
Vitamin D Compounds PRI-2191 and PRI-2205 Enhance Anastrozole Activity in Human Breast Cancer Models

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
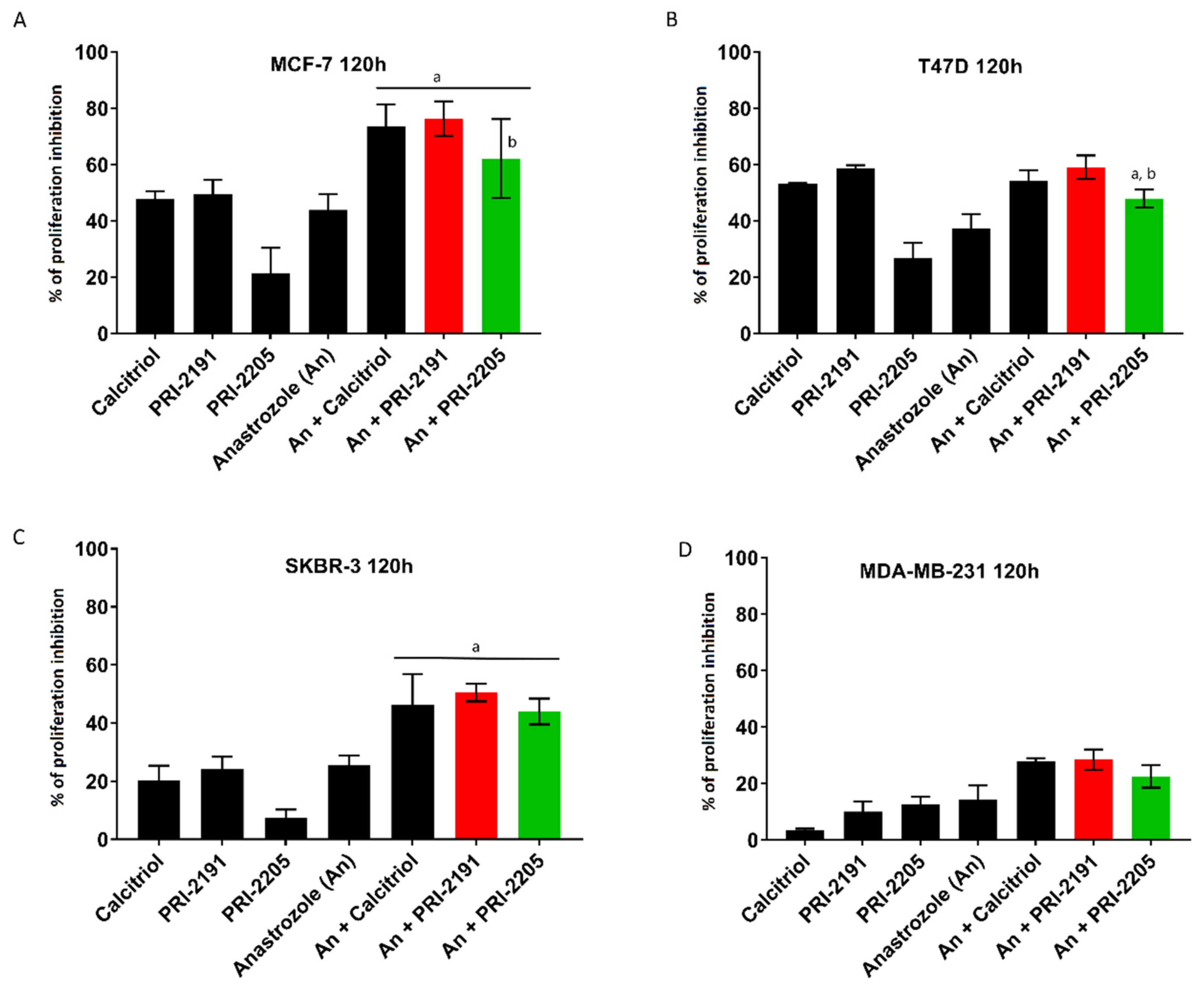
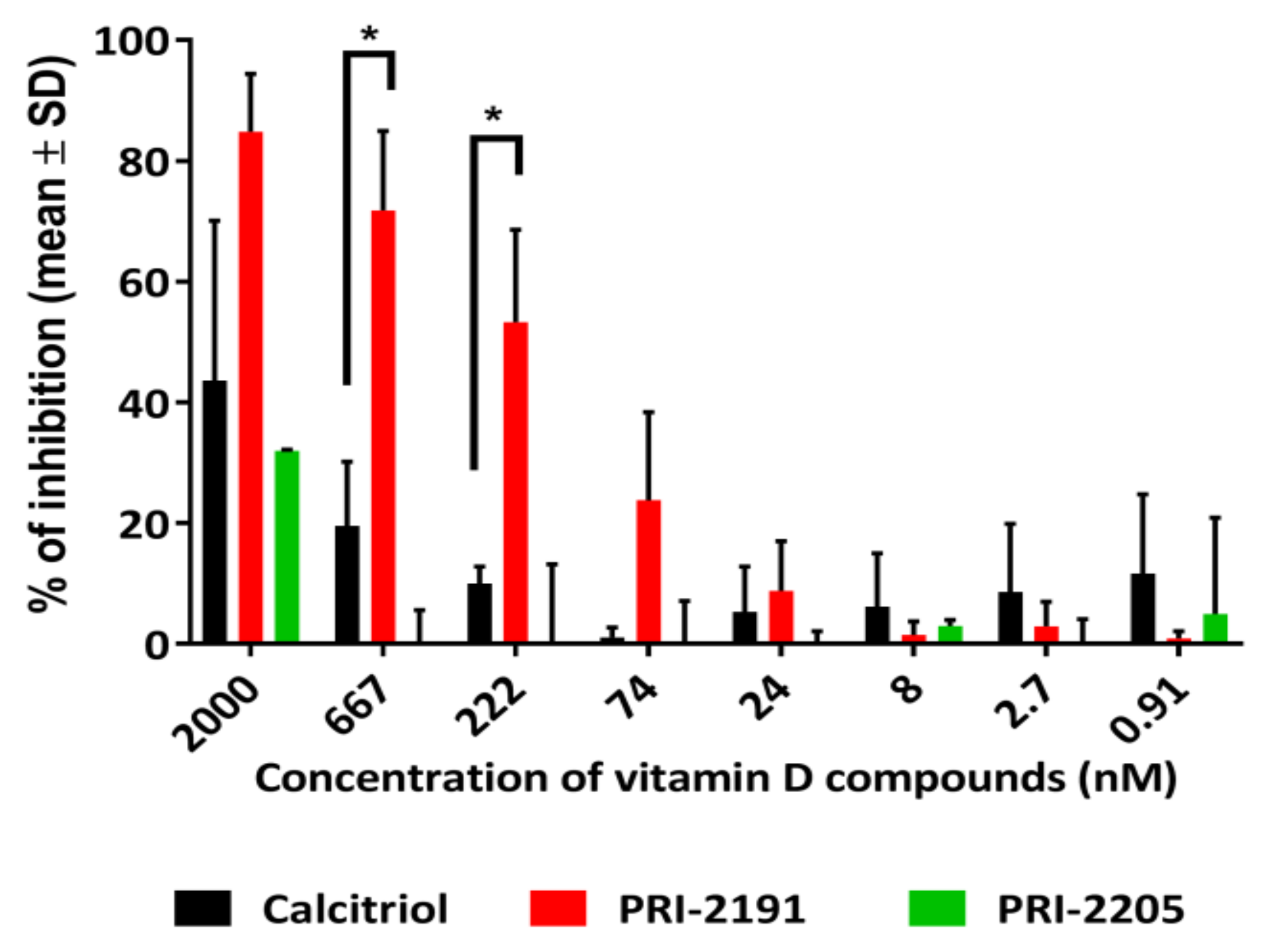
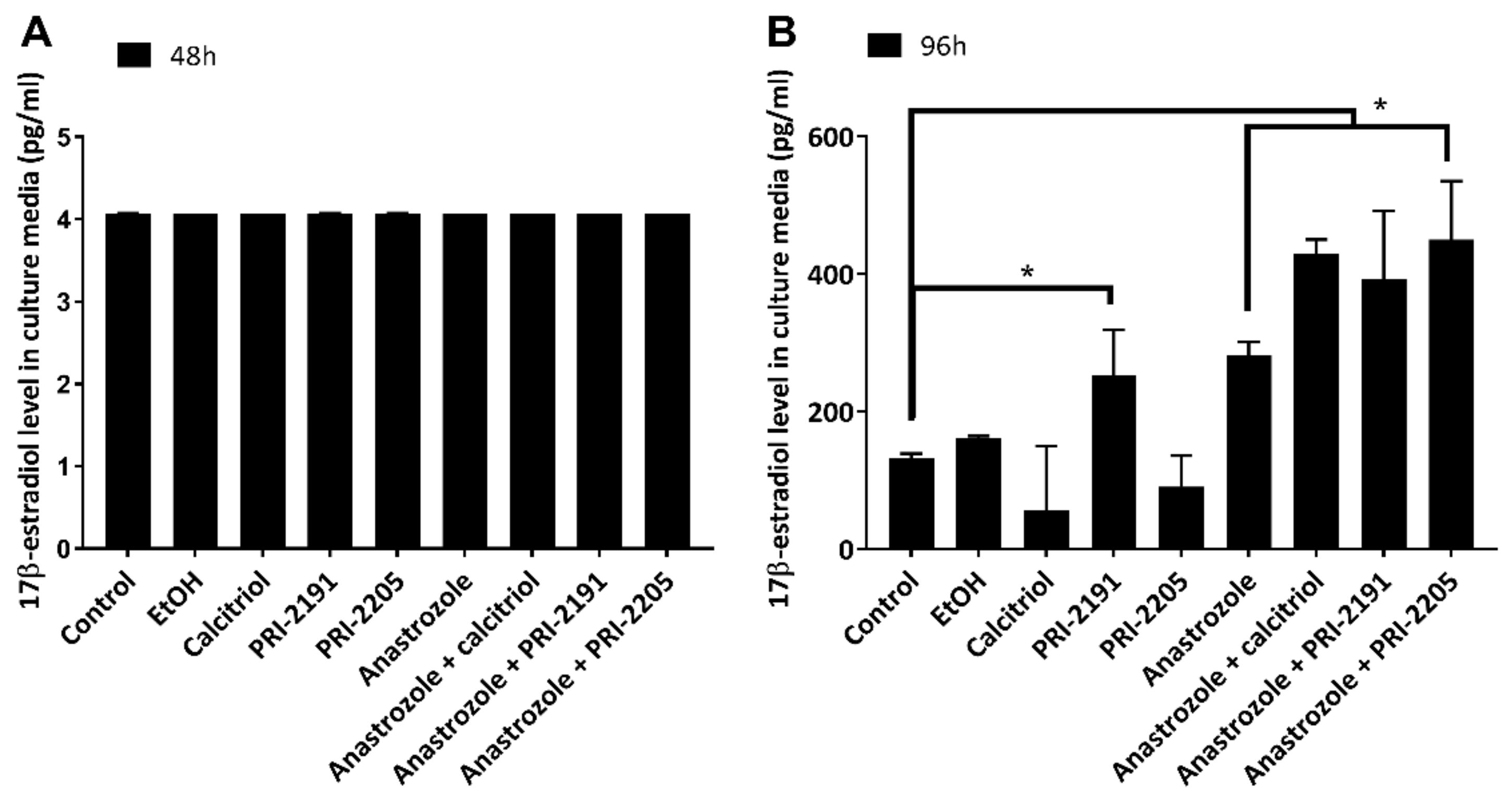
2.1. Calcitriol and Its Analogs Potentiate the Antiproliferative Effect of Anastrozole
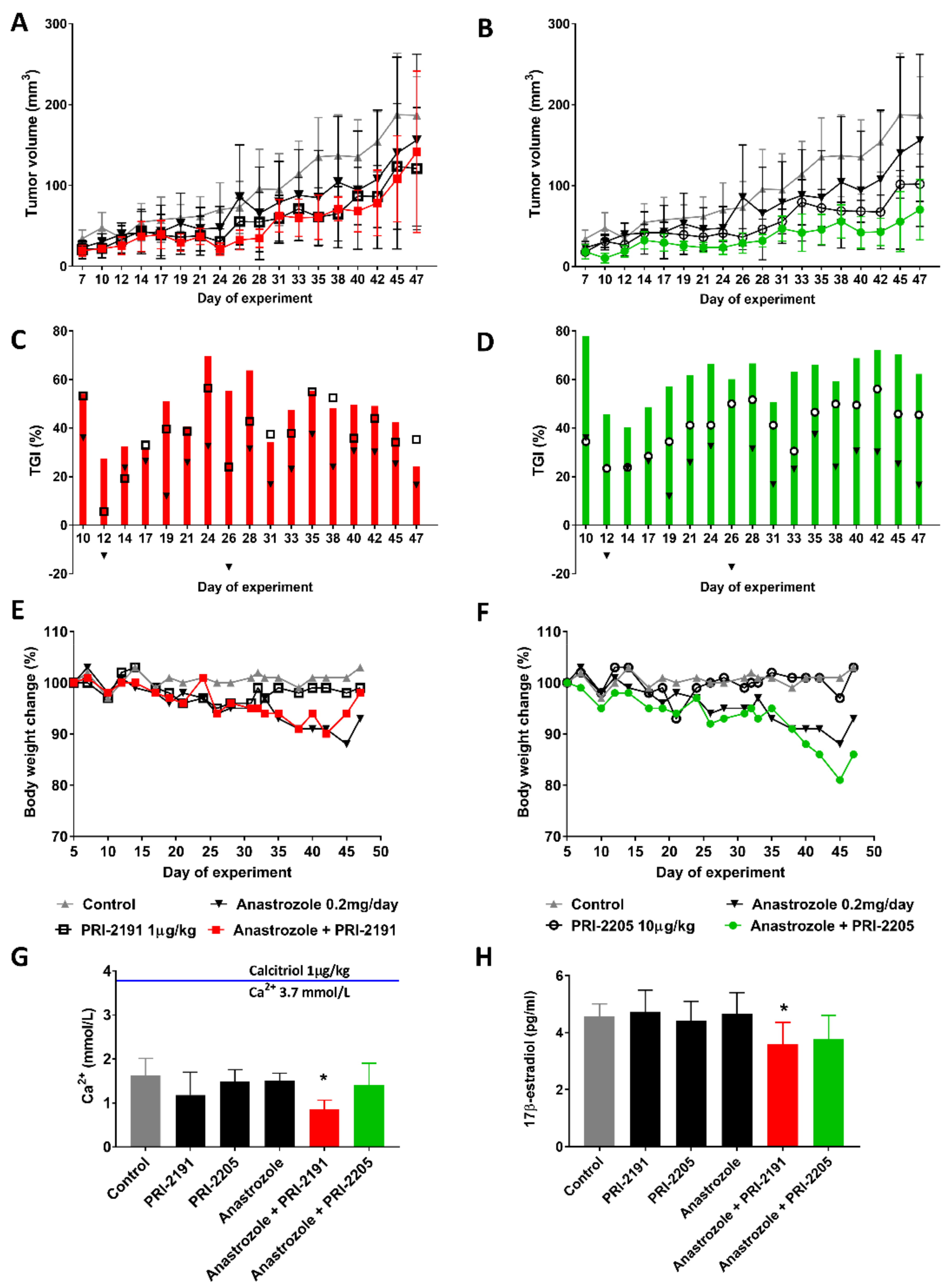
2.2. Effect of Combined Treatment with Anastrozole and Vitamin D Compounds on Mcf-7 Breast Cancer Growth In Vivo
3. Materials and Methods
3.1. Cell Lines
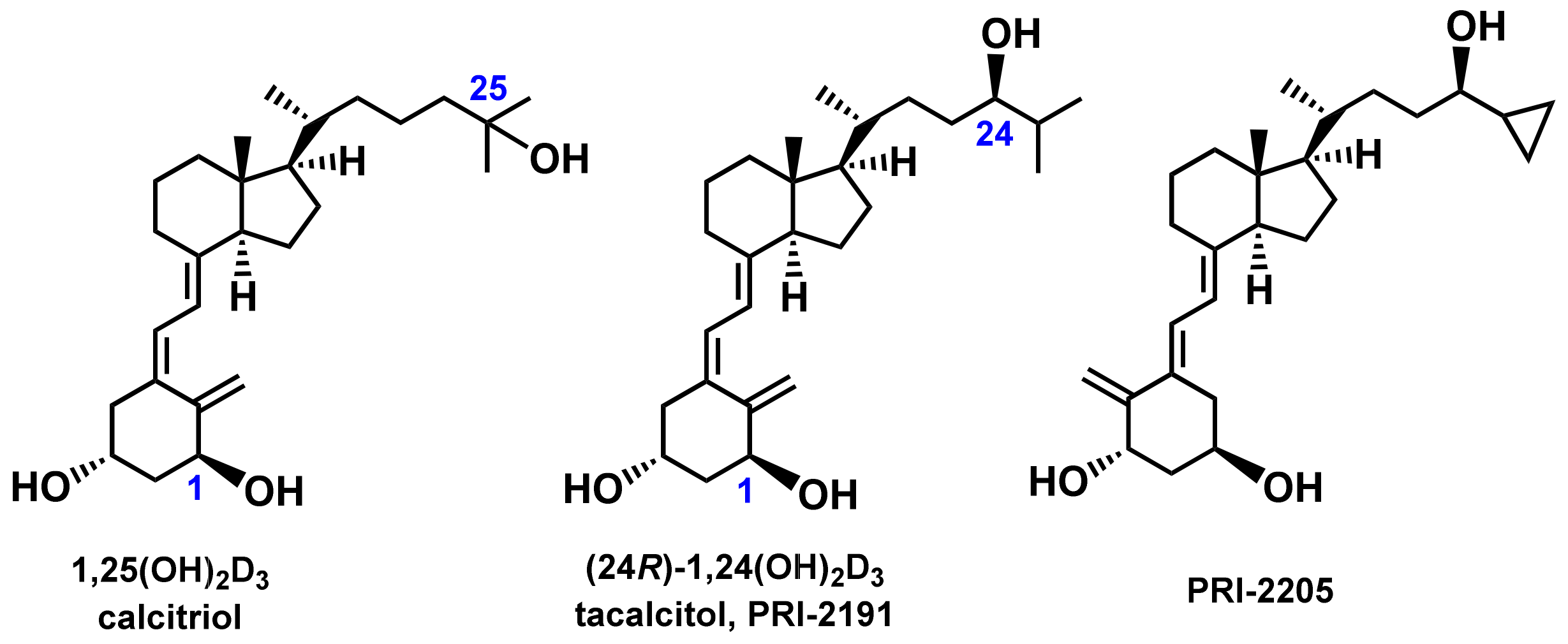
3.2. Compounds
3.3. In Vitro Antiproliferative Assay
3.4. Evaluation of Combination Effects.
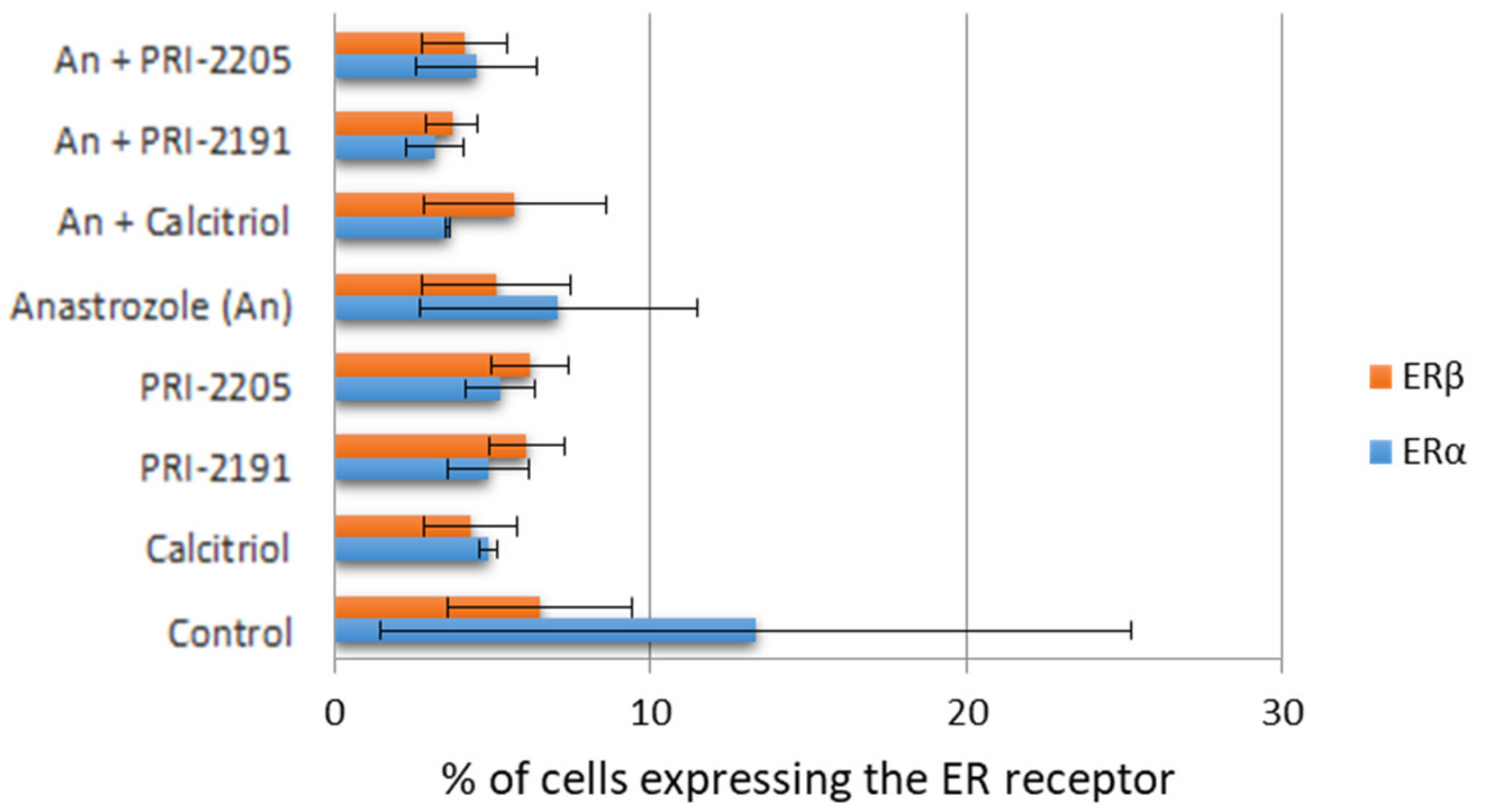
3.5. Estrogen Receptor Expression (Flow Cytometry)
3.6. Estradiol Level Measurement
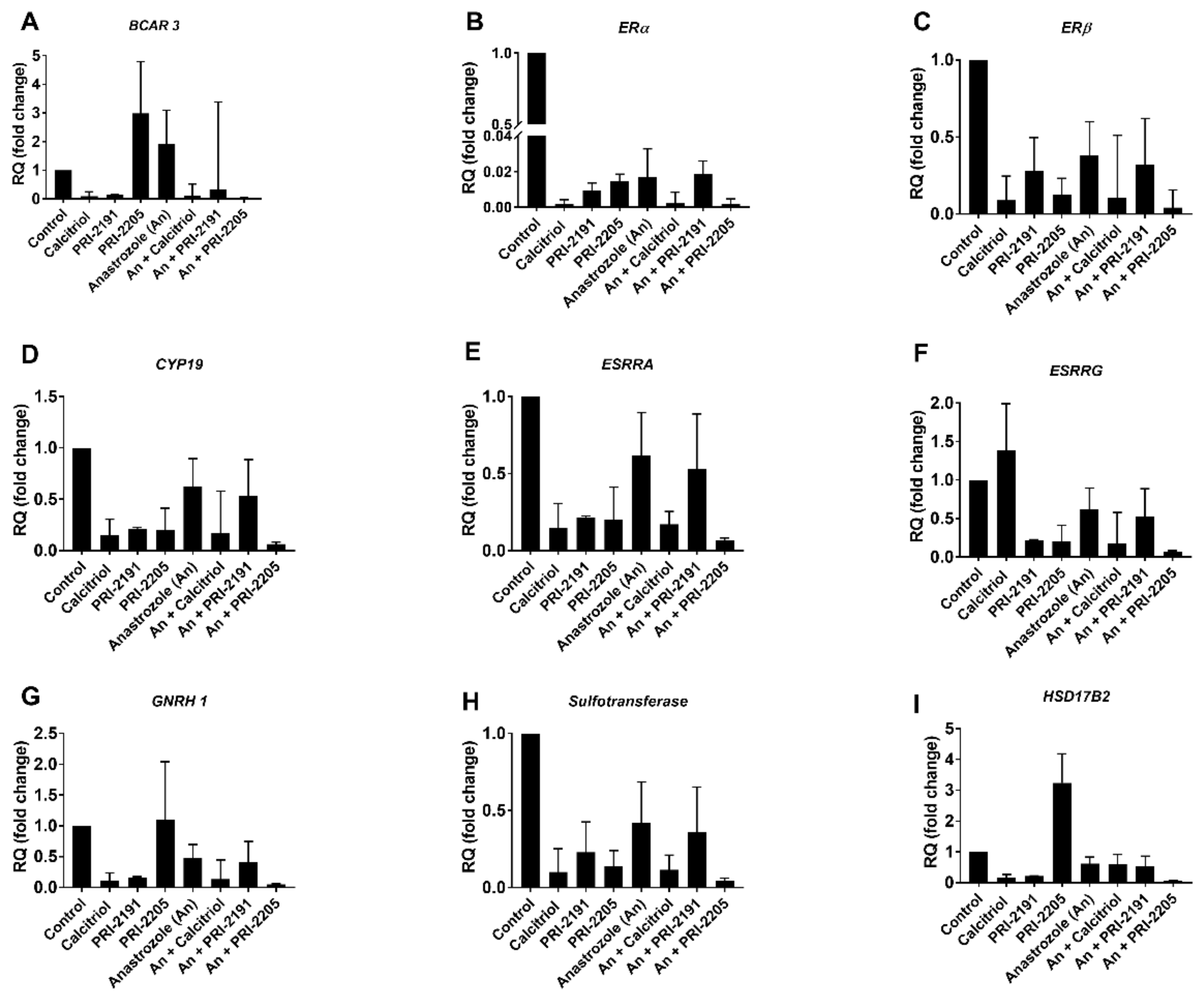
3.7. Estrogen Receptor PCR Array
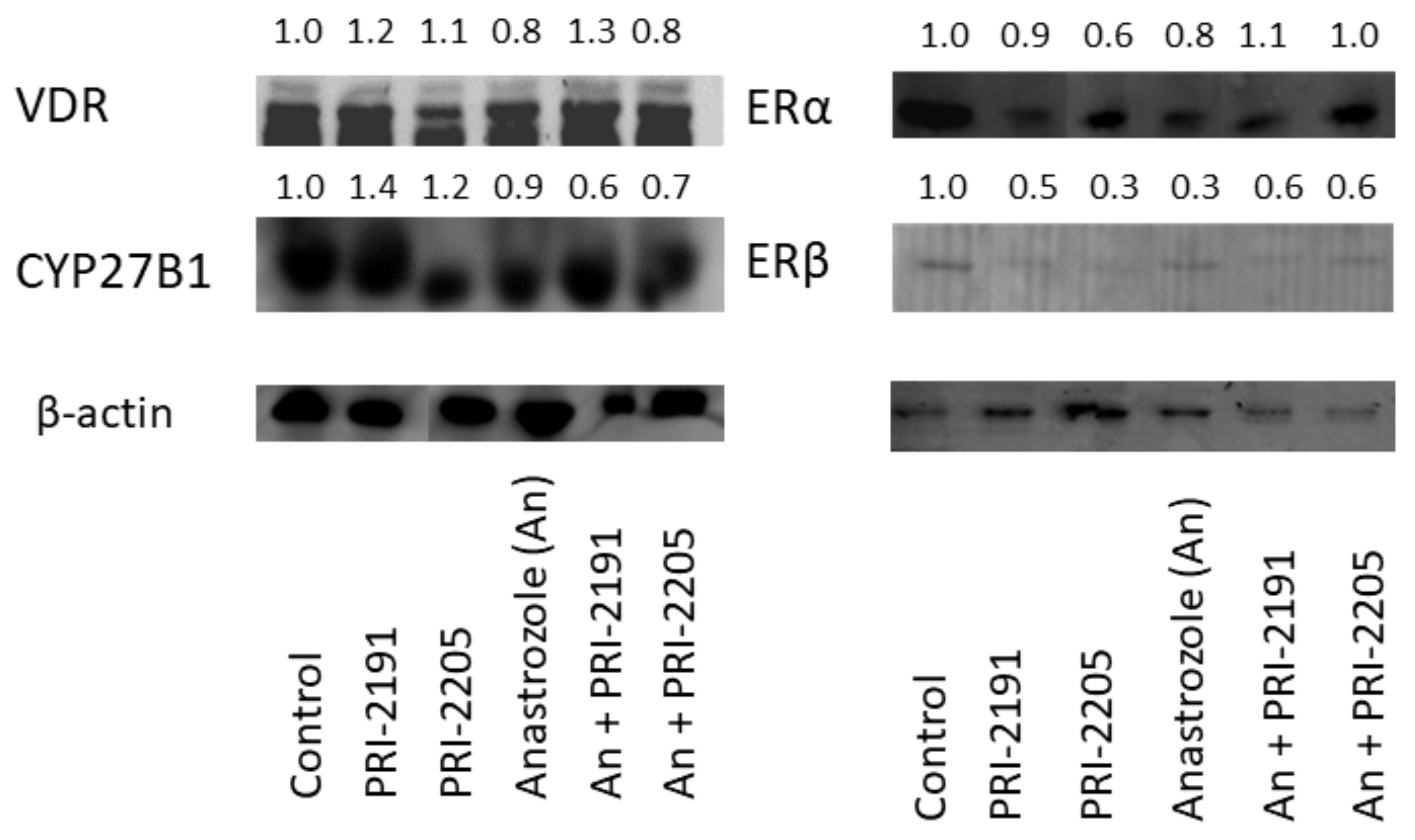
3.8. Western Blot
3.9. Aromatase Activity Assays
3.9.1. Assay I—The Measurement of Estradiol Synthesis (Estrogen Levels in Culture Media or Blood Serum of Treated Mice)
3.9.2. Assay II—Aromatase (CYP19) Enzyme Inhibition by Calcitriol and Its Analogs
3.10. In Vivo Experiments
3.10.1. Mice
3.10.2. Design of In Vivo Experiments
3.10.3. Evaluation of Combination Effects
3.10.4. Body Weight Change
3.10.5. Calcemic Activity
3.10.6. Statistical Evaluation
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AI | Aromatase inhibitor |
| An | Anastrozole |
| ATCC | American Type Culture Collection |
| BWC | Bodyweight change |
| CaSR | Calcium-sensing receptor |
| CYP19 | Gene encoding the cytochrome P450 aromatase |
| CYP27B1 | 25-hydroxyvitamin D3 1α-hydroxylase |
| DMSO | Dimethyl sulfoxide |
| ER | Estrogen receptor |
| EGFR | Epidermal growth factor receptor |
| HER2 | Human epidermal growth factor receptor 2 |
| PBS/PBST | Phosphate-buffered saline/ phosphate-buffered saline + Tween 20 |
| PR | Progesterone receptor |
| PRI | Pharmaceutical Research Institute |
| PVDF | Polyvinyl difluoride |
| SCID | Severe combined immunodeficiency |
| SERD | Selective estrogen receptor downregulator |
| SERM | Selective receptor modulator |
| SRB | Sulforhodamine B |
| TGI | Tumor growth inhibition |
| VDR | Vitamin D receptor |
| VDRE | Vitamin D responsive element |
References
- Kelly, C.M.; Buzdar, A.U. Anastrozole. Exp. Opin. Drug Saf. 2010, 9, 995–1003. [Google Scholar] [CrossRef]
- Shi, Q.; Giordano, S.H.; Lu, H.; Saleeba, A.K.; Malveaux, D.; Cleeland, C.S. Anastrozole-Associated Joint Pain and Other Symptoms in Patients with Breast Cancer. J. Pain 2013, 14, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Romero, S.A.; Su, H.I.; Satagopan, J.; Li, Q.S.; Seluzicki, C.M.; Dries, A.; DeMichele, A.M.; Mao, J.J. Clinical and genetic risk factors for aromatase inhibitor-associated arthralgia in breast cancer survivors. Breast 2020, 49, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, R.E.; Rathbone, E.; Brown, J.E. Management of cancer treatment-induced bone loss. Nat. Rev. Rheumatol. 2013, 9, 365–374. [Google Scholar] [CrossRef]
