
A Bioassay Using a Pentadecanal Derivative to Measure S1P Lyase Activity

Abstract
:1. Introduction
2. Results
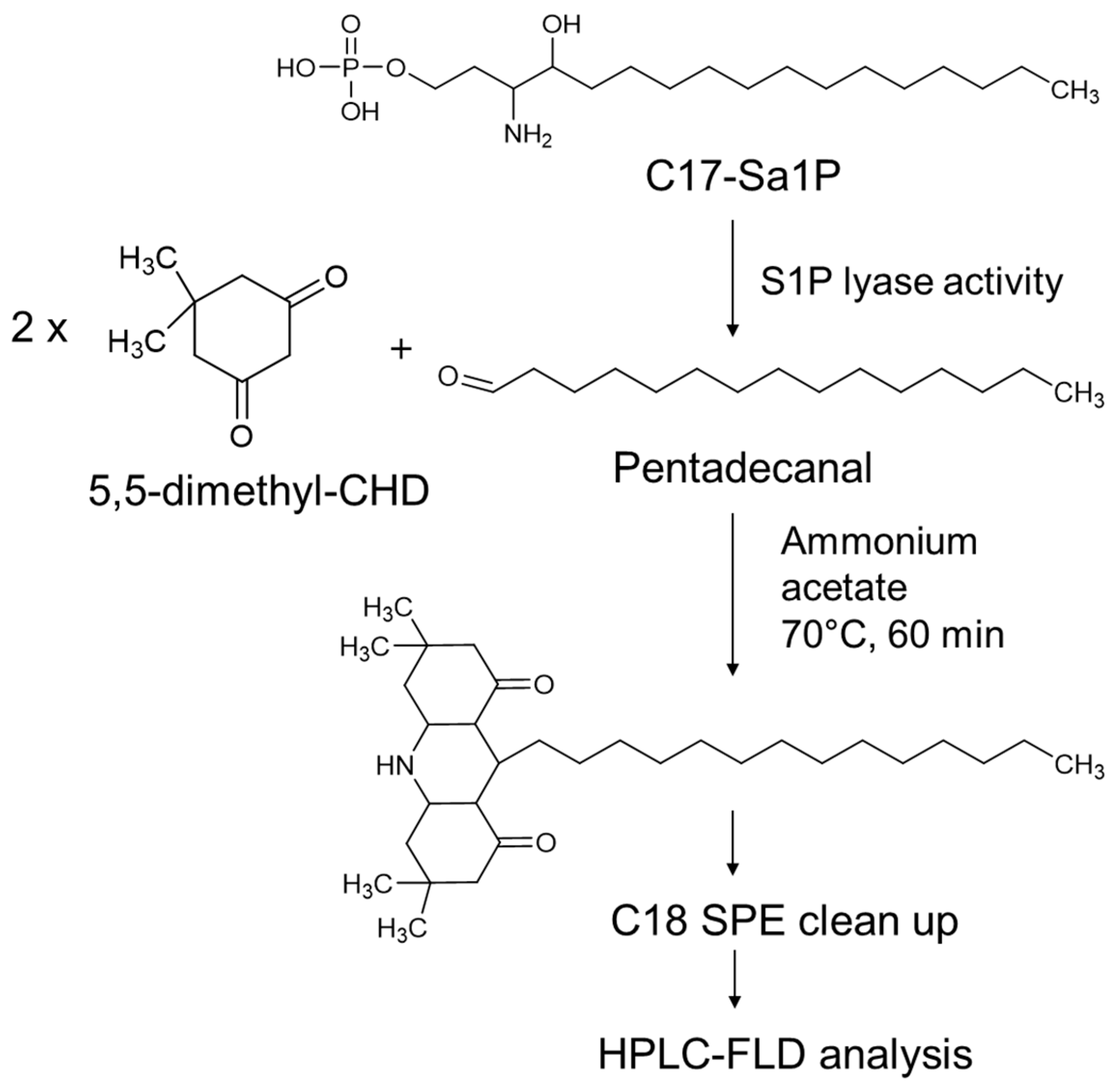
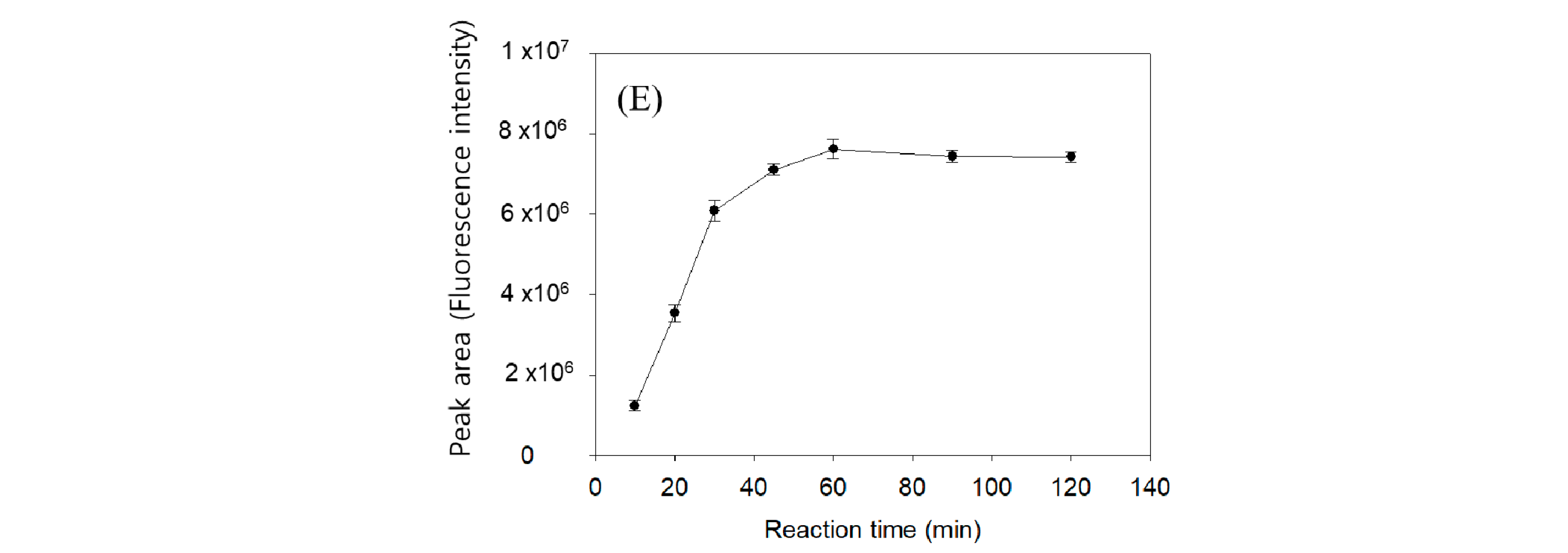
2.1. Optimization of Fluorogenic Derivatization
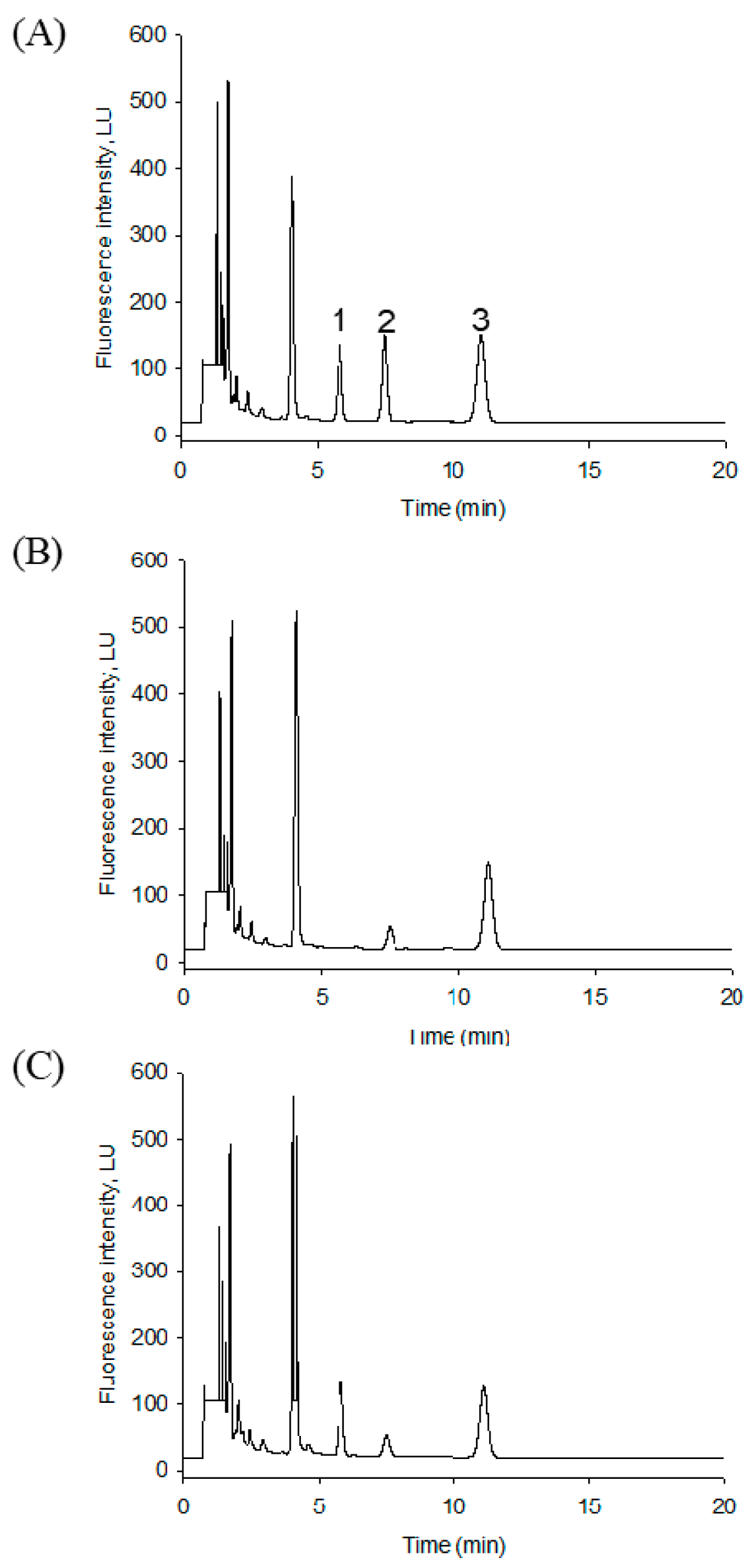
2.2. HPLC Separation of the S1PL Products
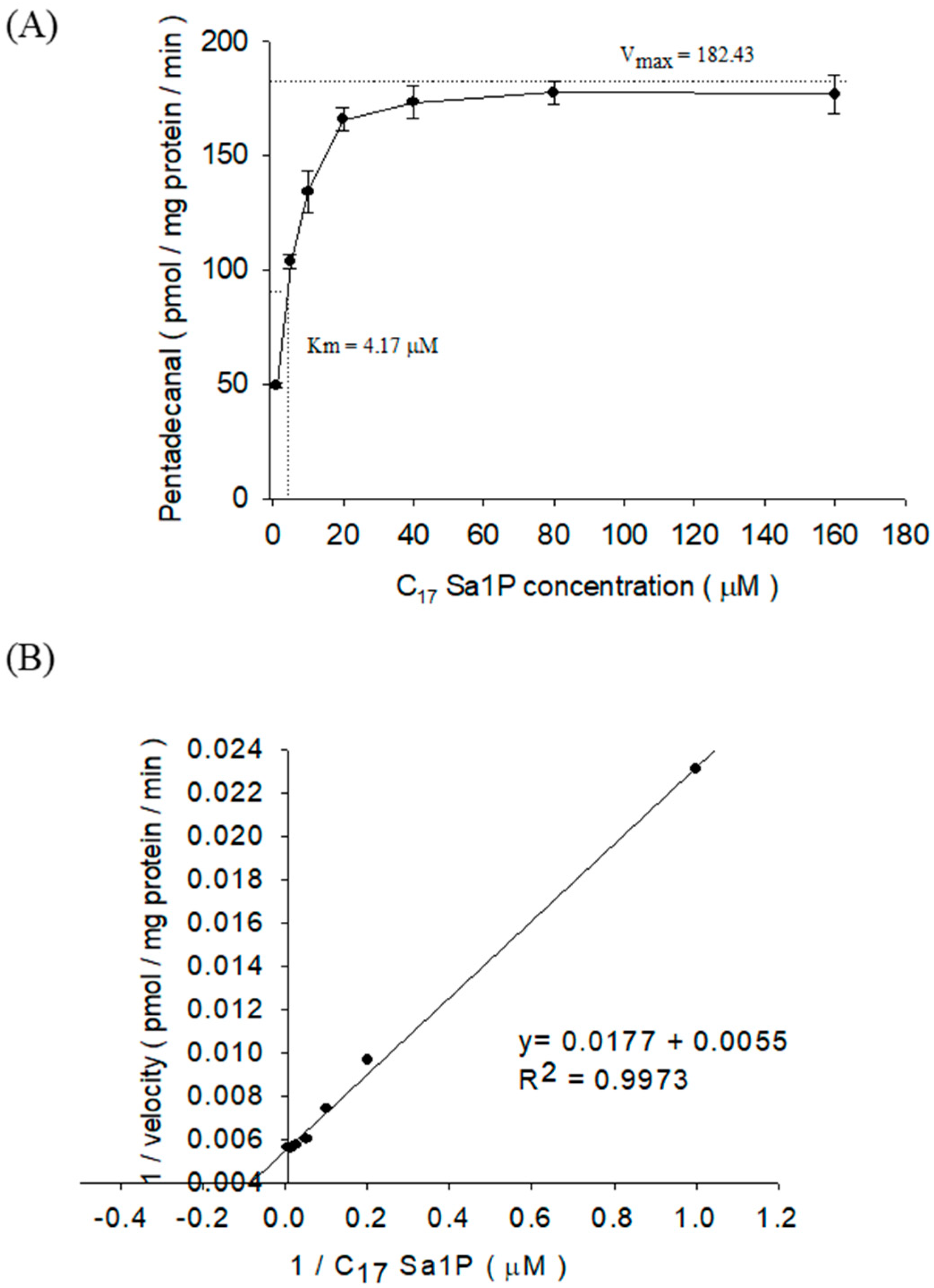
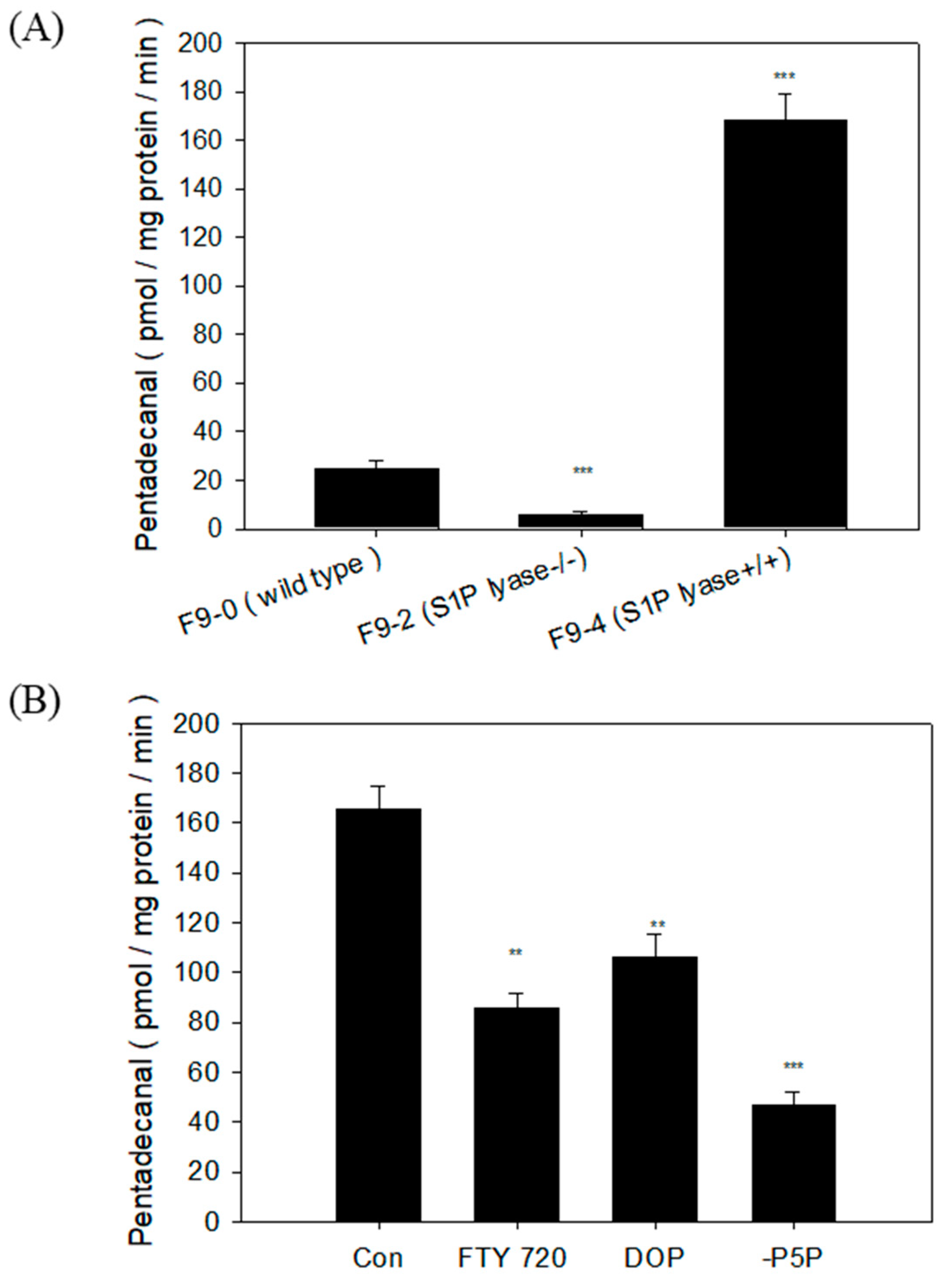
2.3. Characterization of S1PL Activity in Mutant F9 Cell Lysates
2.4. S1PL Activity in Three Mutant F9 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Preparation for S1PL Assay
4.3. Preparation of the Dimedone Reagent
4.4. S1PL Assay
4.5. Measurement of Aldehydes
4.6. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| S1P | Sphingosine-1-phosphate |
| S1PL | S1P lyase |
| Sa1P | Dihydrosphingosine-1-phosphate |
| CHD | Cyclohexanedione |
| DOP | 4-Deoxypyridoxine |
| P5P | Pyridoxal-5-phosphate |
| GPCRs | G-protein-coupled membrane receptors |
| THI | 2-Acetyl-5-tetrahydroxybutyl Imidazole |
| NBD-S1P | 7-Nitrobenz-2-oxa-1,3-diazol S1P |
| C17-Sa1P | C17-Dihydrosphingosine 1-phosphate |
References
