
Genetic Involvement of Mycobacterium avium Complex in the Regulation and Manipulation of Innate Immune Functions of Host Cells


Abstract
:1. Introduction
2. Comparative Genomics of MAC
2.1. Recent Advance in Classification and Identification of MAC Organisms

{kind=link}
{kind=link}
| Species | Strain | Host | Isolate Origin | Genetic Features | Clinical Features | Year of First Description | References | |
|---|---|---|---|---|---|---|---|---|
| IS * | ITS1 ** | |||||||
| I. Mycobacterium avium subspecies | ||||||||
| Mycobacteriumavium subsp. avium | ATCC25291 | Chicken | Denmark | IS901 + IS1245 + | Mav-A | Avian tuberculosis | 1990 | [26] |
| Mycobacteriumavium subsp. hominissuis | IWGMT49 | Pig | Netherlands | IS901 – IS1245 + | Mav-A | Pulmonary disease | 2002 | [21] |
| Mycobacteriumavium subsp. paratuberculosis | ATCC19698 | Cow | USA | IS900 + | Mav-A | Johne’s disease | 1990 | [26] |
| Mycobacterium avium subsp. silvaticum | ATCC49884 | Wood pigeon | France | IS901 + IS1245 + | Mav-A | tuberculosis in birds and paratuberculosis in mammals | 1990 | [26] |
| II. Species/subspecies closely related to Mycobacterium intracellulare | ||||||||
| Mycobacteriumintracellulare | ATCC15985 | Human | - | - | Min-A | Pulmonary disease | 1965 | [10] |
| Mycobacterium chimaera | DSM44623 CIP 107892 | Human | Italy | IS900 – IS901 – IS1245 - | MAC-A | Pulmonary disease | 2004 | [14] |
| Mycobacterium colombiense | CIP108962 | Human | Colombia | MAC-X | Bacteremia; lymphadenopathy | 2006 | [11] | |
| Mycobacterium arosiense | DSM45069 | Human | - | - | - | Disseminated osteomyelitis in immunocompromised child | 2008 | [15] |
| Mycobacterium vulneris | DSM45247 CIP 109859 | Human | Netherlands | MAC-Q | A suppurative wound consequent to a dog bite/ cervical lymphadenitis in a child | 2009 | [16] | |
| Mycobacterium bouchedurhonense | CIP109827 | Human | France | - | - | Pulmonary disease | 2009 | [17] |
| Mycobacterium marseillense | CIP109828 | Human | France | - | - | Pulmonary disease with bilateral bronchiectasis and multiple nodule | 2009 | [17] |
| Mycobacterium timonense | CIP109830 | Human | France | - | MAC-K | Pulmonary disease | 2009 | [17] |
| Mycobacteriumparaintracellulare | KCTC 29084 | Human | Korea | - | MIN-A | Pulmonary disease | 2016 | [18] |
| Mycobacterium intracellulare subsp. yongonense | DSM45126 | Human | Korea | - | - | Pulmonary disease | 2013 | [19] |
| Mycobacterium indicus pranii | MTCC 9506 DSM 45239 | - | India | - | - | Leprosy vaccine candidates (previously referred to Mycobacterium ‘w’) | 2008 | [20] |
2.2. Difference in Evolutionary Modes among MAC Subpsecies
2.3. Genetic Differences in Host Specificity and Physiological Characteristics of MAC
2.4. Genotypic and Genetic Explanation of MAC (Sub)Species in Human Disease
3. Virulence Gene-Associated Adaptation Strategies of MAC during Pathogenesis
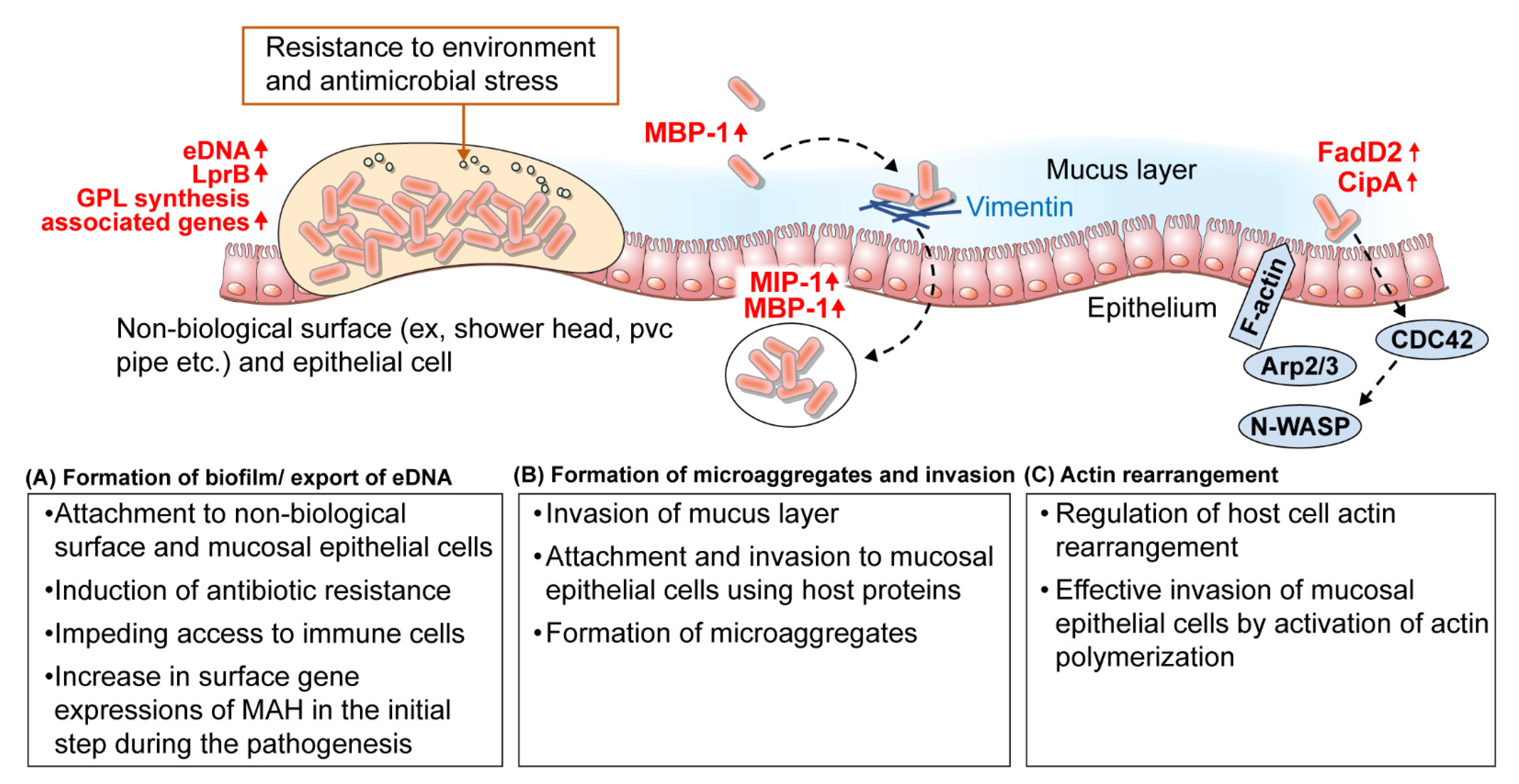
3.1. Mucosal Epithelial Cell Adhesion and Invasion
| Host Defense | Gene | Description | Strategy of MAH | References | ||
|---|---|---|---|---|---|---|
| Action | Description | Strain | ||||
| Strategy 1. Mucosal epithelial cell adhesion and invasion | ||||||
| Mucus layer/mucosal epithelial cell | ag85 mpa51 | - antigen 85 - M. avium MPB51 | attachment | ▪ Binding to fibronectin ▪ Epithelial cell adhesion using host protein | MAH ATCC 15769 | [64,66] |
| MAV_3013 (MBP-1) | - microaggregate binding protein 1 | invasion/ attachment | ▪ Binding to vimentin ▪ Invasion of mucus layer ▪ Epithelial cell adhesion using host protein ▪ Formation of microaggregate | MAH 104 | [61,69] | |
| MAV_4504 | - ABC transporter, ATP-binding protein coordinates | transport | ▪ Translocation of MBP-1 to the bacterial surface | MAH 104 | [69] | |
| MAV_1799 | - hypothetical protein | aggregation | ▪ Rapid recruitment of planktonic bacteria ▪ Formation of microaggregate | MAH 104 | [69] | |
| MAV_0831 (MIP-1) | - microaggregate Invasion Protein-1 | invasion/attachment | ▪ Binding to flaminA ▪ Invasion of epithelial cell | MAH 104 | [69] | |
| fadD2 cipA | - fatty acyl coenzyme A synthase - a domain similar to the PXXP motif of the human piccolo protein | invasion | ▪ Activation of Cdc42 signaling pathway ▪ Rearrangement cytoskeleton | MAH 109 | [72] | |
| accA2, sucA, pstB | - acetyl/propionyl-CoA carboxylase (subunit) - 2-Oxoglutarate dehydrogenase - protein synthetase | attachment | ▪ Association with GPL biosynthesis ▪ Formation of biofilm | MAH A5 | [67] | |
| guaB2 gtf pmmB | - IMP dehydrogenase - glycosyltransferase - Mannose-1-phosphatase | attachment | ▪ Association with GDP-mannose and GPL biosynthesis ▪ Formation of biofilm | MAH A5 | [67] | |
| - Pcd | - hypothetical membrane protein - piperideine-6-carboxylic acid dehydrogenase | attachment | ▪ Association with biosynthesis of aminoadipic acid ▪ Formation of biofilm | MAH A5 | [67] | |
| LprB | - leucine-responsive regulatory protein B | cell wall | ▪ Bacterial cell surface protein ▪ Component of biofilm matrix ▪ Association with antibiotic resistance | MAH 104 | [78] | |
| MAVA5_03380 MAVA5_10375 | - FtsK/ SpoIIIE-like DNA translocation protein | transport | ▪ Export of eDNA ▪ Component of biofilm matrix ▪ Association with antibiotic resistance | MAH A5 | [81] | |
| MAVA5_19945 MAVA5_22765 | - carbonic anhydrase | -* | ▪ Export of eDNA in reseponse to bicarbonate ▪ Component of biofilm matrix | MAH A5 | [82] | |
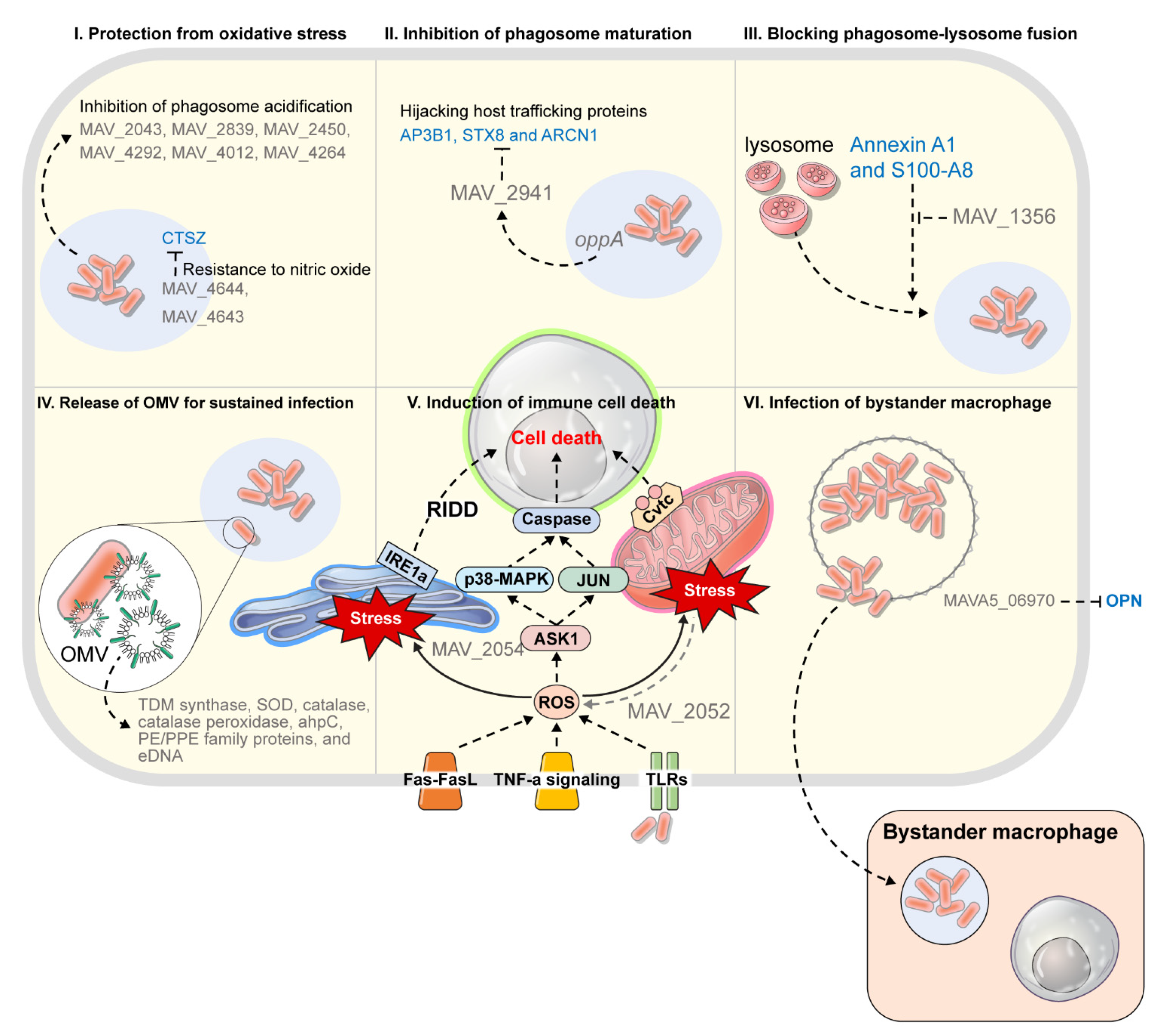
| Strategy 2. Resistance to the phagocytic environment in the immune cells | ||||||