- Singh, S.; Cuzick, J.; Mesher, D.; Richmond, B.; Howell, A. Effect of baseline serum vitamin D levels on aromatase inhibitors induced musculoskeletal symptoms: Results from the IBIS-II, chemoprevention study using anastrozole. Breast Cancer Res. Treat. 2011, 132, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Alhambra, D.; Javaid, M.K.; Servitja, S.; Arden, N.K.; Martinez-García, M.; Diez-Perez, A.; Albanell, J.; Tusquets, I.; Nogues, X. Vitamin D threshold to prevent aromatase inhibitor-induced arthralgia: A prospective cohort study. Breast Cancer Res. Treat. 2010, 125, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto-Alhambra, D.; Servitja, S.; Javaid, M.K.; Garrigós, L.; Arden, N.K.; Cooper, C.; Albanell, J.; Tusquets, I.; Diez-Perez, A.; Nogues, X. Vitamin D threshold to prevent aromatase inhibitor-related bone loss: The B-ABLE prospective cohort study. Breast Cancer Res. Treat. 2012, 133, 1159–1167. [Google Scholar] [CrossRef]
- Niravath, P.; Hilsenbeck, S.G.; Wang, T.; Jiralerspong, S.; Nangia, J.; Pavlick, A.; Ademuyiwa, F.; Frith, A.; Ma, C.; Park, H.; et al. Randomized controlled trial of high-dose versus standard-dose vitamin D3 for prevention of aromatase inhibitor-induced arthralgia. Breast Cancer Res. Treat. 2019, 177, 427–435. [Google Scholar] [CrossRef]
- Martino, G.; Catalano, A.; Agostino, R.M.; Bellone, F.; Morabito, N.; Lasco, C.G.; Vicario, C.M.; Schwarz, P.; Feldt-Rasmussen, U. Quality of life and psychological functioning in postmenopausal women under-going aromatase inhibitor treatment for early breast cancer. PLoS ONE 2020, 15, e0230681. [Google Scholar] [CrossRef] [PubMed]
- Waltman, N.L.; Ott, C.D.; Twiss, J.J.; Gross, G.J.; Lindsey, A.M. Vitamin D Insufficiency and Musculoskeletal Symptoms in Breast Cancer Survivors on Aromatase Inhibitor Therapy. Cancer Nurs. 2009, 32, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Q.J.; Kimler, B.F.; Reddy, P.S.; Sharma, P.; Klemp, J.R.; Nydegger, J.L.; Yeh, H.-W.; Fabian, C.J. Randomized trial of vitamin D3 to prevent worsening of musculoskeletal symptoms in women with breast cancer receiving adjuvant letrozole. The VITAL trial. Breast Cancer Res. Treat. 2017, 166, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.A.; Abdelmoneim, N.A.; Hewala, T.I.; Rashad, M.M.; Abdaallah, S. Vitamin D Receptor in Breast Cancer Tissues and Its Relation to Estrogen Receptor Alpha (ER-α) Gene Expression and Serum 25-hydroxyvitamin D Levels in Egyptian Breast Cancer Patients: A Case-control Study. Clin. Breast Cancer 2019, 19, e407–e414. [Google Scholar] [CrossRef]
- Verma, A.; Schwartz, Z.; Boyan, B.D. 24R,25-dihydroxyvitamin D3 modulates tumorigenicity in breast cancer in an estrogen receptor-dependent manner. Steroids 2019, 150, 108447. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Swami, S.; Peng, L.; Wang, J.; Moreno, J.; Feldman, D. Tissue-selective regulation of aromatase expression by calcitriol: Implications for breast cancer therapy. Endocrinology 2010, 151, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Niravath, P.; Chen, B.; Chapman, J.-A.W.; Agarwal, S.K.; Welschhans, R.L.; Bongartz, T.; Kalari, K.R.; Shepherd, L.E.; Bartlett, J.; Pritchard, K.; et al. Vitamin D Levels, Vitamin D Receptor Polymorphisms, and Inflammatory Cytokines in Aromatase Inhibitor-Induced Arthralgias: An Analysis of CCTG MA.27. Clin. Breast Cancer 2018, 18, 78–87. [Google Scholar] [CrossRef]
