
Reduced Claudin-12 Expression Predicts Poor Prognosis in Cervical Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
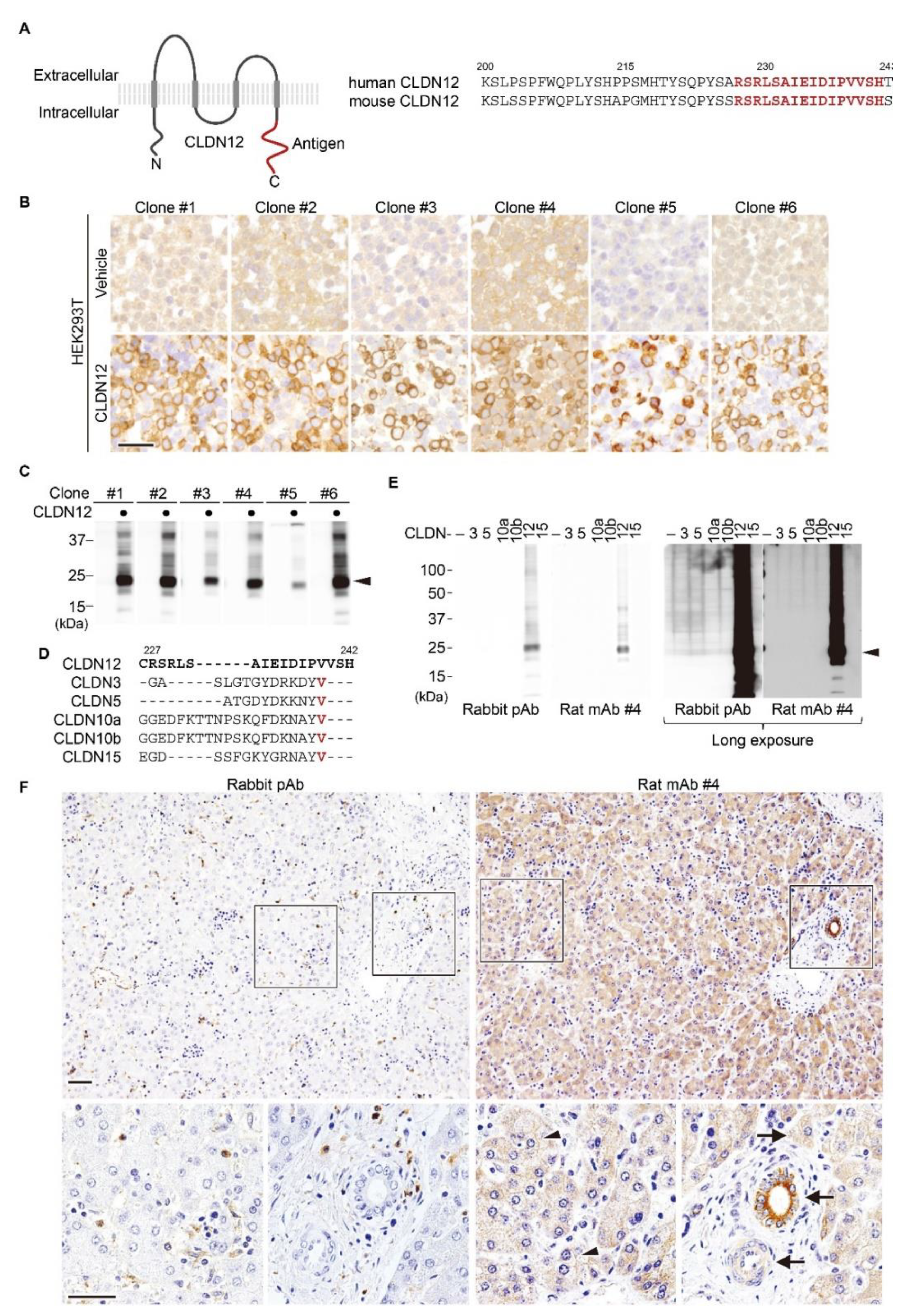
2.1. Establishment of an Anti-Human/Mouse CLDN12 mAb
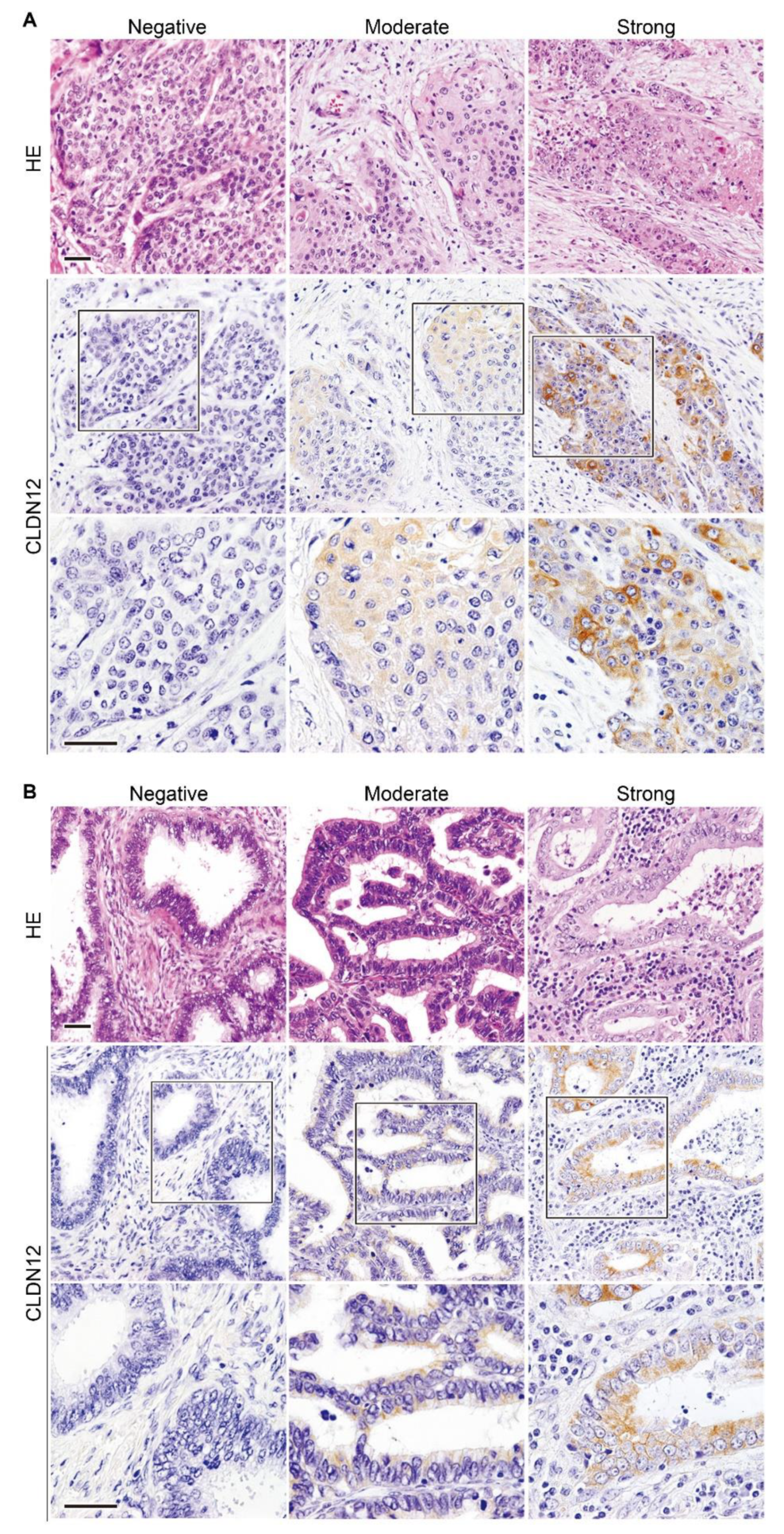
2.2. Expression of CLDN12 Protein in Normal, Premalignant and Malignant Tissues of the Uterine Cervix
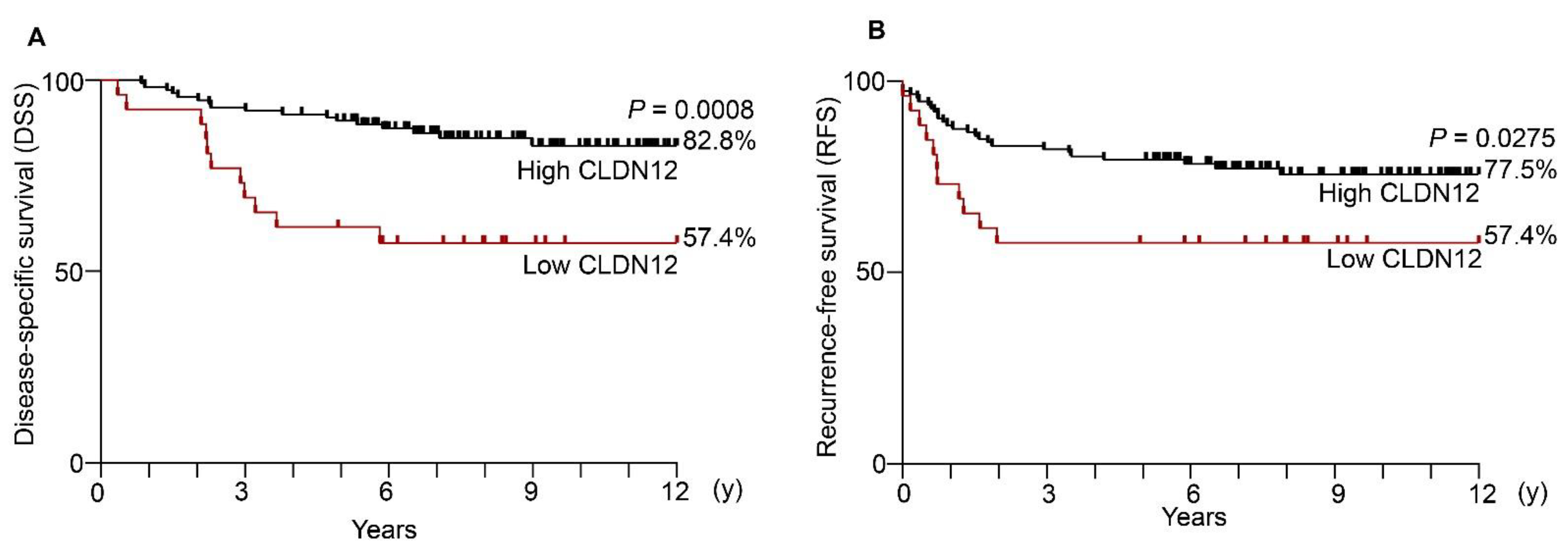
2.3. Low Expression of CLDN12 Correlates with Poor Prognosis and Recurrence in Cervical Cancer
2.4. Reduced CLDN12 Represents an Independent Poor Prognostic Marker for Cervical Cancer
3. Discussion
4. Materials and Methods
4.1. Generation of Antibodies
4.2. Cell Culture, Expression Vectors and Transfection
4.3. Immunoblotting
4.4. Cell Blocks
4.5. Tissue Collection, Immunostaining and Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, C.J.L.M.; Steenbergen, R.D.M. Gynaecological cancer: Novel molecular subtypes of cervical cancer—Potential clinical consequences. Nat. Rev. Clin. Oncol. 2017, 14, 397–398. [Google Scholar] [CrossRef]
- Small, W., Jr.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Schiffman, M.; Wentzensen, N.; Wacholder, S.; Kinney, W.; Gage, J.C.; Castle, P.E. Human papillomavirus testing in the prevention of cervical cancer. J. Natl. Cancer Inst. 2011, 103, 368–383. [Google Scholar] [CrossRef] [Green Version]
- Crosbie, E.J.; Einstein, M.H.; Franceschi, S.; Kitchener, H.C. Human papillomavirus and cervical cancer. Lancet 2013, 382, 889–899. [Google Scholar] [CrossRef]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef]
- Furuse, M.; Tsukita, S. Claudins in occluding junctions of humans and flies. Trends Cell Biol. 2006, 16, 181–188. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Anderson, J.M. Claudins and epithelial paracellular transport. Annu. Rev. Physiol. 2006, 68, 403–429. [Google Scholar] [CrossRef] [PubMed]