| Oxidative stress/ phagosome acidification | MAV_2043 (Cu-Zn SOD) | - Cu-Zn superoxide dismutase | catalysis | ▪ Protection from oxidative stress | MAH 104 | [83] |
| MAV_2839 (ahpC) | - Alkyl hydroperoxide reductase | catalysis | ▪ Protection from oxidative stress | MAH 109 | [84] | |
| MAV_4682 (aceA) | - isocitrate lyase | metabolism | ▪ A key enzyme in glyoxylate shunt ▪ Use of fatty acids and acetates as basic carbon resources under carbon restricted conditions | MAH 109 | [84] | |
| MAV_2450 (pks12) | - Polyketide synthase 12 | cell wall | ▪ Association with susceptibility to oxidative products ▪ Inhibition of phagosome acidification | MAH 104 | [85] | |
| MAV_4292 | - Hypothetical protein | -* | ▪ Association with susceptibility to oxidative products (es, nitric oxide) ▪ Inhibition of phagosome acidification | MAH 104 | [85] | |
| MAV_4012 | - Conserved hypothetical protein | -* | ▪ Association with susceptibility to oxidative products ▪ Inhibition of phagosome acidification | MAH 104 | [85] | |
| MAV_4264 | - Hypothetical protein, homology with bacterial regulatory protein TetR domain | -* | ▪ Regulation of the genes that participate in the inhibition of phagosome acidification (ex, MAV_2450, MAV_4292, MAV_4012) ▪ Inhibition of phagosome acidification | MAH 104 | [85] | |
| MAV_4644 | - putative pore-forming protein that has ADP-ribosyltransferase (ADPRT) activity | interfering with host peptide | ▪ Binding to cathepsin Z ▪ Interfering with CTZ action that induces bacterial death in the presence of NO | MAH 104 | [86] | |
| Phagosome-lysosome fusion | MAV_1356 | - calmodulin-like protein | hijacking host protein | ▪ Hijacking to Annexin A1 and S100-A8 ▪ Regulating the phagocytic membrane ▪ Blocking phagosome-lysosome fusion | MAH 104 | [87] |
| MAV_2928 | - PPE25-MAV | secretion | ▪ Responsible for the Esx-5 region of the Type VII secretion system ▪ Export system for adjacent ESAT family gene, MAV_2921 ▪ Interfering with endosome maturation | MAH 109 | [88,89] | |
| MAV_2941 | - Hypothetical protein, a small protein only present in M. avium | hijacking host protein | ▪ Hijacking host trafficking proteins (ex, AP3B1, STX8 and ARCN1) ▪ Interfering with endosome maturation | MAH 104 | [90] | |
| oppA | - Oligopeptide transporter | transport | ▪ Active transport of oligopeptides and small protein (ex, MAV_2941) | MAH 104 | [91] | |
| Strategy 3. Resistance to antimicrobial peptide | ||||||
| Secretion of antimicrobial peptide | lysX | - lysyl-transferase-lysyl-tRNA synthetase | lysinylation | ▪ Association with GPL expression ▪ Resistance to human beta defensin-1 | MAH 104 | [92,93] |
| MAV_0216 | - Cutinase superfamily protein | -* | ▪ Resistance to antimicrobial peptide (polymyxin B) | MAH 104 | [94] | |
| MAV_3616 | - Long-chain specific acyl-CoA dehydrogenase | -* | ▪ Resistance to antimicrobial peptide (polymyxin B) | MAH 104 | [94] | |
| MAV_2450 | - Erythronolide synthase (polyketide synthase), modules 3 and 4 | -* | ▪ Resistance to antimicrobial peptide (polymyxin B) | MAH 104 | [94] | |
| Strategy 4. Induction of immune cell death and spreading tactics | ||||||
| Cell death | MAV_2052 | - putative cysteine synthase A protein | induction of cell death | ▪ Induction of cell death through TLR4-dependent ROS production and JNK pathway | MAH 104 | [95] |
| MAV_2054 (MMP-1) | - 35-kDa major membrane protein 1 | induction of cell death | ▪ Induction of cell death via ROS production and the mitochondrial pathway | MAH 104 | [96,97] | |
| MAVA5_06970 | - a secreted protein | induction of cell death | ▪ Hijacking OPN to hinder the operation of OPN ▪ Induction of cell death and limitation of the activation of the type I immunity pathway ▪ Enhancement of bystander macrophage | MAH A5 | [59] | |
3.2. Survival Strategy in Phagocytic Immune Cells: Survival Strategy within Phagosomal Environment in the Early Stage of MAH Infection
3.3. Hijacking Tactics of Host Trafficking Protein
3.4. Release of OMV for Sustained Infection
3.5. Genes Related to the Tolerance to Attacks of Host Antimicrobial Peptide
3.6. Induction of Immune Cell Death and Spreading Strategies
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kendall, B.A.; Winthrop, K.L. Update on the Epidemiology of Pulmonary Nontuberculous Mycobacterial Infections. Semin. Respir. Crit. Care Med. 2013, 34, 087–094. [Google Scholar] [CrossRef]
- Yoon, H.J.; Choi, H.Y.; Ki, M. Nontuberculosis mycobacterial infections at a specialized tuberculosis treatment centre in the Republic of Korea. BMC Infect. Dis. 2017, 17, 432. [Google Scholar] [CrossRef] [Green Version]
- Jeon, D. Infection Source and Epidemiology of Nontuberculous Mycobacterial Lung Disease. Tuberc. Respir. Dis. 2019, 82, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Weiss, C.H.; Glassroth, J. Pulmonary disease caused by nontuberculous mycobacteria. Expert Rev. Respir. Med. 2012, 6, 597–613. [Google Scholar] [CrossRef]
- Ko, R.E.; Moon, S.M.; Ahn, S.; Jhun, B.W.; Jeon, K.; Kwon, O.J.; Huh, H.J.; Ki, C.S.; Lee, N.Y.; Koh, W.J. Changing Epidemiology of Nontuberculous Mycobacterial Lung Diseases in a Tertiary Referral Hospital in Korea between 2001 and 2015. J. Korean Med. Sci. 2018, 33, e65. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-I.; Shin, S.J.; Shin, M.-K. Differential Genotyping of Mycobacterium avium Complex and Its Implications in Clinical and Environmental Epidemiology. Microorganisms 2020, 8, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolinsky, E. Nontuberculous mycobacteria and associated diseases. Am. Rev. Respir. Dis. 1979, 119, 107–159. [Google Scholar] [PubMed]
