
Physiologically Active Molecules and Functional Properties of Soybeans in Human Health—A Current Perspective


Abstract
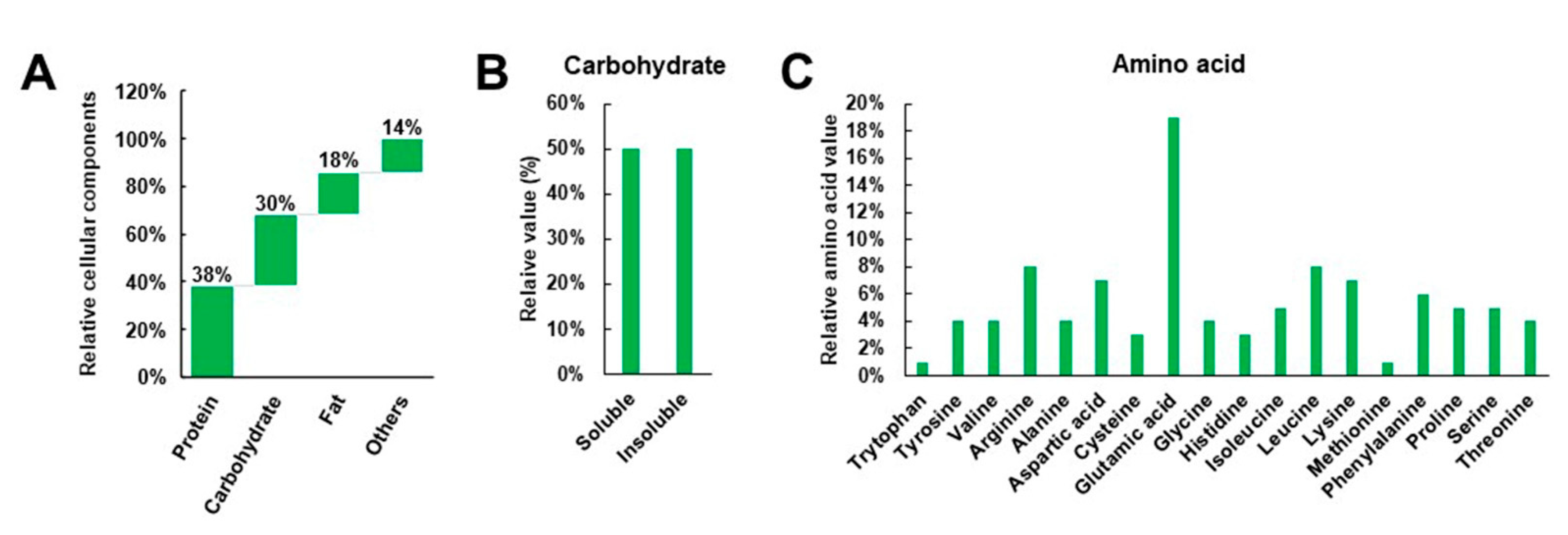
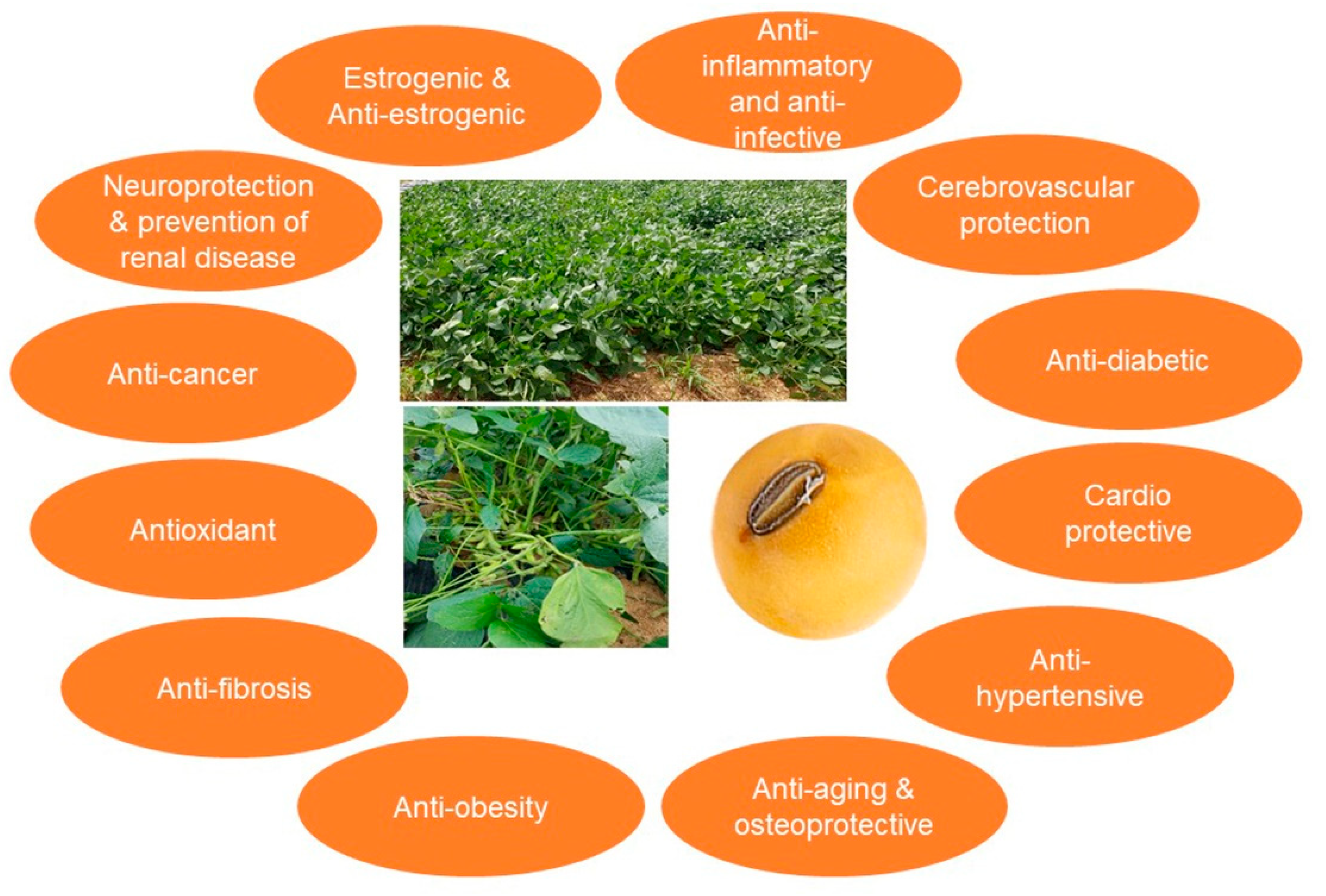
:1. Introduction
