
Prolyl 3-Hydroxylase 2 Is a Molecular Player of Angiogenesis

 , , and
, , and 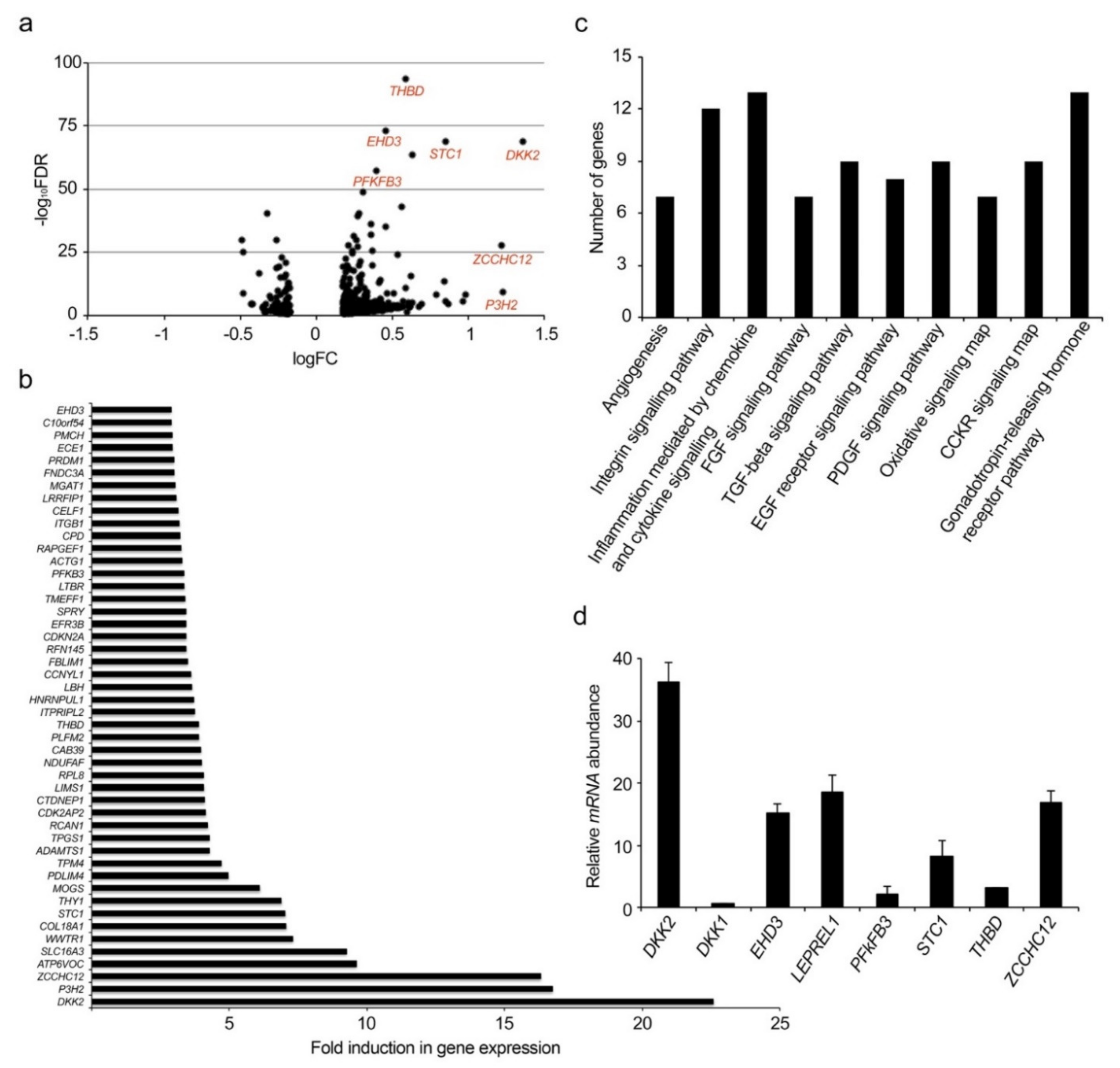{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of New Genes Modulated by VEGF-A
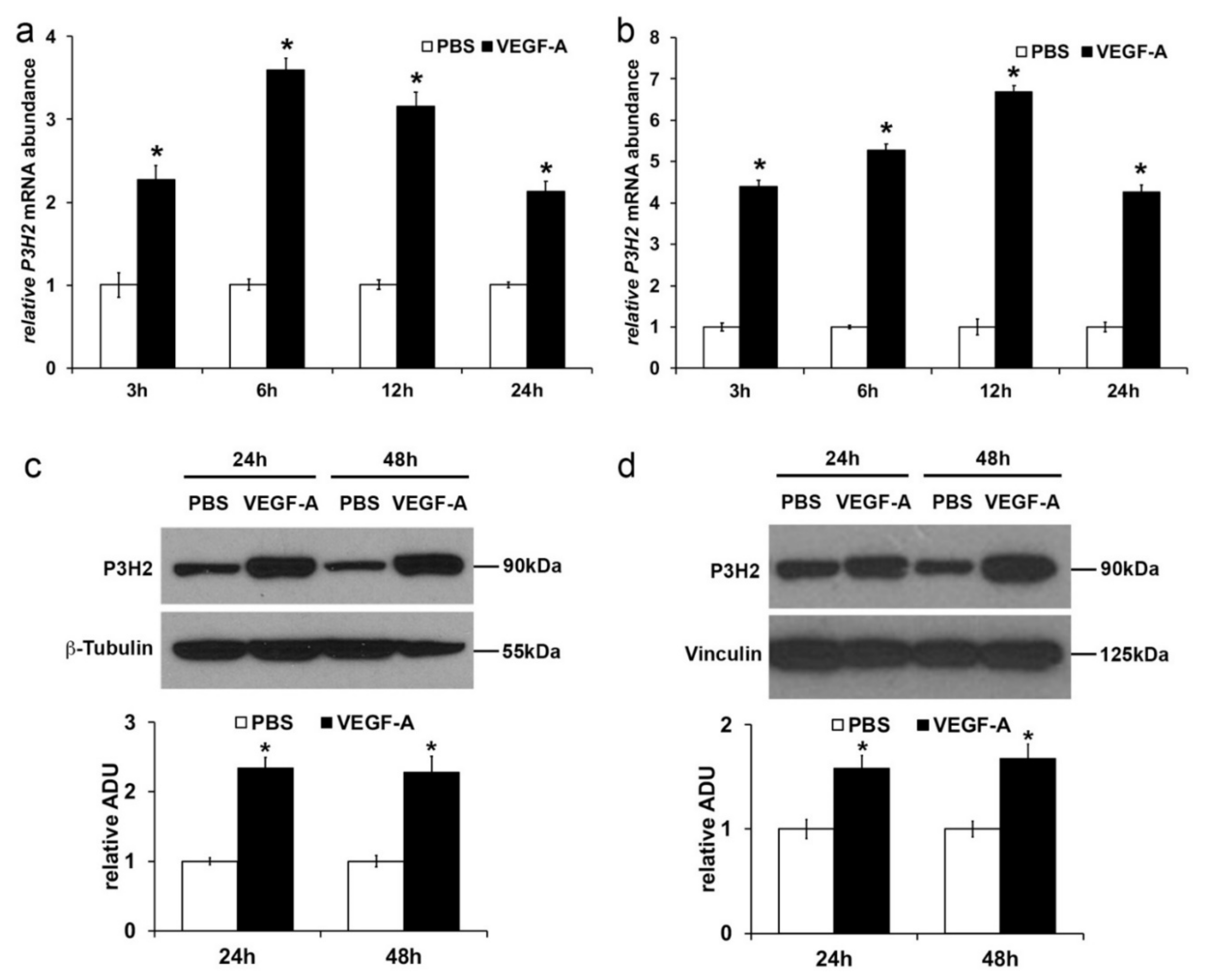
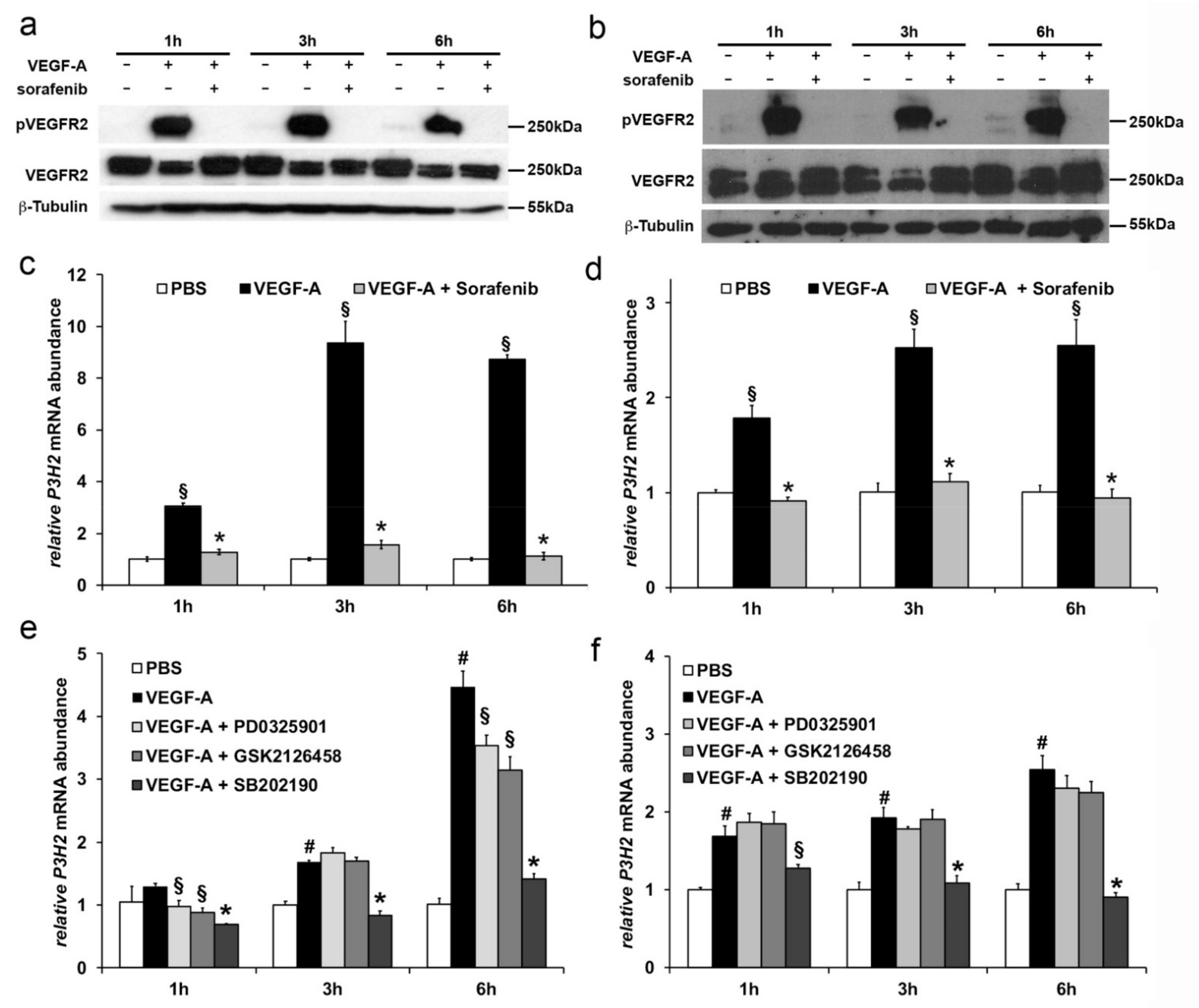
2.2. VEGF-A Stimulates P3H2 Expression in Human Endothelial Cells through VEGFR-2/p38 Signaling
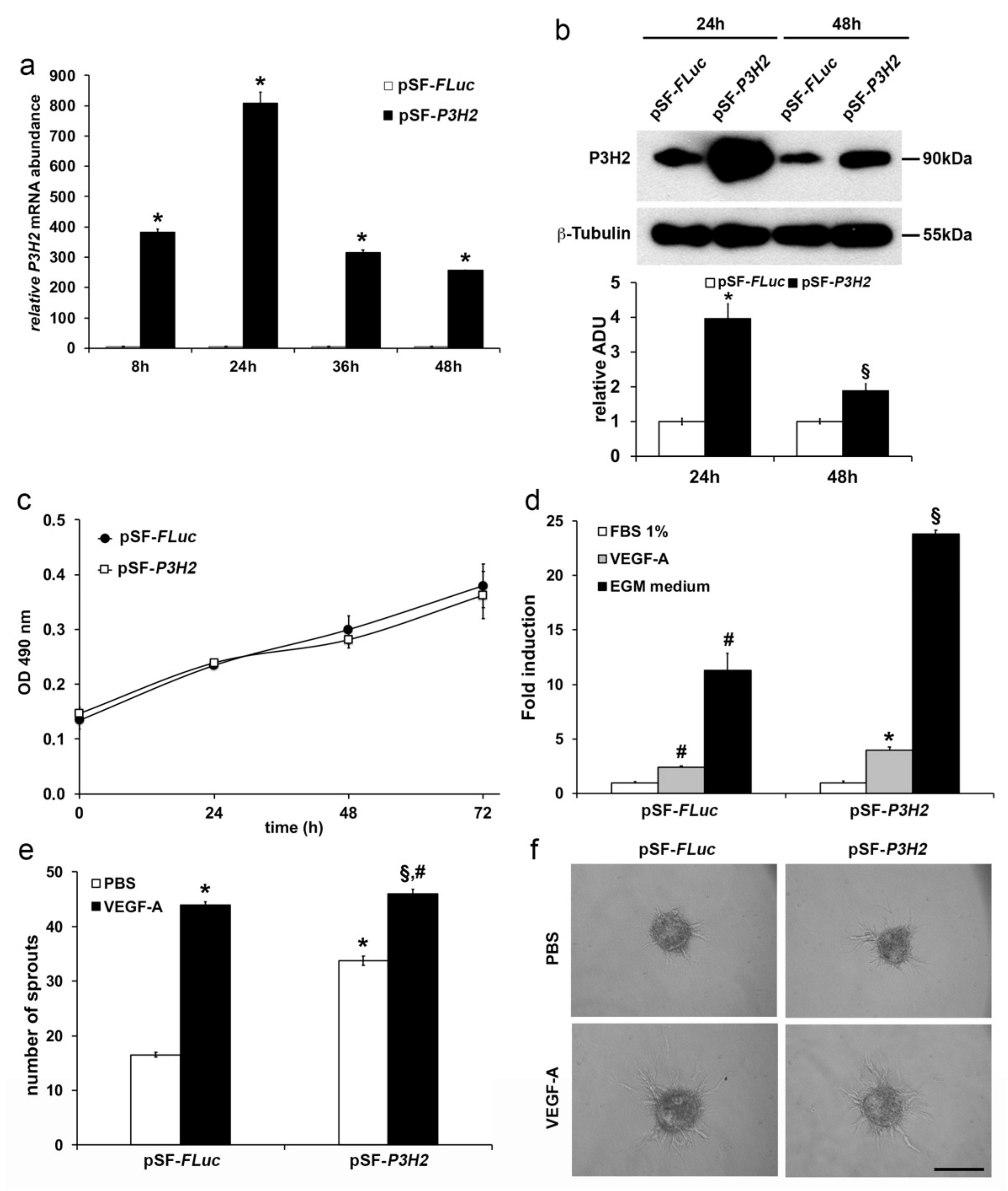
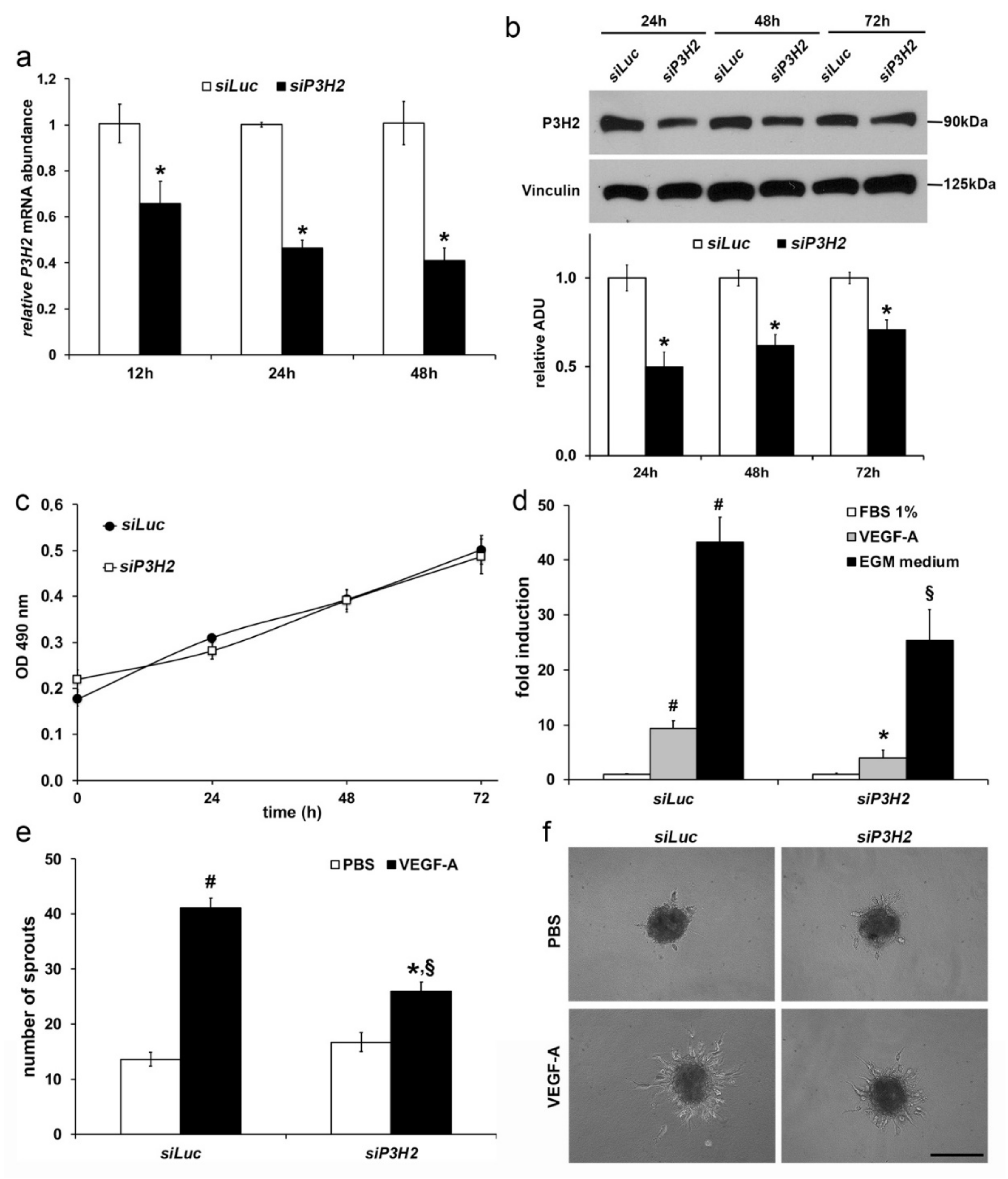
2.3. Modulation of P3H2 Expression Affects Angiogenic Properties of Endothelial Cells
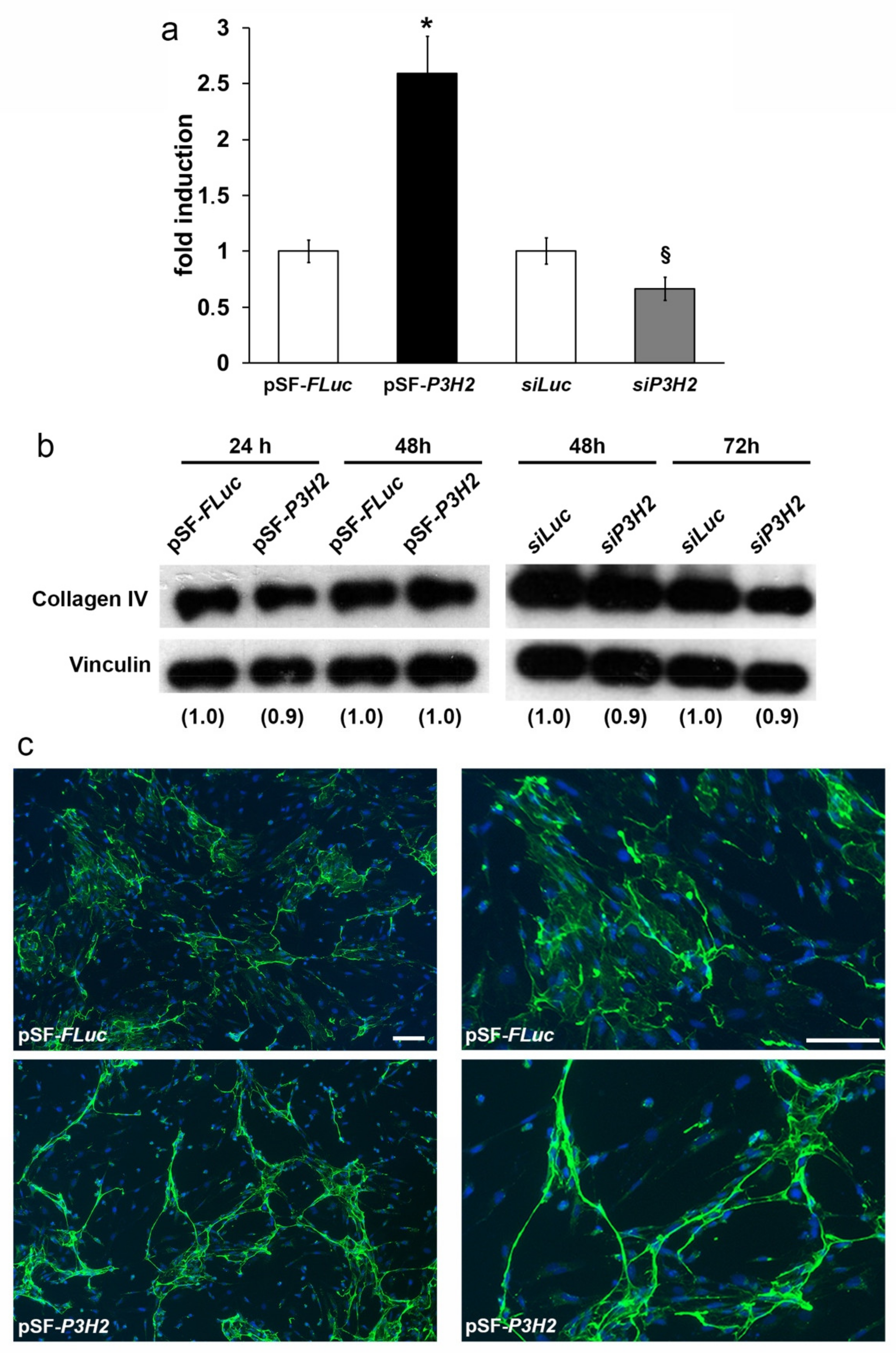
2.4. Overexpression of P3H2 Induces Collagen IV Rearrangement In Vitro
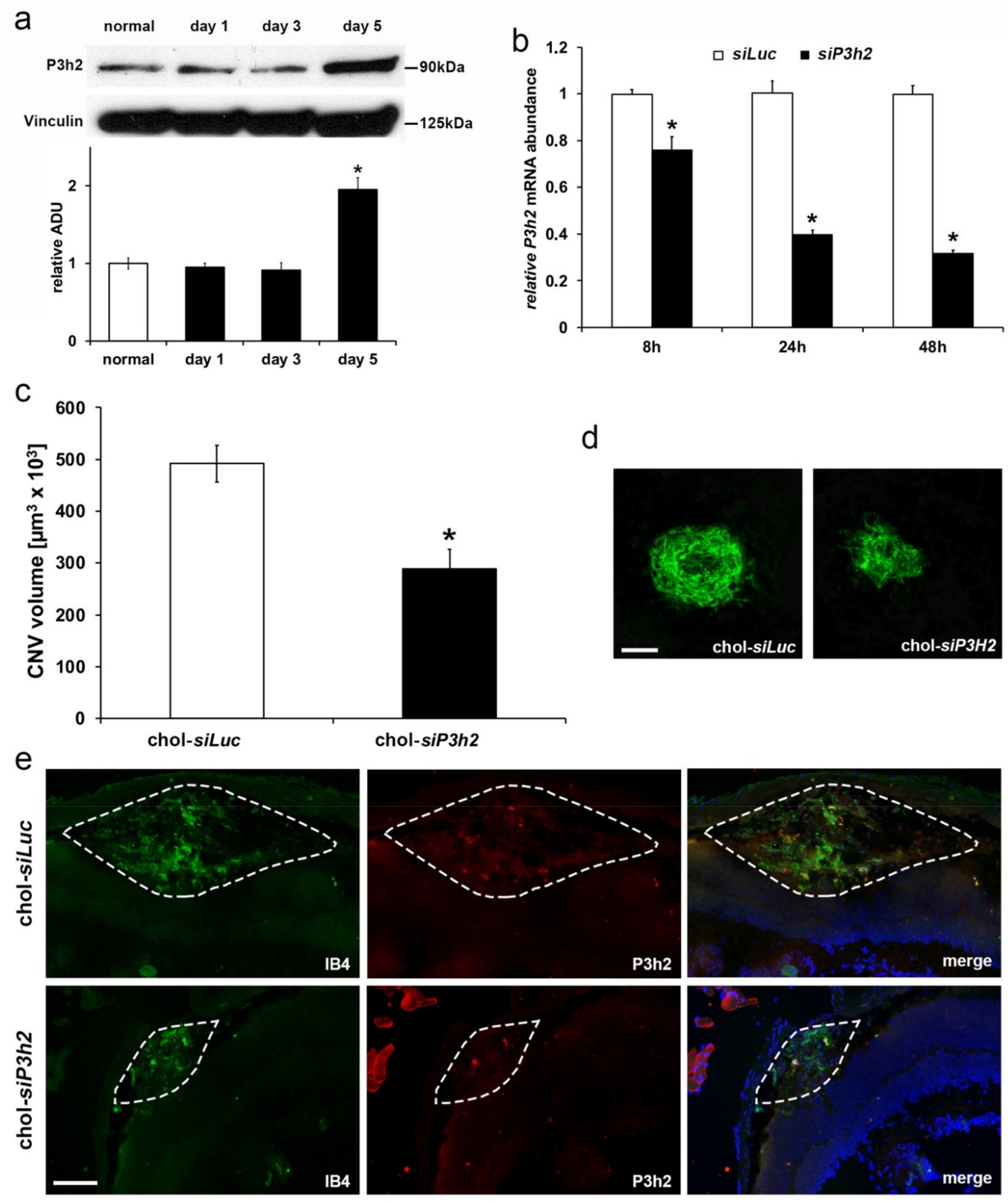
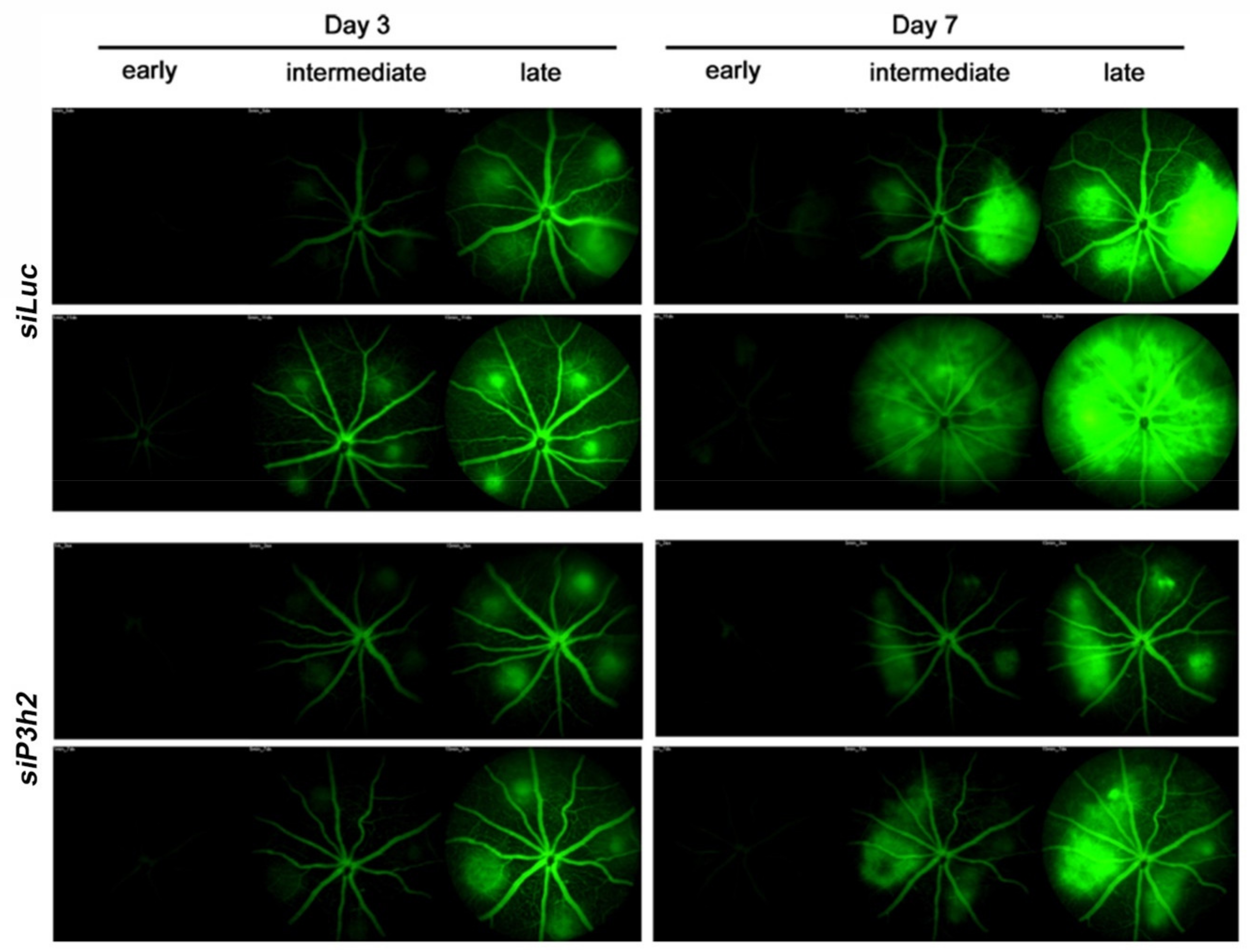
2.5. P3H2 Modulates In Vivo Pathological Angiogenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNAseq
4.3. VEGF-A Stimulation and VEGFR-2 Signaling Inhibition
4.4. Quantitative Real-Time PCR
4.5. Expression Vector
4.6. Transient Transfection
4.7. Western Blot
4.8. Cell Proliferation
4.9. Cell Migration
4.10. Spheroids Sprouting Assay
4.11. Immunofluorescence Analyses
4.12. Animals
4.13. Choroid Neovascularization Model
4.14. Retinal Immunostaining
4.15. Fundus Fluorescein Angiography
4.16. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMD | age-related macular degeneration |
| BM | basement membrane |
| CNV | choroid neovascularization |
| DKK1 | dickkopf WNT signaling pathway inhibitor 1 |
| DKK2 | dickkopf WNT signaling pathway inhibitor 2 |
| ECs | endothelial cells |
| EHD3 | EH-domain containing 3 |
| HMEC | human dermal microvascular endothelial cells |
| HUVEC | human umbilical vein endothelial cells |
| LEPREL1 | Leucine proline-enriched proteoglycan like 1 |
| LV | lentivirus |
| mRPE | mouse retinal pigment epithelial cells |
| P3Hs | prolyl 3-hydroxylases |
| P3H2 | prolyl 3-hydroxylase 2 |
| PFKFB3 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 |
| STC1 | stanniocalcin 1 |
| VEGF-A | vascular endothelial growth factor A |
| VEGFR1 | VEGF receptor 1 |
| VEGFR2 | VEGF receptor 2 |
| ZCCHC12 | Zinc Finger CCHC-Type Containing 12 |
| 3Hyp | 3-hydroxyl-l-proline |
| 4Hyp | 4-hydroxyl-l-proline |
References
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Koch, S.; Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb. Perspect. Med. 2012, 2, a006502. [Google Scholar] [CrossRef] [PubMed]
