1. Introduction
Prostate cancer (PCa) is one of the most common cancers in men. It is the second-leading cause of death for men in the US [
1], and although, for 2020 compared to the last five years, its incidence has decreased, it remains the third-leading cause of death in Europe [
2]. Advanced age, ethnicity, genetic factors and family history are well-established risk factors for PCa [
3,
4], while diet, obesity, physical inactivity, hyperglycemia and environmental pollution are positively associated with prostate cancer [
5,
6]. Usually, monitoring of the plasmatic prostate-specific antigen (PSA) level in the blood is used for the check-up of the health state of the prostate, and if the value is higher than 4 ng/mL, it is a possible risk factor for prostate cancer [
7]. Since high PSA values do not necessary correspond to prostate cancer, a gland biopsy is a mandatory step. The correct staging of prostate cancer represents the starting point to establish which is the best therapeutic strategy, to obtain information about the prognosis and to compare the results of the various therapeutic options. Multiple clinical and biopsy parameters can contribute to staging, especially if added within specific nomograms or through predictive models [
8].
The standard treatment of prostate cancer has different objectives, depending on the anatomical extension and aggressiveness of the disease but, also, on the patient’s life expectancy and the presence of comorbidities that may represent a risk of death higher than that represented by prostate cancer. For this reason, a “watchful waiting” policy (surveillance in the absence of systematic checks) may be indicated in patients with a short life expectancy (generally, less than 10 years). Similarly, patients suffering from a very low low-risk disease, even in the presence of a good life expectancy, can be directed towards an “active surveillance policy”. In patients with metastatic disease, palliation remains the most concretely achievable goal, especially if symptomatic. For these patients, there are currently various hormone therapy (LH-RHa (luteinizing hormone-releasing hormone agonist) ± nonsteroidal antiandrogen ± Docetaxel, LH-RHa + Abiraterone and LH-RHa antagonist) [
9,
10,
11,
12,
13,
14] and chemotherapy (Docetaxel) [
15,
16,
17] options that, together with the most recent forms of radiometabolic therapy (alpha emitters) and bone-targeted therapies, can significantly impact both their quality and their life expectancy. Prostate cancer has long-been considered a low chemo-sensitive tumor, but in the early 2000s, some controlled studies demonstrated the effectiveness of Docetaxel in patients suffering from castration-resistant disease (CRPC) [
18]. For patients affected by CRPC who progress after first-line treatment with deprivation androgenic (ADT), the treatment options available have increased considerably within the past few years and include new chemotherapeutics, in addition to Docetaxel [
19,
20], new hormonal therapies [
21], radio-compounds (Radium-223) [
22,
23] and immunological therapies (Sipuleucel-T, a dendritic cell vaccine) [
24]. However, immunotherapy, with checkpoint inhibitors such as ipilimumab and nivolumab [
25] has so far produced disappointing results in the treatment of prostate cancer, while the phase II study KEYNOTE-199, the results of which were presented at ASCO 2018, would seem to highlight a good therapeutic activity of pembrolizumab monotherapy [
26].
In order to identify a personalized therapy based on the specific characteristics of patients with prostate cancer, an important issue to consider is that PCa shows a high grade of genomic variegation, with different patterns and clinical implications. Usually, the genomic alteration occurs in the early stage of the tumor and accumulates as it progresses; therefore, is possible to distinguish different molecular subtypes of PCa on the basis of gene fusion, gene expression signature and other molecular alteration, but this classification does not correspond to an accurate and precise tumor staging or predictive/prognostic information [
27]. TMPRSS2-ERG fusion is the most common molecular alteration in localized PCa, with a frequency of 40–50% of all prostate cancer diagnosed [
28]. However, other gene fusions are grouped in the ETS-positive subclass. This includes fusions with ETS transcription family genes like ETV1 (10%), ETV4, ETV5 and FLI1 (1–5%). ETS-negative prostate cancer shows recurrent mutations in the SPOP, FOXA1 and IDH genes. Given the high molecular heterogeneity of prostate cancer, a series of both ETS-positive/negative subclasses are generally classified as “others”. In this macro-category, the less recurrent, usually with an unknown molecular meaning, are grouped [
29].
The integrated use of an advanced genome analysis has allowed to recognize and to identify ncRNA, which, even if they do not encode for proteins, have specific biological functions in cancerogenesis and metastasis. Among ncRNAs, the aberrant expression of some long non-coding RNA (lncRNAs) are also correlated with the disease state for PCa [
30,
31] and with a possible role in competing endogenous RNAs (ceRNAs) [
32], suggesting them a role as targets for therapeutic intervention. Long non-coding RNAs are RNAs transcripts >200 nucleotides in length [
33], with a role in cellular differentiation [
34] and in cancer pathway [
35] due to the influence of specific gene expression targets and could undergo a post-transcriptional processing to produce numerous 5′-cappelled small RNA [
36].
The therapeutic approaches against lncRNAs could be direct by affecting lncRNA expression or indirect by targeting protein-coding genes dysregulated by the lncRNA with a consequent perturbation of its molecular pathway and lethal effects on cancer cells [
37]. Briefly, the strategies to target lncRNA encompass the repression of lncRNA transcription by utilizing DNA-binding elements that target its genomic locus, the silencing of the lncRNA to induce transcript degradation, the utilization of small molecules thatmask the binding site for lncRNAs, thereby disrupting the network of interactions responsible for the altered function in disease-related lncRNA, and the utilization of aptamers, which antagonize the lncRNA association with binding partners by folding into a three-dimensional structure with higher affinity and specificity for the same regions [
38]. However, in our opinion, the not well-understood plethora of proteins and, consequently, of cellular pathways that each lncRNA could affect, considering also their ubiquitous presence, exclude the possibility to directly target lncRNAs for the biological effects not easy predictable and perhaps potentially dangerous for noncancer cells.