- Wang, D.; Zhao, Z.; Caperell-Grant, A.; Yang, G.; Mok, S.C.; Liu, J.; Bigsby, R.M.; Xu, Y. S1P differentially regulates migration of human ovarian cancer and human ovarian surface epithelial cells. Mol. Cancer. Ther. 2008, 7, 1993–2002. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Chen, J.; Liu, D.; Li, W.; Wang, H.; Huang, Y.; Gao, C. HDL-S1P protects endothelial function and reduces lung injury during sepsis in vivo and in vitro. Int. J. Biochem. Cell. Biol. 2020, 126, 105819. [Google Scholar] [CrossRef] [PubMed]
- Hymel, L.A.; Ogle, M.E.; Anderson, S.E.; San Emeterio, C.L.; Turner, T.C.; York, W.Y.; Liu, A.Y.; Olingy, C.E.; Sridhar, S.; Lim, H.S.; et al. Modulating local S1P receptor signaling as regenerative immunotherapy after volumetric muscle loss injury. J. Biomed. Mater. Res. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Schwab, S.R.; Pereira, J.P.; Matloubian, M.; Xu, Y.; Huang, Y.; Cyster, J.G. Lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science 2005, 309, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Matloubian, M.; Lo, C.G.; Cinamon, G.; Lesneski, M.J.; Xu, Y.; Brinkmann, V.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 2004, 427, 355–360. [Google Scholar] [CrossRef]