- Chiba, H.; Osanai, M.; Murata, M.; Kojima, T.; Sawada, N. Transmembrane proteins of tight junctions. Biochim. Biophys. Acta 2008, 1778, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Tanaka, H.; Tamura, A. The Claudins: From Tight Junctions to Biological Systems. Trends Biochem. Sci. 2019, 44, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Ichikawa-Tomikawa, N.; Kashiwagi, K.; Endo, C.; Tanaka, S.; Sawada, N.; Watabe, T.; Higashi, T.; Chiba, H. Cell adhesion signals regulate the nuclear receptor activity. Proc. Natl. Acad. Sci. USA 2019, 116, 24600–24609. [Google Scholar] [CrossRef]
- Oliveira, S.S.; Morgado-Díaz, J.A. Claudins: Multifunctional players in epithelial tight junctions and their role in cancer. Cell. Mol. Life Sci. 2007, 64, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Valle, B.L.; Morin, P.J. Chapter 13—Claudins in Cancer Biology. In Current Topics in Membranes; Alan, S.L.Y., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 65, pp. 293–333. [Google Scholar]
- Turksen, K.; Troy, T.-C. Junctions gone bad: Claudins and loss of the barrier in cancer. Biochim. Biophys. Acta 2011, 1816, 73–79. [Google Scholar] [CrossRef]
- Osanai, M.; Takasawa, A.; Murata, M.; Sawada, N. Claudins in cancer: Bench to bedside. Pflugers Arch. 2017, 469, 55–67. [Google Scholar] [CrossRef]
- Tabariès, S.; Siegel, P.M. The role of claudins in cancer metastasis. Oncogene 2017, 36, 1176–1190. [Google Scholar] [CrossRef]
- Zeisel, M.B.; Dhawan, P.; Baumert, T.F. Tight junction proteins in gastrointestinal and liver disease. Gut 2019, 68, 547–561. [Google Scholar] [CrossRef]
- Gowrikumar, S.; Singh, A.B.; Dhawan, P. Role of Claudin Proteins in Regulating Cancer Stem Cells and Chemoresistance-Potential Implication in Disease Prognosis and Therapy. Int. J. Mol. Sci. 2019, 21, 53. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.A.; Syed, N.; Therachiyil, L.; Nisar, S.; Hashem, S.; Macha, M.A.; Yadav, S.K.; Krishnankutty, R.; Muralitharan, S.; Al-Naemi, H.; et al. Claudin-1, A Double-Edged Sword in Cancer. Int. J. Mol. Sci. 2020, 21, 569. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Sugimoto, K.; Tanaka, M.; Endo, Y.; Kato, H.; Honda, T.; Furukawa, S.; Nishiyama, H.; Watanabe, T.; Soeda, S.; et al. Prognostic Significance of Aberrant Claudin-6 Expression in Endometrial Cancer. Cancers 2020, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Sugimoto, K.; Kobayashi, M.; Ichikawa-Tomikawa, N.; Kashiwagi, K.; Watanabe, T.; Soeda, S.; Fujimori, K.; Chiba, H. Aberrant claudin-6-adhesion signaling promotes endometrial cancer progression via estrogen receptor α. Mol. Cancer Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günzel, D.; Yu, A.S.L. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [Green Version]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Plain, A.; Pan, W.; O’Neill, D.; Ure, M.; Beggs, M.R.; Farhan, M.; Dimke, H.; Cordat, E.; Alexander, R.T. Claudin-12 Knockout Mice Demonstrate Reduced Proximal Tubule Calcium Permeability. Int. J. Mol. Sci. 2020, 21, 2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network; Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium Pan-cancer analysis of whole genomes. Nature 2020, 578, 82–93. [CrossRef] [Green Version]
- Fujita, H.; Chiba, H.; Yokozaki, H.; Sakai, N.; Sugimoto, K.; Wada, T.; Kojima, T.; Yamashita, T.; Sawada, N. Differential expression and subcellular localization of claudin-7, -8, -12, -13, and -15 along the mouse intestine. J. Histochem. Cytochem. 2006, 54, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Castro Dias, M.; Coisne, C.; Baden, P.; Enzmann, G.; Garrett, L.; Becker, L.; Hölter, S.M.; German Mouse Clinic Consortium; de Angelis, M.H.; Deutsch, U.; et al. Claudin-12 is not required for blood-brain barrier tight junction function. Fluids Barriers CNS 2019, 16, 30. [Google Scholar] [CrossRef]
