
Engineering a Novel Bivalent Oral Vaccine against Enteric Fever



Prokarium Ltd., London Bioscience Innovation Centre, 2 Royal College Street, London NW1 0NH, UK
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(6), 3287; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063287
Submission received: 27 February 2021
/
Revised: 18 March 2021
/
Accepted: 21 March 2021
/
Published: 23 March 2021
(This article belongs to the Special Issue Antimicrobial Resistance, Molecular Mechanisms and Fight Strategies)
Abstract
:Enteric fever is a major global healthcare issue caused largely by Salmonella enterica serovars Typhi and Paratyphi A. The objective of this study was to develop a novel, bivalent oral vaccine capable of protecting against both serovars. Our approach centred on genetically engineering the attenuated S. Typhi ZH9 strain, which has an excellent safety record in clinical trials, to introduce two S. Paratyphi A immunogenic elements: flagellin H:a and lipopolysaccharide (LPS) O:2. We first replaced the native S. Typhi fliC gene encoding flagellin with the highly homologous fliC gene from S. Paratyphi A using Xer-cise technology. Next, we replaced the S. Typhi rfbE gene encoding tyvelose epimerase with a spacer sequence to enable the sustained expression of O:2 LPS and prevent its conversion to O:9 through tyvelose epimerase activity. The resulting new strain, ZH9PA, incorporated these two genetic changes and exhibited comparable growth kinetics to the parental ZH9 strain. A formulation containing both ZH9 and ZH9PA strains together constitutes a new bivalent vaccine candidate that targets both S. Typhi and S. Paratyphi A antigens to address a major global healthcare gap for enteric fever prophylaxis. This vaccine is now being tested in a Phase I clinical trial (NCT04349553).
1. Introduction
Enteric fever is an infectious disease that causes significant mortality and morbidity in low- and middle-income countries (LMICs), such as those in South Asia and sub-Saharan Africa [1,2,3]. Enteric fever is predominantly caused by two serotypes of Salmonella enterica: S. Typhi, which is responsible for around 10.9 million cases per year, and S. Paratyphi A, which is responsible for around 3.4 million cases per year, based on 2017 data [2].
The widespread use of antibiotics in LMICs has led to the emergence of extensively drug-resistant (XDR) Salmonella strains that are no longer susceptible to multiple lines of antibiotics. For example, in a recent outbreak across three sites in Pakistan, over 90% of both S. Typhi and S. Paratyphi A isolates were resistant to fluoroquinolone [4]. Consequently, the World Health Organization (WHO) has listed fluoroquinolone-resistant Salmonellae as high priority pathogens for the research and development of new antibiotics [5].
Several vaccines are currently licensed for S. Typhi, including a live attenuated oral vaccine (Vivotif™) and parenteral injectable vaccines [6]. However, these do not protect against the S. Paratyphi A strain [7], the prevalence of which is increasing [8]. For example, two hospital-based studies in Nigeria found that a considerable proportion of enteric fever cases (ranging from 17 to 34%) were attributable to S. Paratyphi A [9,10]. Furthermore, several sources agree that enteric fever—in particular, that caused by S. Paratyphi A—is almost certainly under-reported in LMICs [11,12]. Therefore, the development of a bivalent vaccine that protects against both S. Typhi and S. Paratyphi A would address a serious global healthcare need.
Using a live attenuated orally administered vaccine has several potential advantages over injectable subunit Vi vaccines for protecting against enteric fever. These include a longer duration of protection, the establishment of immunological memory and a reactive immune profile that more closely follows that of natural infection [13]. Several groups have constructed live attenuated S. Paratyphi A vaccines. For example, an oral dose of S. Paratyphi A (ΔphoPQ) was well tolerated in rabbits [14], and an oral dose of S. Paratyphi A CVD 1902 (ΔguaBA, ΔclpX) was immunogenic in healthy volunteers [15]. However, these attenuated S. Paratyphi A strains have very limited safety data available in comparison with attenuated S. Typhi strains and do not offer simultaneous protection against both causative agents of enteric fever.
We have taken the approach of engineering a pre-existing oral attenuated Salmonella vaccine technology to enable its rapid adaptation against new disease targets. In clinical formulations, this vaccine is simple to manufacture and is extremely cost-effective. The parental vaccine strain, S. Typhi ZH9, has previously been shown to be safe in multiple clinical trials across a wide range of participant populations, including children living in an endemic LMIC [16,17,18,19,20,21]. In this study, we describe the process of engineering a new S. Typhi strain expressing two S. Paratyphi A antigens (LPS O:2 and H:a flagellin) that can be formulated in equivalent amounts with the S. Typhi ZH9 parental strain to create a bivalent vaccine designed to provide protection against both S. Typhi and S. Paratyphi A. We present data describing the genetic engineering, in vitro characterisation and in vivo immunogenicity of the basic formulation of a new bivalent enteric fever vaccine named Entervax™, which is now in clinical testing (NCT04349553) [22].
2. Results
2.1. Converting Flagellin from H:d to H:a
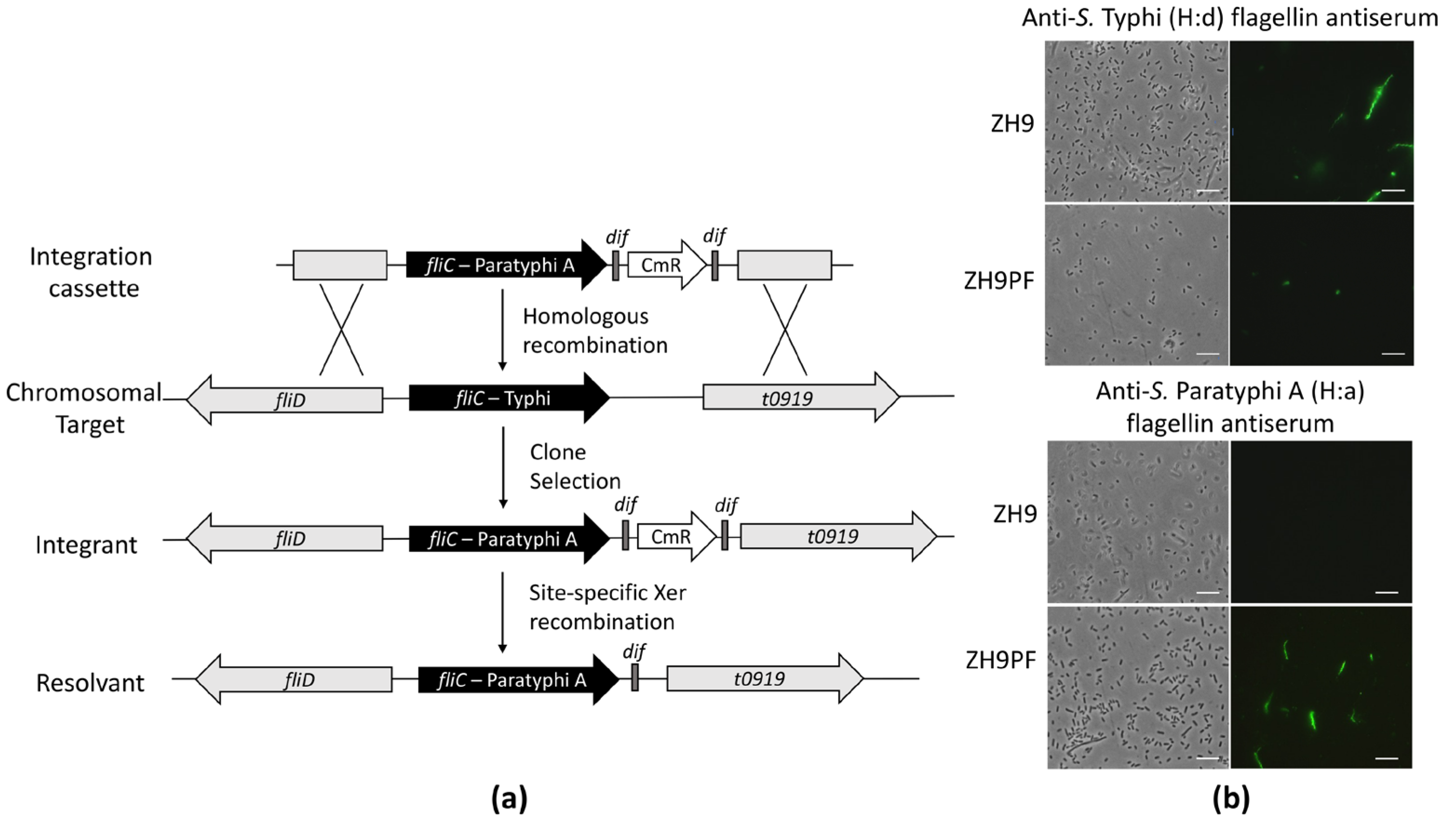
The construction of the parental S. Typhi ZH9 has previously been described, and it represents an attenuated strain with ΔaroC and ΔssaV mutations [17]. To begin constructing a hybrid S. Typhi strain expressing two immunogenic elements from S. Paratyphi A, we first converted the parental ZH9 H:d serotype to a H:a serotype by replacing the native fliC gene encoding flagellin with the S. Paratyphi A fliC gene (Figure 1a; see the methods section for more information).
The successful conversion of the H:d flagellin serotype to H:a in ZH9PF was tested by immunostaining. Bacteria were incubated with H:d antiserum or H:a antiserum plus fluorescent secondary antibodies, followed by visualisation with fluorescence microscopy. The parental ZH9 strain was not reactive against anti-S. Typhi (H:a) flagellin antiserum, while the modified ZH9PF strain showed positive staining (Figure 1b). Conversely, the parental ZH9 strain was reactive against anti-S. Typhi (H:d) flagellin antiserum, while the modified ZH9PF strain was minimally reactive (Figure 1b). The minimal fluorescence observed after staining ZH9PF with H:d flagellin antiserum appeared to be restricted to the membrane, suggesting that this was due to the polyclonal H:d serum recognising surface proteins in addition to its designated target, flagellin. Overall, it was clear that swapping the fliC gene successfully converted the flagellin expressed by ZH9 from an H:d to H:a serotype.