- Simmons, S.; Sasaki, N.; Umemoto, E.; Uchida, Y.; Fukuhara, S.; Kitazawa, Y.; Okudaira, M.; Inoue, A.; Tohya, K.; Aoi, K.; et al. High-endothelial cell-derived S1P regulates dendritic cell localization and vascular integrity in the lymph node. Elife 2019, 8, e41239. [Google Scholar] [CrossRef]
- Leong, W.I.; Saba, J.D. S1P metabolism in cancer and other pathological conditions. Biochimie 2010, 92, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Karuppuchamy, T.; Tyler, C.J.; Lundborg, L.R.; Perez-Jeldres, T.; Kimball, A.K.; Clambey, E.T.; Jedlicka, P.; Rivera-Nieves, J. Sphingosine-1-Phosphate Lyase Inhibition Alters the S1P Gradient and Ameliorates Crohn’s-Like Ileitis by Suppressing Thymocyte Maturation. Inflamm. Bowel. Dis. 2020, 26, 216–228. [Google Scholar] [CrossRef]
- Schwiebs, A.; Herrero San Juan, M.; Schmidt, K.G.; Wiercinska, E.; Anlauf, M.; Ottenlinger, F.; Thomas, D.; Elwakeel, E.; Weigert, A.; Farin, H.F.; et al. Cancer-induced inflammation and inflammation-induced cancer in colon: A role for S1P lyase. Oncogene 2019, 38, 4788–4803. [Google Scholar] [CrossRef]
