
Characterization and Reconstitution of Human Lipoyl Synthase (LIAS) Supports ISCA2 and ISCU as Primary Cluster Donors and an Ordered Mechanism of Cluster Assembly

Abstract
:1. Introduction
2. Results
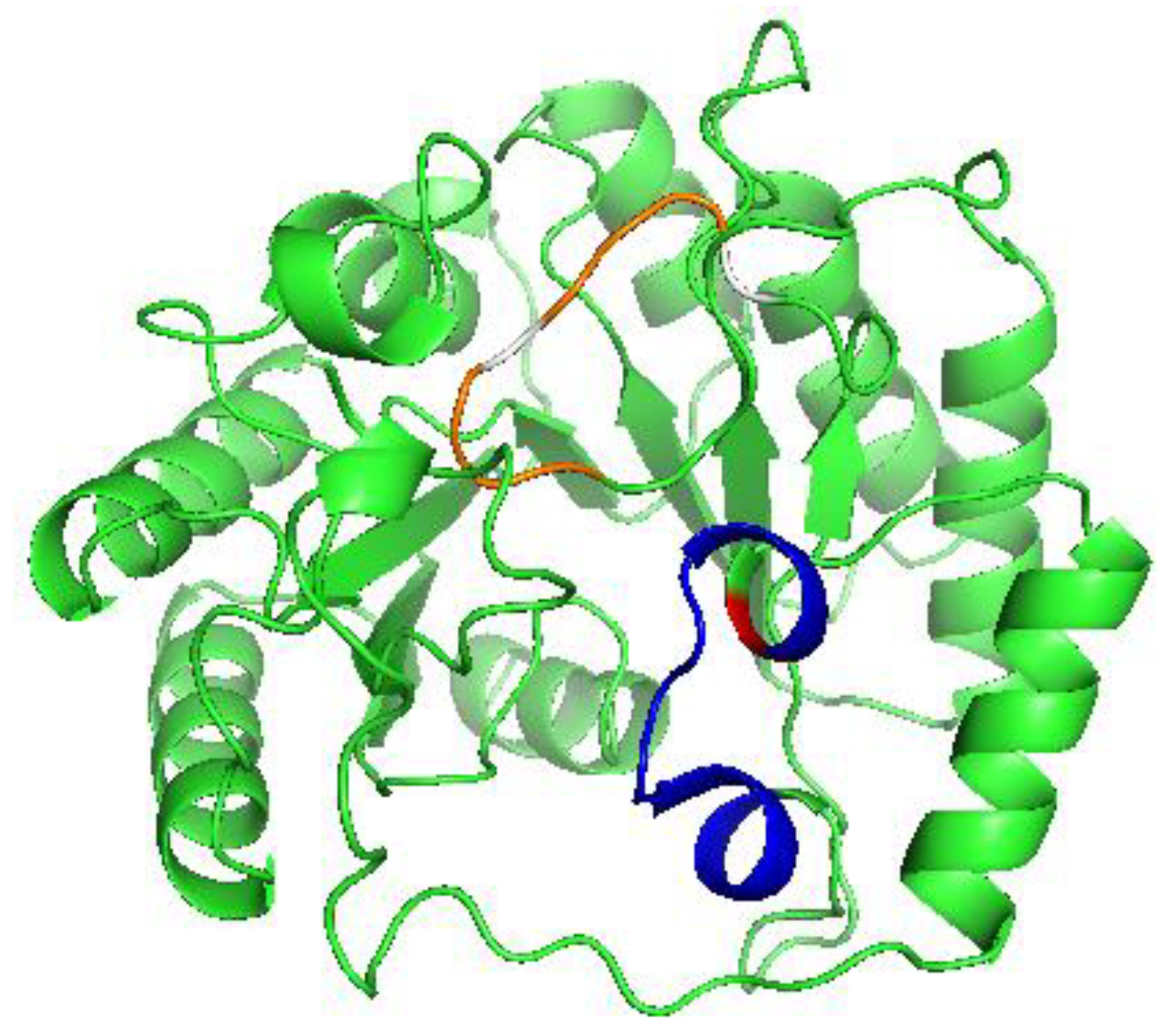
2.1. Characterization of Native Recombinant and Substituted Derivatives of LIAS
2.2. Evaluation of Cluster Donors for LIAS Reconstitution
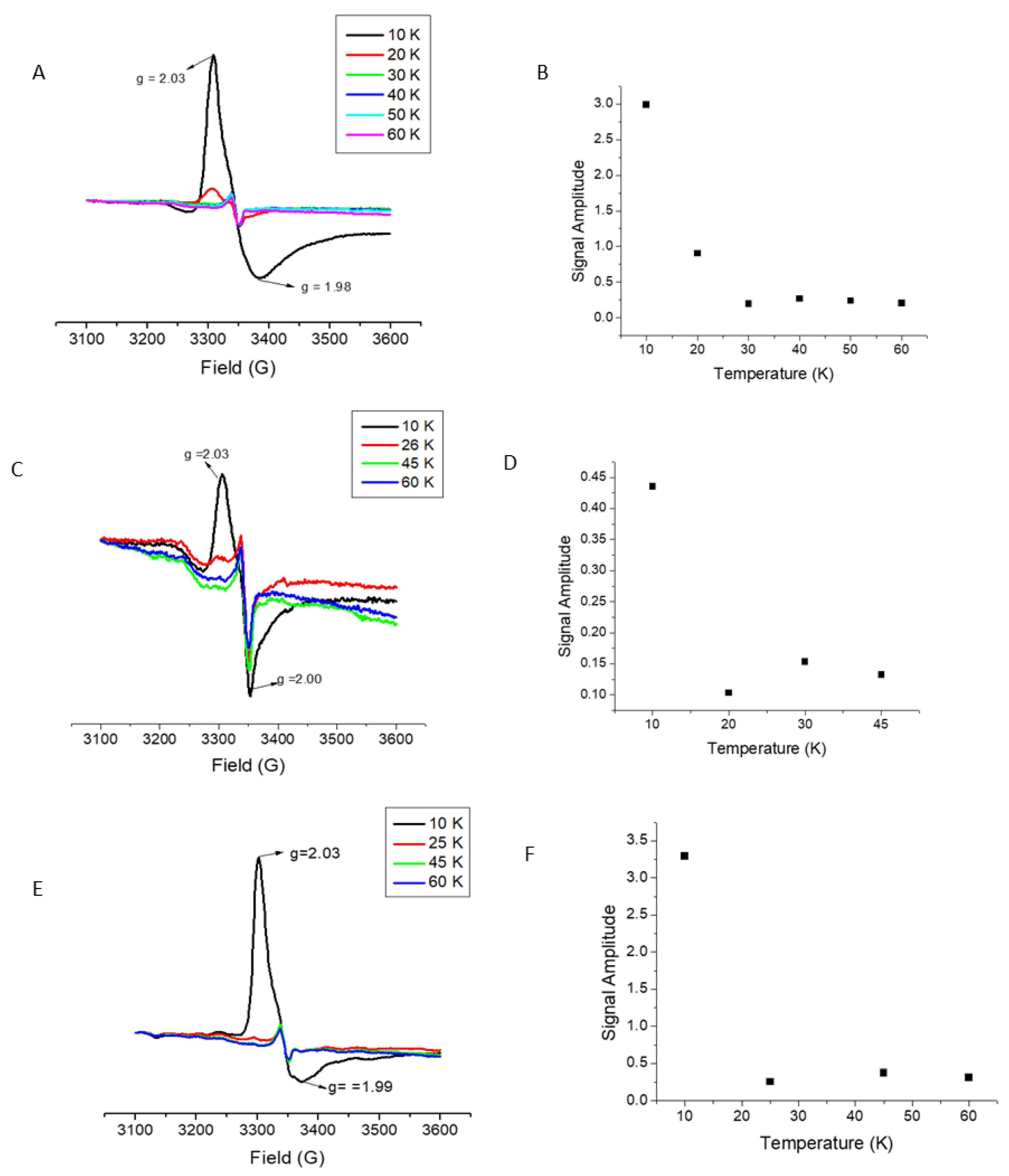
2.3. [Fe–S] Cluster Transfer Monitored via UV–Vis
2.4. Cluster Delivery to LIAS with Substitutions of Reducing and Auxiliary Cluster Ligands
3. Discussion
4. Materials and Methods
4.1. Purchased Materials
4.2. Expression of LIAS
4.3. Site Directed Mutagenesis
4.4. Purification of LIAS
4.5. Expression and Purification of Donor Proteins
4.6. Iron Quantitation
4.7. UV–Vis Characterization
4.8. LC–MS Activity Assays
4.9. Preparation of Electron Paramagnetic Resonance (EPR) Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Packer, L.; Witt, E.H.; Tritschler, H.J. Alpha-Lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Bourguignon, J.; Neuburger, M.; Douce, R. Resolution and characterization of the glycine-cleavage reaction in pea leaf mitcochondria. Biochem. J. 1988, 255, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; DeBusk, B.G. Chemical Nature of an α-lipoic acid conjugate required for oxidation of pyruvate and α-ketoglutarate by an Escherichia Coli mutant. J. Biol. Chem. 1952, 199, 881–888. [Google Scholar] [CrossRef]
- Reed, L.J.; Koike, M.; Levitch, M.E.; Leach, F.R. Studies on the nature and reactions of protein-bound lipoic acid. J. Biol. Chem. 1958, 232, 143–158. [Google Scholar] [CrossRef]
- Lu, W.F.; Cao, J.J.; Guo, Y.J.; Zhong, K.; Zha, G.M.; Wang, L.F.; Yang, G.Y. Expression of the porcine lipoic acid synthase (LIAS) gene in Escherichia coli. Genet. Mol. Res. 2014, 13, 5369–5377. [Google Scholar] [CrossRef]
- Camponeschi, F.; Muzzioli, R.; Ciofi-Baffoni, S.; Piccioli, M.; Banci, L. Paramagnetic (1)H NMR Spectroscopy to Investigate the Catalytic Mechanism of Radical S-Adenosylmethionine Enzymes. J. Mol. Biol. 2019, 431, 4514–4522. [Google Scholar] [CrossRef]