2.2. Modifying LPS from O:9 to O:2
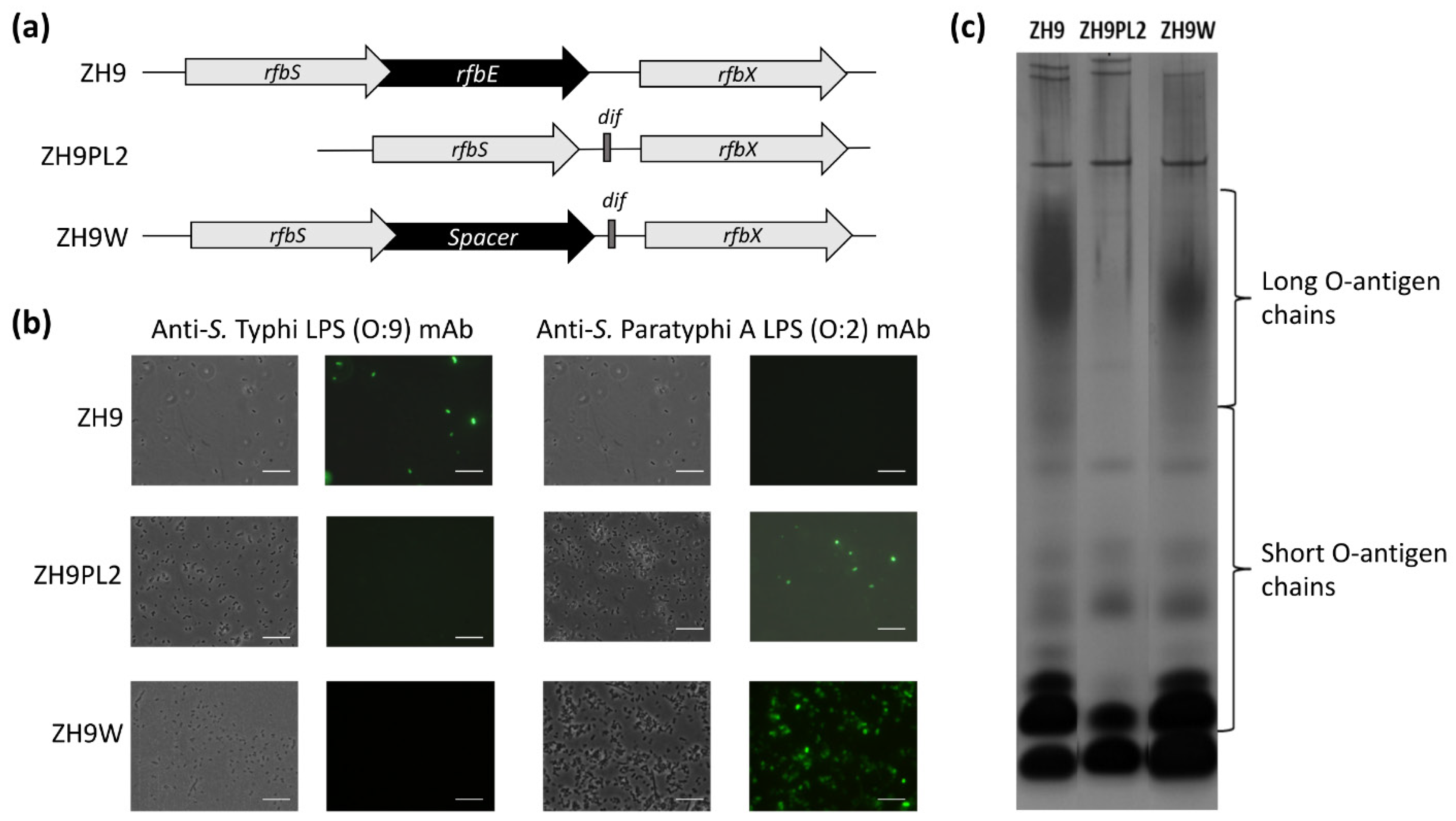
Using the parental ZH9 strain, we next tested two different approaches to convert the native S. Typhi LPS O:9 antigen to the S. Paratyphi A O:2 antigen. The CDP-d-tyvelose 2-epimerase (Tyv) of S. Typhi, encoded by the rfbE gene, catalyses the isomeration of 3,6-dideoxyhexose sugars CDP-paratose to CDP-tyvelose, which is incorporated in the O-antigen, thus conferring serogroup specificity O:9. In S. Paratyphi A, the rfbE gene is mutated, so paratose is incorporated in the O-antigen instead, thus conferring serogroup specificity O:2 [24]. In our first approach, the majority of the rfbE gene was simply deleted; in our second approach, the rfbE gene was fully replaced with the spacer gene, wbdR, which encodes a putative N-acetyltransferase gene that is not functional in Salmonella (Figure 2a). Both approaches followed a similar process for generating a chromosomal integration cassette, as described above for switching fliC genes.
In the first approach to delete the majority of the rfbE gene, the dif-flanked antibiotic resistance gene, cat, was amplified with primers rfbEdelF and rfbEdelR, which comprised homologous sequences to rfbS and rfbX genes, respectively, to generate ZH9PL2. In the second approach to fully replace rfbE with the wbdR spacer cistron, we synthesised the full wdbR spacer cistron, flanked on one side by approximately 700 bp of DNA homologous to the S. Typhi rfbE upstream gene, rfbS, and on the other side, by approximately 700 bp of DNA homologous to the S. Typhi downstream gene, rfbX, to create a deletion cassette and generate ZH9W (Figure 2a).
The conversion of the O:9 to the O:2 serotype of LPS in both ZH9PL2 and ZH9W was tested by direct staining of bacteria with anti-S. Typhi LPS (O:9) or anti-S. Paratyphi A LPS (O:2) monoclonal antibodies (mAbs), followed by visualisation using fluorescence microscopy. Both the ZH9PL2 and ZH9W strains were reactive against the O:2 mAb but not the O:9 mAb, suggesting that both approaches had successfully converted LPS from O:9 to O:2 (Figure 2b). The parental ZH9 strain was reactive against the O:9 mAb but not the O:2 mAb, as expected (Figure 2b). However, SDS-PAGE silver-stain analysis of ZH9PL2 and ZH9W LPS extracts highlighted that only when the rfbE gene had been replaced by the spacer gene, wbdR, was LPS expressed with long O-antigen chains (Figure 2c; Figure S1). Thus, we concluded that the second approach—to replace rfbE with a wbdR spacer gene—generated a strain that was functionally the closest match to the parental strain.
2.3. Constructing the Final New Strain, ZH9PA
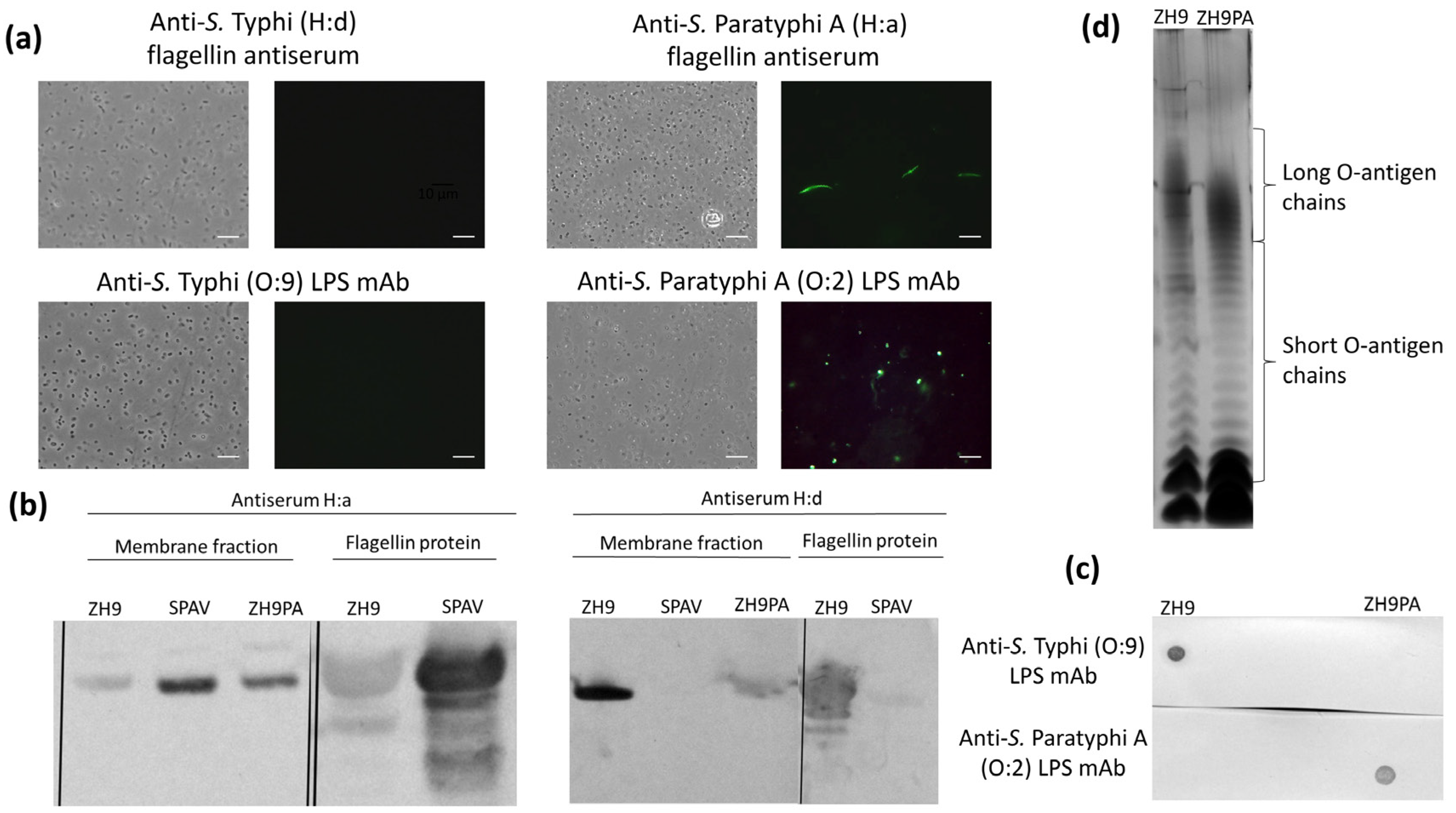
To generate the final ZH9PA strain incorporating both the H:d-to-H:a flagellin and O:9-to-O:2 LPS modifications, the ZH9PF strain (containing the fliC gene replacement) was further modified to replace the rfbE gene with the spacer gene, wbdR. These modifications of the LPS and flagellin loci were verified by sequencing (data not shown), and the expression of LPS and flagellin was analysed using immunostaining followed by fluorescence microscopy, western blot and silver staining techniques.