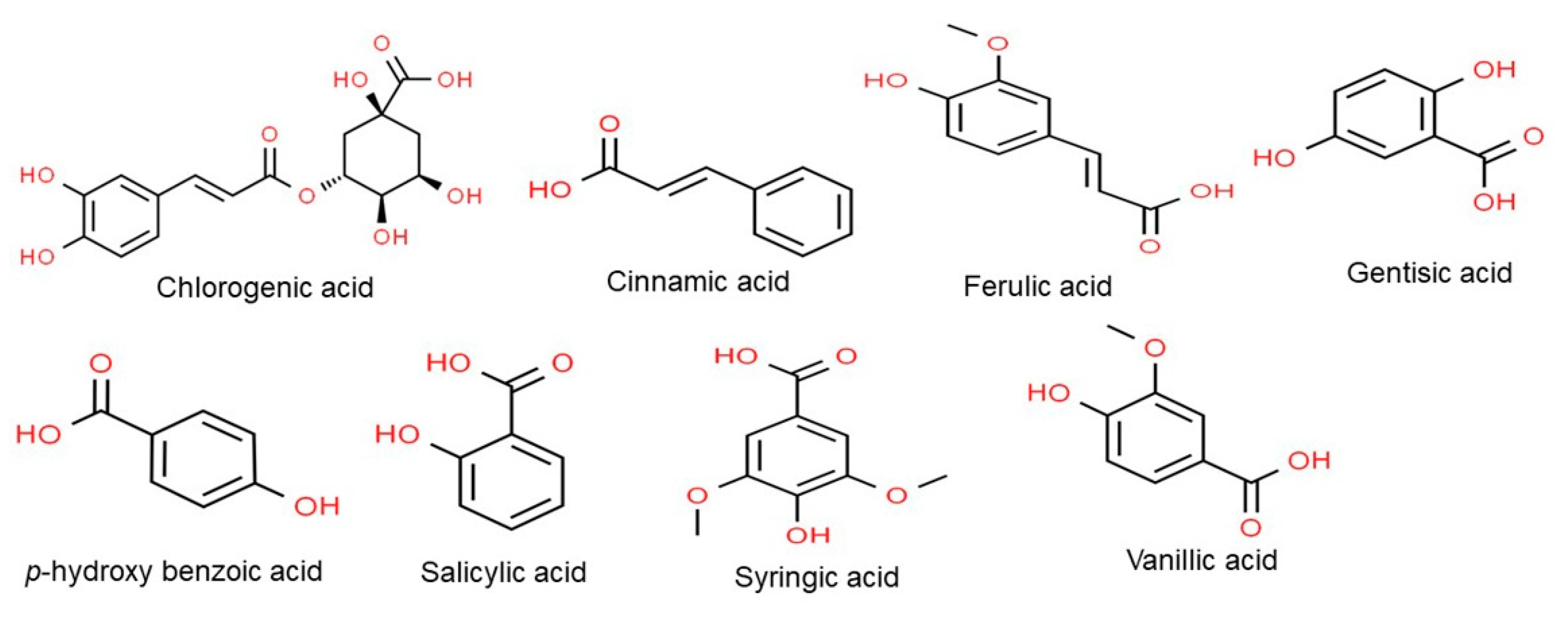
2. Phenolic Compounds
2.1. Phenolic Acids
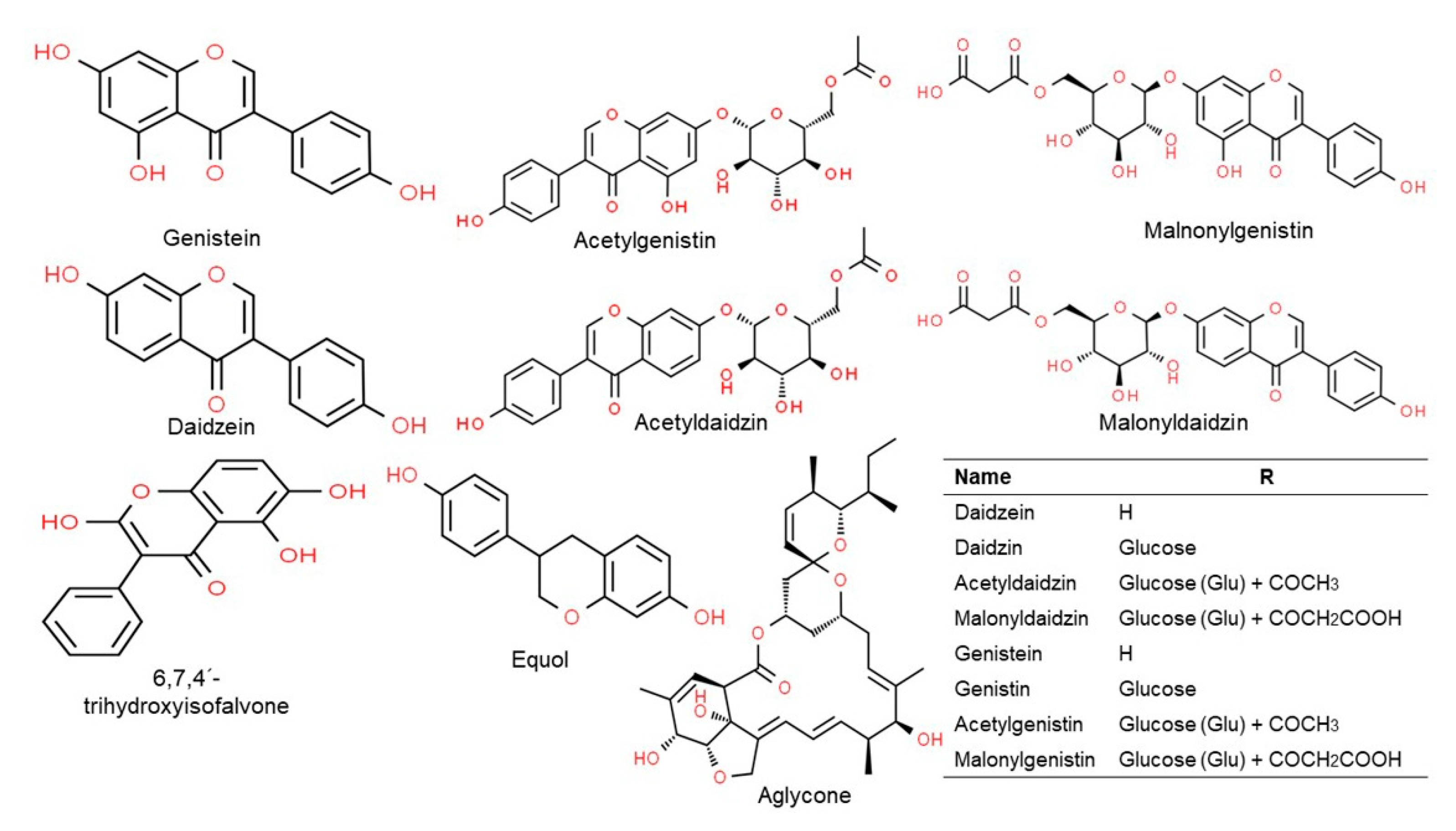