- Ebenezer, D.L.; Fu, P.; Suryadevara, V.; Zhao, Y.; Natarajan, V. Epigenetic regulation of pro-inflammatory cytokine secretion by sphingosine 1-phosphate (S1P) in acute lung injury: Role of S1P lyase. Adv. Biol. Regul. 2017, 63, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Peest, U.; Sensken, S.C.; Andreani, P.; Hanel, P.; Van Veldhoven, P.P.; Graler, M.H. S1P-lyase independent clearance of extracellular sphingosine 1-phosphate after dephosphorylation and cellular uptake. J. Cell Biochem. 2008, 104, 756–772. [Google Scholar] [CrossRef]
- Maeda, Y.; Yagi, H.; Takemoto, K.; Utsumi, H.; Fukunari, A.; Sugahara, K.; Masuko, T.; Chiba, K. S1P lyase in thymic perivascular spaces promotes egress of mature thymocytes via up-regulation of S1P receptor 1. Int. Immunol. 2014, 26, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandhuvula, P.; Honbo, N.; Wang, G.Y.; Jin, Z.Q.; Fyrst, H.; Zhang, M.; Borowsky, A.D.; Dillard, L.; Karliner, J.S.; Saba, J.D. S1P lyase: A novel therapeutic target for ischemia-reperfusion injury of the heart. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1753–H1761. [Google Scholar] [CrossRef] [PubMed]