Based on immunostaining, ZH9PA was reactive against both S. Paratyphi A LPS (O:2) and flagellin (H:a) targets but not S. Typhi LPS (O:9) or flagellin (H:d) (Figure 3a). Similarly, by western blot, the bacterial membrane fractions from ZH9PA were positive for S. Paratyphi A flagellin H:a but only weakly positive for S. Typhi flagellin H:d (Figure 3b; Figure S2). Heat-inactivated bacterial preparations from ZH9PA were also positive for LPS O:2 (but not LPS O:9) by dot blot (Figure 3c; Figure S3). Finally, silver-stained SDS-PAGE analysis confirmed long O-chain expression in ZH9PA, as well as in the ZH9 parent vaccine strain (Figure 3d).
2.4. Evaluating the Growth of ZH9PA
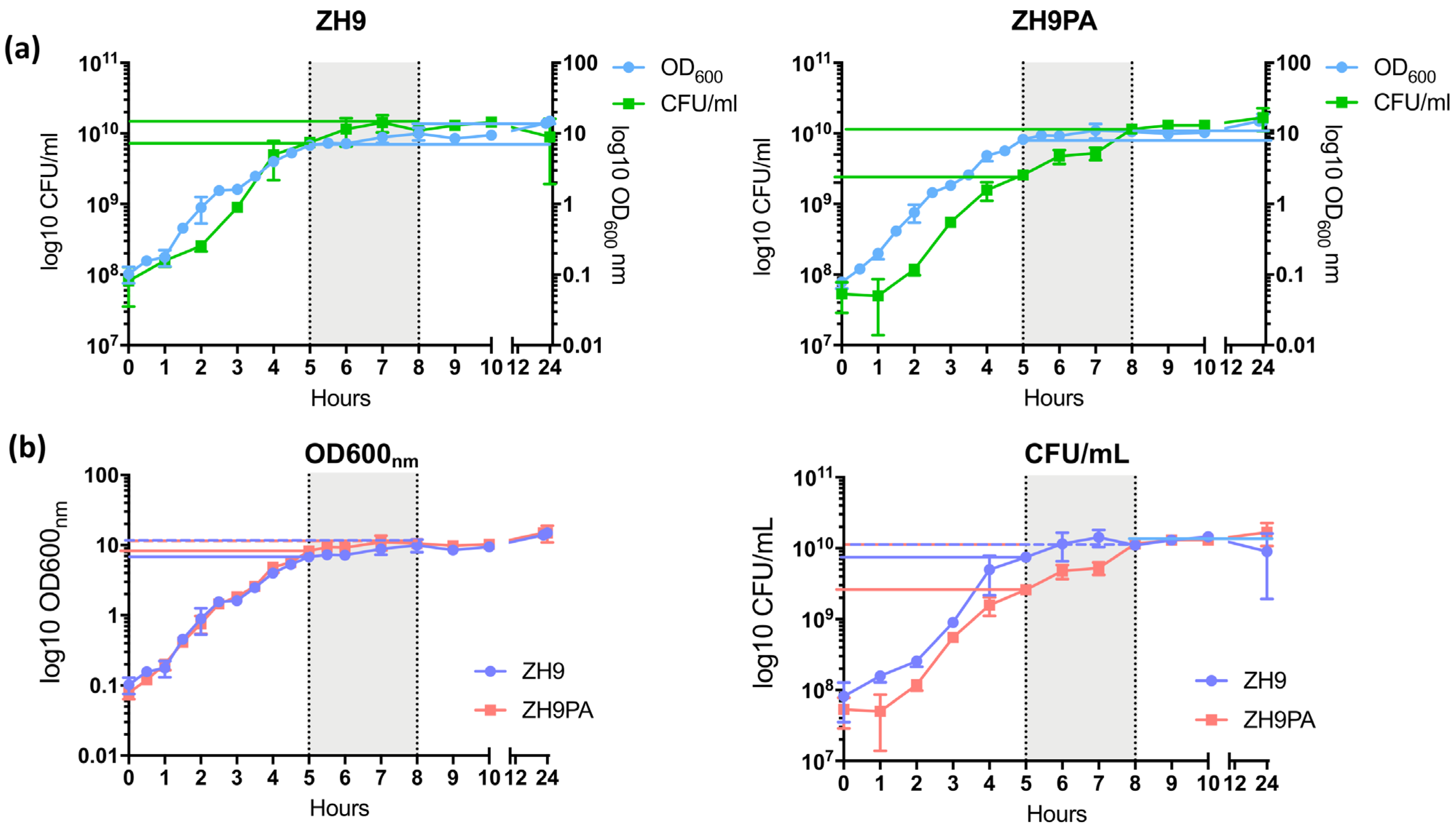
We tested whether the introduction of the S. Paratyphi A flagellin (H:a) and the replacement of S. Typhi rfbE with a spacer gene had an impact on ZH9PA bacterial growth in culture compared to the parental ZH9 strain. For both strains, an OD600 nm = 0.1 was seeded into liquid culture and allowed to grow for 24 h. At regular intervals, samples were taken, and counts were made using both optical density (OD600 nm) and colony-forming unit (CFU/mL) measurements.
We first compared OD600 nm vs. CFU/mL within each individual bacterial strain. The patterns of growth observed over time using both methods suggested that ZH9PA had a slight growth delay together with a longer lag phase compared to the parental ZH9 strain; although by 8 h (the end of the late exponential growth phase, defined as between 5 to 8 h of culture), both strains achieved similar colony counts (1.1 × 1010 vs. 1.15 × 1010 CFU/mL for ZH9 vs. ZH9PA, respectively) (Figure 4a).
Next, we compared OD600 nm or CFU/mL between the two strains (ZH9 vs. ZH9PA). In the late exponential growth phase, the optical density at 600 nm was similar for both strains. This ranged from 7 to 10 for the parental ZH9 strain and from 8 to 11 for the modified ZH9PA strain (Figure 4b). Colony counts within the late exponential growth phase were generally higher for the parental ZH9 strain, ranging from 7.5 × 109 to 1.43 × 1010 for ZH9 and from 2.6 × 109 to 1.15 × 1010 for ZH9PA strain (Figure 4b). No statistically significant differences were observed between the two strains based on either approximate bacterial cell counts (optical density measurements) or absolute bacterial cell counts (CFU/mL) at any timepoint (Figure 4b). Overall, we concluded that the two modifications present in strain ZH9PA had a negligible impact on strain fitness compared to the parental ZH9 strain.
2.5. Evaluating the Immunogenicity of the Basic Bivalent Enteric Fever Vaccine
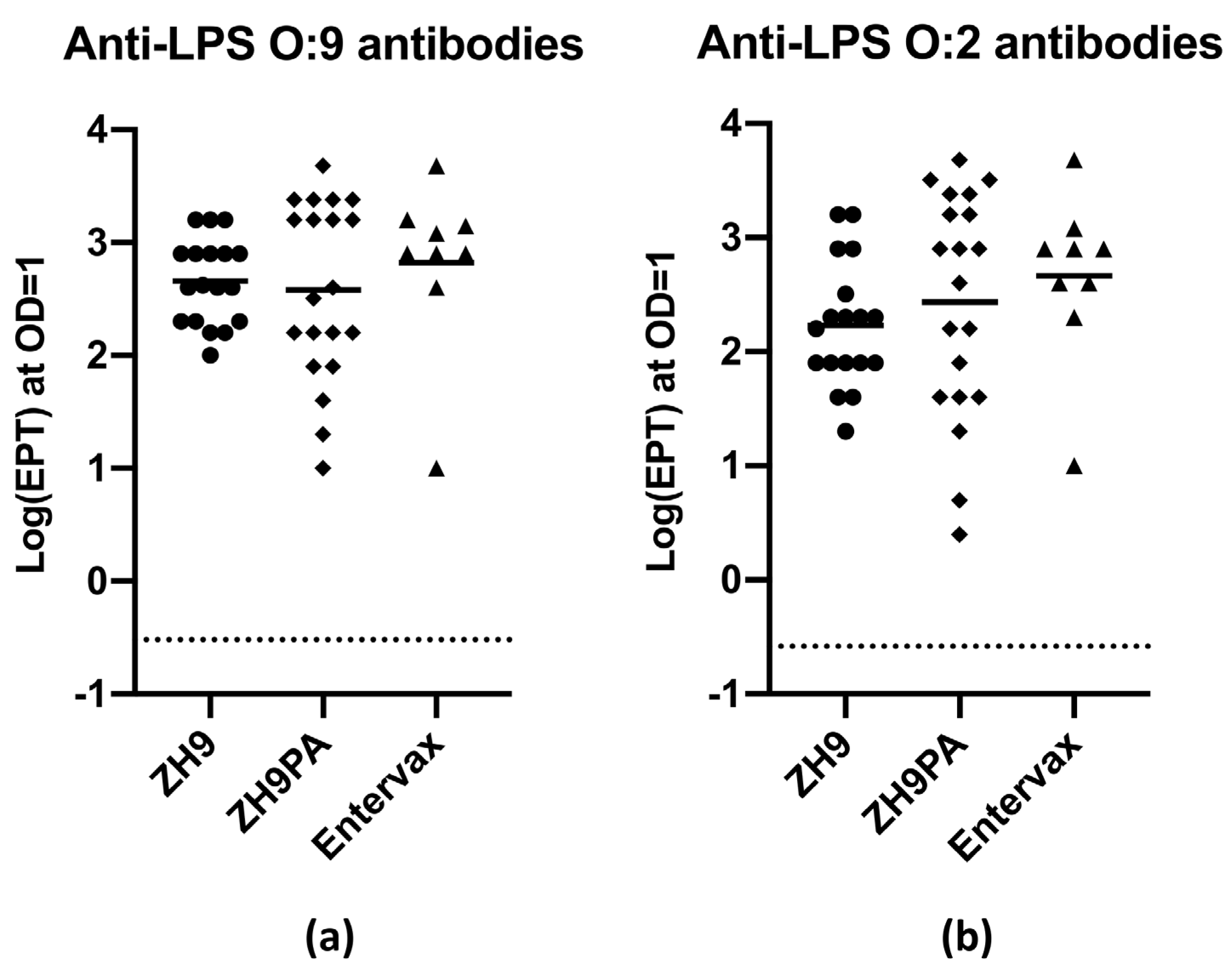
We generated a basic vaccine formulation that mixed equivalent amounts of the ZH9 parental strain and the new ZH9PA strain to create a bivalent vaccine designed to generate immune responses against both S. Typhi and S. Paratyphi A bacteria. In order to assess antibody responses elicited against LPS antigens, and to ensure that the combination of two strains did not impair the response to individual LPS antigens, several groups of naive Balb/c mice were vaccinated subcutaneously with 1 × 108 CFU of ZH9, 1 × 108 CFU of ZH9PA or a basic Entervax™ formulation (a 1:1 combination of 0.5 × 108 CFU of ZH9 and 0.5 × 108 CFU of ZH9PA). Serum samples were obtained before vaccination (d0) and at 35 or 42 days following vaccination and analysed for anti-LPS O:9 or O:2 IgG antibody responses using enzyme-linked immunosorbent assays (ELISA).