- Brown, R.B. Vitamin D, cancer, and dysregulated phosphate metabolism. Endocrinology 2019, 65, 238–243. [Google Scholar] [CrossRef]
- de La Puente-Yagüe, M.; Cuadrado-Cenzual, M.A.; Ciudad-Cabañas, M.J.; Hernández-Cabria, M.; Collado-Yurrita, L. Vitamin D: And its role in breast cancer. Kaohsiung J. Med. Sci. 2018, 34, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Estébanez, N.; Gómez-Acebo, I.; Palazuelos, C.; Llorca, J.; Dierssen-Sotos, T. Vitamin D exposure and Risk of Breast Cancer: A meta-analysis. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Swami, S.; Feldman, D. Vitamin D and breast cancer: Inhibition of estrogen synthesis and signaling. J. Steroid Biochem. Mol. Biol. 2010, 121, 343–348. [Google Scholar] [CrossRef]
- Swami, S.; Krishnan, A.V.; Wang, J.Y.; Jensen, K.; Peng, L.; Albertelli, M.A.; Feldman, D. Inhibitory Effects of Calcitriol on the Growth of MCF-7 Breast Cancer Xenografts in Nude Mice: Selective Modulation of Aromatase Expression in vivo. Horm. Cancer 2011, 2, 190–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milczarek, M.; Rosinska, S.; Psurski, M.; Maciejewska, M.; Kutner, A.; Wietrzyk, J. Combined colonic cancer treatment with vitamin D analogs and irinotecan or oxaliplatin. Anticancer Res. 2013, 33, 433–444. [Google Scholar]
- Wietrzyk, J.; Pełczyńska, M.; Madej, J.; Dzimira, S.; Kuśnierczyk, H.; Kutner, A.; Szelejewski, W.; Opolski, A. Toxicity and antineoplastic effect of (24R)-1,24-dihydroxyvitamin D 3 (PRI-2191). Steroids 2004, 69, 629–635. [Google Scholar] [CrossRef]
- Wietrzyk, J.; Chodyński, M.; Fitak, H.; Wojdat, E.; Kutner, A.; Opolski, A. Antitumor properties of diastereomeric and geometric analogs of vitamin D3. Anti-Cancer Drugs 2007, 18, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Wietrzyk, J.; Milczarek, M.; Kutner, A. The Effect of Combined Treatment on Head and Neck Human Cancer Cell Lines With Novel Analogs of Calcitriol and Cytostatics. Oncol. Res. 2007, 16, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Wietrzyk, J.; Nevozhay, D.; Filip, B.; Milczarek, M.; Kutner, A. The antitumor effect of lowered doses of cytostatics combined with new analogs of vitamin D in mice. Anticancer Res. 2007, 27, 3387–3398. [Google Scholar] [PubMed]
- Wietrzyk, J.; Nevozhay, D.; Milczarek, M.; Filip, B.; Kutner, A. Toxicity and antitumor activity of the vitamin D analogs PRI-1906 and PRI-1907 in combined treatment with cyclophosphamide in a mouse mammary cancer model. Cancer Chemother. Pharmacol. 2008, 62, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Maj, E.; Filip-Psurska, B.; Milczarek, M.; Psurski, M.; Kutner, A.; Wietrzyk, J. Vitamin D derivatives potentiate the anticancer and anti-angiogenic activity of tyrosine kinase inhibitors in combination with cytostatic drugs in an A549 non-small cell lung cancer model. Int. J. Oncol. 2018, 52, 337–366. [Google Scholar] [PubMed]
- Milczarek, M.; Rossowska, J.; Klopotowska, D.; Stachowicz, M.; Kutner, A.; Wietrzyk, J. Tacalcitol increases the sensitivity of colorectal cancer cells to 5-fluorouracil by downregulating the thymidylate synthase. J. Steroid Biochem. Mol. Biol. 2019, 190, 139–151. [Google Scholar] [CrossRef]