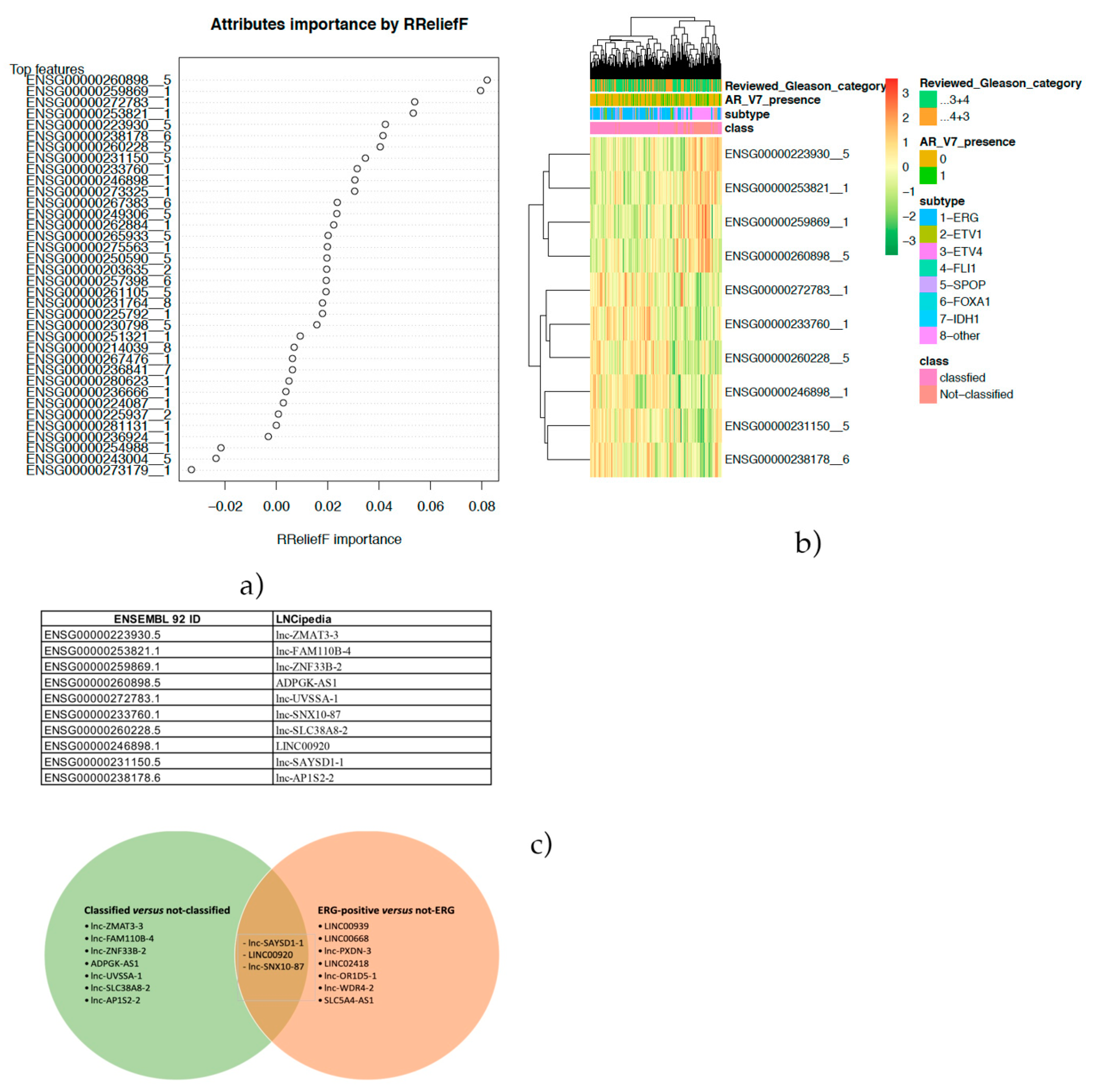
In the present paper, a feature selection approach was applied to a TCGA-PRAD cohort to identify lncRNA able to stratify ERG-positive cases and other subtypes. Moreover, the same approach was also used to focus on the cases with no peculiar genomic alterations.
3. Discussion
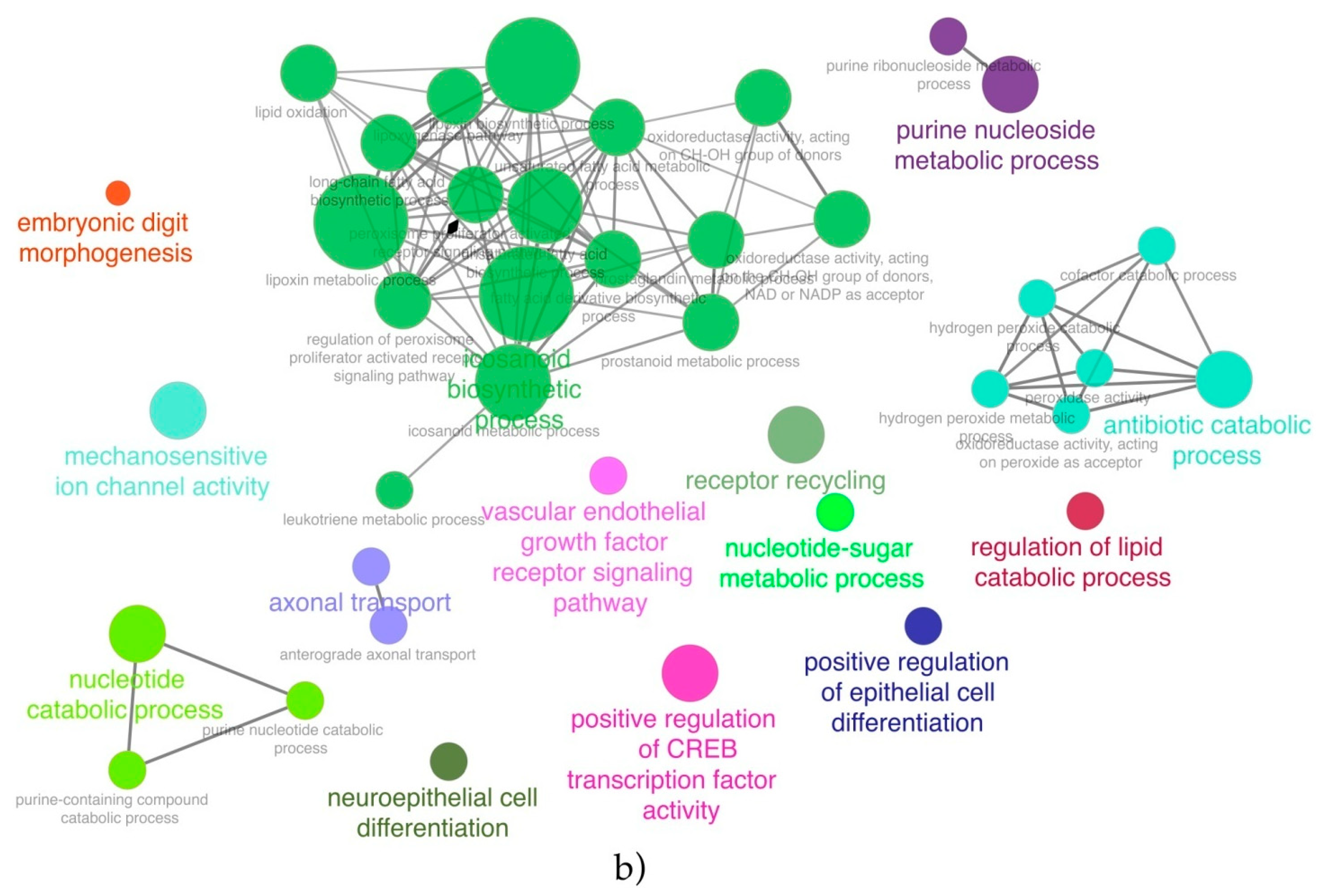
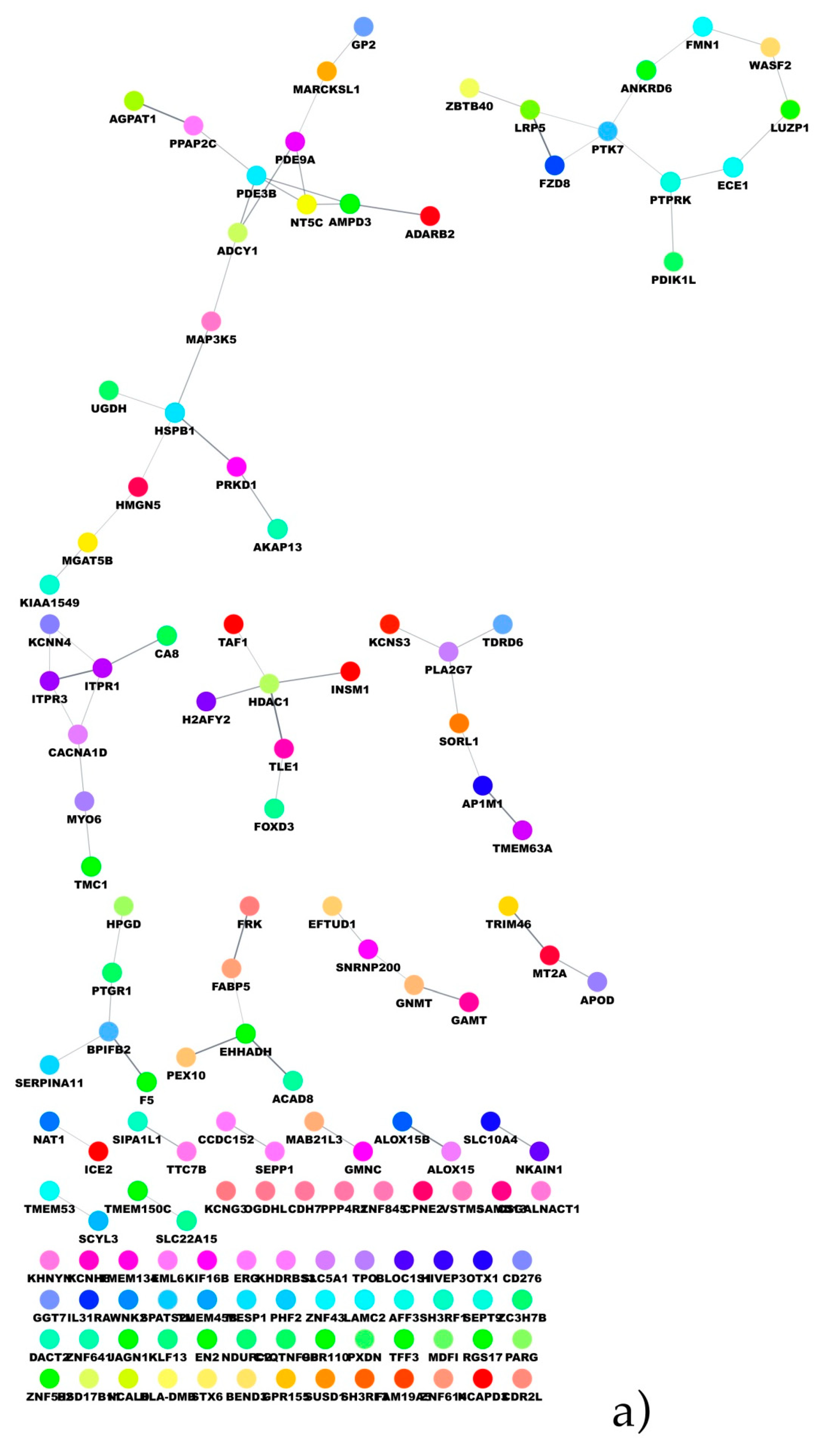
In the present paper, through the application of a feature selection approach, a set of lncRNAs was identified as able to discriminate ERG-positive from other molecular subtypes and classified from not-classified. The chance to gain mechanistic insights on the biological role of the selected lncRNAs was explored through a co-expression analysis and the set-up of a biological network. Such an aspect was explored both by performing a functional enrichment of the co-expression network and the build-up of the PPI network. Moreover, the great amount of molecular data opened the chance to better stratify patients in terms of prognostic/diagnostic predictions.