By ELISA, it was clear that in mice receiving the dual vaccine combination, both the parental ZH9 strain and the new ZH9PA strain could generate equivalent anti-LPS IgG responses against LPS O:9 and O:2, respectively (Figure 5). Importantly, this provided evidence that both strains could be administered together without one strain outcompeting the other in terms of antigenic dominance. Mice vaccinated with each strain individually were also capable of mounting robust IgG responses against LPS targets (Figure 5). Due to the high level of sequence homology between O:9 and O:2 LPS proteins, it was not possible to fully discriminate between the specific anti-O:9 or anti-O:2 antibody responses using currently available antigens and reagents. However, overall, we concluded that immunocompetent mice could mount equivalent immunogenic IgG antibody responses against both ZH9 and ZH9PA, making the combination a promising bivalent vaccine candidate for the prevention of enteric fever.
3. Discussion
We have described a two-step process to genetically engineer a novel strain of Salmonella that has the genotype of attenuated S. Typhi ZH9 but has been modified to express the flagellin and LPS of S. Paratyphi A. This novel strain, ZH9PA, showed similar growth kinetics to the parental ZH9 strain, with a negligible impact on strain fitness. When ZH9PA was administered in equivalent proportions with ZH9 to create the basic formulation for a new vaccine, Entervax™, immunogenic IgG antibody responses were observed against both ZH9 (LPS O:9) and ZH9PA (LPS O:2) following subcutaneous vaccination of immunocompetent mice.
Several groups have attempted to develop vaccines that can protect against either S. Typhi or S. Paratyphi A, but few have focussed on developing a vaccine that protects against both. Those that have pursued a bivalent approach have typically introduced changes to both S. Typhi and S. Paratyphi wild-type strains, requiring safety and environmental validation of two independent strains [25]. Through targeted engineering of the ZH9 strain as the vaccine chassis, we have leveraged an existing profile of robust clinical safety with validated stable attenuating mutations (ΔaroC, ΔssaV) to build a bivalent vaccine formulation capable of eliciting immune responses against both S. Typhi and S. Paratyphi A.
In our study, we used Xer-cise technology to successfully replace the fliC gene encoding S. Typhi ZH9 flagellin with the fliC gene encoding S. Paratyphi A flagellin, and to remove the antibiotic resistance gene used to select the correct mutant from the bacterial chromosome. We used the same recombination technology to replace the rfbE gene encoding CDP-d-tyvelose 2-epimerase with the spacer gene, wbdR. Our results showed that simple deletion of rfbE was not sufficient to maintain the long-O antigens that were a critical feature of the parental strain. This may have been due to disruption of operon transcription, as downstream of rfbE, the gene rfbX encodes a Wzx translocase involved in the translocation of bacterial O-antigen repeat units across the cytoplasmic membrane [26,27]. These findings are in line with those described by Hong et al. 2012, where Wzx translocation was reported to be Salmonella serotype-specific for the repeat-unit structure, and variants with sugar differences were translocated with lower efficiency and minimal production of long-chain O antigens [28]. Therefore, we found it necessary to replace the deleted rfbE gene with a spacer gene to enable native generation of long-O antigens and maintain translocase activity.
The two S. Paratyphi A antigens that we introduced into S. Typhi ZH9 were carefully chosen as robust vaccine components: both the O-antigen from LPS and the H-antigen from flagellin are highly immunogenic and have been used as the basis for serotyping Salmonella serovars together with the Vi antigen as part of the Kauffmann–White scheme for many years [29]. Both are potent pathogen-associated molecular patterns (PAMPs) that stimulate innate immune responses [30]. The glycolipid LPS forms part of the outer leaflet of the exterior Salmonella bacterial membrane, and the O-antigen polysaccharide represents one of the two carbohydrate regions within the LPS structure [29,31,32]. Antibodies generated against O-antigens are highly protective against lethal Salmonella infections in mice [33,34,35]. Similarly, flagellin, a subunit of the bacterial flagellum that enables motility, chemotaxis and invasion, is a potent immune activator [30]. Mice treated with vaccines targeting flagellin proteins are also significantly protected against lethal Salmonella challenges [36,37,38].
There were some technical limitations that we noted in our study. During the initial testing and development of our ELISA assay designed to detect anti-LPS IgG antibodies, it was not possible to fully discriminate between specific anti-S. Typhi O:9 or anti-S. Paratyphi A O:2 antibody responses using currently available antigens and reagents. The LPS moieties of both S. Typhi (O:9) and S. Paratyphi A (O:2) share substantial homology, with only a single major moiety differing between LPS O:9 and LPS O:2 (tyvelose and paratose, respectively) [39]. Notwithstanding this lack of absolute specificity, it was clear from our results that both the individual ZH9 and ZH9PA strains as well as the combination of the two strains (ZH9+ZH9PA) generated anti-LPS O:9 and O:2 IgG responses. Similarly, we noted some cross-reactivity in our immunostaining assays designed to detect flagellin antigens. Bacteria that had been modified to express H:a flagellin (but retain their native LPS; the ZH9PF strain) showed a low level of positive membrane staining when probed with the anti-flagellin H:d antiserum. This suggested that the polyclonal antiserum was recognising surface proteins in addition to the key target, flagellin. This interpretation is supported by the loss of this positive staining when the final ZH9PA strain (where the native versions of both flagellin and LPS were absent) was probed with H:d antiserum. Nevertheless, it remained clear that swapping the fliC gene successfully converted the flagellin expressed by ZH9 from an H:d to H:a serotype. Since S. Typhi is restricted to human hosts only, we used subcutaneous immunisation as a proxy for validating systemic immune responses to the Entervax™ formulation in a mouse model; however, the oral route of administration will be pursued in clinical trials based on strong positive safety and immunogenicity data from the clinical testing of ZH9. Similarly, since both of the Salmonella strains targeted by our vaccine represent biosafety category 3 pathogens [40], we were unable to validate vaccine (antibody) functionality by serum bactericidal assay or opsonophagocytic killing assay; this will be tested in the clinical trial setting using patient material. Finally, it was also not possible to test vaccine protection in an in vivo challenge model, since S. Typhi is a human-specific pathogen, and no robust mouse models exist.
Overall, this study describes a novel bivalent vaccine capable of generating immunogenic antibody responses against both S. Typhi and S. Paratyphi A using a bacterial strain with an established safety record. Our in vitro data confirm the successful genetic engineering of this vaccine, and our in vivo data highlight the immunogenicity of this bivalent vaccine based on IgG antibody responses. Based on this evidence, we have now received regulatory acceptance to initiate a Phase I clinical trial to confirm the safety, tolerability and immunogenicity of the Entervax™ vaccine delivered as an oral formulation in healthy human volunteers (NCT04349553) [22]. This vaccine is positioned to address a major unmet need in global healthcare by providing a new technology that is capable of targeting both S. Typhi and the increasingly prevalent S. Paratyphi A for the prevention of enteric fever.
4. Materials and Methods
4.1. Bacterial Strains and Media
The target strain for chromosomal modifications was Prokarium’s proprietary S. Typhi strain ZH9 (ΔaroC, ΔssaV) derived from S. Typhi Ty2 [17]. This strain was cultured in LB broth containing 2 mg/L 4-aminobenzoic acid, 2 mg/L 2,3-dihydroxybenzoic acid, 8 mg/L L-phenylalanine, 8 mg/L L-tryptophan and 8 mg/L L-tyrosine, and on 1.5% agar plates. SPAV was an attenuated S. Paratyphi A strain (ΔssaV) derived from S. enterica serotype Paratyphi A (NCTC 9322; ECACC, Porton Down, UK). Top10 Escherichia coli bacteria (Life Technologies, Paisley, UK) were used for routine cloning. LB and LB–aro mix media were supplemented with 20 µg/mL chloramphenicol to select transformants and chromosomal integrants. All bacterial cultures were incubated at 37 °C, with shaking at 200 revolutions per minute (RPM) for liquid cultures. The full list of strains and plasmids used in this study are described in Table 1.