- Miller, J.R.; Busby, R.W.; Jordan, S.W.; Cheek, J.; Henshaw, T.F.; Ashley, G.W.; Broderick, J.B.; Cronan, J.E., Jr.; Marletta, M.A. Escherichia coli LipA Is a Lipoyl Synthase: In Vitro Biosynthesis of Lipoylated Pyruvate Dehydrogenase Complex from Octanoyl-Acyl Carrier Protein. Biochemistry 2000, 39, 15166–15178. [Google Scholar] [CrossRef]
- McCarthy, E.; Booker, S. Destruction and reformation on an iron-sulfur cluster during catalysis by lipoyl synthase. Science 2017, 358, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Cicchillo, R.M.; Lee, K.H.; Baleanu-Gogonea, C.; Nesbitt, N.M.; Krebs, C.; Booker, S.J. Escherichia coli Lipoyl Synthase Binds Two Distinct [4Fe-4S] Clusters per Polypeptide. Biochemistry 2004, 43, 11770–11778. [Google Scholar] [CrossRef] [PubMed]
- Ollagnier-de Choudens, S.; Sanakis, Y.; Hewitson, K.; Roach, R.; Munck, E.; Fontecave, M. Reductive Cleavage of S-Adenosylmethionine by Biotin Synthase from Esherichia coli. J. Biol. Chem. 2002, 277, 13449–13454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieder, K.W.; Booker, S.; Ruzicka, F.J.; Beinert, H.; Reed, G.H.; Frey, P.A. S-Adenosylmethionine-Dependent Reduction of Lysine 2,3-Aminomutase and Observation of the Catalytically Functional Iron-Sulfur Centers by Electron Paramagnetic Resonance. Biochemistry 1998, 37, 2578–2585. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.I.; Lanz, N.D.; Goldman, P.J.; Lee, K.-H.; Booker, S.J.; Drennan, C.L. Crystallographic snapshots of sulfur insertion by lipoyl synthase. Proc. Natl. Acad. Sci. USA 2016, 113, 9446–9450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicchillo, R.M.; Iwig, D.; Jones, A.D.; Nesbitt, N.M.; Baleanu-Gogonea, C.; Souder, M.G.; Tu, L.; Booker, S.J. Lipoyl Synthase Requires Two Equivalents of S-Adenosyl-L-Methionine to Synthesize One Equivalent of Lipoic Acid. Biochemistry 2004, 43, 6378–6386. [Google Scholar] [CrossRef] [PubMed]
- Lanz, N.D.; Pandelia, M.E.; Kakar, E.S.; Lee, K.H.; Krebs, C.; Booker, S.J. Evidence for a Catalytically and Kinetically Competent Enzyme− Substrate Cross-Linked Intermediate in Catalysis by Lipoyl Synthase. Biochemistry 2014, 53, 4557–4572. [Google Scholar] [CrossRef]
