K.S. Treatment for hepatorenal syndrome in people with decompensated liver cirrhosis: A network meta-analysis. Cochrane Database Syst. Rev. 2019, 9, CD013103. [Google Scholar] [CrossRef]
- Navasa, M.; Follo, A.; Filella, X.; Jiménez, W.; Francitorra, A.; Planas, R.; Rimola, A.; Arroyo, V.; Rodés, J. Tumor necrosis factor and interleukin-6 in spontaneous bacterial peritonitis in cirrhosis: Relationship with the development of renal impairment and mortality. Hepatology 1998, 27, 1227–1232. [Google Scholar] [CrossRef]
- Tyagi, P.; Sharma, P.; Sharma, B.C.; Puri, A.S.; Kumar, A.; Sarin, S.K. Prevention of hepatorenal syndrome in patients with cirrhosis and ascites: A pilot randomized control trial between pentoxifylline and placebo. Eur. J. Gastroenterol. Hepatol. 2011, 23, 210–217. [Google Scholar] [CrossRef]
- Stine, J.G.; Wang, J.; Cornella, S.L.; Behm, B.W.; Henry, Z.; Shah, N.L.; Caldwell, S.H.; Northup, P.G. Treatment of type-1 hepatorenal syndrome with pentoxifylline: A randomized placebo controlled clinical trial. Ann. Hepatol. 2018, 17, 300–306. [Google Scholar] [CrossRef]
- Majid, D.S.A. Tumor necrosis factor-α and kidney function: Experimental findings in mice. Adv. Exp. Med. Biol. 2011, 691, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Francis, J.; Majid, D.S.A. Tumor necrosis factor-α induces renal vasoconstriction as well as natriuresis in mice. Am. J. Physiol. Renal Physiol. 2008, 295, F1836–F1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.L.; Techasatian, W.; Hato, T.; Liangpunsakul, S. Role of endotoxemia in causing renal dysfunction in cirrhosis. Investig. Med. 2020, 68, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.B.; Zhu, F.Q.; Chen, W.G.; Jiang, L.P.; Chen, J.; Hu, Z.P.; Huang, Y.J.; Zhou, Z.W.; Wang, G.L.; Lin, H.; et al. Oxymatrine improves intestinal epithelial barrier function involving NFκB-mediated signaling pathway in CCl4-induced cirrhotic rats. PLoS ONE 2014, 9, e106082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francés, R.; Chiva, M.; Sanchez, E.; González-Navajas, J.M.; Llover, T.; Zapater, P.; Soriano, G.; Muñoz, C.; Balanzó, J.; Pérez-Mateo, M.; et al. Bacterial translocation is downregulated by anti-TNF-α monoclonal antibody administration in rats with cirrhosis and ascites. J. Hepatol. 2007, 46, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.S.; Aronsohn, A.; Reddy, K.G.; Te, H.S. Rifaximin decreases the incidence and severity of acute kidney injury and hepatorenal syndrome in cirrhosis. Dig. Dis. Sci. 2016, 61, 3621–3626. [Google Scholar] [CrossRef]
- Larrosa, M.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Cerón, J.J.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Effect of low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef]
- Caruso, R.C.; Marafini, I.; Franze, E.; Stolfi, C.; Zorzi, F.; Monteleone, G.; Caprioli, F.; Colantoni, A.; Sarra, M.; Sedda, S.; et al. Defective expression of SIRT1 contributes to sustain inflammatory pathways in the gut. Mucosal Immunol. 2014, 7, 1467–1479. [Google Scholar] [CrossRef]