- Milczarek, M.; Chodyński, M.; Filip-Psurska, B.; Martowicz, A.; Krupa, M.; Krajewski, K.; Kutner, A.; Wietrzyk, J. Synthesis and Biological Activity of Diastereomeric and Geometric Analogs of Calcipotriol, PRI-2202 and PRI-2205, Against Human HL-60 Leukemia and MCF-7 Breast Cancer Cells. Cancers 2013, 5, 1355–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopotowska, D.; Matuszyk, J.; Wietrzyk, J. Steroid hormone calcitriol and its analog tacalcitol inhibit miR-125b expression in a human breast cancer MCF-7 cell line. Steroids 2019, 141, 70–75. [Google Scholar] [CrossRef]
- Subik, K.; Lee, J.-F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.-C.; Bonfiglio, T.; Hicks, D.G.; et al. The Expression Patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by Immunohistochemical Analysis in Breast Cancer Cell Lines. Breast Cancer 2010, 4, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trynda, J.; Turlej, E.; Milczarek, M.; Pietraszek, A.; Chodyński, M.; Kutner, A.; Wietrzyk, J. Antiproliferative Activity and in Vivo Toxicity of Double-Point Modified Analogs of 1,25-Dihydroxyergocalciferol. Int. J. Mol. Sci. 2015, 16, 24873–24894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filip, B.; Milczarek, M.; Wietrzyk, J.; Chodyński, M.; Kutner, A. Antitumor properties of (5E,7E) analogs of vitamin D3. J. Steroid Biochem. Mol. Biol. 2010, 121, 399–402. [Google Scholar] [CrossRef]
- Richard, C.L.; Farach-Carson, M.C.; Rohe, B.; Nemere, I.; Meckling, K.A. Involvement of 1,25D3-MARRS (membrane associated, rapid response steroid-binding), a novel vitamin D receptor, in growth inhibition of breast cancer cells. Exp. Cell Res. 2010, 316, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Beilhartz, G.; Roy, Y.; Richard, C.L.; Curtin, M.; Brown, L.; Cadieux, D.; Coppolino, M.; Farach-Carson, M.C.; Nemere, I.; et al. Nuclear translocation of the 1,25D3-MARRS (membrane associated rapid response to steroids) receptor protein and NFκB in differentiating NB4 leukemia cells. Exp. Cell Res. 2010, 316, 1101–1108. [Google Scholar] [CrossRef]
- Wang, X.; Chen, W.; Singh, N.; Promkan, M.; Liu, G. Effects of potential calcium sensing receptor inducers on promoting chemosensitivity of human colon carcinoma cells. Int. J. Oncol. 2010, 36, 1573–1580. [Google Scholar]
- Segovia-Mendoza, M.; Díaz, L.; Prado-Garcia, H.; Reginato, M.J.; Larrea, F.; García-Becerra, R. The addition of calcitriol or its synthetic analog EB1089 to lapatinib and neratinib treatment inhibits cell growth and promotes apoptosis in breast cancer cells. Am. J. Cancer Res. 2017, 7, 1486–1500. [Google Scholar]
- Segovia-Mendoza, M.; Díaz, L.; González-González, M.E.; Martínez-Reza, I.; García-Quiroz, J.; Prado-Garcia, H.; Ibarra-Sánchez, M.J.; Esparza-López, J.; Larrea, F.; García-Becerra, R. Calcitriol and its analogues enhance the antiproliferative activity of gefitinib in breast cancer cells. J. Steroid Biochem. Mol. Biol. 2015, 148, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Reza, I.; Díaz, L.; Barrera, D.; Segovia-Mendoza, M.; Pedraza-Sánchez, S.; Soca-Chafre, G.; Larrea, F.; García-Becerra, R. Calcitriol inhibits the proliferation of triple-negative breast cancer cells through a mechanism involving the proinflammatory cytokines IL-1β and TNF-α. J. Immunol. Res. 2019, 2019, 6384278. [Google Scholar] [CrossRef] [Green Version]