4.2. Generation of Dif-Flanked Antibiotic Resistance Gene Insertion Cassette
The de novo-synthesised chloramphenicol acetyltransferase (cat) gene from pBRT1Nc was amplified by polymerase chain reaction (PCR) using Q5 polymerase at 1 unit/µL (NEB, Hitchin, UK) and 5NotIdifcat and 3NotIdifcat primers (diluted 1:10 in sterile water from a stock solution of 100 pmol/µL). The DifCAT cassette that was generated was cut with a NotI restriction enzyme (NEB, Hitchin, UK) and ligated to generate Xer-cise plasmids pUCpW_difCAT and pUCpF_difCAT using Quick-Stick Ligase (Bioline Reagents Limited, London, UK) and Top10 E. coli competent cells (see Results) [41]. These were also amplified with primers rfbEdelF and rfbEdelR (designed with homologous 5′ sequences to rfbS and rfbX genes, respectively) and Q5 polymerase (NEB, Hitchin, UK) to create a rfbE chromosomal deletion cassette.
4.3. Chromosomal Integration Procedure
Chromosomal replacement and gene deletion were carried out as previously described [23]. Briefly, S. Typhi ZH9 was first transformed with a pLGBK plasmid coding for λ Red gene functions for integration of linear DNA. Integration cassettes were manufactured by Oxford Genetics (Oxford, UK), linearised with SalI and SacI restriction enzymes, purified and concentrated using gel purification (Zymoclean, Irvine, CA, USA) to add a minimum of 300 ng DNA to 50 µL electrocompetent ZH9(pLGBK) or ZH9PF (pLGBK). After electroporation, cells were resuspended in 1 mL LB–aro mix containing 0.02% arabinose and incubated for 18 h at 37 °C and 200 RPM for recovery and integration. Cells were plated on LB–aro mix containing 20 μg/mL chloramphenicol. Colonies were screened by polymerase chain reaction (PCR) with primers designed on the genomic DNA outside the homologous sequences of the integration cassette using My Taq HS Red mix (Bioline Reagents Limited, London, UK). Positive clones were cultured overnight in 5 mL LB at 37 °C and 200 RPM to delete the cat gene using Xer-cise technology [41]. Glycerol stocks were made and stored at −80 °C, and modified chromosomal sequences were amplified with Q5 polymerase and checked by sequencing using the diagnostic primers listed in Table 2.
4.4. Conversion of Flagellin
The fliC replacement cassette was synthesised to comprise S. Paratyphi A fliC, flanked on one side by approximately 700 bp of DNA homologous to the S. Typhi fliC upstream gene, fliD, and on the other side by approximately 700 bp of DNA homologous to the S. Typhi fliC downstream T0919 pseudogene DNA sequence. A NotI restriction site at the 3′ end of the S. Paratyphi A fliC gene was incorporated to enable insertion of the dif-flanked cat antibiotic resistance marker gene amplified with primers designed with a corresponding NotI restriction site. TOP10 E. coli bacteria were used for transformation to generate a pUCpF_difCAT plasmid. Chromosomal replacement of the S. Typhi fliC gene with S. Paratyphi A fliC was carried out as previously described [23]. Briefly, S. Typhi ZH9 was first transformed with a pLGBK plasmid coding for λ Red gene functions for integration of linear DNA. Electrocompetent ZH9(pLGBK) was transformed, and the replacement cassette was excised from pUCpF-difCAT using SalI and SacI restriction digestion. Transformed colonies were selected on LB–aro mix agar plates supplemented with 20 μg/mL chloramphenicol. Single colonies were isolated and cultured overnight in LB–aro mix broth in the absence of antibiotics. Xer recombination resulted in the deletion of the cat gene to generate chloramphenicol-sensitive colonies of ZH9PF.
4.5. Immunostaining
For immunofluorescence microscopy, a volume of bacterial culture equivalent to an optical density (OD) at A600 nm = 1 was collected, centrifuged at 6000× g for 5 min, and washed in phosphate-buffered saline (PBS). Pellets were resuspended in 10 µL of PBS with 1 µL of primary antibody and incubated for 10 min at room temperature. Flagellin analysis was carried out by staining bacteria with H:d antiserum (SSI Diagnostica) or H:a antiserum (SSI Diagnostica, Oxford Biosystems, Oxford, UK). LPS analysis was carried out by staining bacteria with anti-S. Typhi LPS (clone B348M; GeneTex, Insight Biotechnology, Wembley, UK) or anti-S. Paratyphi A LPS (clone 10B10G; Bio-Rad Laboratories, Hemel Hempstead, UK) monoclonal antibodies. Stained bacterial cells were washed in PBS and pellets were resuspended in 10 µL of PBS with 1 µL of goat anti-mouse secondary antibody (for LPS; Sigma-Aldrich Merck Life Science UK Limited, Gillingham, UK) or goat anti-rabbit secondary antibody (for flagellin; Sigma-Aldrich) conjugated to the Dylight 488 fluorochrome (Bio-Rad Laboratories, Hemel Hempstead, UK) for 10 min at room temperature. Bacterial cells were subsequently washed in PBS and a small volume was applied onto microscope slides to be visualised using a fluorescent microscope (Zeiss Axiophot, Carl Zeiss AG, Oberkochen, Germany) with an attached Zeiss Axiocam camera.
4.6. Western and Dot Blot
To generate protein samples for western blot, bacterial cells were harvested by centrifugation at 10,000× g and 4 °C for 10 min, washed in 10 mM Tris Buffer pH 7.4, then sonicated for 4 min with 30 s bursts at 60% amplitude (Fisherbrand™ 120 Sonic Dismembrator 50/60 Hz (Fisher Scientific UK Ltd., Loughborough, UK)). An initial centrifugation was performed at 6000× g and 4 °C for 10 min to remove unbroken cells; and a second centrifugation was performed at 30,000× g and 4 °C for 45 min to separate the cell membranes and cytoplasmic fractions. Pelleted envelopes were resuspended in 10 mM Tris Buffer pH 7.4 plus 2% (v/v) Triton X-100 (VWR, Lutterworth, UK) and incubated at 25 °C for 15 min to allow solubilisation of the inner membrane. Extracted envelopes were collected by centrifugation at 30,000× g and 4 °C for 45 min. Pellets were washed and resuspend in 10 mM Tris Buffer pH 7.4. Cytoplasmic and membrane fractions were stored at −80 °C prior to western blot analysis. Flagellin H:d and H:a proteins were purified from ZH9 and SPAV strains, respectively (The Native Antigen Company, Kidlington, UK) and used as positive controls; flagellin proteins were stored at −80 °C. Protein separation by molecular mass was performed via sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE) using NuPAGE 4–12% Bis-Tris Protein Gels and MES running buffer (Life Technologies, Paisley, UK) following the manufacturer’s instructions. The SeeBlue™ Plus2 pre-stained protein standard (Life Technologies, Paisley, UK) was used as a molecular marker. Protein gels were transferred onto membranes using a semi-dry trans-blot turbo transfer system (Bio-Rad Laboratories, Hemel Hempstead, UK), when required.
To generate protein samples for dot blot, bacteria were grown at 37 °C for 18 h. 0.5 mL of all tested cultures were heat-inactivated at 95 °C for 10 min. 5 μL of heat-inactivated cultures were dotted onto nitrocellulose membranes.
Membranes were incubated with 5% skim milk diluted in PBS for 1 h at room temperature to minimise non-specific antibody binding prior to adding primary antibodies following the same staining protocol described in the ‘Immunostaining’ section. After three washes with 0.05% Tween-20 (VWR, Lutterworth, UK) diluted in PBS, membranes were incubated with secondary antibodies following the same staining protocol described in the ‘Immunostaining’ section. The horseradish peroxidase (HRP) substrate 3,3′,5,5′-Tetramethylbenzidine (TMB; Sigma-Aldrich, Merck Life Science UK Limited, Gillingham, UK) was added for 10 min. Images were acquired using the GelDoc RX+ system (BioRad Laboratories, Hemel Hempstead, UK).
4.7. Silver Staining
LPS was extracted using a bacterial LPS extraction kit (2B Scientific, Upper Heyford, UK) following manufacturer’s instructions. Briefly, cells were lysed using organic solutions, and the cell membrane phospholipids and proteins were disrupted and released in solution. LPS was then purified from the mixture with a high salt concentration solution and eluted in 10 mM Tris-HCl buffer (pH 8.0) (Sigma-Aldrich, Merck Life Science UK Limited, Gillingham, UK) after washing off salts with 70% ethanol. SDS-PAGE was performed as described in the ‘Western Blotting’ section, and gels were then silver stained using the SilverQuest Silver Staining Kit (Invitrogen, Paisley, UK) following manufacturer’s instructions. Images were acquired using the GelDoc RX+ system (BioRad Laboratories, Hemel Hempstead, UK).
4.8. Growth Studies
Bacteria were seeded into liquid LB medium cultures starting at OD = 0.1 and grown for 24 h. At regular intervals, samples were taken and analysed by spectrophotometry or by titration on agar plates. OD at A600 nm was reported as a measure of growth density. Bacterial titers were measured by 10-fold serial dilutions prepared in sterile PBS; aliquots of three dilutions were plated in triplicate on LB agar plates supplemented with aromix and L-Tyr. The dilution range was selected to culture a reasonable number of bacteria to count per plate (between 30 and 300 colonies per plate). Plates were incubated at 37 °C for ≥18 h for colony counting, and colony-forming units (CFU) per ml were calculated by multiplying the average number of colonies on a plate by the corresponding dilution factor multiplied by 10.
4.9. Murine Immunogenicity
Bacteria for in vivo studies were grown at 37 °C in LB–aro up to the mid-exponential phase, harvested by centrifugation (4100× g at 4 °C for 20 min), washed in sterile cold PBS and concentrated to obtain a final cell density of approximately 6 × 1010 ± 0.5 × 1010 CFU/mL (based on OD600 nm calculations) in PBS containing 10% glycerol (Sigma-Aldrich, Merck Life Science UK Limited, Gillingham, UK). These were frozen at −80 °C until needed. On the day of dosing, vials were thawed at room temperature and vortexed gently for several seconds to obtain a homogeneous suspension. Vials were centrifuged (4100× g at 4 °C for 20 min), supernatants removed, and pellets resuspended in 14 mL sterile cold PBS. This was repeated twice to ensure the pellet was washed three times in total, followed by a final resuspension in 1 mL sterile cold PBS.