- Tugues, S.; Koch, S.; Gualandi, L.; Li, X.; Claesson-Welsh, L. Vascular endothelial growth factors and receptors: Anti-angiogenic therapy in the treatment of cancer. Mol. Asp. Med. 2011, 32, 88–111. [Google Scholar] [CrossRef] [PubMed]
- Tarallo, V.; De Falco, S. The vascular endothelial growth factors and receptors family: Up to now the only target for anti-angiogenesis therapy. Int. J. Biochem. Cell Biol. 2015, 64, 185–189. [Google Scholar] [CrossRef] [PubMed]
- De Falco, S. Antiangiogenesis therapy: An update after the first decade. Korean J. Intern. Med. 2014, 29, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lux, A.; Llacer, H.; Heussen, F.M.; Joussen, A.M. Non-responders to bevacizumab (Avastin) therapy of choroidal neovascular lesions. Br. J. Ophthalmol. 2007, 91, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Sennino, B.; McDonald, D.M. Controlling escape from angiogenesis inhibitors. Nat. Rev. Cancer 2012, 12, 699–709. [Google Scholar] [CrossRef]
- Falavarjani, K.G.; Nguyen, Q.D. Adverse events and complications associated with intravitreal injection of anti-VEGF agents: A review of literature. Eye 2013, 27, 787–794. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.T.; Gorn, M.; Smith, K.; Graham, A.P.; Lau, K.K.; Bicknell, R. Transcriptional profiling of human microvascular endothelial cells in the proliferative and quiescent state using cDNA arrays. Angiogenesis 1999, 3, 211–219. [Google Scholar] [CrossRef]
- Abe, M.; Sato, Y. cDNA microarray analysis of the gene expression profile of VEGF-activated human umbilical vein endothelial cells. Angiogenesis 2001, 4, 289–298. [Google Scholar] [CrossRef]
- Jih, Y.J.; Lien, W.H.; Tsai, W.C.; Yang, G.W.; Li, C.; Wu, L.W. Distinct regulation of genes by bFGF and VEGF-A in endothelial cells. Angiogenesis 2001, 4, 313–321. [Google Scholar] [CrossRef]
- Vasile, E.; Tomita, Y.; Brown, L.F.; Kocher, O.; Dvorak, H.F. Differential expression of thymosin beta-10 by early passage and senescent vascular endothelium is modulated by VPF/VEGF: Evidence for senescent endothelial cells in vivo at sites of atherosclerosis. FASEB J. 2001, 15, 458–466. [Google Scholar] [CrossRef] [Green Version]
