
Perlecan in Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Perlecan is a Modular Proteoglycan
1.2. Perlecan’s Participation in Physiological Processes
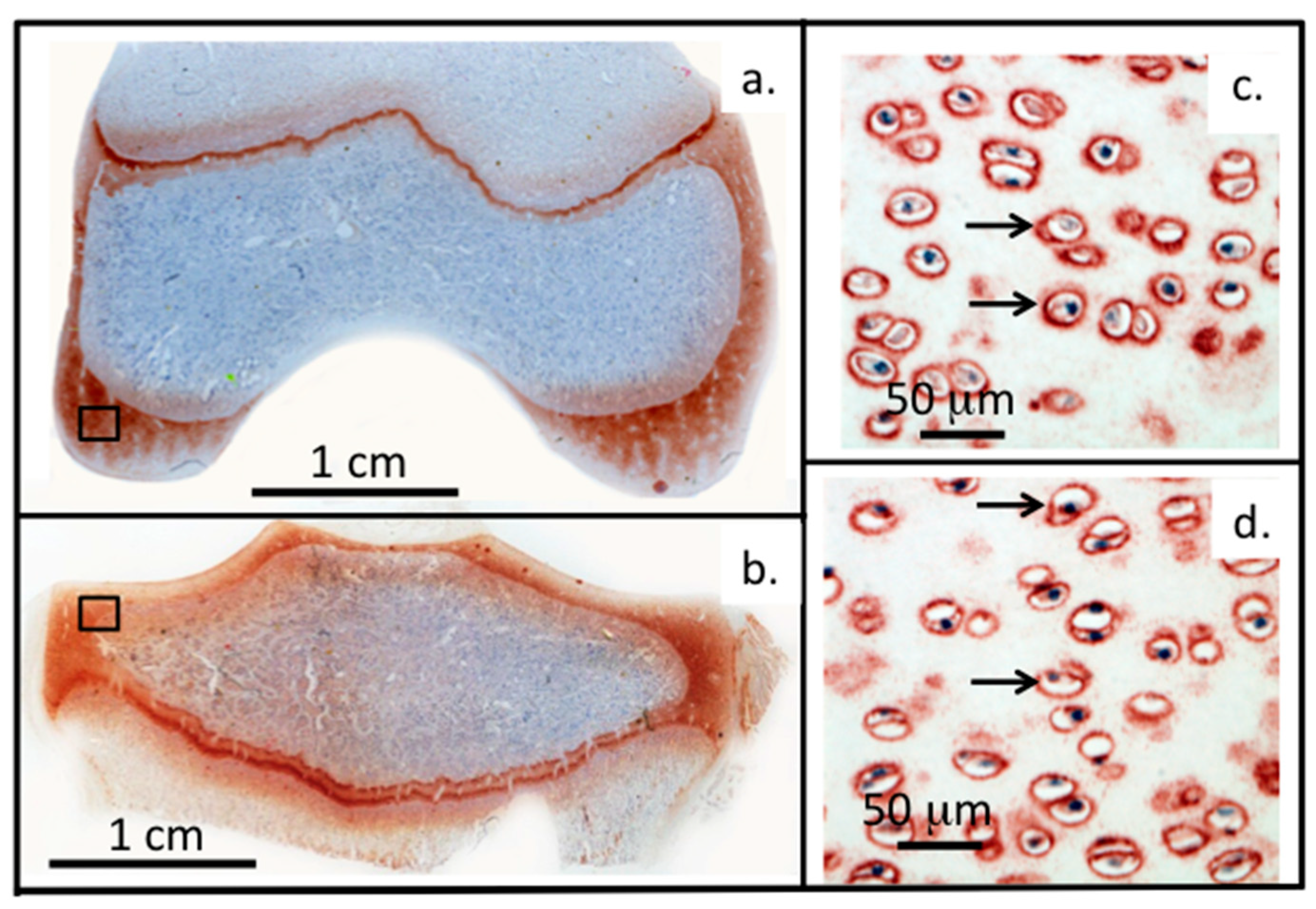
1.3. The Role of Perlecan in Chondrocyte Mechanotransduction
1.4. Perlecan Operates at Several Functional Levels in Load-Bearing Connective Tissues
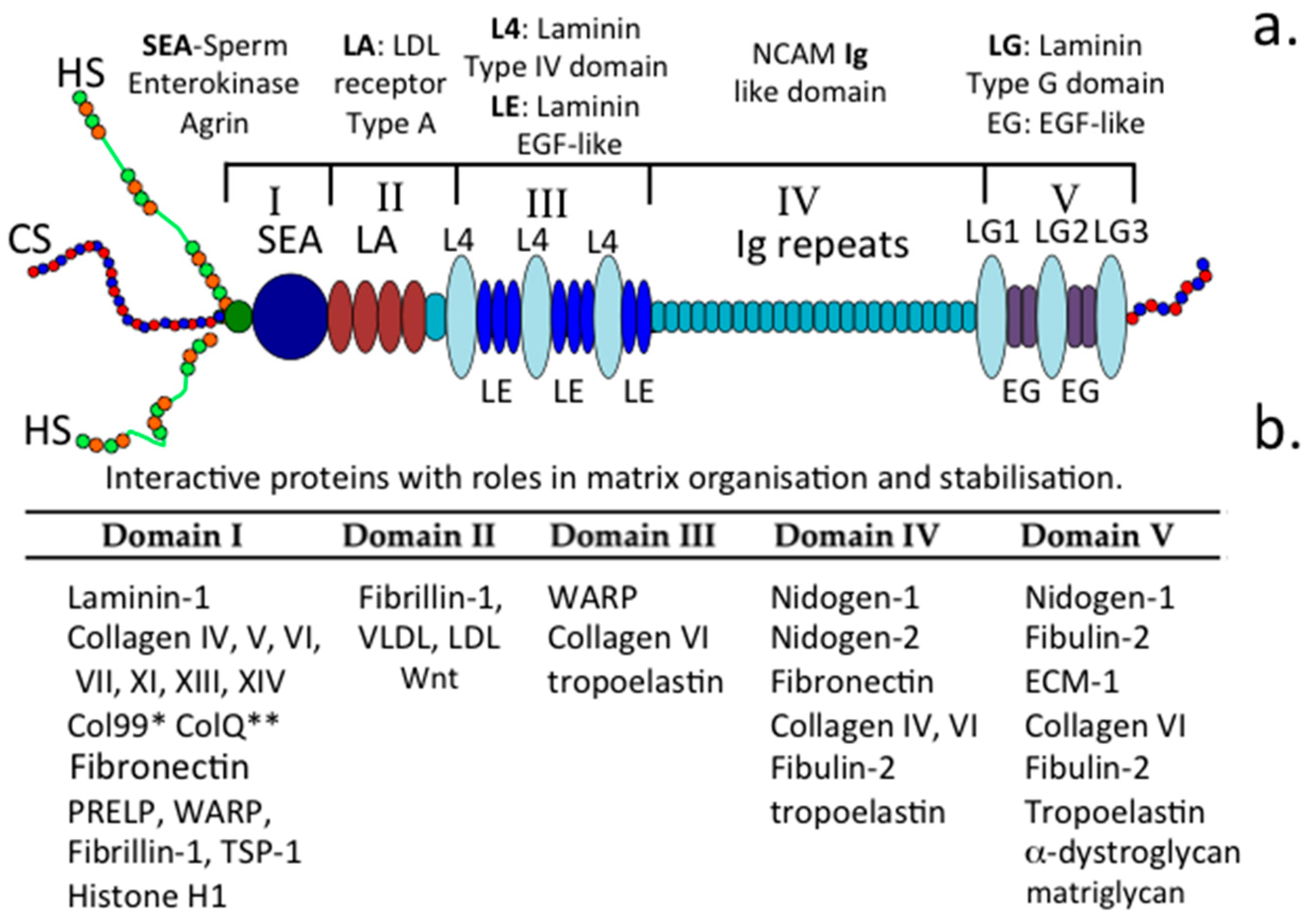
1.5. The Functional Attributes of Perlecan’s Five Domains
1.5.1. Domain-1
1.5.2. Domain II
1.5.3. Domain III
1.5.4. Domain IV
1.5.5. Domain V
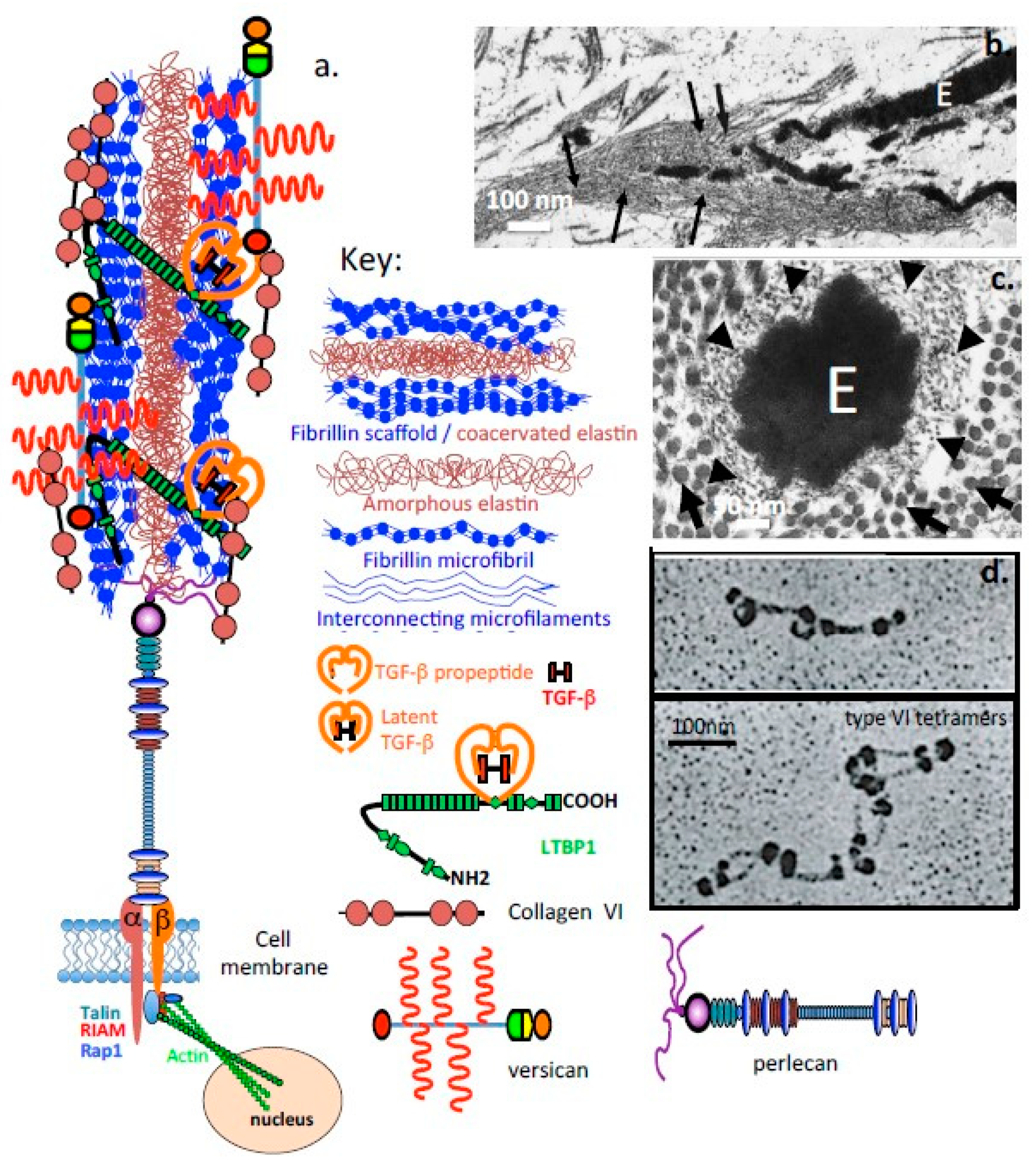
1.6. Fibrillin-1 Stabilizes Tissues and Provides Essential Functional Properties
1.7. Interactive Properties of Perlecan, Assembly of Elastic Microfibrils and Their Functional Attributes
1.8. The Pericellular Matrix, a Dynamic Cell-Matrix Interface
1.9. Elastic Networks in Load-Bearing Connective Tissues
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Melrose, J.; Hayes, A.J.; Whitelock, J.M.; Little, C.B. Perlecan, the “jack of all trades” proteoglycan of cartilaginous weight-bearing connective tissues. Bioessays 2008, 30, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Knox, S.M.; Whitelock, J.M. Perlecan: How does one molecule do so many things? Cell Mol. Life Sci. 2006, 63, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Knox, S.; Fosang, A.J.; Last, K.; Melrose, J.; Whitelock, J. Perlecan from human epithelial cells is a hybrid heparan/chondroitin/keratan sulfate proteoglycan. FEBS Lett. 2005, 579, 5019–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, M.; Lord, M.S.; Cheng, B.; Lyons, J.G.; Alkhouri, H.; Hughes, J.M.; McCarthy, S.J.; Iozzo, R.V.; Whitelock, J.M. Mast cells produce novel shorter forms of perlecan that contain functional endorepellin: A role in angiogenesis and wound healing. J. Biol. Chem. 2013, 288, 3289–3304. [Google Scholar] [CrossRef] [Green Version]
- Rnjak-Kovacina, J.; Tang, F.; Lin, X.; Whitelock, J.M.; Lord, M.S. Recombinant Domain V of Human Perlecan Is a Bioactive Vascular Proteoglycan. Biotechnol. J. 2017, 12, 196. [Google Scholar] [CrossRef]
- Siegel, G.; Malmsten, M.; Ermilov, E. Anionic biopolyelectrolytes of the syndecan/perlecan superfamily: Physicochemical properties and medical significance. Adv. Colloid Interface Sci. 2014, 205, 275–318. [Google Scholar] [CrossRef] [PubMed]
- Gradilla, A.C.; Sanchez-Hernandez, D.; Brunt, L.; Scholpp, S. From top to bottom: Cell polarity in Hedgehog and Wnt trafficking. BMC Biol. 2018, 16, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Nagy, T.L.; Weiner, O.D. Joining forces: Crosstalk between biochemical signalling and physical forces orchestrates cellular polarity and dynamics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170145. [Google Scholar] [CrossRef] [Green Version]
- Thakore, P.; Earley, S. Transient Receptor Potential Channels and Endothelial Cell Calcium Signaling. Compr. Physiol. 2019, 9, 1249–1277. [Google Scholar]
- Ottolini, M.; Hong, K.; Sonkusare, S.K. Calcium signals that determine vascular resistance. Wiley Interdiscip. Rev. Syst. Biol. Med. 2019, 11, e1448. [Google Scholar] [CrossRef]
- Thompson, W.; Modla, S.; Grindel, B.J.; Czymmek KJKirn-Safran, C.B.; Wang, L.; Duncan, R.L.; Farach-Carson, M.C. Perlecan/Hspg2 deficiency alters the pericellular space of the lacunocanalicular system surrounding osteocytic processes in cortical bone. J. Bone Miner Res. 2011, 26, 618–629. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Lai, X.; Price, C.; Thompson, W.R.; Li, W.; Quabili, T.R.; Tseng, W.J.; Liu, X.S.; Zhang, H.; Pan, J.; et al. Perlecan-containing pericellular matrix regulates solute transport and mechanosensing within the osteocyte lacunar-canalicular system. J. Bone Miner Res. 2014, 29, 878–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L. Solute Transport in the Bone Lacunar-Canalicular System (LCS). Curr. Osteoporos. Rep. 2018, 16, 32–41. [Google Scholar] [CrossRef]
- Mow, V.; Bachrach, N.M.; Setton, L.A.; Guilak, F. Stress, Strain, Pressure and Flow Fields in Articular Cartilage and Chondrocytes. In Cell Mechanics and Cellular Engineering; Mow, V.C., Tran-Son-Tay, R., Guilak, F., Hochmuth, R.M., Eds.; Springer LLC: New York, NY, USA, 1994; pp. 345–379. [Google Scholar]