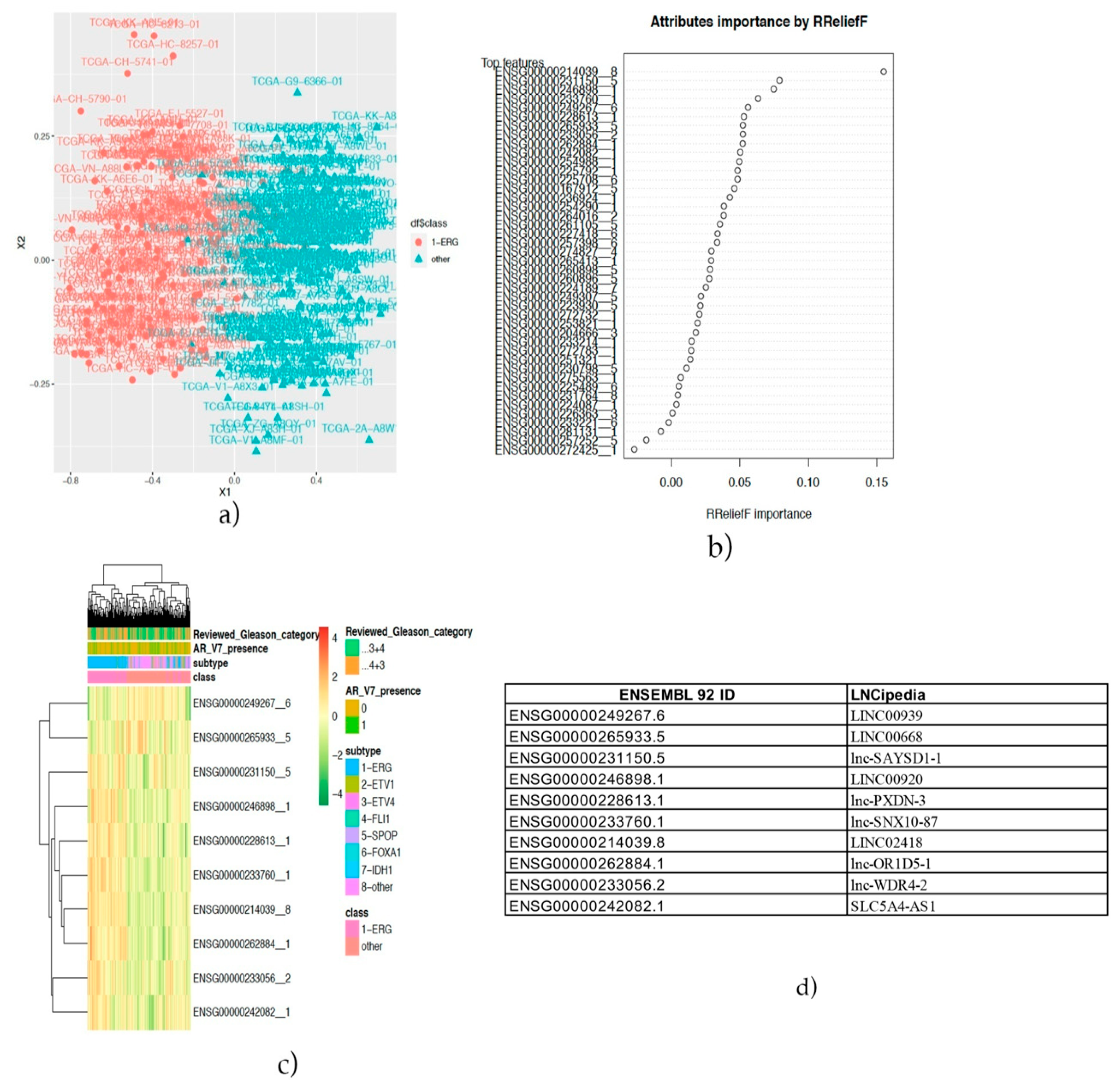
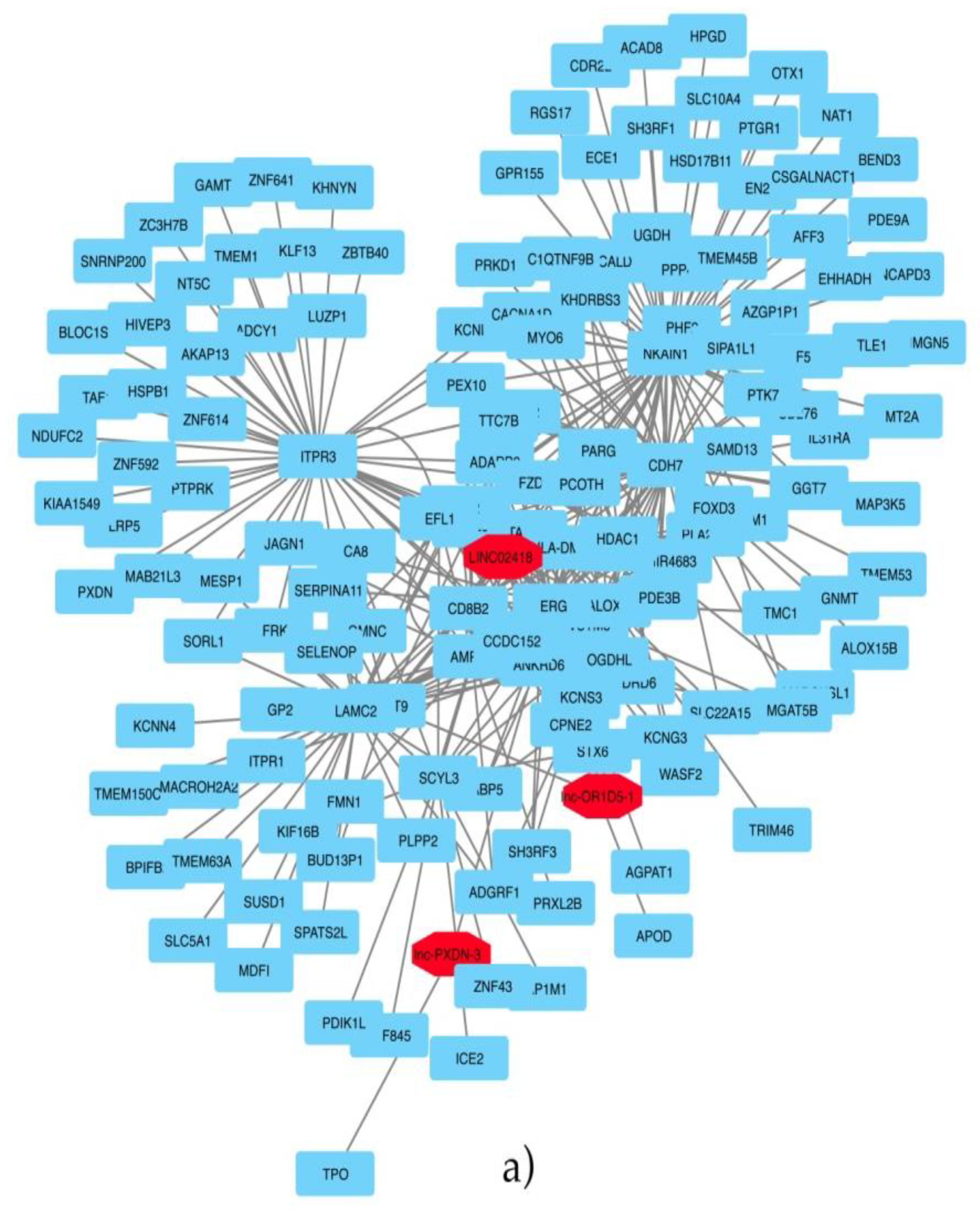
The identification of the three annotated lncRNAs LINC02418, Lnc-OR1D5-1 and Lnc-PXDN-3 discriminating the ERG-positive subtype of prostate cancer vs. others and the analysis of the co-expressed protein-coding genes and these lncRNAs revealed novel promising pharmacological targets in specific subtypes of prostate cancer. The first of them, LINC02418, we found overexpressed in the ERG-positive subtype of prostate cancer and, also, differentially expressed between colorectal cancer (CRC) tissues and noncancerous tissues [
39] and upregulated in NSCLC (non-small cell lung cancer) tissues [
40]. Both authors reported an active role for this lncRNA in tumorigenesis and in a CRC (colorectal cancer) model, and interestingly, Zhao demonstrated that LINC02418 upregulated maternal embryonic leucine zipper kinase (MELK) expression by acting as a ceRNA, which absorbs miR-1273g-3p [
40,
41]. MELK is involved in cancer cell survival and invasiveness and has been already suggested as a novel potential therapeutic target in prostate cancer [
42]. Thus, in ERG-positive high-grade tumors (Gleason greater than 4+3) [
42,
43,
44], expressing a high level of LINC02418, the MELK inhibitor OTS167 could be a promising therapeutic opportunity, because it is already utilized in clinical trials for the treatment of breast cancer and onco-hematological pathologies [
45]. Another speculation, to hypothesize a novel therapeutic strategy in positive ERG prostate cancer patients, was based on the analysis of the co-expressed protein-coding genes and lncRNAs. The upregulation of the Wnt pathway and its inhibition emerged as a promising strategy against prostate cancer [