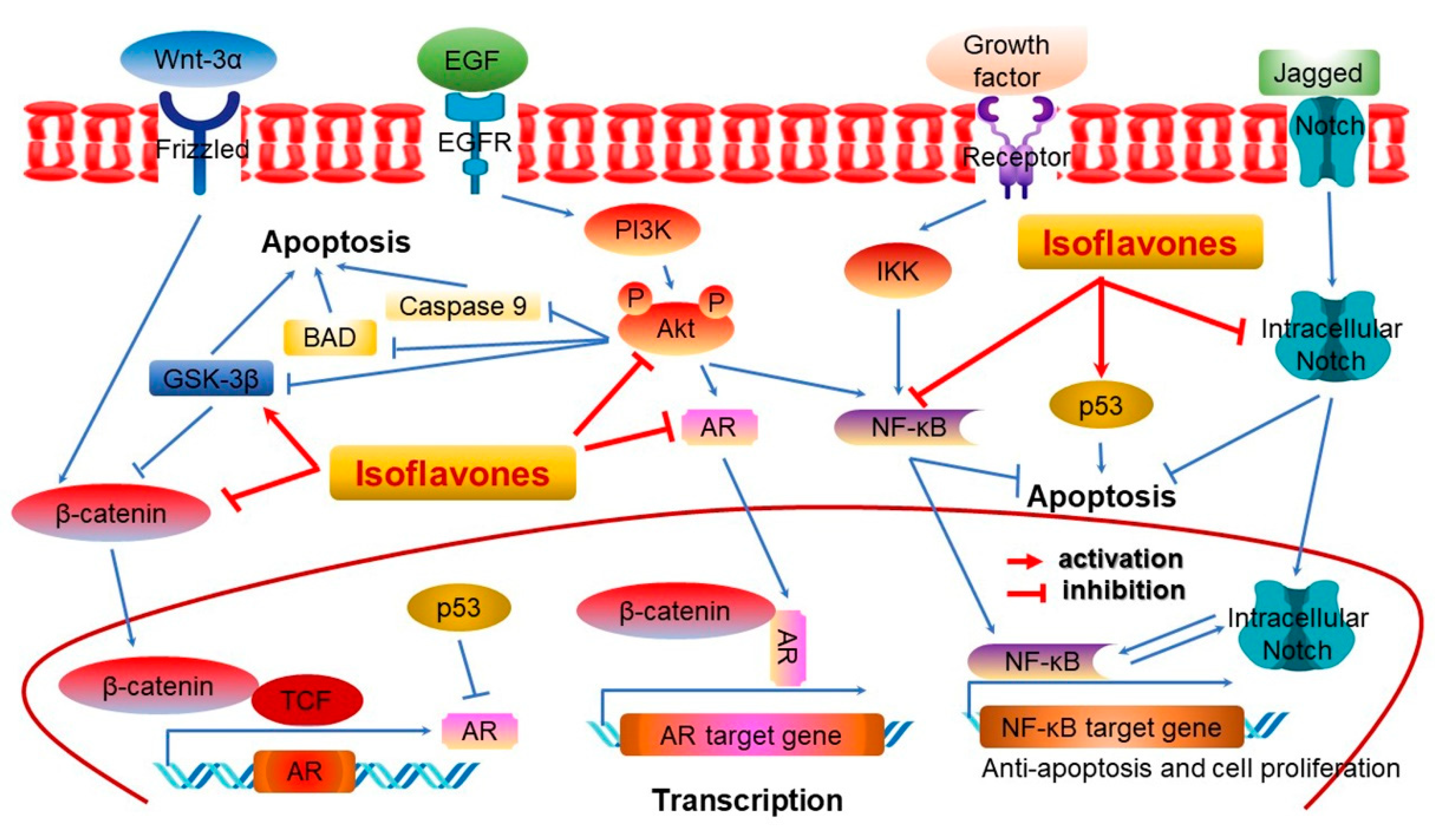
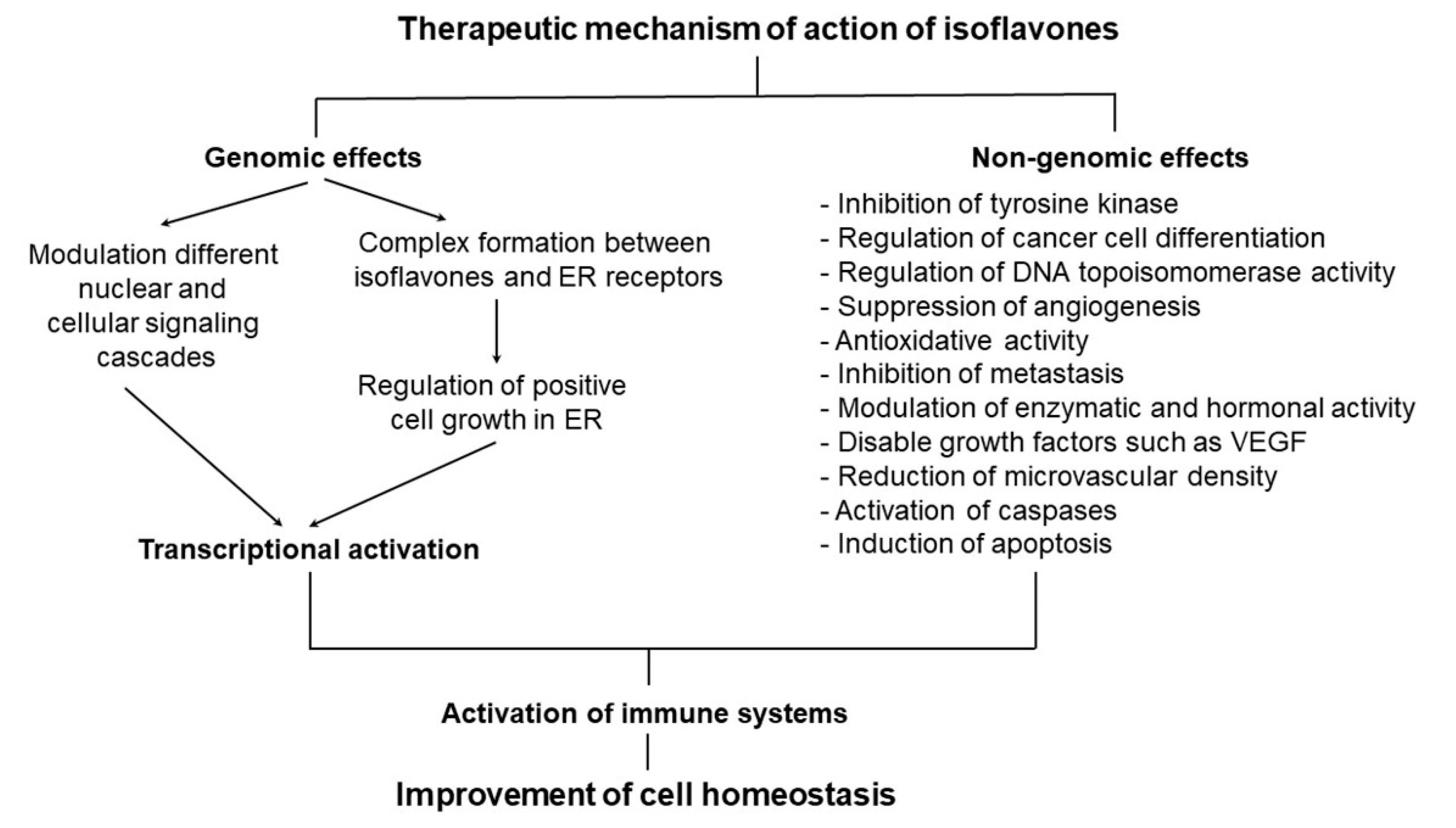
2.2. Isoflavones