Female Balb/c mice (purchased from Charles River, Margate, UK) at 6–8 weeks of age were maintained in sterilised ventilated cages with ad libitum access to food and water and with room conditions set at a temperature of 22 °C (±1 °C), 60% relative humidity and a 12 h light/dark cycle. Vaccines were diluted to the appropriate concentration in sterile cold PBS, followed by subcutaneous immunisation with 100 µL of solution equivalent to 1 × 108 colony-forming units (CFU) of ZH9, 1108 CFU of ZH9PA and a 1:1 combination of 0.5 × 108 CFU of ZH9 and 0.5 × 108 CFU of ZH9PA. For combination injections, the vaccine strains were mixed, and a single injection was performed. Blood samples were collected on day 0 prior to immunisation via tail prick, and on day 35 or 42 following vaccination as a terminal bleed by cardiac puncture. Blood was left to coagulate on ice, and serum was separated by centrifugation for 5 min at 13,000 RPM. The supernatant was removed into a fresh Eppendorf tube, snap frozen and stored at −80 °C. All studies were performed at Evotec (Macclesfield, UK) under UK Home Office Licenses with local ethical committee clearance. All animal experiments were performed by experienced technicians that had completed the UK Home Office Personal License course and held current personal licenses.
4.10. ELISA
Antigens used in enzyme-linked immunosorbent assays (ELISAs) were LPS O:9 purified from S. Typhi ZH9 (The Native Antigen Company, Kidlington, UK) and LPS O:2 purified from attenuated S. Paratyphi A (generated by the Wellcome Sanger Institute, Hinxton, UK; purchased from The Native Antigen Company Kidlington, UK). Nunc F96 Maxisorp Immunoplates (ThermoFisher Scientific, Altrincham, UK) were coated with LPS antigens at an appropriate concentration for each batch after pilot experiments determined the optimal range of coating concentrations using control antibodies and checked batch purity. Mouse serum (collected as described above) was added to wells in serial dilutions. Pre-vaccination (d0) samples were pooled across mice to generate a negative assay control. Bound serum antibodies were detected with HRP-tagged secondary goat anti-mouse IgG antibodies (Sigma-Aldrich, Merck Life Science UK Limited, Gillingham, UK) and developed with TMB substrate (Sigma-Aldrich, Merck Life Science UK Limited, Gillingham, UK) for 10 min in the dark. The reaction was stopped with 1 M sulphuric acid, and absorbance was measured at 450 nm within 15 min of adding the Stop Solution with a Spark plate reader (Tecan Group Ltd., Männedorf, Switzerland). End-point titres were calculated by recording the dilution that intersected the curve at OD = 1 for each serum sample.
4.11. Statistical Analysis
Graphical data was plotted using GraphPad Prism 9 [42]. Data were statistically compared using either a one-way analysis of variance (ANOVA) with Tukey’s multiple comparisons test or a mixed-effects two-way ANOVA with a Bonferroni multiple comparisons test, where appropriate. Results with a p-value <0.05 were considered statistically significant.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/22/6/3287/s1.
Author Contributions
Conceptualization, A.S., L.D. and R.M.C.; formal analysis, A.S., C.P. and M.C.; data curation, A.S. and C.P.; writing—original draft preparation, A.S. and L.D.; writing—review and editing, A.S., C.P., L.D. and R.M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partly funded by Innovate UK (102728). The funders had no role in the study design, conduct or analysis.
Institutional Review Board Statement
All studies were performed at Evotec under UK Home Office Licenses with local ethical committee clearance. All animal experiments were performed by experienced technicians that had completed the UK Home Office Personal License course and held current personal licenses.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original western blot, dot blot and silver staining data presented in this study are available as supplementary figures. All other data presented in this study are available on request from the corresponding author. The data are not publicly available due to the limited scope of the datasets.
Acknowledgments
Annelise Soulier is now employed by Neobe Therapeutics and Rocky Cranenburgh is now employed by Bitrobius Genetics. The authors would like to thank Mike Darsley (MDbioLogic) for consulting on analytical assay development and Gordon Dougan and Derek Pickard at the Sanger Institute for generating the S. Paratyphi A mutant strain.
Conflicts of Interest
All authors are or were employees of Prokarium Ltd. at the time of the design and execution of this study. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Parry, C.M.; Ribeiro, I.; Walia, K.; Rupali, P.; Baker, S.; Basnyat, B. Multidrug resistant enteric fever in South Asia: Unmet medical needs and opportunities. BMJ 2019, 364, k5322. [Google Scholar] [CrossRef] [Green Version]
- GBD. The global burden of typhoid and paratyphoid fevers: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Kariuki, S.; Gordon, M.A.; Feasey, N.; Parry, C.M. Antimicrobial resistance and management of invasive Salmonella disease. Vaccine 2015, 33 (Suppl. 3), C21–C29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, J.; Khan, A.S.; Khan, H.A.; Gilani, S.A.; Akram, S.J.; Ahmad, F.J.; Mehboob, R. Extensively Drug-Resistant (XDR) Typhoid: Evolution, Prevention, and Its Management. Biomed Res. Int. 2020, 2020, 6432580. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Anwar, E.; Goldberg, E.; Fraser, A.; Acosta, C.J.; Paul, M.; Leibovici, L. Vaccines for preventing typhoid fever. Cochrane Database Syst. Rev. 2014. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.B. Vaccines for typhoid fever and other salmonelloses. Curr. Opin. Infect. Dis. 2012, 25, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Sahastrabuddhe, S.; Carbis, R.; Wierzba, T.F.; Ochiai, R.L. Increasing rates of Salmonella Paratyphi A and the current status of its vaccine development. Expert Rev. Vaccines 2013, 12, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, Y.K.; Adedare, T.A.; Ehinmidu, J.O. Antibiotic sensitivity profiles of salmonella organisms isolated from presumptive typhoid patients in Zaria, northern Nigeria. Afr. J. Med. Med. Sci. 2005, 34, 109–114. [Google Scholar] [PubMed]
- Akinyemi, K.O.; Bamiro, B.S.; Coker, A.O. Salmonellosis in Lagos, Nigeria: Incidence of Plasmodium falciparum-associated co-infection, patterns of antimicrobial resistance, and emergence of reduced susceptibility to fluoroquinolones. J. Health Popul. Nutr. 2007, 25, 351–358. [Google Scholar] [PubMed]
- Antillón, M.; Warren, J.L.; Crawford, F.W.; Weinberger, D.M.; Kürüm, E.; Pak, G.D.; Marks, F.; Pitzer, V.E. The burden of typhoid fever in low- and middle-income countries: A meta-regression approach. PLoS Negl. Trop. Dis. 2017, 11, e0005376. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, K.S.; Pak, G.D.; Excler, J.L.; Sahastrabuddhe, S.; Marks, F.; Kim, J.H.; Mogasale, V. Spatial and Temporal Patterns of Typhoid and Paratyphoid Fever Outbreaks: A Worldwide Review, 1990–2018. Clin. Infect. Dis. 2019, 69, S499–S509. [Google Scholar] [CrossRef]
- Guzman, C.A.; Borsutzky, S.; Griot-Wenk, M.; Metcalfe, I.C.; Pearman, J.; Collioud, A.; Favre, D.; Dietrich, G. Vaccines against typhoid fever. Vaccine 2006, 24, 3804–3811. [Google Scholar] [CrossRef]
- Roland, K.L.; Tinge, S.A.; Kochi, S.K.; Thomas, L.J.; Killeen, K.P. Reactogenicity and immunogenicity of live attenuated Salmonella enterica serovar Paratyphi A enteric fever vaccine candidates. Vaccine 2010, 28, 3679–3687. [Google Scholar] [CrossRef] [PubMed]
- Wahid, R.; Kotloff, K.L.; Levine, M.M.; Sztein, M.B. Cell mediated immune responses elicited in volunteers following immunization with candidate live oral Salmonella enterica serovar Paratyphi A attenuated vaccine strain CVD 1902. Clin. Immunol. 2019, 201, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Darton, T.C.; Jones, C.; Blohmke, C.J.; Waddington, C.S.; Zhou, L.; Peters, A.; Haworth, K.; Sie, R.; Green, C.A.; Jeppesen, C.A.; et al. Using a Human Challenge Model of Infection to Measure Vaccine Efficacy: A Randomised, Controlled Trial Comparing the Typhoid Vaccines M01ZH09 with Placebo and Ty21a. PLoS Negl. Trop. Dis. 2016, 10, e0004926. [Google Scholar] [CrossRef] [PubMed]
- Hindle, Z.; Chatfield, S.N.; Phillimore, J.; Bentley, M.; Johnson, J.; Cosgrove, C.A.; Ghaem-Maghami, M.; Sexton, A.; Khan, M.; Brennan, F.R.; et al. Characterization of Salmonella enterica derivatives harboring defined aroC and Salmonella pathogenicity island 2 type III secretion system (ssaV) mutations by immunization of healthy volunteers. Infect. Immun. 2002, 70, 3457–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.D.; McKenzie, R.; O’Neill, J.P.; Larsson, C.J.; Bourgeois, A.L.; Shimko, J.; Bentley, M.; Makin, J.; Chatfield, S.; Hindle, Z.; et al. Evaluation of Salmonella enterica serovar Typhi (Ty2 aroC-ssaV-) M01ZH09, with a defined mutation in the Salmonella pathogenicity island 2, as a live, oral typhoid vaccine in human volunteers. Vaccine 2006, 24, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.D.; Tenney, K.M.; Larsson, C.J.; O’Neill, J.P.; Ventrone, C.; Bentley, M.; Upton, A.; Hindle, Z.; Fidler, C.; Kutzko, D.; et al. The novel oral typhoid vaccine M01ZH09 is well tolerated and highly immunogenic in 2 vaccine presentations. J. Infect. Dis. 2005, 192, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, C.E.; Sadigh, K.S.; Carmolli, M.P.; Harro, C.; Sheldon, E.; Lindow, J.C.; Larsson, C.J.; Martinez, T.; Feller, A.; Ventrone, C.H.; et al. In a randomized, double-blinded, placebo-controlled trial, the single oral dose typhoid vaccine, M01ZH09, is safe and immunogenic at doses up to 1.7 × 10(10) colony-forming units. Vaccine 2010, 28, 3602–3608. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.; Nguyen, T.D.; Nguyen, T.T.; Ninh, T.T.; Tran, N.B.; Nguyen, V.M.; Tran, T.T.; Cao, T.T.; Pham, V.M.; Nguyen, T.C.; et al. A randomised trial evaluating the safety and immunogenicity of the novel single oral dose typhoid vaccine M01ZH09 in healthy Vietnamese children. PLoS ONE 2010, 5, e11778. [Google Scholar] [CrossRef]
- Prokarium. Study to Evaluate the Safety, Tolerability and Immunogenicity of a Potential Enteric Fever Vaccine. 2019. Available online: https://ClinicalTrials.gov/show/NCT04349553 (accessed on 27 February 2021).