- Harmer, J.E.; Hisox, M.J.; Dinis, P.C.; Fox, S.J.; Iliopoulos, A.; Hussey, J.E.; Sandy, J.; Van Beek, F.T.; Wessex, J.W.; Roach, P.L. Structures of Lipoyl Synthase Reveal a Compact Active Site for Controlling Sequential Sulfur Insertion Reactions. Biochem. J. 2014, 464, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Wachnowsky, C.; Fidai, I.; Cowan, J.A. Iron-sulfur cluster biosynthesis and trafficking-impact on human disease conditions. Metallomics 2018, 10, 9–29. [Google Scholar] [CrossRef]
- Cameron, J.M.; Levandovskiy, V.; MacKay, N.; Rouault, T.; Tong, W.; Oglive, I.; Shoubridge, E.A.; Robinson, B.H. Mutations in Iron-Sulfur Cluster Scaffold Genes NFU1 and BOLA3 Cause a Fatal Deficiency of Multiple Respiratory Chain and 2-Oxoacid Dehydrogenase Enzymes. Am. J. Hum. Genet. 2011, 89, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Sastre, A.; Tort, F.; Stehling, O.; Uzarska, M.A.; Arranz, J.A.; Del Toro, M.; Labayru, M.T.; Landa, J.; Font, A.; Garcia-Villoria, J.; et al. A Fatal Mitochondrial Disease Is Associated with Defective NFU1 Function in the Maturation of a Subset of Mitochondrial Fe-S Proteins. Am. J. Hum. Genet. 2011, 89, 656–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, P.R., II; Friederich, M.W.; Swanson, M.A.; Shaikh, T.; Bhattacharya, K.; Scharer, G.H.; Aicher, J.; Creadon-Swindell, G.; Geiger, E.; MacLean, K.N.; et al. Variant non-ketotic hyperglycinemia is caused by mutations in LIAS, BOLA3 and the novel gene GLRX5. Brain 2014, 137, 366–379. [Google Scholar] [CrossRef] [Green Version]
- Mayr, J.A.; Zimmerman, F.A.; Fauth, C.; Bergheim, C.; Merierhofer, D.; Radmayr, D.; Zschocke, J.; Koch, J.; Sperl, W. Lipoic Acid Synthetase Deficiency Causes Neonatal—Onset Epilepsy, Defective Mitochondrial Energy Metabolism, and Glycine Elevation. Am. J. Hum. Genet. 2011, 89, 792–797. [Google Scholar] [CrossRef] [Green Version]
- Muhlenhoff, U.; Richter, N.; Pines, O.; Pierik, A.J.; Lill, R. Specialized Function of Yeast Isa1 and Isa2 Proteins in the Maturation of Mitochondrial [4Fe-4S] Proteins. J. Biol. Chem. 2011, 286, 41205–41216. [Google Scholar] [CrossRef] [Green Version]