- Wellman, A.S.; Metukuri, M.R.; Kazgan, N.; Xu, X.; Xu, Q.; Ren, N.S.X.; Czopik, A.; Shanahan, M.T.; Kang, A.; Chen, W.; et al. Intestinal epithelial sirtuin 1 regulates intestinal inflammation during aging in mice by altering the intestinal microbiota. Gastroenterology 2017, 153, 772–786. [Google Scholar] [CrossRef]
- Gan, Z.; Wei, W.; Li, Y.; Wu, J.; Zhao, Y.; Zhang, L.; Wang, T.; Zhong, X. Curcumin and resveratrol regulate intestinal bacteria and alleviate intestinal inflammation in weaned piglets. Molecules 2019, 24, 1220. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-inflammatory effects of resveratrol, curcumin and simvastatin in acute small intestinal inflammation. PLoS ONE 2010, 5, e15099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajravelu, R.K.; Copelovitch, L.; Osterman, M.T.; Scott, F.I.; Mamtani, R.; Lewis, J.D.; Denburg, M.R. Inflammatory bowel diseases are associated with an increased risk for chronic kidney disease, which decreases with age. Clin. Gastroenterol. Hepatol. 2020, 18, 2262–2268. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Liu, S.M.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Farzaneh, S.H.; Kieffer, D.A.; Adams, S.H.; Martin, R.J. High amylose resistant starch diet ameliorates oxidative stress, inflammation, and progression of chronic kidney disease. PLoS ONE 2014, 9, e114881. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, M.; Zhou, Y.; Wang, C.; Yuan, Y.; Li, L.; Bresette, W.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Resveratrol exerts dose-dependent anti-fibrotic or pro-fibrotic effects in kidneys: A potential risk to individuals with impaired kidney function. Phytomedicine 2019, 57, 223–235. [Google Scholar] [CrossRef]
- Holthoff, J.H.; Wang, Z.; Seely, K.A.; Gokden, N.; Mayeux, P.R. Resveratrol improves renal microcirculation, protects the tubular epithelium, and prolongs survival in a mouse model of sepsis-induced acute kidney injury. Kidney Int. 2012, 81, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Gordish, K.L.; Beierwaltes, W.H. Resveratrol induces acute endothelium-dependent renal vasodilation mediated through nitric oxide and reactive oxygen species scavenging. Am. J. Physiol. Renal Physiol. 2014, 306, F542–F550. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhang, L. Resveratrol provides benefits in mice with type II diabetes-induced chronic renal failure through AMPK signaling pathway. Exp. Med. 2018, 16, 333–341. [Google Scholar] [CrossRef]
- Di Pascoli, M.; Diví, M.; Rodríguez-Vilarrupla, A.; Rosado, E.; Gracia-Sancho, J.; Vilaseca, M.; Bosch, J.; García-Pagán, J.C. Resveratrol improves intrahepatic endothelial dysfunction and reduces hepatic fibrosis and portal pressure in cirrhotic rats. J. Hepatol. 2013, 58, 904–910. [Google Scholar] [CrossRef]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.; Verschoor, C.P.; Loukov, D.; Schenck, L.P.; Jury, J.; Foley, K.P.; et al. Age-associated microbial dysbiosis promotes intestinal permeability, systemic inflammation, and macrophage dysfunction. Cell Host Microbe 2017, 21, 455–466.e4. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, srep08096. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Heuman, D.M.; Hylemon, P.B.; Sanyal, A.J.; White, M.B.; Monteith, P.; Noble, N.A.; Unser, A.B.; Daita, K.; Fisher, A.R.; et al. The cirrhosis dysbiosis ratio defines changes in the gut microbiome associated with cirrhosis and its complications. J. Hepatol. 