- Weston, G.C.; Haviv, I.; Rogers, P.A. Microarray analysis of VEGF-responsive genes in myometrial endothelial cells. Mol. Hum. Reprod. 2002, 8, 855–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Toy, K.; Ingle, G.; Zlot, C.; Williams, P.M.; Fuh, G.; Li, B.; de Vos, A.; Gerritsen, M.E. Vascular endothelial growth factor-induced genes in human umbilical vein endothelial cells: Relative roles of KDR and Flt-1 receptors. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1797–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autiero, M.; Waltenberger, J.; Communi, D.; Kranz, A.; Moons, L.; Lambrechts, D.; Kroll, J.; Plaisance, S.; De Mol, M.; Bono, F.; et al. Role of PlGF in the intra- and intermolecular cross talk between the VEGF receptors Flt1 and Flk1. Nat. Med. 2003, 9, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Tiainen, P.; Pasanen, A.; Sormunen, R.; Myllyharju, J. Characterization of recombinant human prolyl 3-hydroxylase isoenzyme 2, an enzyme modifying the basement membrane Collagen IV. J. Biol. Chem. 2008, 283, 19432–19439. [Google Scholar] [CrossRef] [Green Version]
- Jarnum, S.; Kjellman, C.; Darabi, A.; Nilsson, I.; Edvardsen, K.; Aman, P. LEPREL1, a novel ER and Golgi resident member of the Leprecan family. Biochem. Biophys. Res. Commun. 2004, 317, 342–351. [Google Scholar] [CrossRef]
- Hudson, D.M.; Eyre, D.R. Collagen prolyl 3-hydroxylation: A major role for a minor post-translational modification? Connect. Tissue Res. 2013, 54, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenhove-McCarthy, D.J.; McCarthy, K.J. Molecular characterization of a novel basement membrane-associated proteoglycan, leprecan. J. Biol. Chem. 1999, 274, 25004–25017. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.C.; Sugihara, T.; Yoshida, A.; Nomura, H.; Wadhwa, R. Gros1, a potential growth suppressor on chromosome 1: Its identity to basement membrane-associated proteoglycan, leprecan. Oncogene 2000, 19, 3576–3583. [Google Scholar] [CrossRef] [Green Version]