- Peters, G.; van der Wilt, C.; van Moorsel, C.; Kroep, J.; Bergman, A.; Ackland, S. Basis for effective combination cancer chemotherapy with antimetabolites. Pharmacol. Ther. 2000, 87, 227–253. [Google Scholar] [CrossRef]
- Thill, M.; Hoellen, F.; Becker, S.; Dittmer, C.; Fischer, D.; Kümmel, S.; Salehin, D.; Friedrich, M.; Köster, F.; Diedrich, K.; et al. Expression of prostaglandin- and vitamin D-metabolising enzymes in benign and malignant breast cells. Anticancer Res. 2012, 32, 367–372. [Google Scholar]
- Aparna, R.; Subhashini, J.; Roy, K.R.; Reddy, G.S.; Robinson, M.; Uskokovic, M.R.; Reddanna, P.; Reddy, G.V. Selective inhibition of cyclooxygenase-2 (COX-2) by 1α,25-dihydroxy-16-ene-23-yne-vitamin D3, a less calcemic vitamin D analog. J. Cell. Biochem. 2008, 104, 1832–1842. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, J.; Hansen, S.K.; Lykkesfeldt, A.E. Vitamin D analog EB1089 inhibits aromatase expression by dissociation of co-modulator WSTF from the CYP19A1 promoter-a new regulatory pathway for aromatase. Biochim. Biophys. Acta 2013, 1833, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Merhi, Z.; Doswell, A.; Krebs, K.; Cipolla, M. Vitamin D Alters Genes Involved in Follicular Development and Steroidogenesis in Human Cumulus Granulosa Cells. J. Clin. Endocrinol. Metab. 2014, 99, E1137–E1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, L.J.; Ulrichsen, S.P.; Ahern, T.P.; Goodman, M.; McCullough, L.E.; Waller, L.A.; Christensen, K.B.; Damkier, P.; Hamilton-Dutoit, S.; Lauridsen, K.L.; et al. 17β-Hydroxysteroid dehydrogenase 1:2 and breast cancer recurrence: A Danish population-based study. Acta Oncol. 2020, 59, 329–333. [Google Scholar] [CrossRef]
- Zhang, C.-Y.; Calvo, E.-L.; Yang, C.-Q.; Liu, J.; Sang, X.-Y.; Lin, S.-X. Transcriptome of 17β-hydroxysteroid dehydrogenase type 2 plays both hormone-dependent and hormone-independent roles in MCF-7 breast cancer cells. J. Steroid Biochem. Mol. Biol. 2019, 195, 105471. [Google Scholar] [CrossRef] [PubMed]
- Maj, E.; Filip-Psurska, B.; Świtalska, M.; Kutner, A.; Wietrzyk, J. Vitamin D Analogs Potentiate the Antitumor Effect of Imatinib Mesylate in a Human A549 Lung Tumor Model. Int. J. Mol. Sci. 2015, 16, 27191–27207. [Google Scholar] [CrossRef] [Green Version]
- Welsh, J. Vitamin D and breast cancer: Past and present. J. Steroid Biochem. Mol. Biol. 2018, 177, 15–20. [Google Scholar] [CrossRef]
- Griffin, N.; Dowling, M. Vitamin D supplementation and clinical outcomes in cancer survivorship. Br. J. Nurs. 2018, 27, 1121–1128. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Cohen, D.J.; Schwartz, N.; Muktipaty, C.; Koblinski, J.E.; Boyan, B.D.; Schwartz, Z. 24R,25-Dihydroxyvitamin D3 reg-ulates breast cancer cells in vitro and in vivo. Biochim. Biophys. Acta 2019, 1863, 1498–1512. [Google Scholar] [CrossRef]
- DeLuca, H.F. History of the discovery of vitamin D and its active metabolites. BoneKEy Rep. 2014, 3, 479. [Google Scholar] [CrossRef] [Green Version]