- Biet, F.; Boschiroli, M.L.; Guilloteau, L.A. Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC). Vet. Res. 2005, 36, 411–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignatov, D.; Kondratieva, E.; Azhikina, T.; Apt, A. Mycobacterium avium-triggered diseases: Pathogenomics. Cell. Microbiol. 2012, 14, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Runyon, E.H. Pathogenic mycobacteria. Bibl. Tuberc. 1965, 21, 235–287. [Google Scholar]
- Murcia, M.I.; Tortoli, E.; Menendez, M.C.; Palenque, E.; Garcia, M.J. Mycobacterium colombiense sp. nov., a novel member of the Mycobacterium avium complex and description of MAC-X as a new ITS genetic variant. Int. J. Syst. Evol. Microbiol. 2006, 56, 2049–2054. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.; García, I.G.; Aranaz, A.; Bezos, J.; Romero, B.; de Juan, L.; Mateos, A.; Gómez-Mampaso, E.; Domínguez, L. Genetic Diversity of Mycobacterium avium Isolates Recovered from Clinical Samples and from the Environment: Molecular Characterization for Diagnostic Purposes. J. Clin. Microbiol. 2008, 46, 1246–1251. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.S.; Pal, S.; Sarkar, I.; Roy, A.; das Mohapatra, P.K.; Sen, A. Comparative genomics of Mycobacterium reveals evolutionary trends of M. avium complex. Genomics 2019, 111, 426–435. [Google Scholar] [CrossRef]
- Tortoli, E.; Rindi, L.; Garcia, M.J.; Chiaradonna, P.; Dei, R.; Garzelli, C.; Kroppenstedt, R.M.; Lari, N.; Mattei, R.; Mariottini, A.; et al. Proposal to elevate the genetic variant MAC-A, included in the Mycobacterium avium complex, to species rank as Mycobacterium chimaera sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Bang, D.; Herlin, T.; Stegger, M.; Andersen, A.B.; Torkko, P.; Tortoli, E.; Thomsen, V.O. Mycobacterium arosiense sp. nov., a slowly growing, scotochromogenic species causing osteomyelitis in an immunocompromised child. Int. J. Syst. Evol. Microbiol. 2008, 58, 2398–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ingen, J.; Boeree, M.J.; Kosters, K.; Wieland, A.; Tortoli, E.; Dekhuijzen, P.N.R.; van Soolingen, D. Proposal to elevate Mycobacterium avium complex ITS sequevar MAC-Q to Mycobacterium vulneris sp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 2277–2282. [Google Scholar] [CrossRef] [Green Version]
- Ben-Salah, I.; Cayrou, C.; Raoult, D.; Drancourt, M. Mycobacterium marseillense sp. nov., Mycobacterium timonense sp. nov. and Mycobacterium bouchedurhonense sp. nov., members of the Mycobacterium avium complex. Int. J. Syst. Evol. Microbiol. 2009, 59, 2803–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Kim, B.J.; Kim, H.; Won, Y.S.; Jeon, C.O.; Jeong, J.; Lee, S.H.; Lim, J.H.; Lee, S.H.; Kim, C.K.; et al. Mycobacterium paraintracellulare sp. nov., for the genotype INT-1 of Mycobacterium intracellulare. Int. J. Syst. Evol. Microbiol. 2016, 66, 3132–3141. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-J.; Math, R.K.; Jeon, C.O.; Yu, H.-K.; Park, Y.-G.; Kook, Y.-H.; Kim, B.-J. Mycobacterium yongonense sp. nov., a slow-growing non-chromogenic species closely related to Mycobacterium intracellulare. Int. J. Syst. Evol. Microbiol. 2013, 63, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Talwar, G.P.; Ahmed, N.; Saini, V. The use of the name Mycobacterium w for the leprosy immunotherapeutic bacillus creates confusion with M. tuberculosis-W (Beijing strain): A suggestion. Infect. Genet. Evol. 2008, 8, 100–101. [Google Scholar] [CrossRef]
- Mijs, W.; de Haas, P.; Rossau, R.; van der Laan, T.; Rigouts, L.; Portaels, F.; van Soolingen, D. Molecular evidence to support a proposal to reserve the designation Mycobacterium avium subsp. avium for bird-type isolates and ’M. avium subsp. hominissuis’ for the human/porcine type of M. avium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1505–1518. [Google Scholar]
- Radomski, N.; Thibault, V.C.; Karoui, C.; de Cruz, K.; Cochard, T.; Gutiérrez, C.; Supply, P.; Biet, F.; Boschiroli, M.L. Determination of Genotypic Diversity of Mycobacterium avium Subspecies from Human and Animal Origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism Typing Methods. J. Clin. Microbiol. 2010, 48, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Ronai, Z.; Csivincsik, A.; Dan, A.; Gyuranecz, M. Molecular analysis and MIRU-VNTR typing of Mycobacterium avium subsp. avium, ’hominissuis’ and silvaticum strains of veterinary origin. Infect. Genet. Evol. 2016, 40, 192–199. [Google Scholar] [CrossRef]
- Turenne, C.Y.; Collins, D.M.; Alexander, D.C.; Behr, M.A. Mycobacterium avium subsp. paratuberculosis and M. avium subsp. avium Are Independently Evolved Pathogenic Clones of a Much Broader Group of M. avium Organisms. J. Bacteriol. 2008, 190, 2479–2487. [Google Scholar] [CrossRef] [Green Version]
- Saxegaard, F.; Baess, I. Relationship between Mycobacterium avium, Mycobacterium paratuberculosis and “wood pigeon mycobacteria”. Determinations by DNA-DNA hybridization. APMIS 1988, 96, 37–42. [Google Scholar] [CrossRef]
- Thorel, M.-F.; Krichevsky, M.; Lévy-Frébault, V.V. Numerical Taxonomy of Mycobactin-Dependent Mycobacteria, Emended Description of Mycobacterium avium, and Description of Mycobacterium avium subsp. avium subsp. nov., Mycobacterium avium subsp. paratuberculosis subsp. nov., and Mycobacterium avium subsp. silvaticum subsp. nov. Int. J. Syst. Bacteriol. 1990, 40, 254–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronai, Z.; Csivincsik, A.; Dan, A. Molecular identification of Mycobacterium avium subsp. silvaticum by duplex high-resolution melt analysis and subspecies-specific real-time PCR. J. Clin. Microbiol. 2015, 53, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Hasonova, L.; Pavlik, I. Economic impact of paratuberculosis in dairy cattle herds: A review. Veterinarni. Medicina. 2006, 51, 193–211. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.B.; Barletta, R.G. Mycobacterium avium subsp. paratuberculosis in Veterinary Medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, R.L.J.L.; Lana, R.; Díaz-Rubio, M. Mycobacterium avium subspecies paratuberculosis and its relationship with Crohn’s disease. World J. Gastroenterol. 2009, 15, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Turenne, C.Y.; Wallace, R.; Behr, M.A. Mycobacterium avium in the Postgenomic Era. Clin. Microbiol. Rev. 2007, 20, 205–229. [Google Scholar] [CrossRef] [Green Version]