- Kariya, Y.; Kihara, A.; Ikeda, M.; Kikuchi, F.; Nakamura, S.; Hashimoto, S.; Choi, C.H.; Lee, Y.M.; Igarashi, Y. Products by the sphingosine kinase/sphingosine 1-phosphate (S1P) lyase pathway but not S1P stimulate mitogenesis. Genes Cells 2005, 10, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Kashem, M.A.; Wa, C.; Wolak, J.P.; Grafos, N.S.; Ryan, K.R.; Sanville-Ross, M.L.; Fogarty, K.E.; Rybina, I.V.; Shoultz, A.; Molinaro, T.; et al. A high-throughput scintillation proximity assay for sphingosine-1-phosphate lyase. Assay Drug. Dev. Technol. 2014, 12, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Bandhuvula, P.; Fyrst, H.; Saba, J.D. A rapid fluorescence assay for sphingosine-1-phosphate lyase enzyme activity. J. Lipid. Res. 2007, 48, 2769–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdyshev, E.V.; Goya, J.; Gorshkova, I.; Prestwich, G.D.; Byun, H.S.; Bittman, R.; Natarajan, V. Characterization of sphingosine-1-phosphate lyase activity by electrospray ionization-liquid chromatography/tandem mass spectrometry quantitation of (2E)-hexadecenal. Anal. Biochem. 