- Haider, M.; Schugart, R.C.; Setton, L.A.; Guilak, F. A mechano-chemical model for the passive swelling response of an isolated chondron under osmotic loading. Biomech. Model. Mechanobiol. 2006, 5, 160–171. [Google Scholar] [CrossRef]
- Maroudas, A. Physicochemical Properties of Articular Cartilage. In Adult Articular Cartilage; Mar, F., Ed.; Pitman Medical: Bath, UK, 1979; pp. 215–290. [Google Scholar]
- Urban, J.; Hall, A.C.; Gehl, K.A. Regulation of matrix synthesis rates by the ionic and osmotic environment of articular chondrocytes. J. Cell Physiol. 1993, 154, 262–270. [Google Scholar] [CrossRef]
- Urban, J. The chondrocyte: A cell under pressure. Br. J. Rheumatol. 1994, 33, 901–908. [Google Scholar] [CrossRef]
- Jeremiasse, B.; Matta, C.; Fellows, C.R.; Boocock, D.J.; Smith, J.R.; Liddell, S.; Lafeber, F.; van Spil, W.E.; Mobasheri, A. Alterations in the chondrocyte surfaceome in response to pro-inflammatory cytokines. BMC Mol. Cell Biol. 2020, 21, 47. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.; May, H.; Mobasheri, A.; Barrett-Jolley, R. Chondrocyte channel transcriptomics: Do microarray data fit with expression and functional data? Channels 2013, 7, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleckar, M.; Martin-Vasallo, P.; Giles, W.R.; Mobasheri, A. Physiological Effects of the Electrogenic Current Generated by the Na+/K+ Pump in Mammalian Articular Chondrocytes. Bioelectricity 2020, 2, 2020. [Google Scholar] [CrossRef]
- Mobasheri, A.; Mobasheri, R.; Francis, M.J.; Trujillo, E.; Alvarez de la Rosa, D.; Martín-Vasallo, P. Ion transport in chondrocytes: Membrane transporters involved in intracellular ion homeostasis and the regulation of cell volume, free [Ca2+] and pH. Histol. Histopathol. 1998, 13, 893–910. [Google Scholar] [PubMed]
- Mobasheri, A.; Matta, C.; Uzieliene, I.; Budd, E.; Martín-Vasallo, P.; Bernotiene, E. The chondrocyte channelome: A narrative review. Jt. Bone Spine 2019, 86, 29–35. [Google Scholar] [CrossRef]
- Lee, W.; Leddy, H.A.; Chen, Y.; Lee, S.H.; Zelenski, N.A.; McNulty, A.L.; Wu, J.; Beicker, K.N.; Coles, J.; Zauscher, S.; et al. Synergy between Piezo1 and Piezo2 channels confers high-strain mechanosensitivity to articular cartilage. Proc. Natl. Acad. Sci. USA 2014, 111, E5114–E5122. [Google Scholar] [CrossRef] [Green Version]
- Phan, M.; Leddy, H.A.; Votta, B.J.; Kumar, S.; Levy, D.S.; Lipshutz, D.B.; Lee, S.H.; Liedtke, W.; Guilak, F. Functional characterization of TRPV4 as an osmotically sensitive ion channel in porcine articular chondrocytes. Arthritis Rheumatol. 2009, 60, 3028–3037. [Google Scholar] [CrossRef] [Green Version]
- Mobasheri, A. Regulation of Na+, K+-ATPase density by the extracellular ionic and osmotic environment in bovine articular chondrocytes. Physiol. Res. 1999, 45, 509–512. [Google Scholar]
- Mobasheri, A.; Avila, J.; Co´zar-Castellano, I.; Brownleader, M.D.; Trevan, M.; Francis, M.J.; Lamb, J.F.; Martín-Vasallo, P. Na+, K+-ATPase isozyme diversity; comparative biochemistry and physiological implications of novel functional interactions. Biosci. Rep. 2000, 20, 51–91. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Errington, R.J.; Golding, S.; Hall, A.C.; Urban, J.P. Characterization of the Na+, K(+)-ATPase in isolated bovine articular chondrocytes; molecular evidence for multiple alpha and beta isoforms. Cell Biol. Int. 1997, 21, 212–301. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Hall, A.C.; Urban, J.P.; France, S.J.; Smith, A.L. Immunologic and autoradiographic localisation of the Na+, K(+)-ATPase in articular cartilage: Upregulation in response to changes in extracellular Na+ concentration. Int. J. Biochem. Cell Biol. 1997, 29, 649–657. [Google Scholar] [CrossRef]
- Trujillo, E.; de la Rosa, D.A.; Mobasheri, A.; Avila, J.; González, T.; Martín-Vasallo, P. Sodium transport systems in human chondrocytes. I. Morphological and functional expression of the Na+,K(+)- ATPase alpha and beta subunit isoforms in healthy and arthritic chondrocytes. Histol. Histopathol. 1999, 14, 1011–1022. [Google Scholar]