- Coelho, A.C.; de Lurdes Pinto, M.; Matos, A.; Matos, M.; dos Anjos Pires, M. Mycobacterium avium complex in domestic and wild animals. In Insights from Veterinary Medicine; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar]
- Shah, N.M.; Davidson, J.A.; Anderson, L.F.; Lalor, M.K.; Kim, J.; Thomas, H.L.; Lipman, M.; Abubakar, I. Pulmonary Mycobacterium avium-intracellulare is the main driver of the rise in non-tuberculous mycobacteria incidence in England, Wales and Northern Ireland, 2007–2012. BMC Infect. Dis. 2016, 16, 195. [Google Scholar] [CrossRef] [Green Version]
- Cuttino, J.T.; Mc, C.A. Pure granulomatous nocardiosis, a new fungus disease distinguished by intracellular parasitism; a description of a new disease in man due to a hitherto undescribed organism, Nocardia intracellularis, n. sp., including a study of the biologic and pathogenic properties of this species. Am. J. Pathol. 1949, 25, 1–47. [Google Scholar]
- Thorel, M.; Huchzermeyer, H.; Michel, A. Mycobacterium avium and Mycobacterium intracellulare infection in mammals. Rev. Sci. Tech. l’OIE 2001, 20, 204–218. [Google Scholar] [CrossRef]
- Timms, V.J.; Hassan, K.A.; Mitchell, H.M.; Neilan, B.A. Comparative genomics between human and animal associated subspecies of the Mycobacterium avium complex: A basis for pathogenicity. BMC Genom. 2015, 16, 695. [Google Scholar] [CrossRef]
- MacKenzie, N.; Alexander, D.C.; Turenne, C.Y.; Behr, M.A.; de Buck, J.M. Genomic Comparison of PE and PPE Genes in the Mycobacterium avium Complex. J. Clin. Microbiol. 2009, 47, 1002–1011. [Google Scholar] [CrossRef] [Green Version]
- Turenne, C.Y.; Semret, M.; Cousins, D.V.; Collins, D.M.; Behr, M.A. Sequencing of hsp65 Distinguishes among Subsets of the Mycobacterium avium Complex. J. Clin. Microbiol. 2006, 44, 433–440. [Google Scholar] [CrossRef] [Green Version]
- Frothingham, R.; Wilson, K.H. Sequence-based differentiation of strains in the Mycobacterium avium complex. J. Bacteriol. 1993, 175, 2818–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunze, Z.M.; Wall, S.; Appelberg, R.; Silva, M.T.; Portaels, F.; McFadden, J.J. IS901, a new member of a widespread class of atypical insertion sequences, is associated with pathogenicity in Mycobacterium avium. Mol. Microbiol. 1991, 5, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, C.; Bernasconi, C.; Burki, D.; Bodmer, T.; Telenti, A. A novel insertion element from Mycobacterium avium, IS1245, is a specific target for analysis of strain relatedness. J. Clin. Microbiol. 1995, 33, 304–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frothingham, R.; Meeker-O’Connell, W.A. Genetic diversity in the Mycobacterium tuberculosis complex based on variable numbers of tandem DNA repeats. Microbiology 1998, 144, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannantine, J.P.; Zhang, Q.; Li, L.-L.; Kapur, V. Genomic homogeneity between Mycobacterium avium subsp. avium and Mycobacterium avium subsp. paratuberculosis belies their divergent growth rates. BMC Microbiol. 2003, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Uchiya, K.-I.; Takahashi, H.; Yagi, T.; Moriyama, M.; Inagaki, T.; Ichikawa, K.; Nakagawa, T.; Nikai, T.; Ogawa, K. Comparative Genome Analysis of Mycobacterium avium Revealed Genetic Diversity in Strains that Cause Pulmonary and Disseminated Disease. PLoS ONE 2013, 8, e71831. [Google Scholar] [CrossRef]
- Bannantine, J.P.; Conde, C.; Bayles, D.O.; Branger, M.; Biet, F. Genetic Diversity Among Mycobacterium avium Subspecies Revealed by Analysis of Complete Genome Sequences. Front. Microbiol. 2020, 11, 1701. [Google Scholar] [CrossRef]
- Uchiya, K.-I.; Tomida, S.; Nakagawa, T.; Asahi, S.; Nikai, T.; Ogawa, K. Comparative genome analyses of Mycobacterium avium reveal genomic features of its subspecies and strains that cause progression of pulmonary disease. Sci. Rep. 2017, 7, 39750. [Google Scholar] [CrossRef] [Green Version]
- Uchiya, K.-I.; Takahashi, H.; Nakagawa, T.; Yagi, T.; Moriyama, M.; Inagaki, T.; Ichikawa, K.; Nikai, T.; Ogawa, K. Characterization of a Novel Plasmid, pMAH135, from Mycobacterium avium Subsp. hominissuis. PLoS ONE 2015, 10, e0117797. [Google Scholar] [CrossRef] [Green Version]
- Arruda, S.; Bomfim, G.; Knights, R.; Huima-Byron, T.; Riley, L.W. Cloning of an M. tuberculosis DNA fragment associated with entry and survival inside cells. Science 1993, 261, 1454–1457. [Google Scholar] [CrossRef] [PubMed]