2011, 408, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Luth, A.; Neuber, C.; Kleuser, B. Novel methods for the quantification of (2E)-hexadecenal by liquid chromatography with detection by either ESI QTOF tandem mass spectrometry or fluorescence measurement. Anal. Chim. Acta 2012, 722, 70–79. [Google Scholar] [CrossRef]
- Zhao, P.P.; Zhou, X.F.; Dai, J.J.; Xu, H.J. Catalyst-free reductive amination of aromatic aldehydes with ammonium formate and Hantzsch ester. Org. Biomol. Chem. 2014, 12, 9092–9096. [Google Scholar] [CrossRef]
- Van Veldhoven, P.P.; Mannaerts, G.P. Subcellular localization and membrane topology of sphingosine-1-phosphate lyase in rat liver. J. Biol. Chem. 1991, 266, 12502–12507. [Google Scholar] [CrossRef]
- Serra, M.; Saba, J.D. Sphingosine 1-phosphate lyase, a key regulator of sphingosine 1-phosphate signaling and function. Adv. Enzyme Regul. 2010, 50, 349–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, S.; Ishii, M. Sphingosine-1-phosphate: A master regulator of lymphocyte egress and immunity. Arch. Immunol. Ther. Exp. 2014, 62, 103–115. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Spike Amount (pmol) | Intraday (RSD, %) | Interday (RSD, %) | Intraday (Accuracy, %) | Interday (Accuracy, %) |