2014, 60, 940–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.S.; Betrapally, N.S.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; White, M.B.; Unser, A.; Thacker, L.R.; Sanyal, A.J.; Kang, D.J.; et al. Salivary microbiota reflects changes in gut microbiota in cirrhosis with hepatic encephalopathy. Hepatology 2015, 62, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia muciniphila protects against atherosclerosis by preventing metabolic endotoxemia-Induced Inflammation in Apoe-/- Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.Q.; Zha, J.M.; Wang, Y.T.; Turner, J.R. IgCAM domain 3 is necessary for basal and TNF-induced MLCK1 trafficking in intestinal epithelial cells. FASEB J. 2013, 27, 949. [Google Scholar]
- Jin, Y.; Blikslager, A. The regulation of intestinal mucosal barrier by myosin light chain kinase/rho kinases. Int. J. Mol. Sci. 2020, 21, 3550. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, P.; Haimerl, M.; Mazagova, M.; Brenner, D.A.; Schnabl, B. Toll-like receptor 2–Mediated intestinal injury and enteric tumor necrosis factor receptor i contribute to liver fibrosis in mice. Gastroenterology 2012, 143, 1330–1340.e1. [Google Scholar] [CrossRef] [Green Version]
- Kazgan, N.; Metukuri, M.R.; Purushotham, A.; Lu, J.; Rao, A.; Lee, S.; Pratt-Hyatt, M.; Lickteig, A.; Csanaky, I.L.; Zhao, Y.; et al. Intestine-specific deletion of SIRT1 in mice impairs DCoH2–HNF-1α–FXR signaling and alters systemic bile acid homeostasis. Gastroenterology 2014, 146, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, K.A.; Kapsoritakis, A.N.; Theodoridou, C.; Karangelis, D.; Germenis, A.E.; Stefanidis, I.; Potamianos, S.P. Neutrophil gelatinase-associated lipocalin (NGAL) in inflammatory bowel disease: Association with pathophysiology of inflammation, established markers, and disease activity. J. Gastroenterol. 2011, 47, 519–530. [Google Scholar] [CrossRef]
- Hashikata, A.; Yamashita, A.; Suzuki, S.; Nagayasu, S.; Shinjo, T.; Taniguchi, A.; Fukushima, M.; Nakai, Y.; Nin, K.; Watanabe, N.; et al. The inflammation–lipocalin 2 axis may contribute to the development of chronic kidney disease. Nephrol. Dial. Transpl. 2013, 29, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Li, L.P.; Zeng, Z.; Mu, J.X.; Yang, X.; Zhou, C.; Wang, Z.L.; Zhang, H. Value of urinary KIM-1 and NGAL combined with serum Cys C for predicting acute kidney injury secondary to decompensated cirrhosis. Sci. Rep. 2018, 8, 7962. [Google Scholar] [CrossRef] [Green Version]
- Allegretti, A.S.; Solà, E.; Ginès, P. Clinical application of kidney biomarkers in cirrhosis. Am. J. Kidney Dis. 2020, 76, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Yisireyili, M.; Higashiyama, Y.; Nishijima, F.; Niwa, T. Indoxyl sulfate upregulates renal expression of ICAM-1 via production of ROS and activation of NF-κB and p53 in proximal tubular cells. Life Sci. 2013, 92, 143–148. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitani, Y.; Zhang, L.; Yoshida, M.; Azuma, T.