- Lips, P.; Gielen, E.; van Schoor, N.M. Vitamin D supplements with or without calcium to prevent fractures. BoneKEY Rep. 2014, 3, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisiewicz, A.; Kowalski, K.; Banach, J.; Łabędź, N.; Stachowicz-Suhs, M.; Piotrowska, A.; Milczarek, M.; Kłopotowska, D.; Dzięgiel, P.; Wietrzyk, J. Vitamin D Metabolite Profile in Cholecalciferol- or Calcitriol-Supplemented Healthy and Mammary Gland Tumor-Bearing Mice. Nutrition 2020, 12, 3416. [Google Scholar] [CrossRef]
- Hilborn, E.; Stål, O.; Jansson, A. Estrogen and androgen-converting enzymes 17β-hydroxysteroid dehydrogenase and their involvement in cancer: With a special focus on 17β-hydroxysteroid dehydrogenase type 1, 2, and breast cancer. Oncotarget 2017, 8, 30552–30562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Itoh, S.; Takeuchi, Y. Effectiveness of bisphosphonate combined with activated vitamin D in patients with aro-matase inhibitor-induced osteoporosis after breast cancer operation. Osteoporos. Sarcopenia 2018, 4, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.; Wietzke, J.A.; Zinser, G.M.; Smyczek, S.; Romu, S.; Tribble, E.; Welsh, J.C.; Byrne, B.; Narvaez, C.J. Impact of the Vitamin D3 receptor on growth-regulatory pathways in mammary gland and breast cancer. J. Steroid Biochem. Mol. Biol. 2002, 83, 85–92. [Google Scholar] [CrossRef]
- Kemmis, C.M.; Salvador, S.M.; Smith, K.M.; Welsh, J. Human Mammary Epithelial Cells Express CYP27B1 and Are Growth Inhibited by 25-Hydroxyvitamin D-3, the Major Circulating Form of Vitamin D-3. J. Nutr. 2006, 136, 887–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swami, S.; Krishnan, A.V.; Feldman, D. 1alpha,25-Dihydroxyvitamin D3 down-regulates estrogen receptor abundance and suppresses estrogen actions in MCF-7 human breast cancer cells. Clin. Cancer Res. 2000, 6, 3371–3379. [Google Scholar] [PubMed]
- Anisiewicz, A.; Filip-Psurska, B.; Pawlik, A.; Nasulewicz-Goldeman, A.; Piasecki, T.; Kowalski, K.; Maciejewska, M.; Jarosz, J.; Banach, J.; Papiernik, D.; et al. Calcitriol Analogues Decrease Lung Metastasis but Impair Bone Metabolism in Aged Ovariectomized Mice Bearing 4T1 Mammary Gland Tumours. Aging Dis. 2019, 10, 977–991. [Google Scholar] [CrossRef] [Green Version]
- Psurski, M.; Janczewski, Ł.; Świtalska, M.; Gajda, A.; Goszczyński, T.M.; Oleksyszyn, J.; Wietrzyk, J.; Gajda, T. Novel phosphonate analogs of sulforaphane: Synthesis, in vitro and in vivo anticancer activity. Eur. J. Med. Chem. 2017, 132, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Nevozhay, D. Cheburator Software for Automatically Calculating Drug Inhibitory Concentrations from In Vitro Screening Assays. PLoS ONE 2014, 9, e106186. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Type- | Receptors |
|---|---|---|
| MCF-7 | human breast adenocarcinoma, estrogen receptor positive | VDR (+), ER (+), PR (±), HER2(−), EGFR (+) |
| MDA-MB-231 | human breast adenocarcinoma, grade III, triple-negative | VDR (±), ER (−), PR (−), HER2 (−), EGFR (+) |
| SKBR-3 | human breast adenocarcinoma, HER-2-positive | VDR (±), ER (−), PR (−), HER-2 (+) |