- Kishiro, Y.; Kagawa, M.; Naito, I.; Sado, Y. A novel method of preparing rat-monoclonal antibody-producing hybridomas by using rat medial iliac lymph node cells. Cell Struct. Funct. 1995, 20, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Satohisa, S.; Chiba, H.; Osanai, M.; Ohno, S.; Kojima, T.; Saito, T.; Sawada, N. Behavior of tight-junction, adherens-junction and cell polarity proteins during HNF-4alpha-induced epithelial polarization. Exp. Cell Res. 2005, 310, 66–78. [Google Scholar] [CrossRef]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial claudin: Claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J. Cell Biol. 1999, 147, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, D.; Kayamori, K.; Sakamoto, K.; Tsuchiya, M.; Ikeda, T.; Harada, H.; Yoda, T.; Watabe, T.; Hara-Yokoyama, M. Intracellular claudin-1 at the invasive front of tongue squamous cell carcinoma is associated with lymph node metastasis. Cancer Sci. 2020, 111, 700–712. [Google Scholar] [CrossRef]
- Zhang, W.-N.; Li, W.; Wang, X.-L.; Hu, Z.; Zhu, D.; Ding, W.-C.; Liu, D.; Li, K.-Z.; Ma, D.; Wang, H. CLDN1 expression in cervical cancer cells is related to tumor invasion and metastasis. Oncotarget 2016, 7, 87449–87461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Miller, C.; Mosher, R.; Zhao, X.; Deeds, J.; Morrissey, M.; Bryant, B.; Yang, D.; Meyer, R.; Cronin, F.; et al. Identification of cervical cancer markers by cDNA and tissue microarrays. Cancer Res. 2003, 63, 1927–1935. [Google Scholar] [PubMed]
- Sobel, G.; Páska, C.; Szabó, I.; Kiss, A.; Kádár, A.; Schaff, Z. Increased expression of claudins in cervical squamous intraepithelial neoplasia and invasive carcinoma. Hum. Pathol. 2005, 36, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Lee, S.-J.; Seo, J.; Song, S.Y.; Ahn, G.; Park, C.-S.; Lee, J.-H.; Kim, B.-G.; Bae, D.-S. Increased expressions of claudin-1 and claudin-7 during the progression of cervical neoplasia. Gynecol. Oncol. 2005, 97, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Kiss, A.; Schaff, Z.; Sobel, G. Claudins as diagnostic and prognostic markers in gynecological cancer. Histol. Histopathol. 