- Foley-Thomas, E.M.; Whipple, D.L.; Bermudez, L.E.; Barletta, R.G. Phage infection, transfection and transformation of Mycobacterium avium complex and Mycobacterium paratuberculosis. Microbiology 1995, 141, 1173–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Voss, J.J.; Rutter, K.-L.; Schroeder, B.G.; Su, H.; Zhu, Y.; Barry, C.E. The salicylate-derived mycobactin siderophores of Mycobacterium tuberculosis are essential for growth in macrophages. Proc. Natl. Acad. Sci. USA 2000, 97, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Moolji, J.; Dufort, A.; Staffa, A.; Domenech, P.; Reed, M.B.; Behr, M.A. Iron Acquisition in Mycobacterium avium subsp. paratuberculosis. J. Bacteriol. 2016, 198, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Bannantine, J.P.; Zhang, Q.; Amonsin, A.; May, B.J.; Alt, D.; Banerji, N.; Kanjilal, S.; Kapur, V. The complete genome sequence of Mycobacterium avium subspecies paratuberculosis. Proc. Natl. Acad. Sci. USA 2005, 102, 12344–12349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopinč, R.; Lapanje, A. Antibiotic susceptibility profile of Mycobacterium avium subspecies hominissuis is altered in low-iron conditions. J. Antimicrob. Chemother. 2012, 67, 2903–2907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffrey, B.; Rose, S.J.; Gilbert, K.; Lewis, M.; Bermudez, L.E. Comparative analysis of the genomes of clinical isolates of Mycobacterium avium subsp. hominissuis regarding virulence-related genes. J. Med. Microbiol. 2017, 66, 1063–1075. [Google Scholar] [CrossRef]
- Inderlied, C.B.; Kemper, C.A.; Bermudez, L.E. The Mycobacterium avium complex. Clin. Microbiol. Rev. 1993, 6, 266–310. [Google Scholar] [CrossRef] [PubMed]
- Early, J.; Fischer, K.; Bermudez, L.E. Mycobacterium avium uses apoptotic macrophages as tools for spreading. Microb. Pathog. 2011, 50, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez, L.E.; Danelishvili, L.; Babrack, L.; Pham, T. Evidence for genes associated with the ability of Mycobacterium avium subsp. hominissuis to escape apoptotic macrophages. Front. Cell. Infect. Microbiol. 2015, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, L.E. Differential mechanisms of intracellular killing of Mycobacterium avium and Listeria monocytogenes by activated human and murine macrophages. The role of nitric oxide. Clin. Exp. Immunol. 2008, 91, 277–281. [Google Scholar] [CrossRef]
- Danelishvili, L.; Rojony, R.; Carson, K.L.; Palmer, A.L.; Rose, S.J.; Bermudez, L.E. Mycobacterium avium subsp. hominissuis effector MAVA5_06970 promotes rapid apoptosis in secondary-infected macrophages during cell-to-cell spread. Virulence 2018, 9, 1287–1300. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Catanzaro, A.; Rao, S.P. Apoptosis of human monocytes and macrophages by Mycobacterium avium sonicate. Infect. Immun. 1997, 65, 5262–5271. [Google Scholar] [CrossRef] [Green Version]
- Babrak, L.; Danelishvili, L.; Rose, S.J.; Kornberg, T.; Bermudez, L.E. The Environment of “Mycobacterium avium subsp. hominissuis” Microaggregates Induces Synthesis of Small Proteins Associated with Efficient Infection of Respiratory Epithelial Cells. Infect. Immun. 2014, 83, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Aronson, T.; Holtzman, A.; Glover, N.; Boian, M.; Froman, S.; Berlin, O.G.W.; Hill, H.; Stelma, G. Comparison of Large Restriction Fragments of Mycobacterium avium Isolates Recovered from AIDS and Non-AIDS Patients with Those of Isolates from Potable Water. J. Clin. Microbiol. 1999, 37, 1008–1012. [Google Scholar] [CrossRef] [Green Version]
- Nishiuchi, Y.; Tamura, A.; Kitada, S.; Taguri, T.; Matsumoto, S.; Tateishi, Y.; Yoshimura, M.; Ozeki, Y.; Matsumura, N.; Ogura, H.; et al. Mycobacterium avium complex organisms predominantly colonize in the bathtub inlets of patients’ bathrooms. Jpn. J. Infect. Dis. 2009, 62, 182–186. [Google Scholar] [PubMed]
- Kitaura, H.; Ohara, N.; Naito, M.; Kobayashi, K.; Yamada, T. Fibronectin-binding proteins secreted by Mycobacterium avium. APMIS 2000, 108, 558–564. [Google Scholar] [CrossRef]
- Middleton, A.M.; Chadwick, M.V.; Nicholson, A.G.; Dewar, A.; Groger, R.K.; Brown, E.J.; Wilson, R. The role of Mycobacterium avium complex fibronectin attachment protein in adherence to the human respiratory mucosa. Mol. Microbiol. 2000, 38, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Schorey, J.S.; Holsti, M.A.; Ratliff, T.L.; Allen, P.M.; Brown, E.J. Characterization of the fibronectin-attachment protein of Mycobacterium avium reveals a fibronectin-binding motif conserved among mycobacteria. Mol. Microbiol. 1996, 21, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Danelishvili, L.; Wu, M.; Macnab, M.; Bermudez, L.E. Mycobacterium avium Genes Associated with the Ability to Form a Biofilm. Appl. Environ. Microbiol. 2006, 72, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Danelishvili, L.; Wu, M.; Hidaka, E.; Katsuyama, T.; Stang, B.; Petrofsky, M.; Bildfell, R.; Bermudez, L.E. The ability to form biofilm influences Mycobacterium avium invasion and translocation of bronchial epithelial cells. Cell. Microbiol. 2006, 8, 806–814. [Google Scholar] [CrossRef]
- Babrak, L.; Danelishvili, L.; Rose, S.J.; Bermudez, L.E. Microaggregate-associated protein involved in invasion of epithelial cells by Mycobacterium avium subsp. hominissuis. Virulence 2015, 6, 694–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogel, M.R.; Soni, P.N.; Troken, J.R.; Sitikov, A.; Trejo, H.E.; Ridge, K.M. Vimentin is sufficient and required for wound repair and remodeling in alveolar epithelial cells. FASEB J. 2011, 25, 3873–3883. [Google Scholar] [CrossRef] [Green Version]
- Reddy, V.M.; Kumar, B. Interaction of Mycobacterium avium Complex with Human Respiratory Epithelial Cells. J. Infect. Dis. 2000, 181, 1189–1193. [Google Scholar] [CrossRef]