| T47D | human ductal carcinoma, estrogen receptor-positive | VDR (+), ER (+), PR (+), HER2 (+), EGFR (+) |
| No | Gene and the Product Name |
|---|---|
| 1 | ADCYAP1 adenylate cyclase-activating polypeptide 1 (pituitary) |
| 2 | BCAR1 breast cancer anti-estrogen resistance 1 |
| 3 | BCAR3 breast cancer anti-estrogen resistance 3 |
| 4 | CARM1 coactivator-associated arginine methyltransferase 1 |
| 5 | CEBPB CCAAT/enhancer-binding protein (C/EBP), beta |
| 6 | CYP19A1 cytochrome P450, family 19, subfamily A, polypeptide 1 |
| 7 | CYP1B1 cytochrome P450, family 1, subfamily B, polypeptide 1 |
| 8 | DDX54 DEAD (Asp-Glu-Ala-Asp) box polypeptide 54 |
| 9 | EBAG9 estrogen receptor binding site associated, antigen, 9 |
| 10 | EGF epidermal growth factor |
| 11 | ERG v-ets erythroblastosis virus E26 oncogene homolog (avian) |
| 12 | ESR1 estrogen receptor 1 |
| 13 | ESR2 estrogen receptor 2 (ER beta) |
| 14 | ESRRA estrogen-related receptor alpha |
| 15 | ESRRG estrogen-related receptor gamma |
| 16 | FGF2 fibroblast growth factor 2 (basic) |
| 17 | FOXO1 forkhead box O1 |
| 18 | FSHB follicle-stimulating hormone, beta polypeptide |
| 19 | FSHR follicle-stimulating hormone receptor |
| 20 | GABARAPL1 GABA(A) receptor-associated protein-like 1 |
| 21 | GHRH growth hormone-releasing hormone |
| 22 | GNL3 guanine nucleotide-binding protein-like 3 (nucleolar) |
| 23 | GNRH1 gonadotropin-releasing hormone 1 (luteinizing-releasing hormone) |
| 24 | HMGA1 high mobility group AT-hook 1 |
| 25 | HMGB1 high mobility group box 1 |
| 26 | HMGB2 high mobility group box 2 |
| 27 | HSD17B2 hydroxysteroid (17-beta) dehydrogenase 2 |
| 28 | HSD17B8 hydroxysteroid (17-beta) dehydrogenase 8 |
| 29 | HSPB8 heat shock 22kDa protein 8 |
| 30 | IGF1 insulin-like growth factor 1 (somatomedin C) |
| 31 | IL1A interleukin 1, alpha |
| 32 | IL1B interleukin 1, beta |
| 33 | INHA inhibin, alpha |
| 34 | ISG20 interferon-stimulated exonuclease gene 20 kDa |
| 35 | LHCGR luteinizing hormone/choriogonadotropin receptor |
| 36 | TSKU tsukushi small leucine rich proteoglycan homolog (Xenopus laevis) |
| 37 | MKNK2 MAP kinase interacting serine/threonine kinase 2 |
| 38 | MPG N-methylpurine-DNA glycosylase |
| 39 | MTA1 metastasis associated 1 |
| 40 | NCOA1 nuclear receptor coactivator 1 |
| 41 | NCOA3 nuclear receptor coactivator 3 |
| 42 | NCOA5 nuclear receptor coactivator 5 |
| 43 | NCOA6 nuclear receptor coactivator 6 |
| 44 | NCOA7 nuclear receptor coactivator 7 |
| 45 | NFATC4 nuclear factor of activated T-cells, cytoplasmic, calcineurin-dependent 4 |
| 46 | NFKB1 nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 |
| 47 | NR0B2 nuclear receptor subfamily 0, group B, member 2 |
| 48 | NR1I3 nuclear receptor subfamily 1, group I, member 3 |
| 49 | NR2C2 nuclear receptor subfamily 2, group C, member 2 |
| 50 | NR6A1 nuclear receptor subfamily 6, group A, member 1 |
| 51 | NRG1 neuregulin 1 |
| 52 | NRIP1 nuclear receptor-interacting protein 1 |
| 53 | OVGP1 oviductal glycoprotein 1, 120kDa |
| 54 | PELP1 proline-, glutamate- and leucine-rich protein 1 |
| 55 | PGR progesterone receptor |
| 56 | PHB2 prohibitin 2 |
| 57 | PLG plasminogen |
| 58 | POU4F1 POU class 4 homeobox 1 |