- Erickson, G.; Northrup, D.; Guilak, F. Hypo-osmotic stress induces calcium-dependent actin reorganization in articular chondrocytes. Osteoarth. Cartil. 2003, 11, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Zelenski, N.; Leddy, H.A.; Sanchez-Adams, J.; Zhang, J.; Bonaldo, P.; Liedtke, W.; Guilak, F. Type VI Collagen Regulates Pericellular Matrix Properties, Chondrocyte Swelling, and Mechanotransduction in Mouse Articular Cartilage. Arthritis Rheumatol. 2015, 67, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- Gilchrist, C.; Leddy, H.A.; Kaye, L.; Case, N.D.; Rothenberg, K.E.; Little, D.; Liedtke, W.; Hoffman, B.D.; Guilak, F. TRPV4-mediated calcium signaling in mesenchymal stem cells regulates aligned collagen matrix formation and vinculin tension. Proc. Natl. Acad. Sci. USA 2019, 116, 1992–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, T.; Saklatvala, J. Basic fibroblast growth factor: An extracellular mechanotransducer in articular cartilage? Biochem. Soc. Trans. 2006, 34, 456–457. [Google Scholar] [CrossRef]
- Vincent, T.; McLean, C.J.; Full, L.E.; Peston, D.; Saklatvala, J. FGF-2 is bound to perlecan in the pericellular matrix of articular cartilage, where it acts as a chondrocyte mechanotransducer. Osteoarthr. Cartil. 2007, 15, 752–763. [Google Scholar] [CrossRef] [Green Version]
- Danalache, M.; Kleinert, R.; Schneider, J.; Erler, A.L.; Schwitalle, M.; Riester, R.; Traub, F.; Hofmann, U.K. Changes in stiffness and biochemical composition of the pericellular matrix as a function of spatial chondrocyte organisation in osteoarthritic cartilage. Osteoarthr. Cartil. 2019, 27, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Wiberg, C.; Hedbom, E.; Khairullina, A.; Lamande, S.R.; Oldberg, A.; Timpl, R.; Morgelin, M.; Heinegard, D. Biglycan and decorin bind close to the n-terminal region of the collagen VI triple helix. J. Biol. Chem. 2001, 276, 18947–18952. [Google Scholar] [CrossRef] [Green Version]
- Hayes, A.J.; Shu, C.C.; Lord, M.S.; Little, C.B.; Whitelock, J.M.; Melrose, J. Pericellular colocalisation and interactive properties of type VI collagen and perlecan in the intervertebral disc. Eur. Cell Mater. 2016, 32, 40–57. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Isaacs, M.D.; Smith, S.M.; Hughes, C.E.; Little, C.B.; Caterson, B.; Hayes, A.J. Chondroitin sulphate and heparan sulphate sulphation motifs and their proteoglycans are involved in articular cartilage formation during human foetal knee joint development. Histochem. Cell Biol. 2012, 138, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Roughley, P.; Knox, S.; Smith, S.; Lord, M.; Whitelock, J. The structure, location, and function of perlecan, a prominent pericellular proteoglycan of fetal, postnatal, and mature hyaline cartilages. J. Biol. Chem. 2006, 281, 36905–36914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melrose, J.; Shu, C.; Whitelock, J.M.; Lord, M.S. The cartilage extracellular matrix as a transient developmental scaffold for growth plate maturation. Matrix Biol. 2016, 52–54, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Shu, C.; Melrose, J. Comparative immunolocalisation of perlecan with collagen II and aggrecan in human foetal, newborn and adult ovine joint tissues demonstrates perlecan as an early developmental chondrogenic marker. Histochem. Cell Biol. 2010, 134, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, R.E.; Defrate, L.E.; Guilak, F. A biomechanical role for perlecan in the pericellular matrix of articular cartilage. Matrix Biol. 2012, 31, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Chery, D.; Han, B.; Zhou, Y.; Wang, C.; Adams, S.M.; Chandrasekaran, P.; Kwok, B.; Heo, S.J.; Enomoto-Iwamoto, M.; Lu, X.L.; et al. Decorin Regulates Cartilage Pericellular Matrix Micromechanobiology. Matrix Biol. 2021, 96, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Li, Q.; Wang, C.; Patel, P.; Adams, S.M.; Doyran, B.; Nia, H.T.; Oftadeh, R.; Zhou, S.; Li, C.Y.; et al. Decorin Regulates the Aggrecan Network Integrity and Biomechanical Functions of Cartilage Extracellular Matrix. ACS Nano 2019, 13, 11320–11333. [Google Scholar] [CrossRef] [PubMed]