2009, 24, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, T.; Takasawa, A.; Murata, M.; Kojima, Y.; Takasawa, K.; Nojima, M.; Aoyama, T.; Hiratsuka, Y.; Ono, Y.; Tanaka, S.; et al. Analysis of the expression and localization of tight junction transmembrane proteins, claudin-1, -4, -7, occludin and JAM-A, in human cervical adenocarcinoma. Histol. Histopathol. 2016, 31, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Benczik, M.; Galamb, Á.; Koiss, R.; Kovács, A.; Járay, B.; Székely, T.; Szekerczés, T.; Schaff, Z.; Sobel, G.; Jeney, C. Claudin-1 as a Biomarker of Cervical Cytology and Histology. Pathol. Oncol. Res. 2016, 22, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Hagen, S.J.; Ang, L.-H.; Zheng, Y.; Karahan, S.N.; Wu, J.; Wang, Y.E.; Caron, T.J.; Gad, A.P.; Muthupalani, S.; Fox, J.G. Loss of Tight Junction Protein Claudin 18 Promotes Progressive Neoplasia Development in Mouse Stomach. Gastroenterology 2018, 155, 1852–1867. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Chimge, N.-O.; Zhou, B.; Flodby, P.; Castaldi, A.; Firth, A.L.; Liu, Y.; Wang, H.; Yang, C.; Marconett, C.N.; et al. CLDN18.1 attenuates malignancy and related signaling pathways of lung adenocarcinoma in vivo and in vitro. Int. J. Cancer 2018, 143, 3169–3180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Flodby, P.; Luo, J.; Castillo, D.R.; Liu, Y.; Yu, F.-X.; McConnell, A.; Varghese, B.; Li, G.; Chimge, N.-O.; et al. Claudin-18-mediated YAP activity regulates lung stem and progenitor cell homeostasis and tumorigenesis. J. Clin. Investig. 2018, 128, 970–984. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Sentani, K.; Tanaka, H.; Yano, T.; Suzuki, K.; Oshima, M.; Yasui, W.; Tamura, A.; Tsukita, S. Deficiency of Stomach-Type Claudin-18 in Mice Induces Gastric Tumor Formation Independent of H pylori Infection. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Remmele, W.; Hildebrand, U.; Hienz, H.A.; Klein, P.J.; Vierbuchen, M.; Behnken, L.J.; Heicke, B.; Scheidt, E. Comparative histological, histochemical, immunohistochemical and biochemical studies on oestrogen receptors, lectin receptors, and Barr bodies in human breast cancer. Virchows Arch. A Pathol. Anat. Histopathol. 1986, 409, 127–147. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (n = 138) | p-Value | ||
|---|---|---|---|---|
| High CLDN12 (n = 112) | Low CLDN12 (n = 26) | |||
| Age | ||||
| <50 | 74 (66%) | 15 (58%) | 0.4964 | |
| ≥50 | 38 (34%) | 11 (42%) | ||
| Histological type | ||||
| Adenocarcinoma | 19 (17%) | 1 (4%) | SCC vs. non-SCC 0.3533 | |
| Adenosquamous carcinoma | 14 (13%) | 4 (15%) | ||
| Squamous cell carcinoma | 73 (65%) | 20 (77%) | ||
| Other | 6 (5%) | 1 (4%) | ||
| FIGO Stage | ||||
| I | 71 (63%) | 13 (50%) | I vs. II/III/IV 0.2653 | |
| II | 37 (33%) | 12 (46%) | ||
| III | 0 (0%) | 0 (0%) | ||
| IV | 4 (4%) | 1 (4%) | ||
| Tumor size | ||||
| 1 | 68 (61%) | 12 (46%) | pT1 vs. pT2/3/4 0.1921 | |
| 2 | 43 (38%) | 13 (50%) | ||
| 3 | 0 (0%) | 0 (0%) | ||
| 4 | 0 (0%) | 1 (4%) | ||
| Unknown | 1 (1%) | 0 (0%) | ||
| Vascular involvement | ||||
| No | 60 (54%) | 12 (46%) | 0.5088 | |
| Yes | 47 (42%) | 13 (50%) | ||
| Unknown | 5 (4%) | 1 (4%) | ||
| Lymphatic involvement | ||||
| No | 51 (46%) | 10 (38%) | 0.6567 | |
| Yes | 57 (51%) | 15 (58%) | ||
| Unknown | 4 (4%) | 1 (4%) | ||
| Lymph node metastasis | ||||
| N0 | 72 (64%) | 17 (65%) | 1.0000 | |
| N1 | 32 (29%) | 7 (27%) | ||
| NX | 8 (7%) | 2 (8%) | ||
| Distant metastasis | ||||
| M0 | 105 (94%) | 25 (96%) | 1.0000 | |
| M1 | 4 (4%) | 0 (0%) | ||
| MX | 3 (3%) | 1 (4%) | ||
| Chemoradiotherapy | ||||
| Chemotherapy | 16 (14%) | 4 (15%) | no vs. yes 0.7971 | |
| Radiotherapy | 29 (26%) | 7 (27%) | ||
| Concurrent chemoradiotherapy | 39 (35%) | 10 (38%) | ||
| Other | 1 (1%) | 0 (0%) | ||
| None | 27 (24%) | 5 (19%) | ||
| Recurrence | ||||
| No | 91 (81%) | 16 (62%) | 0.0385 | |
| Yes | 21 (19%) | 10 (38%) | ||