| 59 | PPARA peroxisome proliferator-activated receptor alpha |
| 60 | PPARGC1A peroxisome proliferator-activated receptor gamma, coactivator 1 alpha |
| 61 | PPARGC1B peroxisome proliferator-activated receptor gamma, coactivator 1 beta |
| 62 | PPID peptidylprolyl isomerase D |
| 63 | RERG RAS-like, estrogen-regulated, growth inhibitor |
| 64 | RLN1 relaxin 1 |
| 65 | SAFB scaffold attachment factor B |
| 66 | SAFB2 scaffold attachment factor B2 |
| 67 | SRD5A2 steroid-5-alpha-reductase, alpha polypeptide 2 (3-oxo-5 alpha-steroid delta 4-dehydrogenase alpha 2) |
| 68 | SREBF1 sterol regulatory element-binding transcription factor 1 |
| 69 | STS In multiple Geneids |
| 70 | SULT1E1 sulfotransferase family 1E, estrogen-preferring, member 1 |
| 71 | TAF10 RNA polymerase II, TATA box binding protein (TBP)-associated factor, 30kDa |
| 72 | TCF7 transcription factor 7 (T-cell specific, HMG-box) |
| 73 | TFF1 trefoil factor 1 |
| 74 | TNF tumor necrosis factor |
| 75 | TRIM16 tripartite motif containing 16 |
| 76 | TRIM25 tripartite motif containing 25 |
| 77 | UGT1A8 UDP glucuronosyltransferase 1 family, polypeptide A8 |
| 78 | UGT1A3 UDP glucuronosyltransferase 1 family, polypeptide A3 |
| 79 | UGT1A4 UDP glucuronosyltransferase 1 family, polypeptide A4 |
| 80 | UGT2A1 UDP glucuronosyltransferase 2 family, polypeptide A1, complex locus |
| 81 | UGT2B15 UDP glucuronosyltransferase 2 family, polypeptide B15 |
| 82 | UGT2B4 UDP glucuronosyltransferase 2 family, polypeptide B4 |
| 83 | UGT2B7 UDP glucuronosyltransferase 2 family, polypeptide B7 |
| 84 | NR0B1 nuclear receptor subfamily 0, group B, member 1 |
| 85 | GNRH2 gonadotropin-releasing hormone 2 |
| 86 | NRG3 neuregulin 3 |
| 87 | NRG4 neuregulin 4 |
| 88 | AR androgen receptor |
| 89 | ACTB Actin, beta |
| 90 | B2M Beta-2-microglobulin |
| 91 | GAPDH Glyceraldehyde-3-phosphate dehydrogenase |
| 92 | GUSB Glucuronidase, beta |
| 93 | HPRT1 Hypoxanthine phosphoribosyltransferase 1 |
| 94 | PGK1 Phosphoglycerate kinase 1 |
| 95 | PPIA Peptidylprolyl isomerase A |
| 96 | RPL13A Ribosomal protein L13a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filip-Psurska, B.; Psurski, M.; Anisiewicz, A.; Libako, P.; Zbrojewicz, E.; Maciejewska, M.; Chodyński, M.; Kutner, A.; Wietrzyk, J. Vitamin D Compounds PRI-2191 and PRI-2205 Enhance Anastrozole Activity in Human Breast Cancer Models. Int. J. Mol. Sci. 2021, 22, 2781. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052781
Filip-Psurska B, Psurski M, Anisiewicz A, Libako P, Zbrojewicz E, Maciejewska M, Chodyński M, Kutner A, Wietrzyk J. Vitamin D Compounds PRI-2191 and PRI-2205 Enhance Anastrozole Activity in Human Breast Cancer Models. International Journal of Molecular Sciences. 2021; 22(5):2781. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052781
Chicago/Turabian StyleFilip-Psurska, Beata, Mateusz Psurski, Artur Anisiewicz, Patrycja Libako, Ewa Zbrojewicz, Magdalena Maciejewska, Michał Chodyński, Andrzej Kutner, and Joanna Wietrzyk. 2021. "Vitamin D Compounds PRI-2191 and PRI-2205 Enhance Anastrozole Activity in Human Breast Cancer Models" International Journal of Molecular Sciences 22, no. 5: 2781. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052781