- Bloor, A.E.; Cranenburgh, R.M. An efficient method of selectable marker gene excision by Xer recombination for gene replacement in bacterial chromosomes. Appl. Environ. Microbiol. 2006, 72, 2520–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars; WHO Collaborating Centre for Reference and Research on Salmonella, Institut Pasteur: Paris, France, 2007. [Google Scholar]
- Liu, Q.; Li, P.; Luo, H.; Curtiss, R., 3rd; Kong, Q. Attenuated Salmonella Typhimurium expressing Salmonella Paratyphoid A O-antigen induces protective immune responses against two Salmonella strains. Virulence 2019, 10, 82–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Cole, R.A.; Reeves, P.R. An O-antigen processing function for Wzx (RfbX): A promising candidate for O-unit flippase. J. Bacteriol. 1996, 178, 2102–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, C.; Sherwood, R.; Gheesling, L.L.; Brenner, F.W.; Fields, P.I. Molecular analysis of the rfb O antigen gene cluster of Salmonella enterica serogroup O:6,14 and development of a serogroup-specific PCR assay. Appl. Environ. Microbiol. 2003, 69, 6099–6105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Cunneen, M.M.; Reeves, P.R. The Wzx translocases for Salmonella enterica O-antigen processing have unexpected serotype specificity. Mol. Microbiol. 2012, 84, 620–630. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin-a potent immunomodulatory agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.; Pál, T. Lipopolysaccharide: A tool and target in enterobacterial vaccine development. Biol. Chem. 2008, 389, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Lee, J.O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colwell, D.E.; Michalek, S.M.; Briles, D.E.; Jirillo, E.; McGhee, J.R. Monoclonal antibodies to Salmonella lipopolysaccharide: Anti-O-polysaccharide antibodies protect C3H mice against challenge with virulent Salmonella typhimurium. J. Immunol. 1984, 133, 950–957. [Google Scholar]
- Forbes, S.J.; Martinelli, D.; Hsieh, C.; Ault, J.G.; Marko, M.; Mannella, C.A.; Mantis, N.J. Association of a protective monoclonal IgA with the O antigen of Salmonella enterica serovar Typhimurium impacts type 3 secretion and outer membrane integrity. Infect. Immun. 2012, 80, 2454–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michetti, P.; Mahan, M.J.; Slauch, J.M.; Mekalanos, J.J.; Neutra, M.R. Monoclonal secretory immunoglobulin A protects mice against oral challenge with the invasive pathogen Salmonella typhimurium. Infect. Immun. 1992, 60, 1786–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, J.S.; Seok Kim, J.; Im Jang, J.; Kim, B.H.; Young Yoo, S.; Hyeon Choi, J.; Bang, I.S.; Lee, I.S.; Keun Park, Y. Enhancement of host immune responses by oral vaccination to Salmonella enterica serovar Typhimurium harboring both FliC and FljB flagella. PLoS ONE 2013, 8, e74850. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Tennant, S.M.; Wang, J.Y.; Schmidlein, P.J.; Lees, A.; Ernst, R.K.; Pasetti, M.F.; Galen, J.E.; Levine, M.M. Salmonella enterica serovar enteritidis core O polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. enteritidis. Infect. Immun. 2011, 79, 4240–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobat, S.; Flores-Langarica, A.; Hitchcock, J.; Marshall, J.L.; Kingsley, R.A.; Goodall, M.; Gil-Cruz, C.; Serre, K.; Leyton, D.L.; Letran, S.E.; et al. Soluble flagellin, FliC, induces an Ag-specific Th2 response, yet promotes T-bet-regulated Th1 clearance of Salmonella typhimurium infection. Eur. J. Immunol. 2011, 41, 1606–1618. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Knirel, Y.A.; Feng, L.; Perepelov, A.V.; Senchenkova, S.N.; Reeves, P.R.; Wang, L. Structural diversity in Salmonella O antigens and its genetic basis. FEMS Microbiol. Rev. 2014, 38, 56–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- HSE. The Approved List of Biological Agents: Advisory Committee on Dangerous Pathogens; Health and Safety Executive: Bootle, UK, 2013. [Google Scholar]
- Boudehen, Y.M.; Wallat, M.; Rousseau, P.; Neyrolles, O.; Gutierrez, C. An improved Xer-cise technology for the generation of multiple unmarked mutants in Mycobacteria. Biotechniques 2020, 68, 106–110. [Google Scholar] [CrossRef] [PubMed]
- GraphPad-Prism. Software for Data Analysis and Graphing Solutions, Version 9; GraphPad Software: San Diego, CA, USA, 2020. [Google Scholar]
Figure 1.
Replacing the S. Typhi (H:d) flagellin with S. Paratyphi A (H:a) flagellin. (a) The genetic engineering process to generate S. Typhi ZH9 expressing S. Paratyphi A flagellin (ZH9PF). Adapted with permission from Bloor and Cranenburgh, 2006 [23]. (b) Fluorescence microscopy with S. Typhi ZH9 and the derivative strain, ZH9PF, probed with H:d antiserum (anti-S. Typhi) or H:a antiserum (anti-S. Paratyphi A) plus Dylight 488 secondary antibodies; the left column images are phase contrast images, and the right column images are immuno-fluorescence images. Images were taken at 100× magnification. Scale bars represent 10 µm. Representative images were based on three independent experimental repeats.
Figure 1.
Replacing the S. Typhi (H:d) flagellin with S. Paratyphi A (H:a) flagellin. (a) The genetic engineering process to generate S. Typhi ZH9 expressing S. Paratyphi A flagellin (ZH9PF). Adapted with permission from Bloor and Cranenburgh, 2006 [23]. (b) Fluorescence microscopy with S. Typhi ZH9 and the derivative strain, ZH9PF, probed with H:d antiserum (anti-S. Typhi) or H:a antiserum (anti-S. Paratyphi A) plus Dylight 488 secondary antibodies; the left column images are phase contrast images, and the right column images are immuno-fluorescence images. Images were taken at 100× magnification. Scale bars represent 10 µm. Representative images were based on three independent experimental repeats.

Figure 2.
Modifying LPS (O:9) to LPS (O:2). (a) Part of the wild-type O-antigen locus from S. Typhi ZH9 was modified using two test approaches: by deleting the majority of the rfbE cistron to generate S. Typhi ZH9PL2 or by replacing the rfbE cistron with a spacer DNA sequence to maintain the original reading frame to generate S. Typhi ZH9W. (b) Fluorescence microscopy images showing the parental S. Typhi ZH9 and derivative strains, ZH9PL2 and ZH9W, probed with anti-S. Typhi LPS (O:9) or anti-S. Paratyphi A LPS (O:2) monoclonal antibodies followed by Dylight 488 secondary antibodies; the left column images are phase contrast images and the right column images are immuno-fluorescence micrographs. Images were taken at 100× magnification. Scale bars represent 10 µm. Representative images based on three independent experimental repeats. (c) Silver-stained polyacrylamide gel of LPS extracts from the parental S. Typhi ZH9 and derivative strains, ZH9PL2 and ZH9W, indicating the short and long O-antigen chains. LPS = lipopolysaccharide; mAb = monoclonal antibody.
Figure 2.
Modifying LPS (O:9) to LPS (O:2). (a) Part of the wild-type O-antigen locus from S. Typhi ZH9 was modified using two test approaches: by deleting the majority of the rfbE cistron to generate S. Typhi ZH9PL2 or by replacing the rfbE cistron with a spacer DNA sequence to maintain the original reading frame to generate S. Typhi ZH9W. (b) Fluorescence microscopy images showing the parental S. Typhi ZH9 and derivative strains, ZH9PL2 and ZH9W, probed with anti-S. Typhi LPS (O:9) or anti-S. Paratyphi A LPS (O:2) monoclonal antibodies followed by Dylight 488 secondary antibodies; the left column images are phase contrast images and the right column images are immuno-fluorescence micrographs. Images were taken at 100× magnification. Scale bars represent 10 µm. Representative images based on three independent experimental repeats. (c) Silver-stained polyacrylamide gel of LPS extracts from the parental S. Typhi ZH9 and derivative strains, ZH9PL2 and ZH9W, indicating the short and long O-antigen chains. LPS = lipopolysaccharide; mAb = monoclonal antibody.

Figure 3.