- Bilton, M.; Thambyah, A.; Clarke, R.J. How changes in interconnectivity affect the bulk properties of articular cartilage: A fibre network study. Biomech. Model. Mechanobiol. 2018, 17, 1297–1315. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Ojanen, S.; Mohammadi, A.; Finnilä, M.A.; Joukainen, A.; Kröger, H.; Saarakkala, S.; Korhonen, R.K.; Tanska, P. Elastic, Viscoelastic and Fibril-Reinforced Poroelastic Material Properties of Healthy and Osteoarthritic Human Tibial Cartilage. Ann. Biomed. Eng. 2019, 47, 953–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottardi, R.; Hansen, U.; Raiteri, R.; Loparic, M.; Düggelin, M.; Mathys, D.; Friederich, N.F.; Bruckner, P.; Stolz, M. Supramolecular Organization of Collagen Fibrils in Healthy and Osteoarthritic Human Knee and Hip Joint Cartilage. PLoS ONE 2016, 11, e0163552. [Google Scholar] [CrossRef] [PubMed]
- Nickien, M.; Thambyah, A.; Broom, N.D. How a decreased fibrillar interconnectivity influences stiffness and swelling properties during early cartilage degeneration. J. Mech. Behav. Biomed. Mater. 2017, 75, 390–398. [Google Scholar] [CrossRef]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Perlecan displays variable spatial and temporal immunolocalisation patterns in the articular and growth plate cartilages of the ovine stifle joint. Histochem. Cell Biol. 2005, 123, 561–571. [Google Scholar] [CrossRef]
- Hayes, A.J.; Gibson, M.A.; Shu, C.; Melrose, J. Confocal microscopy demonstrates association of LTBP-2 in fibrillin-1 microfibrils and colocalisation with perlecan in the disc cell pericellular matrix. Tissue Cell 2014, 46, 185–197. [Google Scholar] [CrossRef]
- Hayes, A.J.; Smith, S.M.; Gibson, M.A.; Melrose, J. Comparative immunolocalization of the elastin fiber-associated proteins fibrillin-1, LTBP-2, and MAGP-1 with components of the collagenous and proteoglycan matrix of the fetal human intervertebral disc. Spine 2011, 36, E1365–E1372. [Google Scholar] [CrossRef]
- Hayes, A.J.; Smith, S.M.; Melrose, J. Comparative immunolocalisation of fibrillin-1 and perlecan in the human foetal, and HS-deficient hspg2 exon 3 null mutant mouse intervertebral disc. Histochem. Cell Biol. 2013, 139, 1–11. [Google Scholar] [CrossRef]
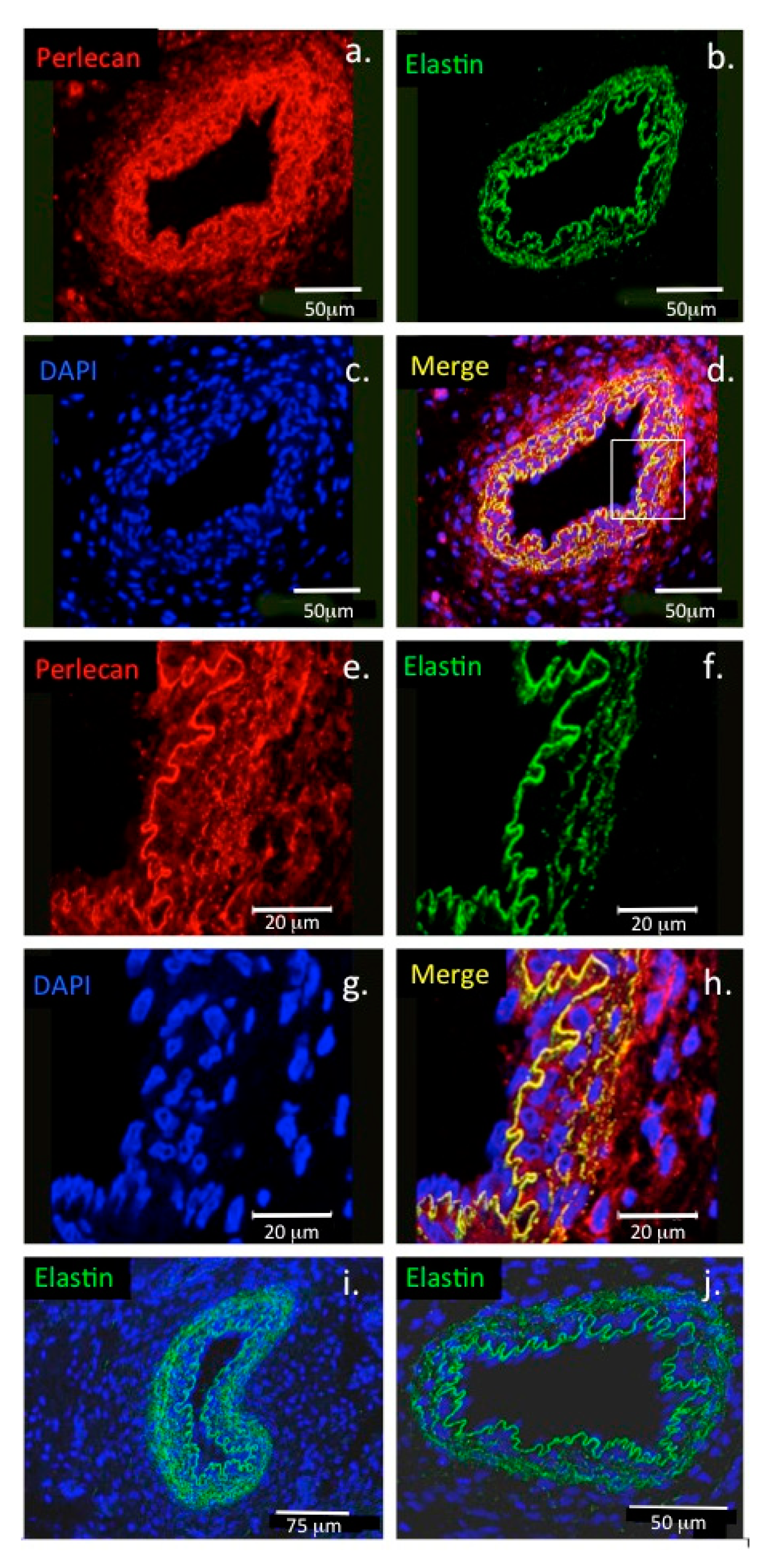
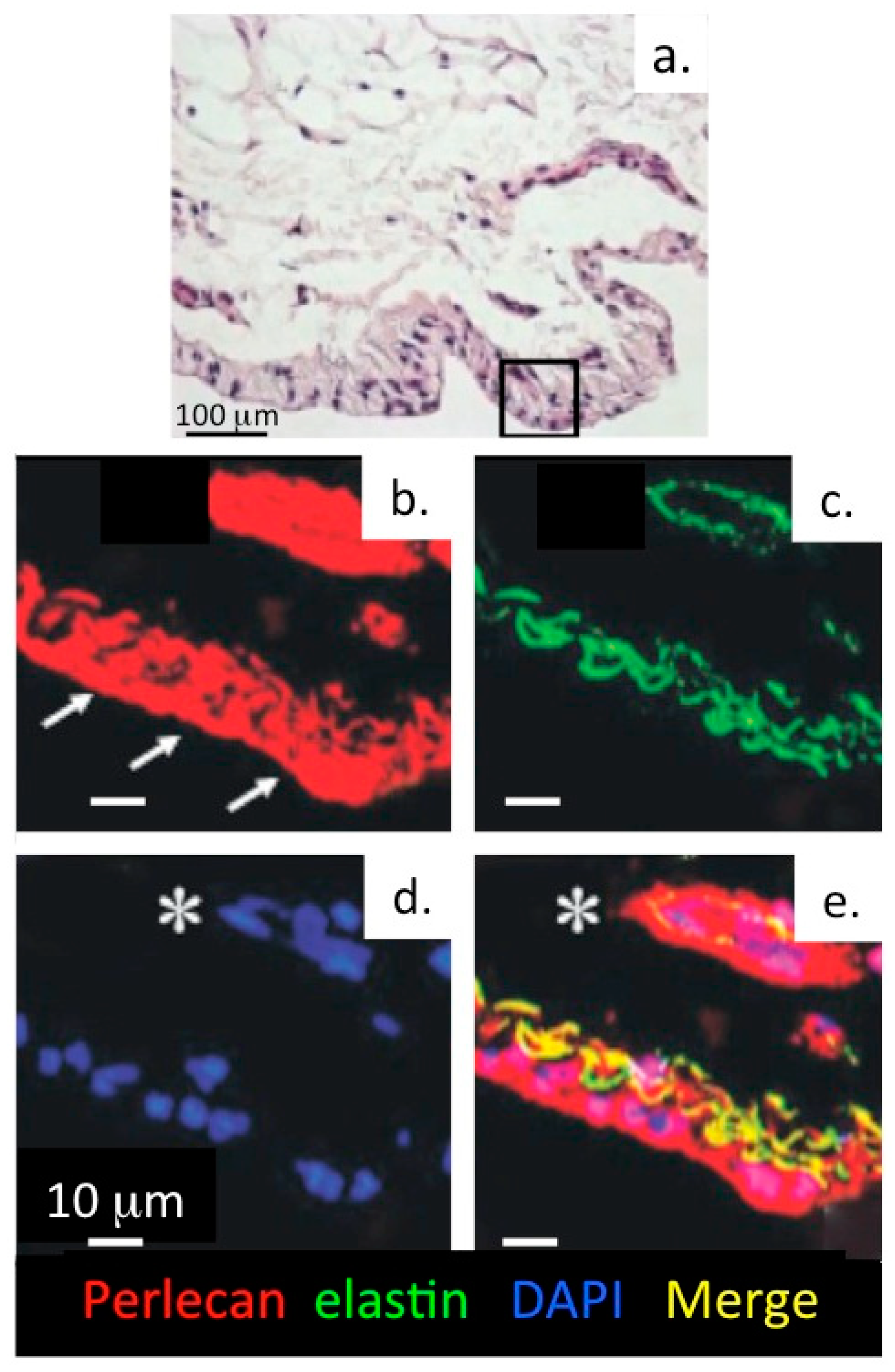
- Hayes, A.J.; Lord, M.S.; Smith, S.M.; Smith, M.M.; Whitelock, J.M.; Weiss, A.S.; Melrose, J. Colocalization in vivo and association in vitro of perlecan and elastin. Histochem. Cell Biol. 2011, 136, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Montes, G. Structural biology of the fibres of the collagenous and elastic systems. Cell Biol. Int. 1996, 20, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Engvall, E.; Hessle, H.; Klier, G. Molecular assembly, secretion, and matrix deposition of type VI collagen. J. Cell Biol. 1986, 102, 703–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fearing, B.; Hernandez, P.A.; Setton, L.A.; Chahine, N.O. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef] [PubMed]
- Hickey, D.; Hukins, D.W. Relation between the structure of the annulus fibrosus and the function and failure of the intervertebral disc. Spine 1980, 5, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Molladavoodi, S.; McMorran, J.; Gregory, D. Mechanobiology of annulus fibrosus and nucleus pulposus cells in intervertebral discs. Cell Tissue Res. 2020, 379, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Elliott, D.M.; Costi, J.J. Structure and mechanical function of the inter-lamellar matrix of the annulus fibrosus in the disc. J. Orthop. Res. 2016, 34, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Smith, S.M.; Appleyard, R.C.; Little, C.B. Aggrecan, versican and type VI collagen are components of annular translamellar crossbridges in the intervertebral disc. Eur. Spine J. 2008, 17, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Schollum, M.L.; Wade, K.R.; Broom, N.D.; Urban, J.P. ISSLS Prize Winner: A Detailed Examination of the Elastic Network Leads to a New Understanding of Annulus Fibrosus Organization. Spine 2015, 40, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