- Kefalides, N.A. Basement membranes: Structural and biosynthetic considerations. J. Investig. Dermatol. 1975, 65, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, T.; Vega-Montoto, L.; Zimmerman, L.J.; Tabb, D.L.; Hudson, B.G.; Vanacore, R.M. Comprehensive Characterization of Glycosylation and Hydroxylation of Basement Membrane Collagen IV by High-Resolution Mass Spectrometry. J. Proteome Res. 2016, 15, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, W.C.; Herman, I.M. Extracellular matrix modulation of endothelial cell shape and motility following injury in vitro. J. Cell Sci. 1985, 73, 19–32. [Google Scholar] [PubMed]
- Herbst, T.J.; McCarthy, J.B.; Tsilibary, E.C.; Furcht, L.T. Differential effects of laminin, intact type IV collagen, and specific domains of type IV collagen on endothelial cell adhesion and migration. J. Cell Biol. 1988, 106, 1365–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madri, J.A.; Pratt, B.M.; Yannariello-Brown, J. Matrix-driven cell size change modulates aortic endothelial cell proliferation and sheet migration. Am. J. Pathol. 1988, 132, 18–27. [Google Scholar]
- Madri, J.A.; Williams, S.K. Capillary endothelial cell cultures: Phenotypic modulation by matrix components. J. Cell Biol. 1983, 97, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Kleinman, H.K.; Martin, G.R.; Lawley, T.J. Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. J. Cell Biol. 1988, 107, 1589–1598. [Google Scholar] [CrossRef]
- Nicosia, R.F.; Madri, J.A. The microvascular extracellular matrix. Developmental changes during angiogenesis in the aortic ring-plasma clot model. Am. J. Pathol. 1987, 128, 78–90. [Google Scholar]
- Bonanno, E.; Iurlaro, M.; Madri, J.A.; Nicosia, R.F. Type IV collagen modulates angiogenesis and neovessel survival in the rat aorta model. In Vitro Cell Dev. Biol. Anim. 2000, 36, 336–340. [Google Scholar] [CrossRef]
- Park, H.; Jung, H.Y.; Choi, H.J.; Kim, D.Y.; Yoo, J.Y.; Yun, C.O.; Min, J.K.; Kim, Y.M.; Kwon, Y.G. Distinct roles of DKK1 and DKK2 in tumor angiogenesis. Angiogenesis 2014, 17, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Law, A.Y.; Wong, C.K. Stanniocalcin-1 and -2 promote angiogenic sprouting in HUVECs via VEGF/VEGFR2 and angiopoietin signaling pathways. Mol. Cell Endocrinol. 2013, 374, 73–81. [Google Scholar] [CrossRef] [PubMed]