46], although thinking about the most appropriate approach is imperative for the pervasive role of the Wnt pathway in normal tissue homeostasis. Several approaches have been taken into account for inhibiting the Wnt pathway, such as LGK974, a drug that targets the Wnt-specific acyltransferase porcupine, and the tankyrase inhibitor XAV939 [
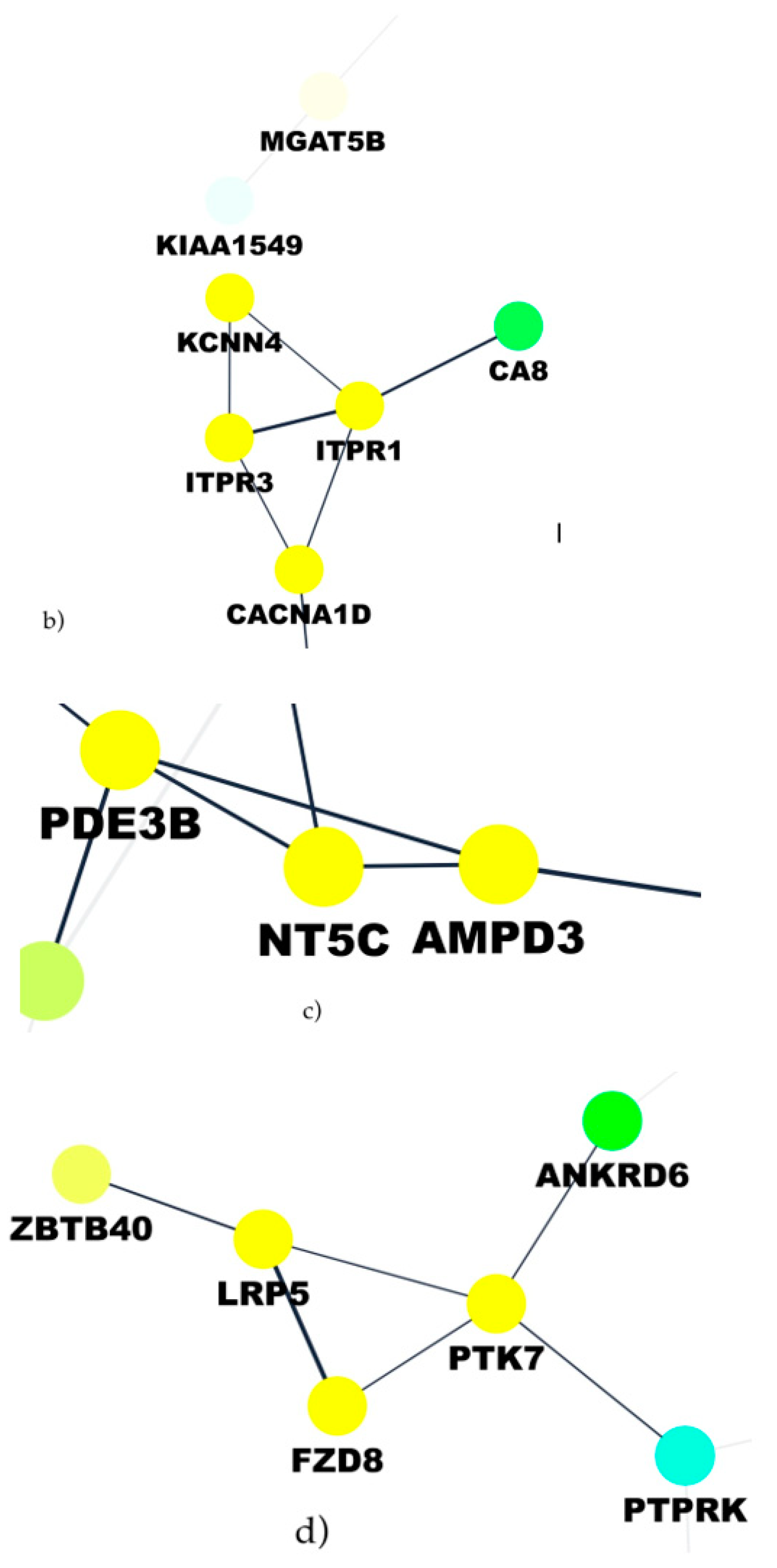
46], and the growing interest in the inhibition of the Wnt pathway in prostate cancer models confirms the validity of the therapeutic hypothesis. Another pathway deregulated in ERG-positive patients and correlated with the upregulation of the three lncRNAs is the purine pathway, with mainly NT5C and AMPD3 as correlated genes which opens up the possibility to utilize purine and pyrimidine antimetabolites [
47] for anticancer treatments of such patients. In prostate cancer, pemetrexed, which inhibits the three enzymes used in purine and pyrimidine synthesis, has been used in combination with drugs, such as docetaxel, showing only modest clinical activity [
39]. It is our opinion that a better selection of patients, stratified not only by degree of disease (Gleason value) but, also, by genomic alteration, could lead to better results in clinical trials.
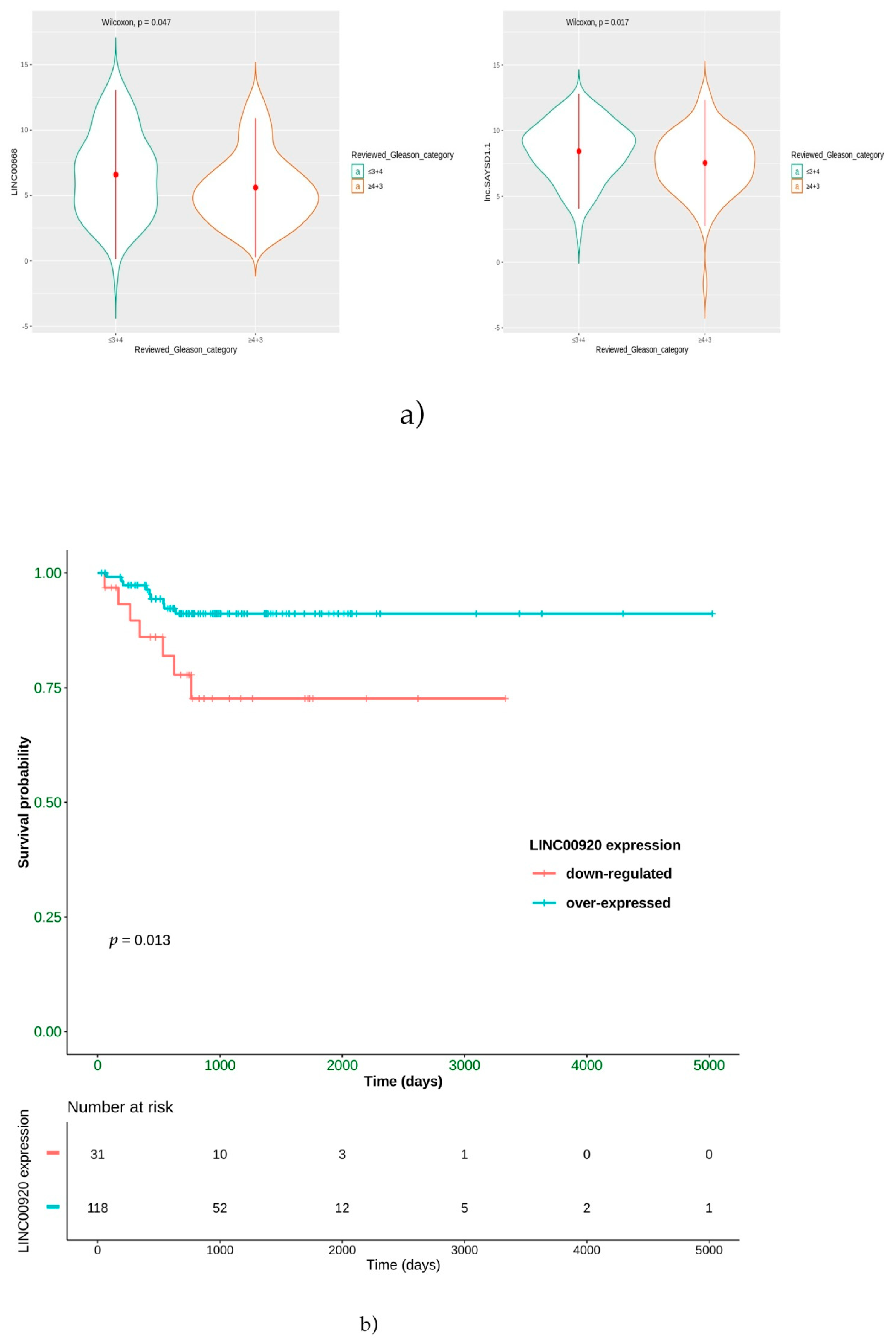
Moreover, a positive prognostic role for LINC00668 and lnc-SAYSD1-1 was highlighted. In particular, we found that the overexpression of these two lncRNAs is significantly correlated with low-grade ERG-positive patients. LINC00668 is a lncRNA known for its role in cell cycle alterations. Its overexpression has already been described as a poor prognostic factor in gastric carcinoma [