- Beilschmidt, L.K.; de Choudens, S.O.; Fournier, M.; Sanakis, I.; Hograindleur, M.A.; Clemancey, M.; Blondin, G.; Schmucker, S.; Eisenmann, A.; Weiss, A.; et al. ISCA1 is essential for mitochondrial Fe4S4 biogenesis in vivo. Nat. Commun. 2017, 8, 1–2. [Google Scholar] [CrossRef]
- Banci, L.; Brancaccio, D.; Ciofi-Baffoni, S.; Del Conte, R.; Gadepalli, R.; Mikolajczyk, M.; Neri, S.; Piccioli, M.; Winkelmann, J. [2Fe-2S] cluster transfer in iron-sulfur protein biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 6203–6208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachnowsky, C.; Hendricks, A.L.; Wesley, N.A.; Ferguson, C.; Fidai, I.; Cowan, J.A. Understanding the Mechanism of [4Fe-4S] Cluster Assembly on Eukaryotic Mitochondrial and Cytosolic Aconitase. Inorg. Chem. 2019, 58, 13686–13695. [Google Scholar] [CrossRef] [PubMed]
- Uzarska, M.A.; Nasta, V.; Weiler, B.D.; Spantgar, F.; Ciofi-Baffoni, S.; Saviello, M.R.; Gonnelli, L.; Muhlenhoff, U.; Banci, L.; Lill, R. Mitochondrial Bol1 and Bol3 function as assembly factors for specific iron-sulfur proteins. eLife 2016, 5, e16673. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Rao, B.; Wachnowsky, C.; Cowan, J.A. Cluster exchange reactivity of [2Fe-2S] cluster-bridged complexes of BOLA3 with monothiol glutaredoxins. Metallomics 2018, 10, 1282–1290. [Google Scholar] [CrossRef]
- Nasta, V.; Giachetti, A.; Ciofi-Baffoni, S.; Banci, L. Structural insights into the molecular function human [2Fe-2S] BOLA1-GRX5 and [2Fe-2S] BOLA3-GRX5 complexes. BBA Gen Subj. 2017, 1861, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Stehling, O.; Lill, R. The Role of Mitochondria in Cellular Iron-Sulfur Protein Biogenesis: Mechanisms, Connected Processes, and Diseases. CSH Perspect. Biol. 2013, 5, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, E.L.; Rankin, A.N.; Dill, Z.R.; Booker, S.J. The A-type domain in Escherichia coli NfuA is required for regenerating the auxiliary [4Fe-4S] cluster in Escherichia coli lipoyl synthase. J. Biol. Chem. 2019, 294, 1609–1617. [Google Scholar] [CrossRef] [Green Version]