- Dam, T.; Danelishvili, L.; Wu, M.; Bermudez, L.E. The fadD2 gene is required for efficient Mycobacterium avium invasion of mucosal epithelial cells. J. Infect. Dis. 2006, 193, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Harriff, M.J.; Danelishvili, L.; Wu, M.; Wilder, C.; McNamara, M.; Kent, M.L.; Bermudez, L.E. Mycobacterium avium Genes MAV_5138 and MAV_3679 Are Transcriptional Regulators That Play a Role in Invasion of Epithelial Cells, in Part by Their Regulation of CipA, a Putative Surface Protein Interacting with Host Cell Signaling Pathways. J. Bacteriol. 2008, 191, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Krzywinska, E.; Schorey, J.S. Characterization of genetic differences between Mycobacterium avium subsp. avium strains of diverse virulence with a focus on the glycopeptidolipid biosynthesis cluster. Veter. Microbiol. 2003, 91, 249–264. [Google Scholar] [CrossRef]
- Rose, S.J.; Bermudez, L.E. Mycobacterium avium Biofilm Attenuates Mononuclear Phagocyte Function by Triggering Hyperstimulation and Apoptosis during Early Infection. Infect. Immun. 2013, 82, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.; Wu, M.; Drummond, D.C.; Bermudez, L.E. Characterization of biofilm formation by clinical isolates of Mycobacterium avium. J. Med. Microbiol. 2003, 52, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Rojony, R.; Martin, M.; Campeau, A.; Wozniak, J.M.; Gonzalez, D.J.; Jaiswal, P.; Danelishvili, L.; Bermudez, L.E. Quantitative analysis of Mycobacterium avium subsp. hominissuis proteome in response to antibiotics and during exposure to different environmental conditions. Clin. Proteom. 2019, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNabe, M.; Tennant, R.; Danelishvili, L.; Young, L.; Bermudez, L.E. Mycobacterium avium ssp. hominissuis biofilm is composed of distinct phenotypes and influenced by the presence of antimicrobials. Clin. Microbiol. Infect. 2011, 17, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, G.; Young, L.S.; Bermudez, L.E. A Subinhibitory Concentration of Clarithromycin Inhibits Mycobacterium avium Biofilm Formation. Antimicrob. Agents Chemother. 2004, 48, 4907–4910. [Google Scholar] [CrossRef] [Green Version]
- Rose, S.J.; Babrak, L.M.; Bermudez, L.E. Mycobacterium avium Possesses Extracellular DNA that Contributes to Biofilm Formation, Structural Integrity, and Tolerance to Antibiotics. PLoS ONE 2015, 10, e0128772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.J.; Bermudez, L.E. Identification of Bicarbonate as a Trigger and Genes Involved with Extracellular DNA Export in Mycobacterial Biofilms. mBio 2016, 7, e01597-16. [Google Scholar] [CrossRef] [Green Version]
- McNamara, M.; Tzeng, S.-C.; Maier, C.; Wu, M.; Bermudez, L.E. Surface-exposed proteins of pathogenic mycobacteria and the role of cu-zn superoxide dismutase in macrophages and neutrophil survival. Proteome Sci. 2013, 11, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, M.; Tzeng, S.-C.; Maier, C.S.; Zhang, L.; Bermudez, L.E. Surface Proteome of “Mycobacterium avium subsp. hominissuis” during the Early Stages of Macrophage Infection. Infect. Immun. 2012, 80, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-J.; Danelishvili, L.; Wagner, D.; Petrofsky, M.; Bermudez, L.E. Identification of virulence determinants of Mycobacterium avium that impact on the ability to resist host killing mechanisms. J. Med. Microbiol. 2010, 59, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.S.; Danelishvili, L.; Rose, S.J.; Bermudez, L.E. MAV_4644 Interaction with the Host Cathepsin Z Protects Mycobacterium avium subsp. hominissuis from Rapid Macrophage Killing. Microorganisms 2019, 7, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinison, J.J.; Danelishvili, L.; Gupta, R.; Rose, S.J.; Babrak, L.M.; Bermudez, L.E. Identification of Mycobacterium avium subsp. hominissuis secreted proteins using an in vitro system mimicking the phagosomal environment. BMC Microbiol. 2016, 16, 270. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Miltner, E.; Wu, M.; Petrofsky, M.; Bermudez, L.E. A Mycobacterium avium PPE gene is associated with the ability of the bacterium to grow in macrophages and virulence in mice. Cell. Microbiol. 2004, 7, 539–548. [Google Scholar] [CrossRef]
- McNamara, M.; Danelishvili, L.; Bermudez, L.E. The Mycobacterium avium ESX-5 PPE protein, PPE25-MAV, interacts with an ESAT-6 family Protein, MAV_2921, and localizes to the bacterial surface. Microb. Pathog. 2012, 52, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danelishvili, L.; Bermudez, L.E. Mycobacterium avium MAV_2941 mimics phosphoinositol-3-kinase to interfere with macrophage phagosome maturation. Microbes Infect. 2015, 17, 628–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danelishvili, L.; Stang, B.; Bermudez, L.E. Identification of Mycobacterium avium genes expressed during in vivo infection and the role of the oligopeptide transporter OppA in virulence. Microb. Pathog. 2014, 76, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Kirubakar, G.; Murugaiyan, J.; Schaudinn, C.; Dematheis, F.; Holland, G.; Eravci, M.; Weise, C.; Roesler, U.; Lewin, A. Proteome Analysis of a M. avium Mutant Exposes a Novel Role of the Bifunctional Protein LysX in the Regulation of Metabolic Activity. J. Infect. Dis. 2018, 218, 291–299. [Google Scholar] [CrossRef]