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. Intestinal Anti-Inflammatory Activity of Lentinan: Influence on IL-8 and TNFR1 Expression in Intestinal Epithelial Cells. PLoS ONE 2013, 8, e62441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, A.; Islam, M.T.; Prieto, M.C.; Majid, D.S.A. Tumor necrosis factor-α receptor type 1, not type 2, mediates its acute responses in the kidney. Am. J. Physiol. Renal Physiol. 2012, 302, F1650–F1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Stärkel, P.; Turner, J.R.; Ho, S.B.; Schnabl, B. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology 2015, 61, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Murakoshi, M.; Gohda, T.; Suzuki, Y. Circulating tumor necrosis factor receptors: A potential biomarker for the progression of diabetic kidney disease. Int. J. Mol. Sci. 2020, 21, 1957. [Google Scholar] [CrossRef] [Green Version]
- Solé, C.; Solà, E.; Huelin, P.; Carol, M.; Moreira, R.; Cereijo, U.; Mas, J.; Graupera, I.; Pose, E.; Napoleone, L.; et al. Characterization of inflammatory response in hepatorenal syndrome: Relationship with kidney outcome and survival. Liver Int. 2019, 39, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.L.; Liu, C.W.; Hsu, C.F.; Huang, C.C.; Lin, M.W.; Huang, S.F.; Li, T.H.; Lee, K.C.; Hsieh, Y.C.; Yang, Y.Y.; et al. Obeticholic acid ameliorates hepatorenal syndrome in ascitic cirrhotic rats by down-regulating the renal 8-iso-PGF2α-activated COX-TXA2 pathway. Clin. Sci. 2020, 134, 2055–2073. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Hsieh, S.L.; Lee, P.; Yeh, Y.; Lee, K.C.; Hsieh, Y.C.; Wang, Y.W.; Lee, T.Y.; Huang, Y.H.; Chan, C.C.; et al. Long-term cannabinoid type 2 receptor agonist therapy decreases bacterial translocation in rats with cirrhosis and ascites. J. Hepatol. 2014, 6, 1004–1013. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Fausta, K.; Raes, J. CoNet app: Inference of biological association networks using cytoscape. F1000Research 2016, 5, 1519. [Google Scholar] [CrossRef]
- Constanze, B.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Evidence that TNF-β suppresses osteoblast differentiation of mesenchymal stem cells and resveratrol reverses it through modulation of NF-kB, Sirt1 and Runx2. Cell Tissue Res. 2020, 381, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Busch, F.; Shayan, P.; Shakibaei, M.; Teijido, O.; Rappaport, S.M.; Chamberlin, A.; Noskov, S.Y.; Aguilella, V.M.; Rostovtseva, T.K.; et al. Sirtuin-1 (SIRT1) is required for promoting chondrogenic differentiation of mesenchymal stem cells. J. Biol. Chem. 2014, 289, 22048–22062. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Shayan, P.; Popper, B.; Goel, A.; Shakibaei, M. Sirt1 is required for resveratrol-mediated chemopreventive effects in colorectal cancer cells. Nutrients 2016, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Song, X.; Zhang, D.; Guo, N.; Wu, C.; Chen, K.; Liu, Y.; Yuan, L.; Chen, X.; Huang, X. Resveratrol improves left ventricular remodeling in chronic kidney disease via Sirt1-mediated regulation of FoxO1 activity and MnSOD expression. BioFactors 2019, 46, 168–179. [Google Scholar] [CrossRef]