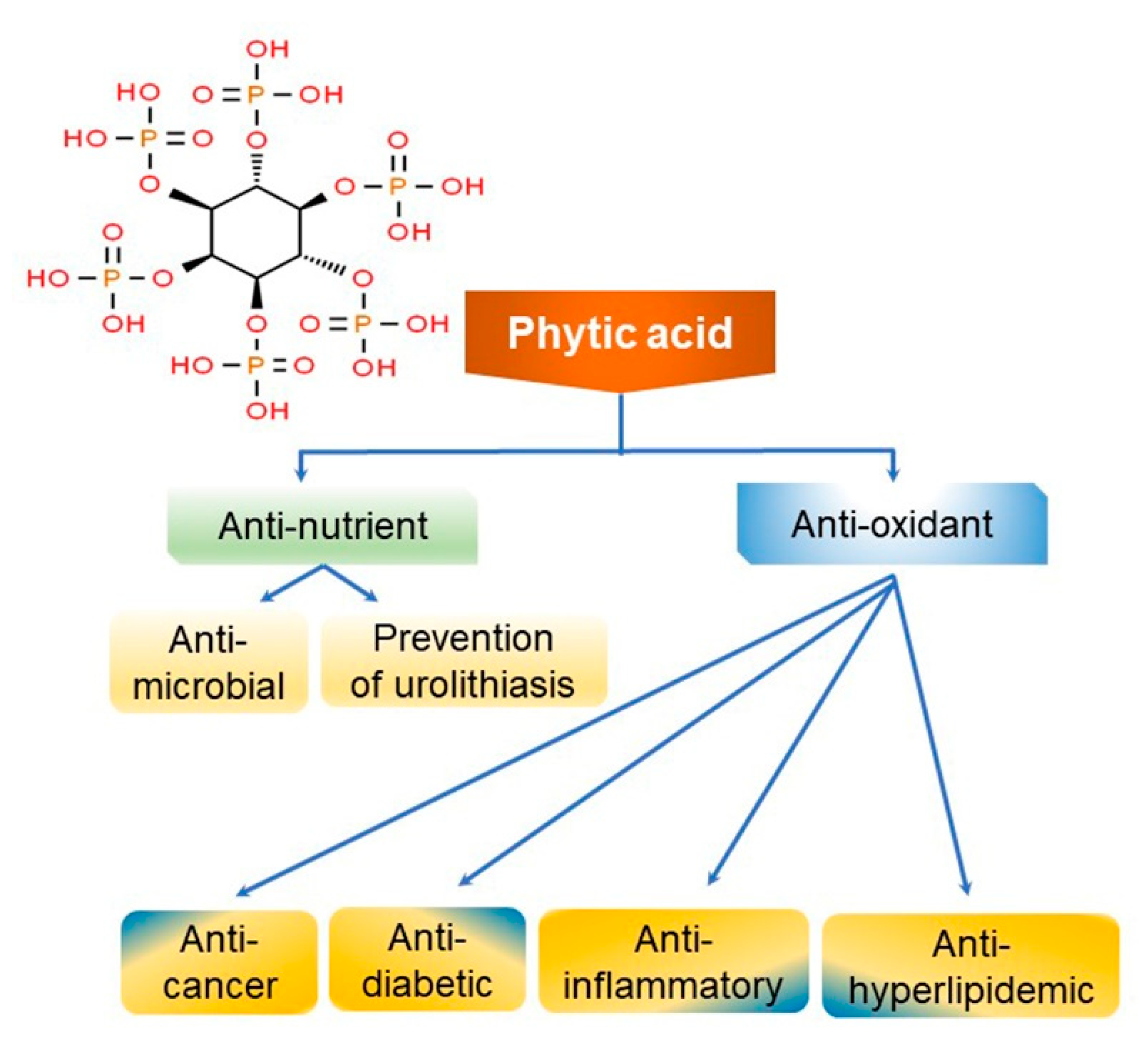
3. Phytic Acid
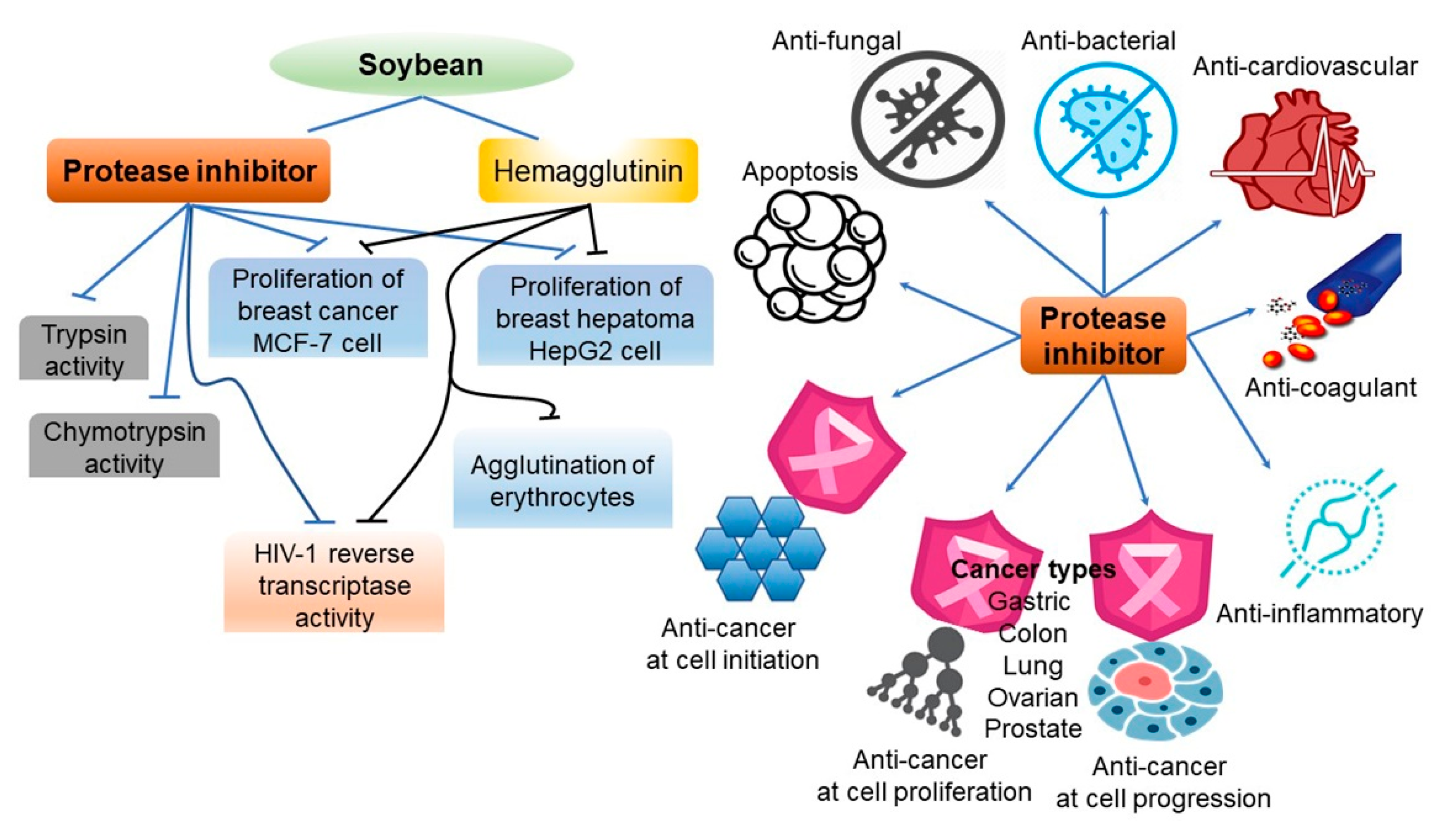