| Variable | p-Value | Hazard Ratio | 95% CI | |
|---|---|---|---|---|
| CLDN12 | Low | 0.002 | 3.412 | 1.595–7.300 |
| Age | ≥50 | 0.603 | 0.810 | 0.366–1.792 |
| Histological type | SCC | 0.336 | 0.689 | 0.323–1.472 |
| FIGO Stage | ≥2b | <0.001 | 4.866 | 2.277–10.400 |
| Tumor size | >4 cm | <0.001 | 3.945 | 1.862–8.360 |
| Vascular involvement | Yes | 0.005 | 3.509 | 1.465–8.409 |
| Lymphatic involvement | Yes | 0.019 | 2.973 | 1.193–7.408 |
| Lymph node metastasis | Yes | 0.355 | 1.452 | 0.659–3.199 |
| Distant metastasis | Yes | 0.004 | 5.915 | 1.780–19.66 |
| Chemoradiotherapy | Yes | 0.041 | 4.495 | 1.066–18.949 |
| Recurrence | Yes | <0.001 | 19.787 | 7.969–49.135 |
| Variable | p-Value | Hazard Ratio | 95% CI | |
|---|---|---|---|---|
| CLDN12 | Low | 0.029 | 2.615 | 1.101–6.214 |
| FIGO Stage | ≥2b | 0.002 | 4.075 | 1.679–9.891 |
| Recurrence | Yes | <0.001 | 14.852 | 5.847–37.726 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.; Kobayashi, M.; Sugimoto, K.; Endo, Y.; Kojima, M.; Furukawa, S.; Watanabe, T.; Soeda, S.; Hashimoto, Y.; Fujimori, K.; et al. Reduced Claudin-12 Expression Predicts Poor Prognosis in Cervical Cancer. Int. J. Mol. Sci. 2021, 22, 3774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073774
Rahman A, Kobayashi M, Sugimoto K, Endo Y, Kojima M, Furukawa S, Watanabe T, Soeda S, Hashimoto Y, Fujimori K, et al. Reduced Claudin-12 Expression Predicts Poor Prognosis in Cervical Cancer. International Journal of Molecular Sciences. 2021; 22(7):3774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073774
Chicago/Turabian StyleRahman, Abidur, Makoto Kobayashi, Kotaro Sugimoto, Yuta Endo, Manabu Kojima, Shigenori Furukawa, Takafumi Watanabe, Shu Soeda, Yuko Hashimoto, Keiya Fujimori, and et al. 2021. "Reduced Claudin-12 Expression Predicts Poor Prognosis in Cervical Cancer" International Journal of Molecular Sciences 22, no. 7: 3774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073774