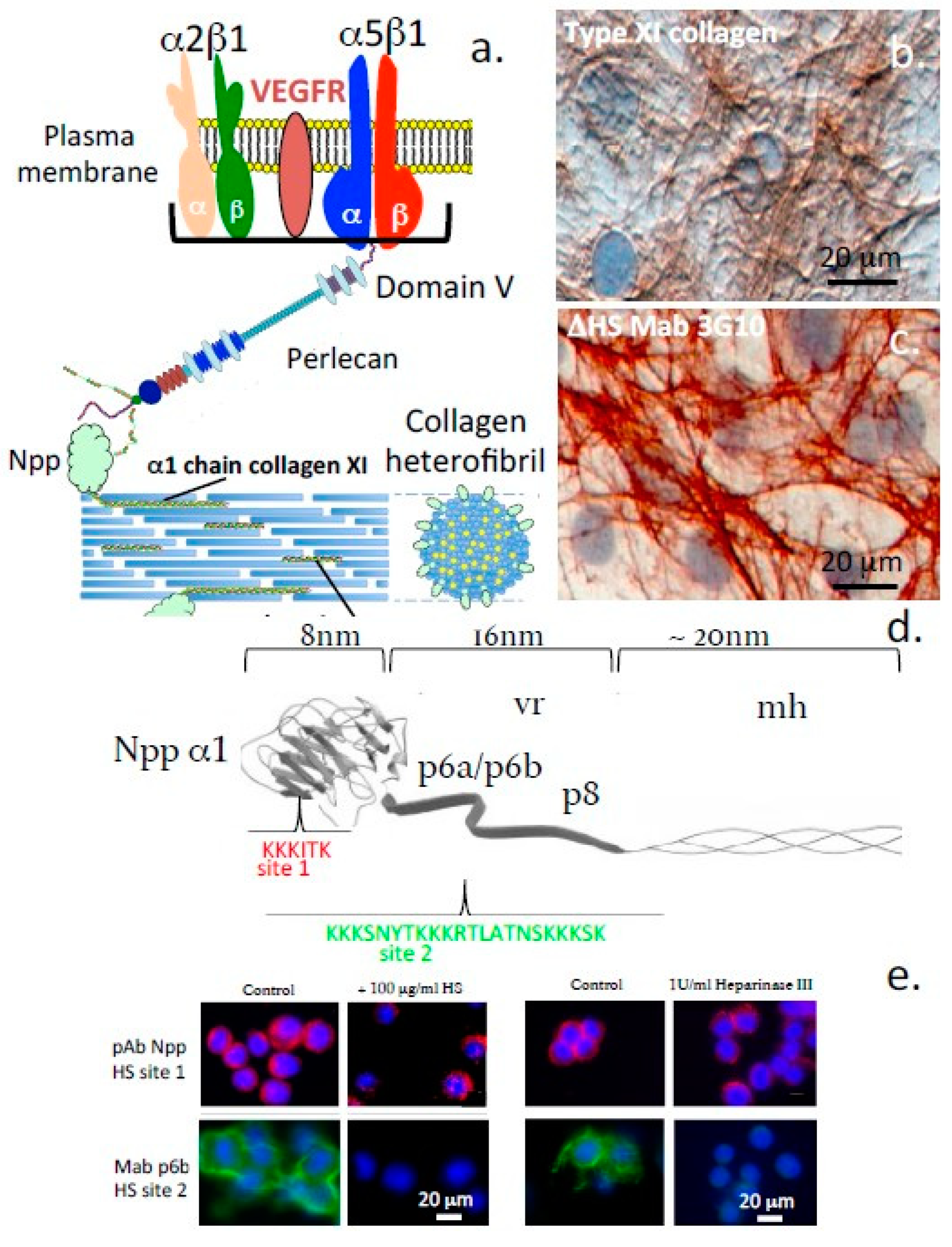
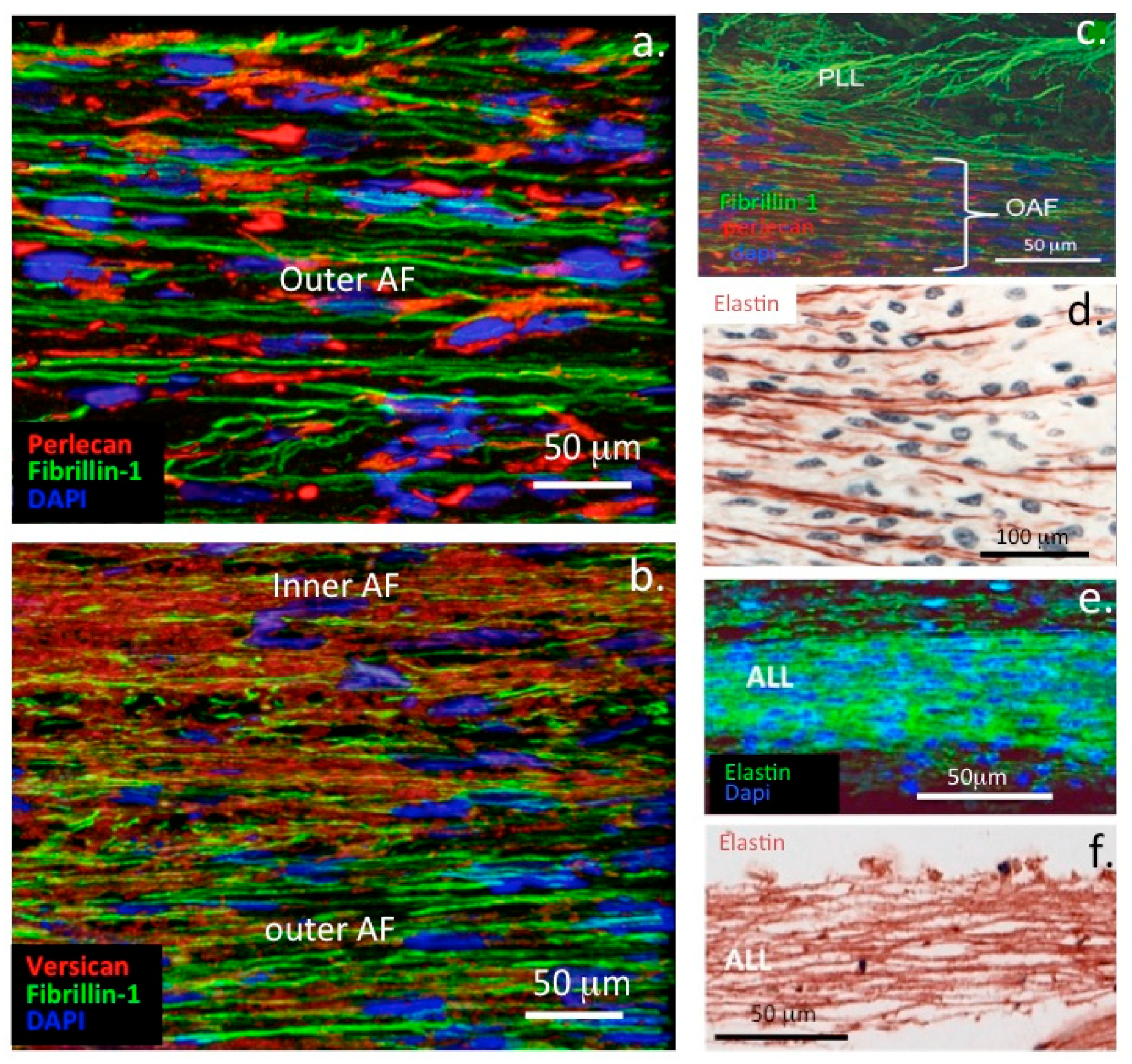
- Smith, S.; Melrose, J. Type XI collagen-perlecan-HS interactions stabilise the pericellular matrix of annulus fibrosus cells and chondrocytes providing matrix stabilisation and homeostasis. J. Mol. Histol. 2019, 50, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Warner, L.; Brown, R.J.; Yingst, S.M.; Oxford, J.T. Isoform-specific heparan sulfate binding within the amino-terminal noncollagenous domain of collagen alpha1(XI). J. Biol. Chem. 2006, 281, 39507–39516. [Google Scholar] [CrossRef] [Green Version]
- Ida-Yonemochi, H.; Ahsan, M.S.; Saku, T. Differential expression profiles between α-dystroglycan and integrin β1 in ameloblastoma: Two possible perlecan signalling pathways for cellular growth and differentiation. Histopathology 2011, 58, 234–245. [Google Scholar] [CrossRef]
- Kamimura, K.; Ueno, K.; Nakaawa, J.; Hamada, R.; Saitoe, M.; Maeda, N. Perlecan regulates bidirectional Wnt signaling at the Drosophila neuromuscular junction. J. Cell Biol. 2013, 200, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Singhal, N.; Martin, P.T. Role of extracellular matrix proteins and their receptors in the development of the vertebrate neuromuscular junction. Dev. Neurobiol. 2011, 71, 982–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Adams, J.; Wilusz, R.E.; Guilak, F. Atomic force microscopy reveals regional variations in the micromechanical properties of the pericellular and extracellular matrices of the meniscus. J. Orthop. Res. 2013, 31, 1218–1225. [Google Scholar] [CrossRef] [Green Version]
- Wilusz, R.; Guilak, F. High resistance of the mechanical properties of the chondrocyte pericellular matrix to proteoglycan digestion by chondroitinase, aggrecanase, or hyaluronidase. J. Mech. Behav. Biomed. Mater. 2014, 38, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, E.; Mörgelin, M.; Sasaki, T.; Timpl, R.; Heinegård, D.; Aspberg, A. The leucine-rich repeat protein PRELP binds perlecan and collagens and may function as a basement membrane anchor. J. Biol. Chem. 2002, 277, 15061–15068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.; Zamurs, L.; Brachvogel, B.; Schlötzer-Schrehardt, U.; Hansen, U.; Lamandé, S.R.; Rowley, L.; Fitzgerald, J.; Bateman, J.F. Mice lacking the extracellular matrix protein WARP develop normally but have compromised peripheral nerve structure and function. J. Biol. Chem. 2009, 284, 12020–12030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farach-Carson, M.; Brown, A.J.; Lynam, M.; Safran, J.B.; Carson, D.D. A novel peptide sequence in perlecan domain IV supports cell adhesion, spreading and FAK activation. Matrix Biol. 2008, 27, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murdoch, A.; Dodge, G.R.; Cohen, I.; Tuan, R.S.; Iozzo, R.V. Primary structure of the human heparan sulfate proteoglycan from basement membrane (HSPG2/perlecan). A chimeric molecule with multiple domains homologous to the low density lipoprotein receptor, laminin, neural cell adhesion molecules, and epidermal growth factor. J. Biol. Chem. 1992, 267, 8544–8557. [Google Scholar]
- Noonan, D.; Fulle, A.; Valente, P.; Cai, S.; Horigan, E.; Sasaki, M.; Yamada, Y.; Hassell, J.R. The complete sequence of perlecan, a basement membrane heparan sulfate proteoglycan, reveals extensive similarity with laminin A chain, low density lipoprotein-receptor, and the neural cell adhesion molecule. J. Biol. Chem. 1991, 266, 22939–22947. [Google Scholar] [CrossRef]
- Grindel, B.; Li, Q.; Arnold, R.; Petros, J.; Zayzafoon, M.; Muldoon, M.; Stave, J.; Chung, L.W.; Farach-Carson, M.C. Perlecan/HSPG2 and matrilysin/MMP-7 as indices of tissue invasion: Tissue localization and circulating perlecan fragments in a cohort of 288 radical prostatectomy patients. Oncotarget 2016, 7, 10433–40447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, L.; Tellman, T.V.; Farach-Carson, M.C. Flipping the Molecular Switch: Influence of Perlecan and Its Modifiers in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 133–146. [Google Scholar] [PubMed]