4. Protease Inhibitors
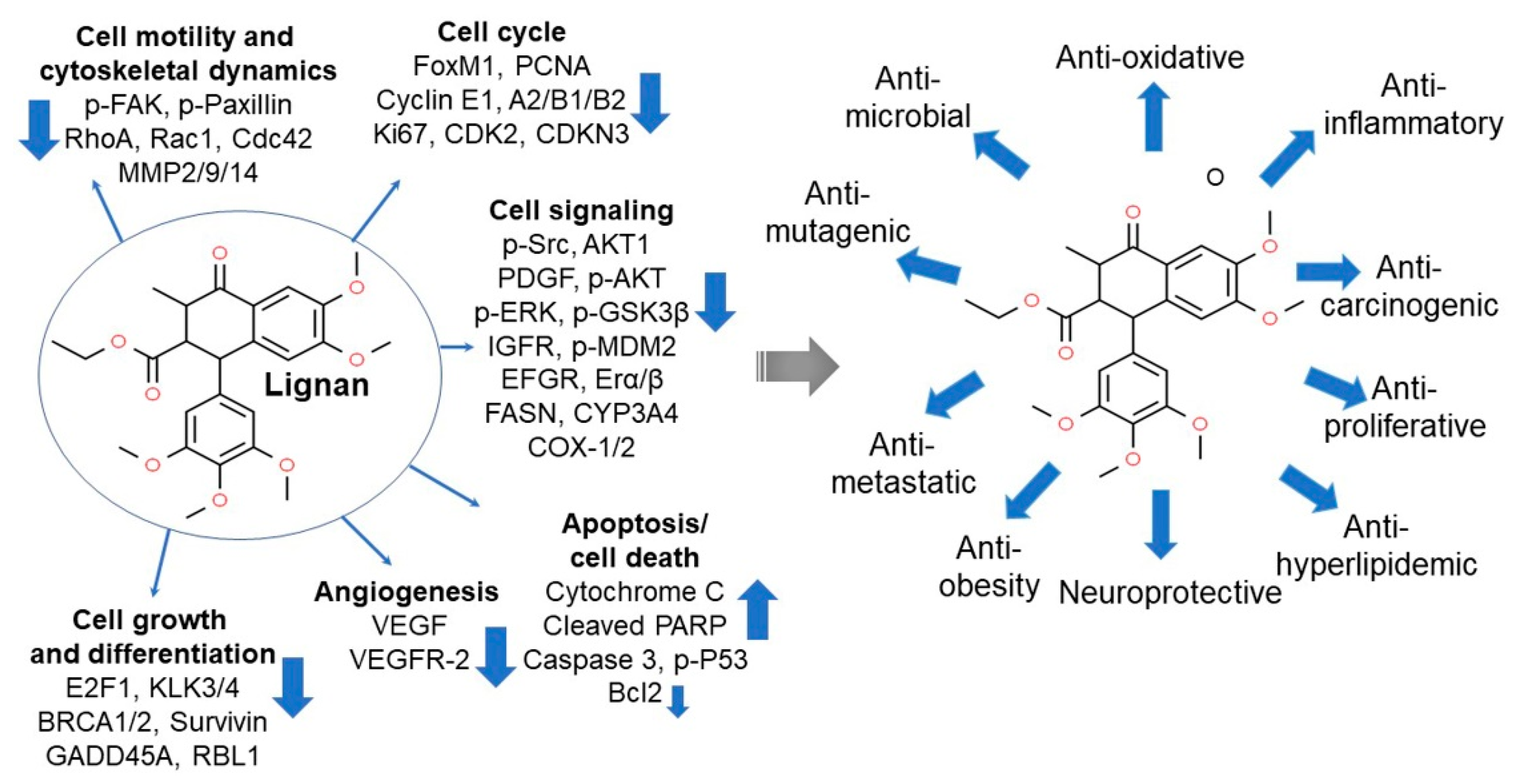
5. Lignans
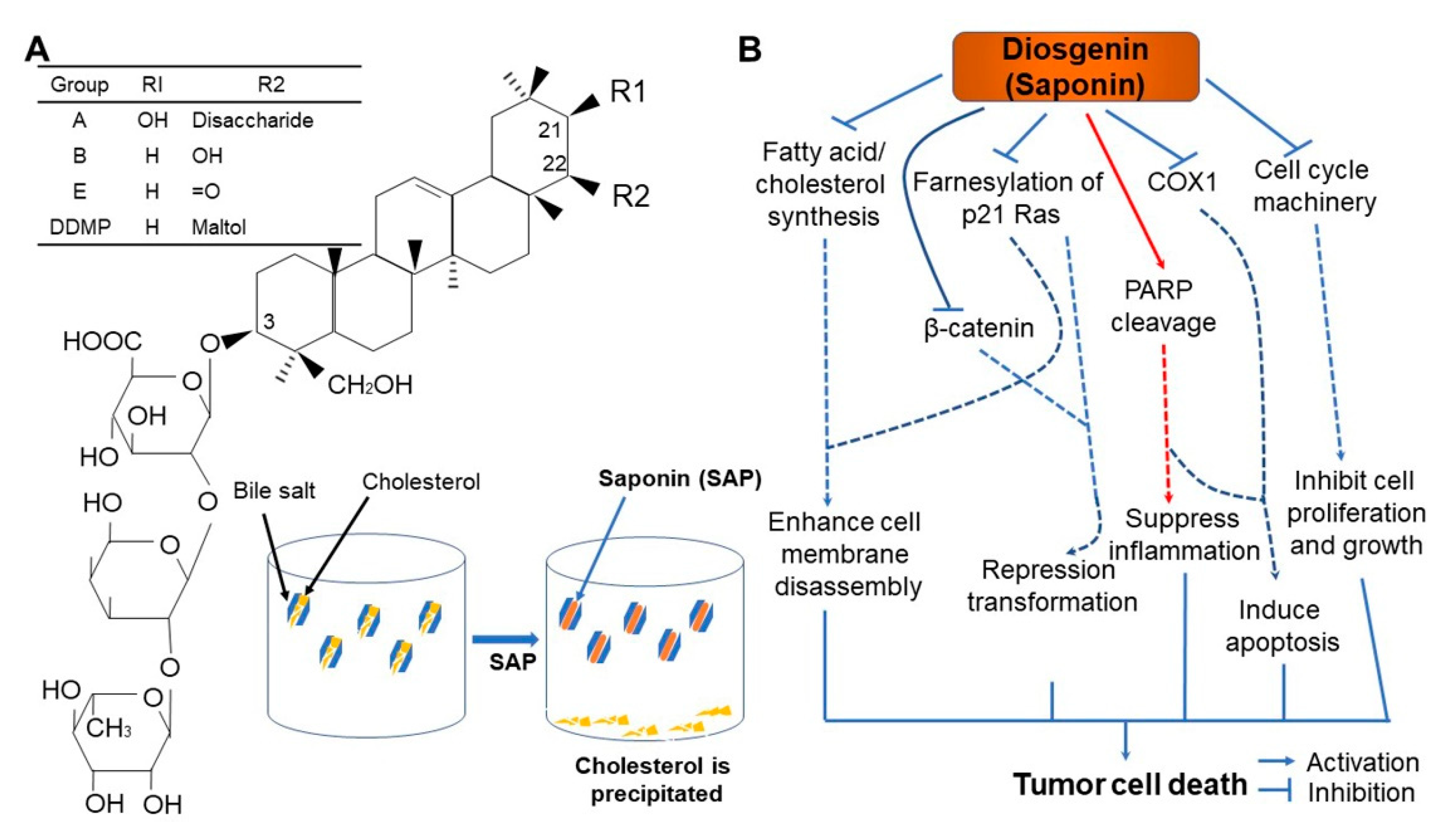