48], in breast cancer progression [
49] and in the progression of colorectal cancer (CRC) [
50]. If a role as a miR188-5p sponge is hypothesized in CRC, in gastric carcinoma, it could play a role in cyclin-dependent protein kinase inhibitors (CKIs) by means of epigenetic regulation. Both effects are contemplated in the lncRNA functions [
51,
52]. Our evidence on LINC00688 is in contrast with those reported in gastric, breast and colon cancers [
48]. Therefore, further investigation is needed to verify the role of this lncRNA in ERG-positive prostate tumors, in which it is reported that the expression of cell cycle-related genes is negatively regulated by ERG [
53]. Likewise, the lnc-SAYSD1-1 also bioinformatically allows to distinguish the ERG-positive from all the other subclasses of prostate cancer with a Gleason score ≤ 3+4. However, to date, no data have been reported on this lncRNA; therefore, we can only speculate about a possible biological role. This lncRNA maps chromosome 6 and partially overlaps the opposite strand with the DNAH8 gene (NM_001206927.2). The overexpression of this gene promotes androgen receptor activity and is associated with prostate cancer progression [
54]. Therefore, we can speculate that lncSAYSD1-1 could be a cis-NAT (natural antisense transcripts) [
55], and the overexpression of DNAH8 could be controlled primarily by the action of lnc-SAYSD1-1 through microRNA (miRNA) mechanisms. This would explain an overexpression of lnc-SAYSD1-1 in an early stage of the tumor, such as that represented by Gleason ≤ 3+4, while the absence in subjects suffering from cancer with Gleason ≥ 4+3 could be understood as a positive prognostic factor.