- Kuma’novics, A.; Chen, O.S.; Li, L.; Bagley, D.; Adkins, E.M.; Lin, H.; Dingra, N.N.; Outten, C.E.; Keller, G.; Winge, D.; et al. Identification of FRA1 and FRA2 as Genes Involved in Regulating the Yeast Iron Regulon in Response to Mitochondrial Iron-Sulfur Cluster Synthesis. J. Biol. Chem. 2008, 283, 10276–10286. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cowan, J.A. Iron sulfur biosythesis. Human NFU mediates sulfide delivery to ISU in the final step of of [2Fe-2S] cluster assembly. Chem. Comm. 2007, 3192–3194. [Google Scholar]
- Wachnowsky, C.; Fidai, I.; Cowan, J.A. Iron-sulfur cluster exchange reactions mediated by the human Nfu protein. J. Biol. Inorg. Chem. 2016, 21, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Nuth, M.; Yoon, T.; Cowan, J.A. Iron-sulfur cluster biosynthesis: Characterization of iron nucleation sites for assembly of the [2Fe-2S]2+ cluster core in IscU proteins. J. Am. Chem. Soc. 2002, 124, 8774–8775. [Google Scholar] [CrossRef]
- Mansy, S.S.; Xiong, Y.; Hemann, C.; Hille, R.; Sundaralingam, M.; Cowan, J.A. Crystal structure and stability studies of C77S HiPIP: A serine ligated [4Fe-4S] cluster. Biochemistry 2002, 41, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Fidai, I.; Wachnowsky, C.; Cowan, J.A. Glutathione-complexed [2Fe-2S] clusters function in Fe-S cluster storage and trafficking. J. Biol. Inorg. Chem. 2016, 21, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Olive, J.A.; Cowan, J.A. Role of the HSPA9/HSC20 chaperone pair in promoting directional human iron-sulfur cluster exchange involving monothiol glutaredoxin 5. J. Inorg. Biochem. 2018, 184, 100–107. [Google Scholar] [CrossRef]
- Foster, M.W.; Sheref, S.S.; Hwang, J.; Penner-Hahn, J.E.; Surerus, K.K.; Cowan, J.A. A Mutant Human IscU Protein contains a Stable [2Fe-2S]2+ Center of Possible Functional Significance. J. Am. Chem. Soc. 2000, 122, 6805–6806. [Google Scholar] [CrossRef]
- Wu, S.P.; Wu, G.; Surerus, K.K.; Cowan, J.A. Iron-sulfur cluster biosynthesis. Kinetic analysis of [2Fe-2S] cluster transfer from holo ISU to apo Fd: Role of redox chemistry and a conserved aspartate. Biochemistry 2002, 41, 8876–8885. [Google Scholar] [CrossRef]
- Moulis, J.M.; Meyer, J. Characterization of the selenium-substituted 2 [4Fe-4Se] ferredoxin from Clostridium pasteurianum. Biochemistry 1982, 21, 4762–4771. [Google Scholar] [CrossRef]