- Bix, G. Perlecan domain V therapy for stroke: A beacon of hope? ACS Chem. Neurosci. 2013, 4, 370–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trout, A.; Kahle, M.P.; Roberts, J.M.; Marcelo, A.; de Hoog, L.; Boychuk, J.A.; Grupke, S.L.; Berretta, A.; Gowing, E.K.; Boychuk, C.R.; et al. Perlecan Domain-V Enhances Neurogenic Brain Repair After Stroke in Mice. Transl. Stroke Res. 2020, 12, 72–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapanadechopone, P.; Hassell, J.R.; Rigatti, B.; Couchman, J.R. Localization of glycosaminoglycan substitution sites on domain V of mouse perlecan. Biochem. Biophys. Res. Commun. 1999, 265, 680–690. [Google Scholar] [CrossRef]
- Kielty, C. Fell-Muir Lecture: Fibrillin microfibrils: Structural tensometers of elastic tissues? Int. J. Exp. Pathol. 2017, 98, 172–190. [Google Scholar] [CrossRef] [Green Version]
- Godwin, A.; Singh, M.; Lockhart-Cairns, M.P.; Alanazi, Y.F.; Cain, S.A.; Baldock, C. The role of fibrillin and microfibril binding proteins in elastin and elastic fibre assembly. Matrix Biol. 2019, 84, 17–30. [Google Scholar] [CrossRef]
- Kozel, B.; Mecham, R.P. Elastic fiber ultrastructure and assembly. Matrix Biol. 2019, 84, 31–40. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. 3D Distribution of Perlecan within Intervertebral Disc Chondrons Suggests Novel Regulatory Roles for this Multifunctional Modular Heparan Sulphate Proteoglycan. Eur. Cells Mater. 2020, 41, 73–89. [Google Scholar] [CrossRef]
- Yadin, D.; Robertson, I.B.; McNaught-Davis, J.; McNaught-Davis, J.; Evans, P.; Stoddart, D.; Handford, P.A.; Jensen, S.A.; Redfield, C. Structure of the fibrillin-1 N-terminal domains suggests that heparan sulfate regulates the early stages of microfibril assembly. Structure 2013, 21, 1743–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.; Yanagisawa, H. Recent updates on the molecular network of elastic fiber formation. Essays Biochem. 2019, 63, 365–376. [Google Scholar]
- Thomson, J.; Singh, M.; Eckersley, A.; Cain, S.A.; Sherratt, M.J.; Baldock, C. Fibrillin microfibrils and elastic fibre proteins: Functional interactions and extracellular regulation of growth factors. Semin. Cell Dev. Biol. 2019, 89, 109–117. [Google Scholar] [CrossRef]
- Cao, L.; Guilak, F.; Setton, L.A. Three-dimensional morphology of the pericellular matrix of intervertebral disc cells in the rat. J. Anat. 2007, 211, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Guilak, F.; Setton, L.A. Pericellular Matrix Mechanics in the Anulus Fibrosus Predicted by a Three-Dimensional Finite Element Model and In Situ Morphology. Cell Mol. Bioeng. 2009, 2, 306–319. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Guilak, F.; Setton, L.A. Three-dimensional finite element modeling of pericellular matrix and cell mechanics in the nucleus pulposus of the intervertebral disk based on in situ morphology. Biomech. Model. Mechanobiol. 2011, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Isogai, Z.; Aspberg, A.; Keene, D.R.; Ono, R.N.; Reinhardt, D.P.; Sakai, L.Y. Versican interacts with fibrillin-1 and links extracellular microfibrils to other connective tissue networks. J. Biol. Chem. 2002, 277, 4565–4572. [Google Scholar] [CrossRef] [Green Version]
- Kinsey, R.; Williamson, M.R.; Chaudhry, S.; Mellody, K.T.; McGovern, A.; Takahashi, S.; Shuttleworth, C.A.; Kielty, C.M. Fibrillin-1 microfibril deposition is dependent on fibronectin assembly. J. Cell Sci. 2008, 121, 2696–2704. [Google Scholar] [CrossRef] [Green Version]