- De Bock, K.; Georgiadou, M.; Schoors, S.; Kuchnio, A.; Wong, B.W.; Cantelmo, A.R.; Quaegebeur, A.; Ghesquiere, B.; Cauwenberghs, S.; Eelen, G.; et al. Role of PFKFB3-driven glycolysis in vessel sprouting. Cell 2013, 154, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Cho, G.; Lim, Y.; Zand, D.; Golden, J.A. Sizn1 is a novel protein that functions as a transcriptional coactivator of bone morphogenic protein signaling. Mol. Cell Biol. 2008, 28, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, U.; Smith, J.S.; Tachibana, I.; Ueki, K.; Lee, H.K.; Ramaswamy, S.; Wu, Q.; Mohrenweiser, H.W.; Jenkins, R.B.; Louis, D.N. EHD2, EHD3, and EHD4 encode novel members of a highly conserved family of EH domain-containing proteins. Genomics 2000, 63, 255–262. [Google Scholar] [CrossRef]
- Gotink, K.J.; Verheul, H.M. Anti-angiogenic tyrosine kinase inhibitors: What is their mechanism of action? Angiogenesis 2010, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Simons, M.; Alitalo, K.; Annex, B.H.; Augustin, H.G.; Beam, C.; Berk, B.C.; Byzova, T.; Carmeliet, P.; Chilian, W.; Cooke, J.P.; et al. American Heart Association Council on Basic Cardiovascular, S.; Council on Cardiovascular, S.; Anesthesia, State-of-the-Art Methods for Evaluation of Angiogenesis and Tissue Vascularization: A Scientific Statement From the American Heart Association. Circ. Res. 2015, 116, e99–e132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambati, J.; Fowler, B.J. Mechanisms of age-related macular degeneration. Neuron 2012, 75, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Muranaka, K.; Yanagi, Y.; Tamaki, Y.; Takahashi, H.; Usui, T.; Ohashi, K.; Matsuoka, H.; Senda, T. Suppression of laser-induced choroidal neovascularization by oral administration of SA3443 in mice. FEBS Lett. 2005, 579, 6084–6088. [Google Scholar] [CrossRef] [Green Version]
- Kleinman, M.E.; Yamada, K.; Takeda, A.; Chandrasekaran, V.; Nozaki, M.; Baffi, J.Z.; Albuquerque, R.J.; Yamasaki, S.; Itaya, M.; Pan, Y.; et al. Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature 2008, 452, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Masson-Gadais, B.; Houle, F.; Laferriere, J.; Huot, J. Integrin alphavbeta3, requirement for VEGFR2-mediated activation of SAPK2/p38 and for Hsp90-dependent phosphorylation of focal adhesion kinase in endothelial cells activated by VEGF. Cell Stress Chaperones 2003, 8, 37–52. [Google Scholar] [CrossRef]