- Kirubakar, G.; Schäfer, H.; Rickerts, V.; Schwarz, C.; Lewin, A. Mutation on lysX from Mycobacterium avium hominissuis impacts the host–pathogen interaction and virulence phenotype. Virulence 2020, 11, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Motamedi, N.; Danelishvili, L.; Bermudez, L.E. Identification of Mycobacterium avium genes associated with resistance to host antimicrobial peptides. J. Med. Microbiol. 2014, 63, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-I.; Choi, H.-G.; Son, Y.-J.; Whang, J.; Kim, K.; Jeon, H.S.; Park, H.-S.; Back, Y.W.; Choi, S.; Kim, S.-W.; et al. Mycobacterium avium MAV2052 protein induces apoptosis in murine macrophage cells through Toll-like receptor 4. Apoptosis 2016, 21, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-I.; Whang, J.; Choi, H.-G.; Son, Y.-J.; Jeon, H.S.; Back, Y.W.; Park, H.-S.; Paik, S.; Park, J.-K.; Choi, C.H.; et al. Mycobacterium avium MAV2054 protein induces macrophage apoptosis by targeting mitochondria and reduces intracellular bacterial growth. Sci. Rep. 2016, 6, 37804. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.-R.; Lee, K.-S.; Lee, K.I.; Shim, T.S.; Koh, W.-J.; Jeon, H.S.; Son, Y.-J.; Shin, S.-J.; Kim, H.-J. Serodiagnostic Potential of Mycobacterium avium MAV2054 and MAV5183 Proteins. Clin. Vaccine Immunol. 2012, 20, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Ojha, A.K.; Baughn, A.D.; Sambandan, D.; Hsu, T.; Trivelli, X.; Guerardel, Y.; Alahari, A.; Kremer, L.; Jacobs, W.R., Jr.; Hatfull, G.F. Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria. Mol. Microbiol. 2008, 69, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Freeman, R.; Geier, H.; Weigel, K.M.; Do, J.; Ford, T.E.; Cangelosi, G.A. Roles for Cell Wall Glycopeptidolipid in Surface Adherence and Planktonic Dispersal of Mycobacterium avium. Appl. Environ. Microbiol. 2006, 72, 7554–7558. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-W.; Schmoller, S.K.; Bannantine, J.P.; Eckstein, T.M.; Inamine, J.M.; Livesey, M.; Albrecht, R.; Talaat, A.M. A novel cell wall lipopeptide is important for biofilm formation and pathogenicity of Mycobacterium avium subspecies paratuberculosis. Microb. Pathog. 2009, 46, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Aung, T.T.; Yam, J.K.H.; Lin, S.; Salleh, S.M.; Givskov, M.; Liu, S.; Lwin, N.C.; Yang, L.; Beuerman, R.W. Biofilms of Pathogenic Nontuberculous Mycobacteria Targeted by New Therapeutic Approaches. Antimicrob. Agents Chemother. 2016, 60, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, N.B.; Oliveira, F.S.; Duraes, F.V.; de Almeida, L.A.; Florido, M.; Prata, L.O.; Caliari, M.V.; Appelberg, R.; Oliveira, S.C. Toll-like receptor 9 is required for full host resistance to Mycobacterium avium infection but plays no role in induction of Th1 responses. Infect. Immun. 2011, 79, 1638–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, S.S.; Danelishvili, L.; Wagner, D.; Maser, J.; Li, Y.-J.; Morić, I.; Vogt, S.; Yamazaki, Y.; Lai, B.; E Bermudez, L. Virulence-related Mycobacterium avium subsp. hominissuis MAV_2928 gene is associated with vacuole remodeling in macrophages. BMC Microbiol. 2010, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Vergne, I.; Chua, J.; Singh, S.B.; Deretic, V. Cell biology of Mycobacterium tuberculosis phagosome. Annu Rev. Cell Dev. Biol. 2004, 20, 367–394. [Google Scholar] [CrossRef]
- Sharma, V.; Sharma, S.; Zu-Bentrup, K.H.; McKinney, J.D.; Russell, D.G.; Jacobs, W.R.; Sacchettini, J.C. Structure of isocitrate lyase, a persistence factor of Mycobacterium tuberculosis. Nat. Genet. 2000, 7, 663–668. [Google Scholar] [CrossRef]
- Honer Zu Bentrup, K.; Miczak, A.; Swenson, D.L.; Russell, D.G. Characterization of activity and expression of isocitrate lyase in Mycobacterium avium and Mycobacterium tuberculosis. J. Bacteriol. 1999, 181, 7161–7167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrofsky, M.; Bermudez, L.E. Neutrophils from Mycobacterium avium-infected mice produce TNF-alpha, IL-12, and IL-1 beta and have a putative role in early host response. Clin. Immunol. 1999, 91, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Master, S.S.; Springer, B.; Sander, P.; Boettger, E.C.; Deretic, V.; Timmins, G.S. Oxidative stress response genes in Mycobacterium tuberculosis: Role of ahpC in resistance to peroxynitrite and stage-specific survival in macrophages. Microbiology 2002, 148, 3139–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontán, P.; Aris, V.; Ghanny, S.; Soteropoulos, P.; Smith, I. Global Transcriptional Profile of Mycobacterium tuberculosis during THP-1 Human Macrophage Infection. Infect. Immun. 2007, 76, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Malik, Z.A.; Iyer, S.S.; Kusner, D.J. Mycobacterium tuberculosis Phagosomes Exhibit Altered Calmodulin-Dependent Signal Transduction: Contribution to Inhibition of Phagosome-Lysosome Fusion and Intracellular Survival in Human Macrophages. J. Immunol. 2001, 166, 3392–3401. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.M.; Weiss, D.G.; Gerke, V.; Kuznetsov, S.A.; Ahmad, S.F. Annexin A1 is a new functional linker between actin filaments and phagosomes during phagocytosis. J. Cell Sci. 2011, 124, 578–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Pittius, N.C.G.; Sampson, S.L.; Lee, H.; Kim, Y.; van Helden, P.D.; Warren, R.M. Evolution and expansion of the Mycobacterium tuberculosis PE and PPE multigene families and their association with the duplication of the ESAT-6 (esx) gene cluster regions. BMC Evol. Biol. 2006, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Guinn, K.M.; Hickey, M.J.; Mathur, S.K.; Zakel, K.L.; Grotzke, J.E.; Lewinsohn, D.M.; Smith, S.; Sherman, D.R. Individual RD1-region genes are required for export of ESAT-6/CFP-10 and for virulence of Mycobacterium tuberculosis. Mol. Microbiol. 2004, 51, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, I.; Velasco, G.; Pendás, A.M.; Fueyo, A.; López-Otín, C. Cathepsin Z, a Novel Human Cysteine Proteinase with a Short Propeptide Domain and a Unique Chromosomal Location. J. Biol. Chem. 1998, 273, 16816–16823. [Google Scholar] [CrossRef] [Green Version]