|---|---|---|---|---|---|
| 2.5 | 7.23 | 9.12 | 93.60 ± 6.75 | 91.92 ± 8.21 | |
| 5 | 3.25 | 3.18 | 95.29 ± 3.08 | 96.07 ± 3.28 | |
| Pentadecanal | |||||
| 50 | 2.34 | 2.47 | 103.41 ± 3.08 | 102.74 ± 2.67 | |
| 400 | 1.64 | 1.78 | 98.54 ± 1.96 | 100.06 ± 2.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.-O.; Shamshiddinova, M.; Lee, J.-N.; Lee, K.-S.; Lee, Y.-M. A Bioassay Using a Pentadecanal Derivative to Measure S1P Lyase Activity. Int. J. Mol. Sci. 2021, 22, 1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031438
Shin K-O, Shamshiddinova M, Lee J-N, Lee K-S, Lee Y-M. A Bioassay Using a Pentadecanal Derivative to Measure S1P Lyase Activity. International Journal of Molecular Sciences. 2021; 22(3):1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031438
Chicago/Turabian StyleShin, Kyong-Oh, Maftuna Shamshiddinova, Jung-No Lee, Kwang-Sik Lee, and Yong-Moon Lee. 2021. "A Bioassay Using a Pentadecanal Derivative to Measure S1P Lyase Activity" International Journal of Molecular Sciences 22, no. 3: 1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031438