- Mahabeleshwar, G.H.; Feng, W.; Phillips, D.R.; Byzova, T.V. Integrin signaling is critical for pathological angiogenesis. J. Exp. Med. 2006, 203, 2495–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, K.M.; Mei, R. Basement Membrane Type IV Collagen and Laminin: An Overview of Their Biology and Value as Fibrosis Biomarkers of Liver Disease. Anat. Rec. 2017, 300, 1371–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozzi, A.; Yurchenco, P.D.; Iozzo, R.V. The nature and biology of basement membranes. Matrix Biol. 2017, 57–58, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedchenko, V.; Zent, R.; Hudson, B.G. Alpha(v)beta3 and alpha(v)beta5 integrins bind both the proximal RGD site and non-RGD motifs within noncollagenous (NC1) domain of the alpha3 chain of type IV collagen: Implication for the mechanism of endothelia cell adhesion. J. Biol. Chem. 2004, 279, 2772–2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.E.; Senger, D.R. Endothelial extracellular matrix: Biosynthesis, remodeling, and functions during vascular morphogenesis and neovessel stabilization. Circ. Res. 2005, 97, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- De Smet, F.; Segura, I.; De Bock, K.; Hohensinner, P.J.; Carmeliet, P. Mechanisms of vessel branching: Filopodia on endothelial tip cells lead the way. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Mundel, T.M.; Kalluri, R. Type IV collagen-derived angiogenesis inhibitors. Microvasc. Res. 2007, 74, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.J.; Farnand, A.W.; Traeger, G.R.; Weis, M.A.; Eyre, D.R. A role for prolyl 3-hydroxylase 2 in post-translational modification of fibril-forming collagens. J. Biol. Chem. 2011, 286, 30662–30669. [Google Scholar] [CrossRef] [Green Version]
- Kivirikko, K.I.; Ryhanen, L.; Anttinen, H.; Bornstein, P.; Prockop, D.J. Further hydroxylation of lysyl residues in collagen by protocollagen lysyl hydroxylase in vitro. Biochemistry 1973, 12, 4966–4971. [Google Scholar] [CrossRef]
- Risteli, J.; Kivirikko, K.I. Activities of prolyl hydroxylase, lysyl hydroxylase, collagen galactosyltransferase and collagen glucosyltransferase in the liver of rats with hepatic injury. Biochem. J. 1974, 144, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myllyla, R.; Alitalo, K.; Vaheri, A.; Kivirikko, K.I. Regulation of collagen post-translational modification in transformed human and chick-embryo cells. Biochem. J. 1981, 196, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, K.; Hayashi, T.; Peyton, D.H.; Bachinger, H.P. The peptides acetyl-(Gly-3(S)Hyp-4(R)Hyp)10-NH2 and acetyl-(Gly-Pro-3(S)Hyp)10-NH2 do not form a collagen triple helix. J. Biol. Chem. 2004, 279, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.L.; Bretscher, L.E.; Guzei, I.A.; Raines, R.T. Effect of 3-hydroxyproline residues on collagen stability. J. Am. Chem. Soc. 2003, 125, 6422–6427. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Mizuno, K.; Bachinger, H.P. The crystal structure of a collagen-like polypeptide with 3(S)-hydroxyproline residues in the Xaa position forms a standard 7/2 collagen triple helix. J. Biol. Chem. 2006, 281, 27566–27574. [Google Scholar] [CrossRef] [Green Version]
- Mordechai, S.; Gradstein, L.; Pasanen, A.; Ofir, R.; El Amour, K.; Levy, J.; Belfair, N.; Lifshitz, T.; Joshua, S.; Narkis, G.; et al. High myopia caused by a mutation in LEPREL1, encoding prolyl 3-hydroxylase 2. Am. J. Hum. Genet. 2011, 89, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Tong, P.; Peng, Y.; Wang, T.; Liu, Y.; Chen, J.; Li, Y.; Tian, Q.; Hu, Y.; Zheng, Y.; et al. Homozygous loss-of-function mutation of the LEPREL1 gene causes severe non-syndromic high myopia with early-onset cataract. Clin. Genet. 2014, 86, 575–579. [Google Scholar] [CrossRef]
- Hudson, D.M.; Joeng, K.S.; Werther, R.; Rajagopal, A.; Weis, M.; Lee, B.H.; Eyre, D.R. Post-translationally abnormal collagens of prolyl 3-hydroxylase-2 null mice offer a pathobiological mechanism for the high myopia linked to human LEPREL1 mutations. J. Biol. Chem. 2015, 290, 8613–8622. [Google Scholar] [CrossRef] [Green Version]
- Pokidysheva, E.; Boudko, S.; Vranka, J.; Zientek, K.; Maddox, K.; Moser, M.; Fassler, R.; Ware, J.; Bachinger, H.P. Biological role of prolyl 3-hydroxylation in type IV collagen. Proc. Natl. Acad. Sci. USA 2014, 111, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Kuo, D.S.; Labelle-Dumais, C.; Gould, D.B. COL4A1 and COL4A2 mutations and disease: Insights into pathogenic mechanisms and potential therapeutic targets. Hum. Mol. Genet. 2012, 21, R97–R110. [Google Scholar] [CrossRef] [Green Version]