Converting flagellin and LPS in the final new strain, ZH9PA. (a) Fluorescence microscopy images showing the S. Typhi ZH9 derivative strain, ZH9PA, probed with anti-S. Typhi (H:d) or anti-S. Paratyphi A (H:a) flagellin antiserum and anti-S. Typhi (O:9) or anti-S. Paratyphi A (O:2) LPS mAbs; the left images are phase contrast images and right images are immuno-fluorescence micrographs. Images were taken at 100× magnification. Scale bars represent 10µm. Representative images based on three independent experimental repeats. (b) Western blots of membrane fractions probed with anti-S. Typhi (H:d) or anti-S. Paratyphi A (H:a) flagellin antisera using ZH9 or SPAV as positive controls, respectively. Purified flagellin proteins were also included as a positive control. (c) Dot blot probed with anti-S. Typhi and anti-S. Paratyphi A LPS mAbs. (d) Silver-stained polyacrylamide gel of LPS preparations from S. Typhi ZH9 and derivative strains, ZH9PA, indicating the short and long O-antigen chains. LPS = lipopolysaccharide; mAb = monoclonal antibody; SPAV = attenuated S. Paratyphi A.
Figure 3.
Converting flagellin and LPS in the final new strain, ZH9PA. (a) Fluorescence microscopy images showing the S. Typhi ZH9 derivative strain, ZH9PA, probed with anti-S. Typhi (H:d) or anti-S. Paratyphi A (H:a) flagellin antiserum and anti-S. Typhi (O:9) or anti-S. Paratyphi A (O:2) LPS mAbs; the left images are phase contrast images and right images are immuno-fluorescence micrographs. Images were taken at 100× magnification. Scale bars represent 10µm. Representative images based on three independent experimental repeats. (b) Western blots of membrane fractions probed with anti-S. Typhi (H:d) or anti-S. Paratyphi A (H:a) flagellin antisera using ZH9 or SPAV as positive controls, respectively. Purified flagellin proteins were also included as a positive control. (c) Dot blot probed with anti-S. Typhi and anti-S. Paratyphi A LPS mAbs. (d) Silver-stained polyacrylamide gel of LPS preparations from S. Typhi ZH9 and derivative strains, ZH9PA, indicating the short and long O-antigen chains. LPS = lipopolysaccharide; mAb = monoclonal antibody; SPAV = attenuated S. Paratyphi A.

Figure 4.
Comparison of growth profiles. Bacteria were seeded into LB broth cultures at OD600 nm = 0.1 and grown for 24 h. At regular intervals, samples were taken and analysed by spectrophotometry or by titration on agar plates. Optical density (a measure of growth density) and bacterial titre were plotted for ZH9 (the parental strain) and ZH9PA (the modified strain). The late exponential growth phase (5 to 8 h) is shown in grey. (a) OD600 nm and CFU/mL measurements compared within each individual strain. (b) OD600 nm or CFU/mL measurements compared between both strains. Statistical comparisons were made using a two-way ANOVA, based on triplicate cultures in a single experiment. CFU = colony-forming units; ml = millilitres; nm = nanometres.
Figure 4.
Comparison of growth profiles. Bacteria were seeded into LB broth cultures at OD600 nm = 0.1 and grown for 24 h. At regular intervals, samples were taken and analysed by spectrophotometry or by titration on agar plates. Optical density (a measure of growth density) and bacterial titre were plotted for ZH9 (the parental strain) and ZH9PA (the modified strain). The late exponential growth phase (5 to 8 h) is shown in grey. (a) OD600 nm and CFU/mL measurements compared within each individual strain. (b) OD600 nm or CFU/mL measurements compared between both strains. Statistical comparisons were made using a two-way ANOVA, based on triplicate cultures in a single experiment. CFU = colony-forming units; ml = millilitres; nm = nanometres.

Figure 5.
Anti-LPS IgG antibody responses following in vivo vaccination. (a) Specific IgG antibody responses against S. Typhi LPS (O:9). (b) Specific IgG antibody responses against S. Paratyphi A LPS (O:2). Antibody responses were evaluated by ELISA in Balb/c mouse serum at 35 or 42 days following subcutaneous vaccination with 108 CFU ZH9 (•), 108 CFU ZH9PA (♦) or a 1:1 mix of 0.5 × 108 CFU of ZH9 and 0.5 × 108 CFU of ZH9PA (Entervax™ basic formulation (▲)). Pre-vaccination (d0) samples were pooled across individual mice to generate the negative assay control (dotted line). Each data point represents an individual mouse, and data were pooled across three independent experiments. Mean values are represented by the horizontal bar. Statistical comparisons were made using a one-way ANOVA. ELISA = enzyme-linked immunosorbent assay; EPT = end point titre; IgG = immunoglobulin G; LPS = lipopolysaccharide; OD = optical density.
Figure 5.
Anti-LPS IgG antibody responses following in vivo vaccination. (a) Specific IgG antibody responses against S. Typhi LPS (O:9). (b) Specific IgG antibody responses against S. Paratyphi A LPS (O:2). Antibody responses were evaluated by ELISA in Balb/c mouse serum at 35 or 42 days following subcutaneous vaccination with 108 CFU ZH9 (•), 108 CFU ZH9PA (♦) or a 1:1 mix of 0.5 × 108 CFU of ZH9 and 0.5 × 108 CFU of ZH9PA (Entervax™ basic formulation (▲)). Pre-vaccination (d0) samples were pooled across individual mice to generate the negative assay control (dotted line). Each data point represents an individual mouse, and data were pooled across three independent experiments. Mean values are represented by the horizontal bar. Statistical comparisons were made using a one-way ANOVA. ELISA = enzyme-linked immunosorbent assay; EPT = end point titre; IgG = immunoglobulin G; LPS = lipopolysaccharide; OD = optical density.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains and plasmids used directly in this study.
| Strain or Plasmid | Description | Source or Reference |
|---|---|---|
| Strains | ||
| Top10 E. coli | F– mcrA Δ(mrr-hsdRMS-mcrBC) φ80lacZΔM15 ΔlacX74 recA1 araD139 Δ(ara-leu)7697 galU galK λ– rpsL(StrR) endA1 nupG | Life Technologies, Paisley, UK |
| S. Typhi ZH9 | Ty2 ΔaroC ΔssaV | Prokarium |
| S. Typhi ZH9PF | Ty2 ΔaroC ΔssaV Δt0918::SPA0911 | This work |
| S. Typhi ZH9PL2 | Ty2 ΔaroC ΔssaV ΔrfbE | This work |
| S. Typhi ZH9PA | Ty2 ΔaroC ΔssaV ΔrfbE::wbdR Δt0918::SPA0911 | This work |
| Plasmids | ||
| pBRT1Nc | Synthesised with Chloramphenical resistance gene | This work |
| pUCFlic2 | Synthesised S. Paratyphi A fliC | This work |
| pUCWbdR | Synthesised E. coli wbdR | This work |
| pUCpF-difcat | Precursor S. Paratyphi A fliC integration Xer-cise plasmid | This work |
| pUCpW-difcat | Precursor wbdR integration Xer-cise plasmid | This work |
| pL2-difcat | Precursor rfbE deletion Xer-cise cassette | This work |
| pLGBK | Lambda Red helper plasmid | Prokarium |
Abbreviations: E. coli = Escherichia coli; PCR = polymerase chain reaction.
Table 2.
PCR and sequencing primers used in this study.
| Primer Name | Primer Sequence (5′-3′) | Primer Function |
|---|---|---|
| 5NotIdifcat | taagcggccgcATTTAACATAATATACATTATGCGCACCgcccgaacaccac | Primers designed to include NotI restriction sites at the 5′ and 3′ end respectively. The lowercase letters represent the region of homology to pBRT1Nc; the NotI restriction site is underlined and dif sites are in capital letters |
| 3NotIdifcat | ggcggccgcGGTGCGCATAATGTATATTATGTTAAATgggcgagtttacatctcaaaaccg | |
| rfbE del F | AATAGGATGGAAAAGAGAGTTCTCTCTTGTTGATGCATTAACTGAAATAATTGAAGAGGAAGGGAAATGAAAAGCTTGGTACCGAGCTCG | rfbE deletion |
| rfbE del R | TTTGAAAGCCAAGAGGAAGCGGCAATAATAAGATGTCTTGGAATTCTAACCAACCTCAGTTTCCTCACTCTAGATGCATGCTCGAGCGGC | rfbE deletion |
| L1 | AGGCTTGACTACAGAGCATTTAGATTATGTAG | Diagnostic primer for LPS locus |
| L6 | ACATACTTCTACAATTAAGGAGTGAGAAGATTGATTATTAATACT | Diagnostic primer for LPS locus |
| L2 | TCACGACTTACATCCTACTTCG | Diagnostic primer for LPS locus |
| L3 | TGTTCCTGCCGGTATAACTG | Diagnostic primer for LPS locus |
| L4 | CAGTTTCCTCACGTCAGCTT | Diagnostic primer for LPS locus |
| L5 | CTGGCCATAATGCTTGTAATACCGCA | Diagnostic primer for LPS locus |
| F1 | GCTGACTTGCGCATAAGCTTTGA | Diagnostic primer for Flagellin locus |
| F8 | AACAGCCCTGCGTTAAATGAGT | Diagnostic primer for Flagellin locus |
| F3 | TATTGCTCTGACGCTCAATG | Diagnostic primer for Flagellin locus |
| F6 | ACGGTGATTTCTTTCATTACACAG | Diagnostic primer for Flagellin locus |
| F5 | TTCAGCAGTATCAGCGCCGGT | Diagnostic primer for Flagellin locus |
Abbreviations: E. coli = Escherichia coli; PCR = polymerase chain reaction.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Soulier, A.; Prevosto, C.; Chol, M.; Deban, L.; Cranenburgh, R.M. Engineering a Novel Bivalent Oral Vaccine against Enteric Fever. Int. J. Mol. Sci. 2021, 22, 3287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063287
AMA Style
Soulier A, Prevosto C, Chol M, Deban L, Cranenburgh RM. Engineering a Novel Bivalent Oral Vaccine against Enteric Fever. International Journal of Molecular Sciences. 2021; 22(6):3287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063287
Chicago/Turabian StyleSoulier, Annelise, Claudia Prevosto, Mary Chol, Livija Deban, and Rocky M. Cranenburgh. 2021. "Engineering a Novel Bivalent Oral Vaccine against Enteric Fever" International Journal of Molecular Sciences 22, no. 6: 3287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063287
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.