6. Saponins
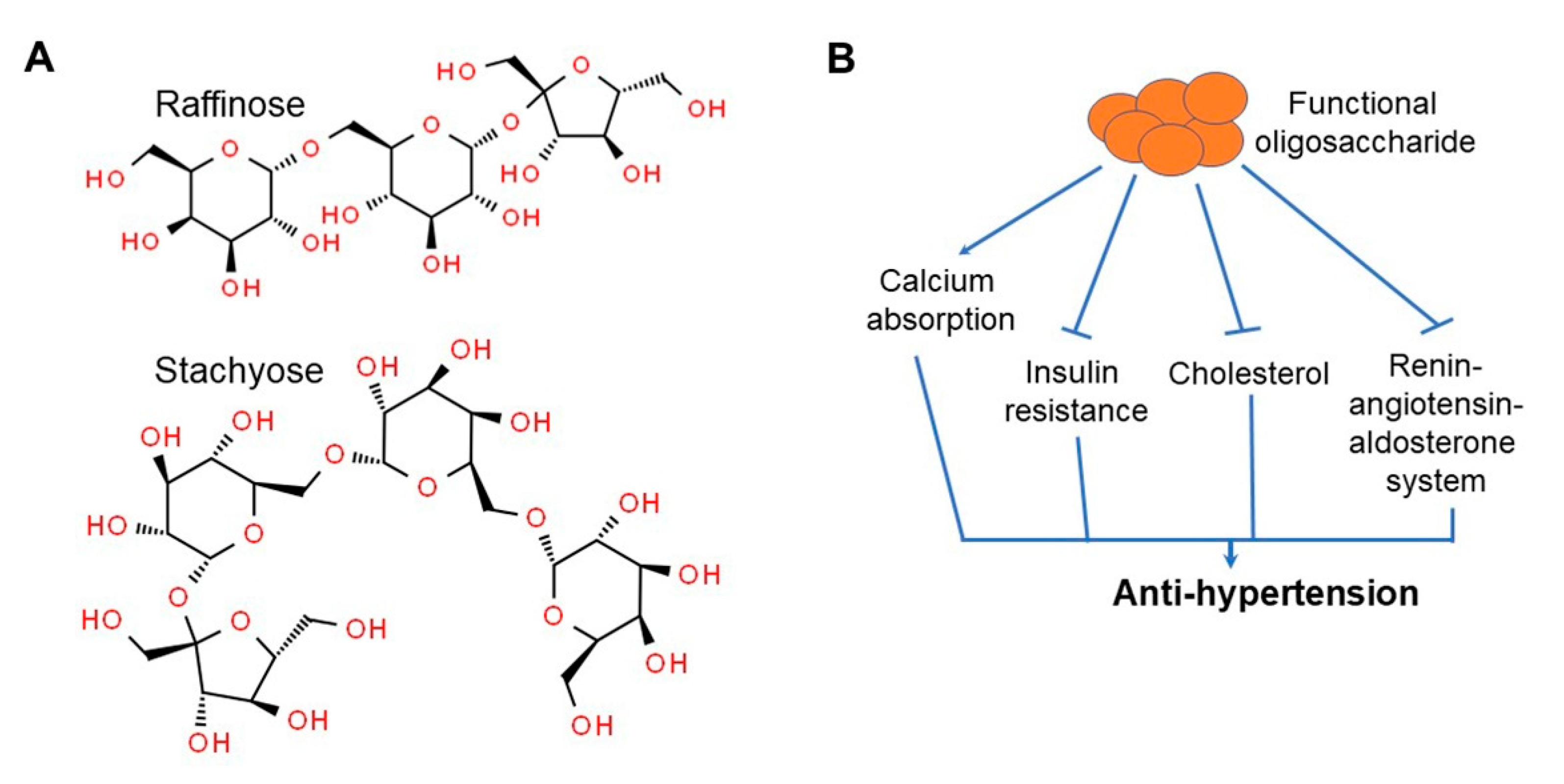
7. Dietary Fiber and Soy Oligosaccharides