- Wachnowsky, C.; Fidai, I.; Cowan, J.A. Cytosolic iron-sulfur cluster transfer-a proposed kinetic pathway for reconstitution of glutaredoxin 3. FEBS Lett. 2016, 590, 4531–4540. [Google Scholar] [CrossRef] [Green Version]
- Fidai, I.; Wachnowsky, C.; Cowan, J.A. Mapping cellular Fe-S cluster uptake and exchange reactions—Divergent pathways for iron-sulfur cluster delivery to human ferredoxins. Metallomics 2016, 8, 1283–1293. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| [2Fe–2S] Cluster Donor | Apparent Second Order Rate Constants |
|---|---|
| H. sapiens ISCA1 | no transfer |
| H. sapiens ISCA2 | 3400 ± 200 M−1min−1 |
| H. sapiens ISCA2 (truncated) | 1130 ± 70 M−1min−1 |
| H. sapiens NFU | no transfer |
| H. sapiens BOLA3 | negligible transfer |
| H. sapiens ISCU | 780 ± 50 M−1min−1 |
| H. sapiens GLRX5 | no transfer |
| H. sapiens Glrx5/BOLA3 heterodimer | no transfer |
| S. cerevisiae GLRX3 | no transfer |
| H. sapiens Nfu/ISCA2 heterodimer | no transfer |
| H. sapiens ISCA1/ISCA2 heterodimer | no transfer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hendricks, A.L.; Wachnowsky, C.; Fries, B.; Fidai, I.; Cowan, J.A. Characterization and Reconstitution of Human Lipoyl Synthase (LIAS) Supports ISCA2 and ISCU as Primary Cluster Donors and an Ordered Mechanism of Cluster Assembly. Int. J. Mol. Sci. 2021, 22, 1598. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041598
Hendricks AL, Wachnowsky C, Fries B, Fidai I, Cowan JA. Characterization and Reconstitution of Human Lipoyl Synthase (LIAS) Supports ISCA2 and ISCU as Primary Cluster Donors and an Ordered Mechanism of Cluster Assembly. International Journal of Molecular Sciences. 2021; 22(4):1598. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041598
Chicago/Turabian StyleHendricks, Amber L., Christine Wachnowsky, Brian Fries, Insiya Fidai, and James A. Cowan. 2021. "Characterization and Reconstitution of Human Lipoyl Synthase (LIAS) Supports ISCA2 and ISCU as Primary Cluster Donors and an Ordered Mechanism of Cluster Assembly" International Journal of Molecular Sciences 22, no. 4: 1598. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041598