- Sabatier, L.; Djokic, J.; Fagotto-Kaufmann, C.; Chen, M.; Annis, D.S.; Mosher, D.F.; Reinhardt, D.P. Complex contributions of fibronectin to initiation and maturation of microfibrils. Biochem. J. 2013, 456, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massam-Wu, T.; Chiu, M.; Choudhury, R.; Chaudhry, S.S.; Baldwin, A.K.; McGovern, A.; Baldock, C.; Shuttleworth, C.A.; Kielty, C.M. Assembly of fibrillin microfibrils governs extracellular deposition of latent TGF beta. J. Cell Sci. 2010, 123, 3006–3018. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, F.; Dietz, H.C. Fibrillin-rich microfibrils: Structural determinants of morphogenetic and homeostatic events. J. Cell Physiol. 2007, 213, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, F.; Carta, L.; Lee-Arteaga, S.; Liu, C.; Nistala, H.; Smaldone, S. Fibrillin-rich microfibrils—Structural and instructive determinants of mammalian development and physiology. Connect. Tissue Res. 2008, 49, 1–6. [Google Scholar] [CrossRef]
- Ramirez, F.; Rifkin, D.B. Extracellular microfibrils: Contextual platforms for TGFbeta and BMP signaling. Curr. Opin. Cell Biol. 2009, 21, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Sengle, G.; Sakai, L.Y. The fibrillin microfibril scaffold: A niche for growth factors and mechanosensation? Matrix Biol. 2015, 47, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hubmacher, D.; Apte, S.S. The biology of the extracellular matrix: Novel insights. Curr. Opin. Rheumatol. 2013, 25, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Halper, J.; Kjaer, M. Basic components of connective tissues and extracellular matrix: Elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. Adv. Exp. Med. Biol. 2014, 802, 31–47. [Google Scholar] [PubMed]
- Wilusz, R.; Sanchez-Adams, J.; Guilak, F. The structure and function of the pericellular matrix of articular cartilage. Matrix Biol. 2014, 39, 25–32. [Google Scholar] [CrossRef]
- Liu, C.; Wang, B.; Xiao, L.; Li, Y.; Xu, L.; Zhao, Z.; Zhang, L. Protective effects of the pericellular matrix of chondrocyte on articular cartilage against the development of osteoarthritis. Histol. Histopathol. 2018, 33, 757–764. [Google Scholar]
- Ocken, A.; Ku, M.M.; Kinzer-Ursem, T.L.; Calve, S. Perlecan knockdown significantly alters extracellular matrix composition and organization during cartilage development. Mol. Cell Proteom. 2020, 19, 1220–1235. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.; Steidle, J.; Danalache, M.; Bonnaire, F.; Walter, C.; Rolauffs, B. Chondrocyte death after mechanically overloading degenerated human intervertebral disk explants is associated with a structurally impaired pericellular matrix. J. Tissue Eng. Regen. Med. 2018, 12, 2000–2010. [Google Scholar] [CrossRef] [PubMed]
- Felka, T.; Rothdiener, M.; Bast, S.; Uynuk-Ool, T.; Zouhair, S.; Ochs, B.G.; de Zwart, P.; Stoeckle, U.; Aicher, W.K.; Hart, M.L.; et al. Loss of spatial organization and destruction of the pericellular matrix in early osteoarthritis in vivo and in a novel in vitro methodology. Osteoarthr. Cartil. 2016, 24, 1200–1209. [Google Scholar] [CrossRef] [Green Version]
- Danalache, M.; Erler, A.L.; Wolfgart, J.M.; Schwitalle, M.; Hofmann, U.K. Biochemical changes of the pericellular matrix and spatial chondrocyte organization-Two highly interconnected hallmarks of osteoarthritis. J. Orthop. Res. 2020, 38, 2170–2180. [Google Scholar] [CrossRef]
- Guilak, F.; Nims, R.J.; Dicks, A.; Wu, C.L.; Meulenbelt, I. Osteoarthritis as a disease of the cartilage pericellular matrix. Matrix Biol. 2018, 71–72, 40–50. [Google Scholar] [CrossRef]
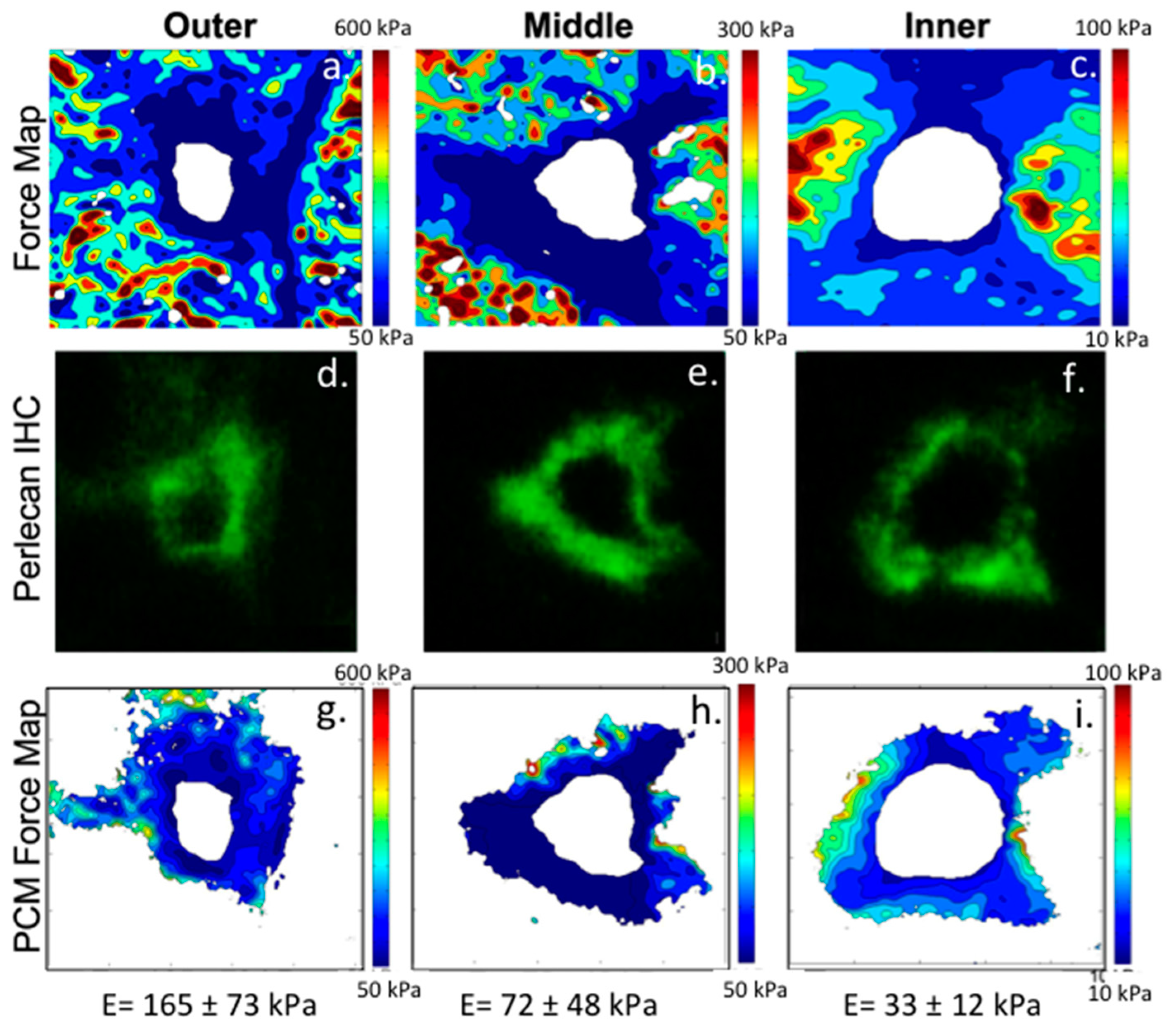
- Wilusz, R.; DeFrate, L.E.; Guilak, F. Immunofluorescence-guided atomic force microscopy to measure the micromechanical properties of the pericellular matrix of porcine articular cartilage. J. R. Soc. Interface 2012, 9, 2997–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilusz, R.; Zauscher, S.; Guilak, F. Micromechanical mapping of early osteoarthritic changes in the pericellular matrix of human articular cartilage. Osteoarthr. Cartil. 2013, 21, 1895–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fermor, H.; McLure, S.W.; Taylor, S.D.; Russell, S.L.; Williams, S.; Fisher, J.; Ingham, E. The elastin network: Its relationship with collagen and cells in articular cartilage as visualized by multiphoton microscopy. Biomed. Mater. Eng. 2015, 25, 381–395. [Google Scholar] [PubMed]
- Zhao, Z.; Li, Y.; Wang, M.; Zhao, S.; Zhao, Z.; Fang, J. Mechanotransduction pathways in the regulation of cartilage chondrocyte homoeostasis. J. Cell Mol. Med. 2020, 24, 5408–5419. [Google Scholar] [CrossRef] [Green Version]