- Zhang, J.; Bi, R.; Meng, Q.; Wang, C.; Huo, X.; Liu, Z.; Wang, C.; Sun, P.; Sun, H.; Ma, X.; et al. Catalpol alleviates adriamycin-induced nephropathy by activating the SIRT1 signalling pathway in vivo and in vitro. Br. J. Pharmacol. 2019, 176, 4558–4573. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT-BDL (n = 7) | SIRT1IEC-KO-BDL (n = 7) | WT-Sham (n = 7) | SIRT1IEC-KO-Sham (n = 7) | |
|---|---|---|---|---|
| Body weight (gram) | 27.9 ± 2.4 * | 27.4 ± 3.2 | 30.8 ± 2.7 | 29.8 ± 4.6 |
| Mean arterial blood pressure (MAP, mmHg) | 92 ± 8 * | 91 ± 10 | 119 ± 14 | 112 ± 7 # |
| Heart rate (beats/min) | 458 ± 44 | 461 ± 32 | 422 ± 51 | 430 ± 55 |
| cardiac output (CO, mL/min) | 7.4 ± 0.2 * | 4.5 ± 0.2 # | 11.2 ± 0.3 | 10.9 ± 0.4 # |
| cardiac index (CI, mL.min/100 g) | 26.5 ± 1.7 * | 16.7 ± 1.5 # | 36.3 ± 2.2 | 35.9 ± 2.9 # |
| LPS binding protein (LBP) (ng/mL) | 27.8 ± 1.2 * | 34.7 ± 0.9 | 9.3 ± 0.6 | 12.4 ± 1.1 |
| Culture % rate of MLN | 43% * | 71% # | 0 | 14% |
| Name of Gene | Sequence of Sense Primer (5′-3′) | Sequence of Anti-Sense Primer (3′-5′) |
|---|---|---|
| 18S | GTAACCCGTTGAACCCCATT | CCATCCAATCGGTAGTAGCG |
| SIRT1 | GCAACAGCATCTTGCCGAT | GTGCTACTGGTCTCACTT |
| TNFR1 | TGACCCTCTCCTCTACGGA | CCATCCACCACAGCATACA |
| TNFR2 | GACTGGCGAACTGCTT | AACTGGGTGCTGTGGTCAAT |
| Nox-2 | GGGAACTGGGCTGTGAATGA | CAGTGCTGACCCAAGGAGTT |
| Nox-4 | ACAGTCCTGGCTTACCTTCG | TTCTGGGATCCTCATTCTGG |
| CuZnSOD (superoxide dismutase) | GCGGTGAACCAGTTGTGTTGTC | CCTCTGGACCCGTTACACTGAC |
| MnSOD (superoxide dismutase) | ATGTTACAACTCAGGTCGCTCTTC | CCTCTCAACGACCTCCGATAGT |
| Catalase | CCGACCAGGGCATCAAAA | GAGGCCATAATCCGGATCTTC |
| MLCK | AAT GGT GTT GCT GGA GAT CGA GGT | CTCAAAGTTACCACCGCTGCTG |
| ZO-1 | CGGGACTGTTGGTATTGGCTAGA | GGCCAGGGCCATAGTAAAGTTTG |
| Occludin | TCCTATAAATCCACGCCGGTTC | CTCAAAGTTACCACCGCTGCTG |
| E-cadherin | TCA ACG ATC CTG ACC AGC AGT TCG- | GGT GAA CCA TCA TCT GTG GCG ATG |
| KIM-1 | TGGCACTGTGACATCCTCAGA | GCAACGGACATGCCAACATA |
| IL-18 | CTTTGGAAGCCTGCTATAATCC | GGTCAAGAGGAAGTGATTTGGA |
| Lipocalin-2 | TGGCCACTTGCACATTGTAG | ATGTCACCTCCATCCTGGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, Y.-T.; Liu, T.-T.; Yang, U.-C.; Huang, C.-C.; Liu, C.-W.; Huang, S.-F.; Li, T.-H.; Liu, H.-M.; Lin, M.-W.; Yang, Y.-Y.; et al. Intestinal SIRT1 Deficiency-Related Intestinal Inflammation and Dysbiosis Aggravate TNFα-Mediated Renal Dysfunction in Cirrhotic Ascitic Mice. Int. J. Mol. Sci. 2021, 22, 1233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031233
Chou Y-T, Liu T-T, Yang U-C, Huang C-C, Liu C-W, Huang S-F, Li T-H, Liu H-M, Lin M-W, Yang Y-Y, et al. Intestinal SIRT1 Deficiency-Related Intestinal Inflammation and Dysbiosis Aggravate TNFα-Mediated Renal Dysfunction in Cirrhotic Ascitic Mice. International Journal of Molecular Sciences. 2021; 22(3):1233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031233
Chicago/Turabian StyleChou, Yu-Te, Tze-Tze Liu, Ueng-Cheng Yang, Chia-Chang Huang, Chih-Wei Liu, Shiang-Fen Huang, Tzu-Hao Li, Hsuan-Miao Liu, Ming-Wei Lin, Ying-Ying Yang, and et al. 2021. "Intestinal SIRT1 Deficiency-Related Intestinal Inflammation and Dysbiosis Aggravate TNFα-Mediated Renal Dysfunction in Cirrhotic Ascitic Mice" International Journal of Molecular Sciences 22, no. 3: 1233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031233