- Chiplunkar, S.S.; Silva, C.A.; Bermudez, L.E.; Danelishvili, L. Characterization of membrane vesicles released by Mycobacterium avium in response to environment mimicking the macrophage phagosome. Future Microbiol. 2019, 14, 293–313. [Google Scholar] [CrossRef] [Green Version]
- Kalscheuer, R.; Weinrick, B.; Veeraraghavan, U.; Besra, G.S.; Jacobs, W.R., Jr. Trehalose-recycling ABC transporter LpqY-SugA-SugB-SugC is essential for virulence of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 107, 21761–21766. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-M.; Jo, E.-K. Antimicrobial Peptides in Innate Immunity against Mycobacteria. Immune Netw. 2011, 11, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Van Der Does, A.M.; Bergman, P.; Agerberth, B.; Lindbom, L. Induction of the human cathelicidin LL-37 as a novel treatment against bacterial infections. J. Leukoc. Biol. 2012, 92, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Sturgill-Koszycki, S.; Schaible, U.E.; Russell, D.G. Mycobacterium-containing phagosomes are accessible to early endosomes and reflect a transitional state in normal phagosome biogenesis. EMBO J. 1996, 15, 6960–6968. [Google Scholar] [CrossRef]
- Bermudez, L.E.M.; Young, L.S.; Gupta, S. 1,25 Dihydroxyvitamin D3-dependent inhibition of growth or killing of Mycobacterium avium complex in human macrophages is mediated by TNF and GM-CSF. Cell. Immunol. 1990, 127, 432–441. [Google Scholar] [CrossRef]
- Schorey, J.S.; Sweet, L. The mycobacterial glycopeptidolipids: Structure, function, and their role in pathogenesis. Glycobiology 2008, 18, 832–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, E.; Stankowska, D.; Zhang, J.; Fol, M.; Cheng, Q.-J.; Lun, S.; Bishai, W.R.; Rajagopalan, M.; Chatterjee, D.; Madiraju, M.V. The Two-Domain LysX Protein of Mycobacterium tuberculosis Is Required for Production of Lysinylated Phosphatidylglycerol and Resistance to Cationic Antimicrobial Peptides. PLoS Pathog. 2009, 5, e1000534. [Google Scholar] [CrossRef]
- Fratazzi, C.; Arbeit, R.D.; Carini, C.; Remold, H.G. Programmed cell death of Mycobacterium avium serovar 4-infected human macrophages prevents the mycobacteria from spreading and induces mycobacterial growth inhibition by freshly added, uninfected macrophages. J. Immunol. 1997, 158, 4320–4327. [Google Scholar]
- Pais, T.F.; Appelberg, R. Induction of Mycobacterium avium growth restriction and inhibition of phagosome-endosome interactions during macrophage activation and apoptosis induction by picolinic acid plus IFNgamma. Microbiology 2004, 150, 1507–1518. [Google Scholar] [CrossRef] [Green Version]
- Briken, V.; Miller, J.L. Living on the edge: Inhibition of host cell apoptosis by Mycobacterium tuberculosis. Futur. Microbiol. 2008, 3, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Kabara, E.; Coussens, P.M. Infection of Primary Bovine Macrophages with Mycobacterium avium Subspecies paratuberculosis Suppresses Host Cell Apoptosis. Front. Microbiol. 2012, 3, 215. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, L.E.; Parker, A.; Petrofsky, M. Apoptosis of Mycobacterium avium-infected macrophages is mediated by both tumour necrosis factor (TNF) and Fas, and involves the activation of caspases. Clin. Exp. Immunol. 1999, 116, 94–99. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Pathak, S.; Basak, C.; Law, S.; Kundu, M.; Basu, J. Execution of Macrophage Apoptosis by Mycobacterium avium through Apoptosis Signal-regulating Kinase 1/p38 Mitogen-activated Protein Kinase Signaling and Caspase 8 Activation. J. Biol. Chem. 2003, 278, 26517–26525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, D.; Lee, J.; Choi, J.; Cho, S.; Kim, S.; Son, S.; Song, C. Reactive oxygen species-mediated endoplasmic reticulum stress response induces apoptosis of Mycobacterium avium-infected macrophages by activating regulated IRE1-dependent decay pathway. Cell. Microbiol. 2019, 21, e13094. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, L.E.; Petrofsky, M.; Sangari, F.J. Intracellular phenotype of Mycobacterium avium enters macrophages primarily by a macropinocytosis-like mechanism and survives in a compartment that differs from that with extracellular phenotype. Cell Biol. Int. 2004, 28, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, L.E.; Parker, A.; Goodman, J.R. Growth within macrophages increases the efficiency of Mycobacterium avium in invading other macrophages by a complement receptor-independent pathway. Infect. Immun. 1997, 65, 1916–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nau, G.J.; Chupp, G.L.; Emile, J.-F.; Jouanguy, E.; Berman, J.S.; Casanova, J.-L.; Young, R.A. Osteopontin Expression Correlates with Clinical Outcome in Patients with Mycobacterial Infection. Am. J. Pathol. 2000, 157, 37–42. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, M.-K.; Shin, S.J. Genetic Involvement of Mycobacterium avium Complex in the Regulation and Manipulation of Innate Immune Functions of Host Cells. Int. J. Mol. Sci. 2021, 22, 3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063011
Shin M-K, Shin SJ. Genetic Involvement of Mycobacterium avium Complex in the Regulation and Manipulation of Innate Immune Functions of Host Cells. International Journal of Molecular Sciences. 2021; 22(6):3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063011
Chicago/Turabian StyleShin, Min-Kyoung, and Sung Jae Shin. 2021. "Genetic Involvement of Mycobacterium avium Complex in the Regulation and Manipulation of Innate Immune Functions of Host Cells" International Journal of Molecular Sciences 22, no. 6: 3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063011