- Khoshgoftar, M.; Torzilli, P.A.; Maher, S.A. Influence of the pericellular and extracellular matrix structural properties on chondrocyte mechanics. J. Orthop. Res. 2018, 36, 721–729. [Google Scholar] [CrossRef]
- Chen, C.; Tambe, D.T.; Deng, L.; Yang, L. Biomechanical properties and mechanobiology of the articular chondrocyte. Am. J. Physiol. Cell Physiol. 2013, 305, C1202–C1208. [Google Scholar] [CrossRef] [Green Version]
- Whitelock, J.M.; Melrose, J.; Iozzo, R.V. Diverse cell signaling events modulated by perlecan. Biochemistry 2008, 47, 11174–11183. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Comparative spatial and temporal immunolocalisation of perlecan, aggrecan and type I, II and IV collagen in the ovine meniscus: An ageing study. Histochem. Cell Biol. 2005, 124, 225–235. [Google Scholar] [CrossRef]
- Green, E.; Mansfield, J.C.; Bell, J.S.; Winlove, C.P. The structure and micromechanics of elastic tissue. Interface Focus 2014, 4, 20130058. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Tirlapur, U.; Fairbank, J.; Handford, P.; Roberts, S.; Winlove, C.P.; Cui, Z.; Urban, J. Microfibrils, elastin fibres and collagen fibres in the human intervertebral disc and bovine tail disc. J. Anat. 2007, 210, 460–471. [Google Scholar] [CrossRef]
- Li, B.; Urban, J.P.; Yu, J. The distribution of fibrillin-2 and LTBP-2, and their co-localisation with fibrillin-1 in adult bovine tail disc. J. Anat. 2012, 220, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Wu, J.P.; Xu, J.; Day, R.E.; Kirk, T.B. Microstructural and compositional features of the fibrous and hyaline cartilage on the medial tibial plateau imply a unique role for the hopping locomotion of kangaroo. PLoS ONE 2013, 8, e74303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Wu, J.P.; Chim, S.M.; Xu, J.; Kirk, T.B. Microstructural analysis of collagen and elastin fibres in the kangaroo articular cartilage reveals a structural divergence depending on its local mechanical environment. Osteoarthr. Cartil. 2013, 21, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, T.; Thibbotuwawa, N.; Gu, Y.; Momot, K.I. MRI magic-angle effect in femorotibial cartilages of the red kangaroo. Magn. Reson. Imaging 2017, 43, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Urban, J.P. The elastic network of articular cartilage: An immunohistochemical study of elastin fibres and microfibrils. J. Anat. 2010, 216, 533–541. [Google Scholar] [CrossRef]
- Mithieux, S.; Weiss, A.S. Elastin. Adv. Protein Chem. 2005, 70, 437–461. [Google Scholar]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Ritty, T.; Ditsios, K.; Starcher, B.C. Distribution of the elastic fiber and associated proteins in flexor tendon reflects function. Anat. Rec. 2002, 268, 430–440. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Wu, J.P.; Kirk, T.B.; Carrino, J.A.; Xiang, C.; Xu, J. High-resolution measurements of the multilayer ultra-structure of articular cartilage and their translational potential. Arthritis Res. Ther. 2014, 16, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, A.; Melrose, J. Glycosaminoglycan and Proteoglycan Biotherapeutics in Articular Cartilage Protection and Repair Strategies: Novel Approaches to Visco-supplementation in Orthobiologics. Adv. Ther. 2019, 2, 1900034. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guilak, F.; Hayes, A.J.; Melrose, J. Perlecan in Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues. Int. J. Mol. Sci. 2021, 22, 2716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052716
Guilak F, Hayes AJ, Melrose J. Perlecan in Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues. International Journal of Molecular Sciences. 2021; 22(5):2716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052716
Chicago/Turabian StyleGuilak, Farshid, Anthony J. Hayes, and James Melrose. 2021. "Perlecan in Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues" International Journal of Molecular Sciences 22, no. 5: 2716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052716