- Van Agtmael, T.; Schlotzer-Schrehardt, U.; McKie, L.; Brownstein, D.G.; Lee, A.W.; Cross, S.H.; Sado, Y.; Mullins, J.J.; Poschl, E.; Jackson, I.J. Dominant mutations of Col4a1 result in basement membrane defects which lead to anterior segment dysgenesis and glomerulopathy. Hum. Mol. Genet. 2005, 14, 3161–3168. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.B.; Marchant, J.K.; Savinova, O.V.; Smith, R.S.; John, S.W. Col4a1 mutation causes endoplasmic reticulum stress and genetically modifiable ocular dysgenesis. Hum. Mol. Genet. 2007, 16, 798–807. [Google Scholar] [CrossRef]
- Jeanne, M.; Jorgensen, J.; Gould, D.B. Molecular and Genetic Analyses of Collagen Type IV Mutant Mouse Models of Spontaneous Intracerebral Hemorrhage Identify Mechanisms for Stroke Prevention. Circulation 2015, 131, 1555–1565. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Miyamura, N.; Ninomiya, Y.; Handa, J.T. Distribution of the Collagen IV isoforms in human Bruch’s membrane. Br. J. Ophthalmol. 2003, 87, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Dilworth, D.J.; Weng, Y.C.; Gould, D.B. Developmental distribution of Collagen IV isoforms and relevance to ocular diseases. Matrix Biol. 2009, 28, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavi, M.V.; Mao, M.; Pawlikowski, B.T.; Kvezereli, M.; Duncan, J.L.; Libby, R.T.; John, S.W.; Gould, D.B. Col4a1 mutations cause progressive retinal neovascular defects and retinopathy. Sci. Rep. 2016, 6, 18602. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pignata, P.; Apicella, I.; Cicatiello, V.; Puglisi, C.; Magliacane Trotta, S.; Sanges, R.; Tarallo, V.; De Falco, S. Prolyl 3-Hydroxylase 2 Is a Molecular Player of Angiogenesis. Int. J. Mol. Sci. 2021, 22, 3896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083896
Pignata P, Apicella I, Cicatiello V, Puglisi C, Magliacane Trotta S, Sanges R, Tarallo V, De Falco S. Prolyl 3-Hydroxylase 2 Is a Molecular Player of Angiogenesis. International Journal of Molecular Sciences. 2021; 22(8):3896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083896
Chicago/Turabian StylePignata, Paola, Ivana Apicella, Valeria Cicatiello, Caterina Puglisi, Sara Magliacane Trotta, Remo Sanges, Valeria Tarallo, and Sandro De Falco. 2021. "Prolyl 3-Hydroxylase 2 Is a Molecular Player of Angiogenesis" International Journal of Molecular Sciences 22, no. 8: 3896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083896