Another interesting result from our bioinformatics analysis is that LINC00920 is overexpressed in ERG-positive patients that showed a better survival in terms of PFS to hormonal treatment, suggesting a role for this lncRNA as a predictive factor. The Lnc00920 has been reported to positively impact pathways related to the cell cycle, cell division, apoptosis and cell movement [
56] by sequestering FOXO1, which functionally suppress the androgen receptor expression [
57]. Then, we can hypothesize that, in low-grade ERG-positive tumors, the higher the expression of Lnc00920, the better the response to hormonal treatment, because Lnc00920 may remove the FOXO1-dependent suppression of androgen receptor expression [
58].
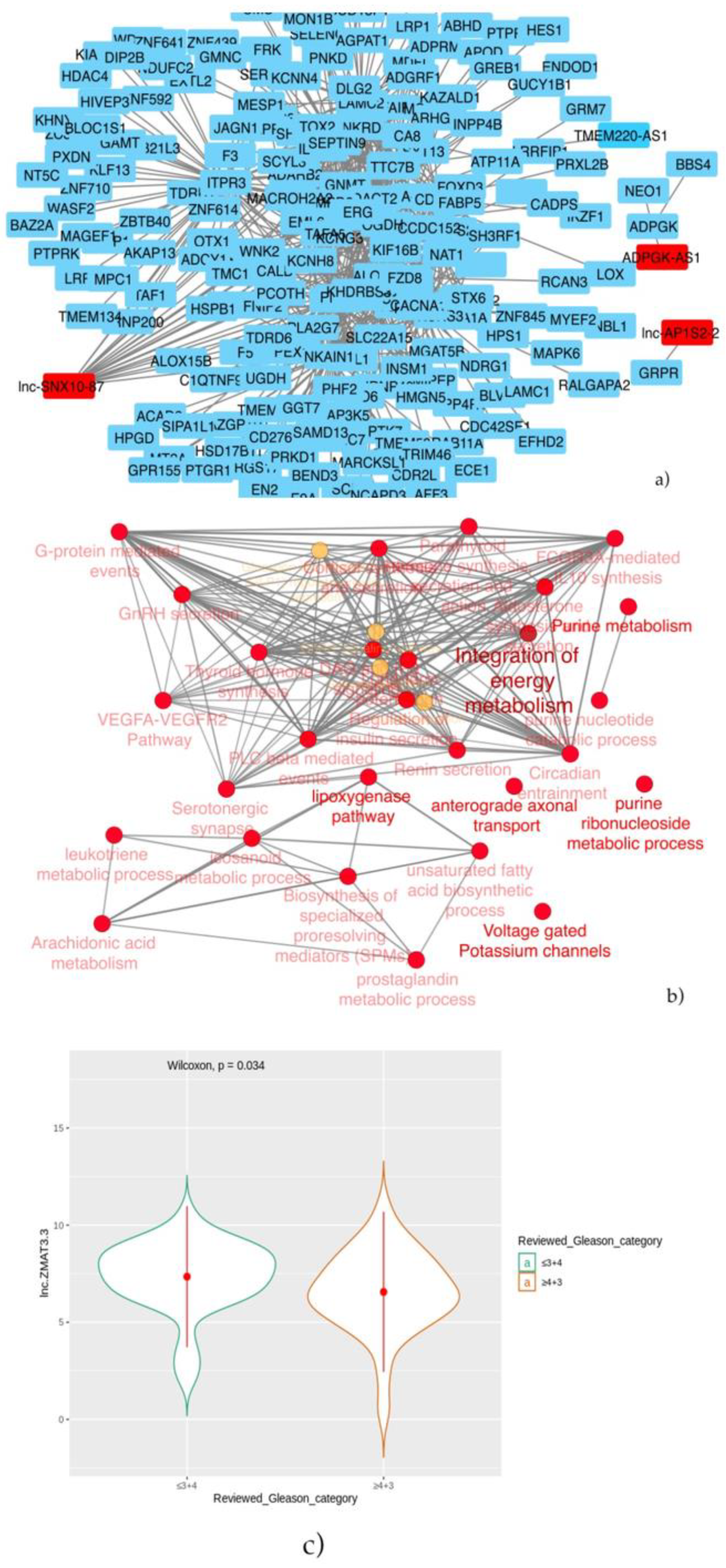
Then, patients were stratified into two groups: prostate cancer with a known genomic alteration vs. unknown, and the co-expression analysis showed that three lncRNAs: lnc-SNX10-87, lnc-AP1S2-2 and ADPGK-AS1 have significant correlation values with other coding genes. As regards the lnc-SNX10-87 alias of LOC100506289 (uncharacterized LOC100506289), it is an RNA gene affiliated with the lncRNA class and localized on chr7:26551822-26557200 [
59]. To date, nothing has been reported on the possible predictive or prognostic role of this specific lncRNA in prostate cancer. However, we found its co-expression with several protein-coding genes, among which, in particular, was the histone deacetylase 1 (HDAC1) gene, whose protein is a component of the histone deacetylase complex, a key element in the control of cell proliferation and differentiation [
59]. It has been reported that HDAC activity is highly increased in metastatic cells compared with noninvasive cancer cells [
60]. In particular, HDAC1 is upregulated in hormone refractory prostate cancer, and the overexpression of HDAC1 leads to an increase in prostate cancer cell proliferation [
61,
62]. Recently, histone deacetylase (HDAC) inhibitors have emerged as a promising new class of anticancer agents that act through a variety of mechanisms, including growth inhibition, cell cycle arrest, differentiation and apoptosis, in cancer cell lines. We suggest that these drugs may have a role in the treatment of prostate cancer patients with unknown genetic alteration. Their effectiveness has already been demonstrated in in vitro and in vivo prostate cancer studies [
52,
61,
62]. Among these MHY219 was shown to inhibit the migration of human prostate cancer cells in the study conducted by De et al. [
61]. Moreover, histone deacetylase inhibitors have been evaluated in castration-resistant prostate cancer (CRPC) or chemotherapy-resistant prostate cancer due to their effects on the expression of the androgen receptor gene. Recent clinical trials of vorinostat (ClinicalTrials.gov Identifier codes: NCT00330161 and NCT01174199), Entinostat (ClinicalTrials.gov Identifier code: NCT03829930), romidepsin and panobinostat (ClinicalTrials.gov Identifier codes: NCT00878436, NCT00493766 and NCT00663832) have provided cautious optimism towards improved outcomes using these novel therapeutic agents for CRPC patients [
63]. In addition, a phase II study of SB939 in patients with recurrent or metastatic castration-resistant prostate cancer concluded in February 2020 (ClinicalTrials.gov Identifier: NCT01075308) [
64]. Another lncRNA in the top 10 best-selected features by the stratification of prostate cancer patients with known genomic alteration vs. unknown is lnc-AP1S2-2. This lncRNA sequence is 454 nucleotides long and located on chrX:16153220-16165121 and has been annotated by four databases (NONCODE, LNCipedia, LncBook and GeneCards) and described by two different groups of researchers [
65,
66]. We found a co-expression network between lnc-AP1S2-2 and the gastrin-releasing peptide receptor (GRPR) gene, which has been extensively investigated as a molecular target in experimental anticancer therapy [
67]. An important characteristic of GRPR is that it is overexpressed in prostatic tumor cells, but only low levels of receptors were found on normal prostate tissues [
68,
69,
70,
71]. Moreover, several authors studied the correlation between GRPR overexpression in PC tissues and the tumor grade or stage, leading to different results. In particular, a significant positive correlation between the GRPR expression and Gleason score in 51 PC patients in contrast to no correlation with the patient’s age, serum PSA level, pathological stage or lymph node status was demonstrated by Nagasaki et al. [
72]. Beer et al. [
68], who analyzed the expression of GRPR both in benign and in malignant prostate samples from 530 PC patients, found a significant inverse correlation with GRPR and a higher Gleason score, PSA value and tumor size, so GRPR was more highly overexpressed in lower-grade cancer and smaller-sized tumors. A positive association between the GRP expression and relapse or advanced tumor stages was reported by Constantinides et al. [
73]. With the aim to develop a personalized management of PC patients, several ligands of GRPR, such as radiolabeled bombesin analogs (ClinicalTrials.gov Identifiers: NCT02440308 and NCT03724253) [
72] able to guide the diagnosis, as well as treatment of PC, are being tested in clinical trials. However, larger prospective clinical trials are needed to strengthen the correlation between preclinical studies in mouse tumor models and the preliminary in vivo performance in cancer patients [
71]. ADPGK-AS1, an antisense lncRNA gene mapped on chr15:72783884-72792963, was found differentially expressed in patients with known genomic alteration vs. unknown ones. Although nothing has been yet reported in the literature on the role of this lncRNA in the development and progression of prostate cancer, there is evidence of its role in the progression of pancreatic and gastric cancers. In particular, Song et al. [
72] found that ADPGK-AS1 is involved in pancreatic cancer progression through activating zinc finger E-box-binding homeobox 1(ZEB1)-mediated epithelial–mesenchymal transition. Huang et al. [
72] demonstrated that ADPGK-AS1 could promote gastric cancer progression via sponging miR-3196 and, therefore, upregulating the KDM1B gene, providing a novel prognostic biomarker and therapeutic target for GC patients. Moreover, ADPGK-AS1 was found co-expressed with the ADP-dependent glucokinase gene (ADPGK), codifying for ADPGK, which catalyzes the ADP-dependent phosphorylation of glucose to glucose-6-phosphate and may play a role in glycolysis, possibly during ischemic conditions [
72].
Finally, in order to define if these lncRNAs could have a role as prognostic or predictive biomarkers, in our analysis, we found that only lnc-ZMAT3-3 is statistically higher in patients with Gleason scores ≤ 3+4 in the “not-classified” group. Thus, it could be a positive prognosticator, meaning that its levels decrease as the disease progresses. However, nothing has been reported in the literature on the role of this lncRNA in cancer. There is no other hypothesis we can formulate, due to the chromosomal position providing little food for thought and the absence of data currently present in the literature. However, this could be a stimulus for future in vitro studies.
In conclusion, high-throughput data allowed to explore the unknown biology of tumors, increasing the burden of hypothesis-generating information. The feature selection approach used in the present study shed light on the chance to identify lncRNAs as potentially predictive biomarkers in terms of prognostication and druggable targets. Prostate cancer is a malignancy that, given its high molecular heterogeneity, could benefit from such a bioinformatic approach, targeting future studies on the selected robust lncRNAs.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}