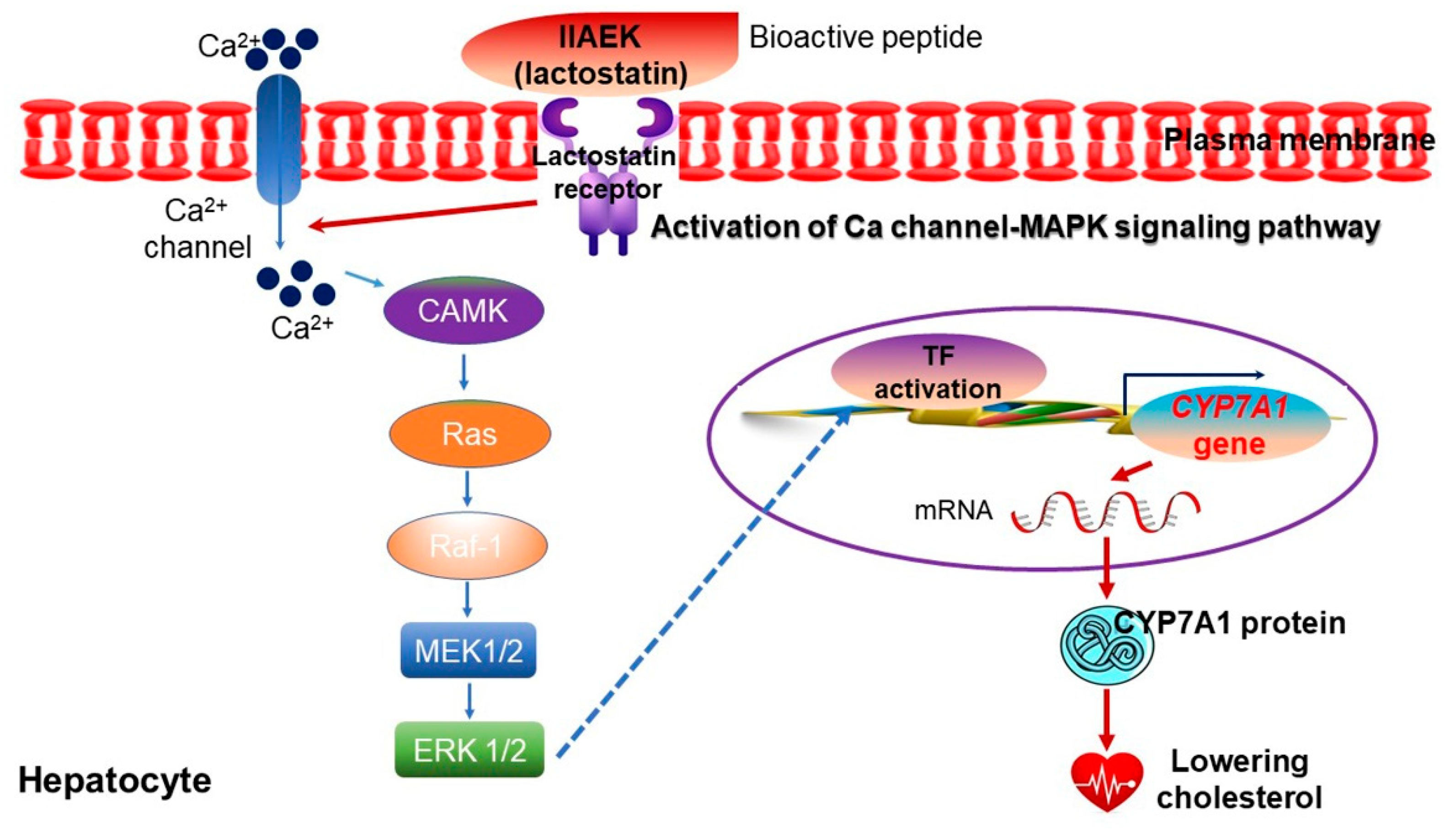
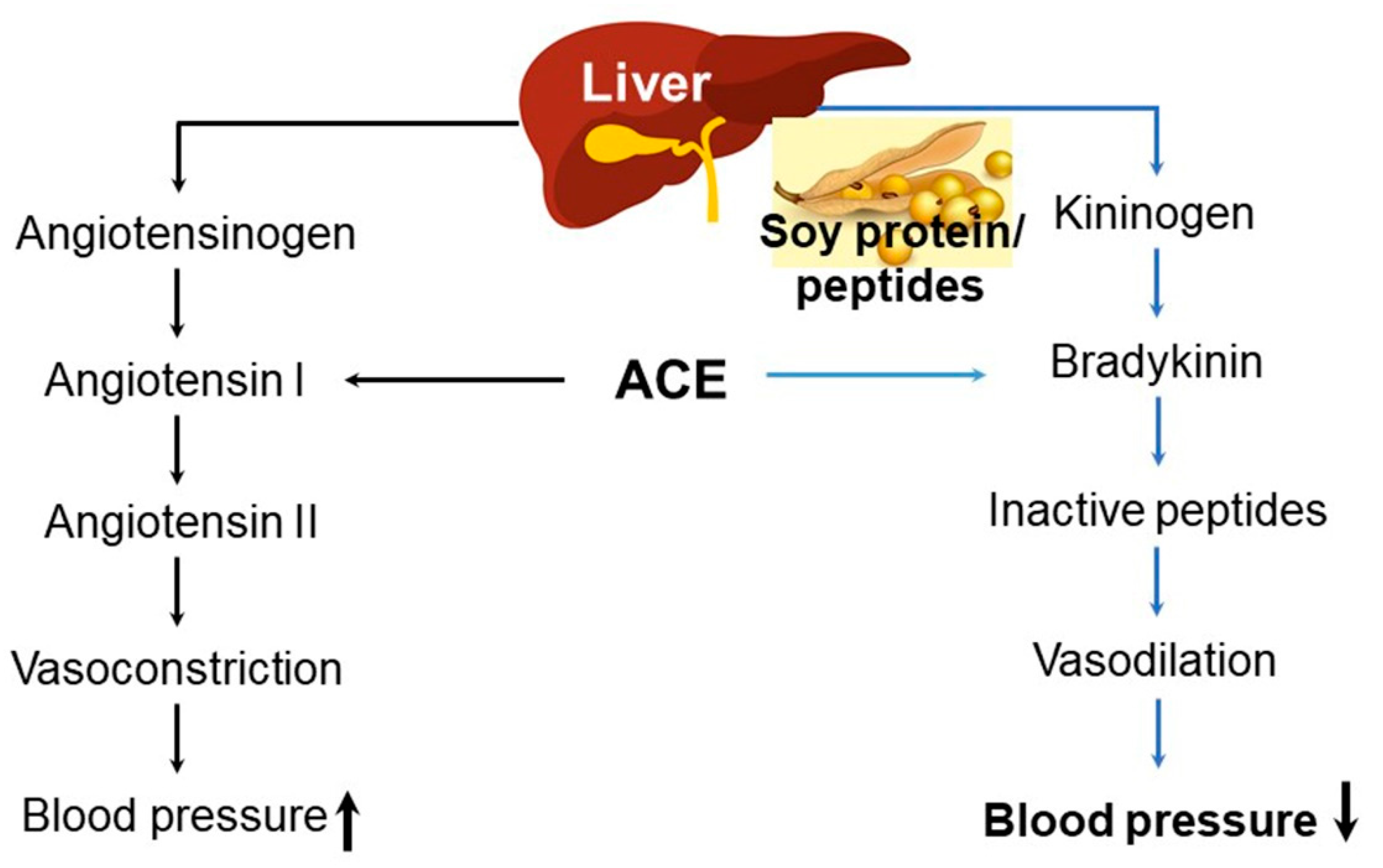
8. Soy Proteins and Peptides
9. Lecithin
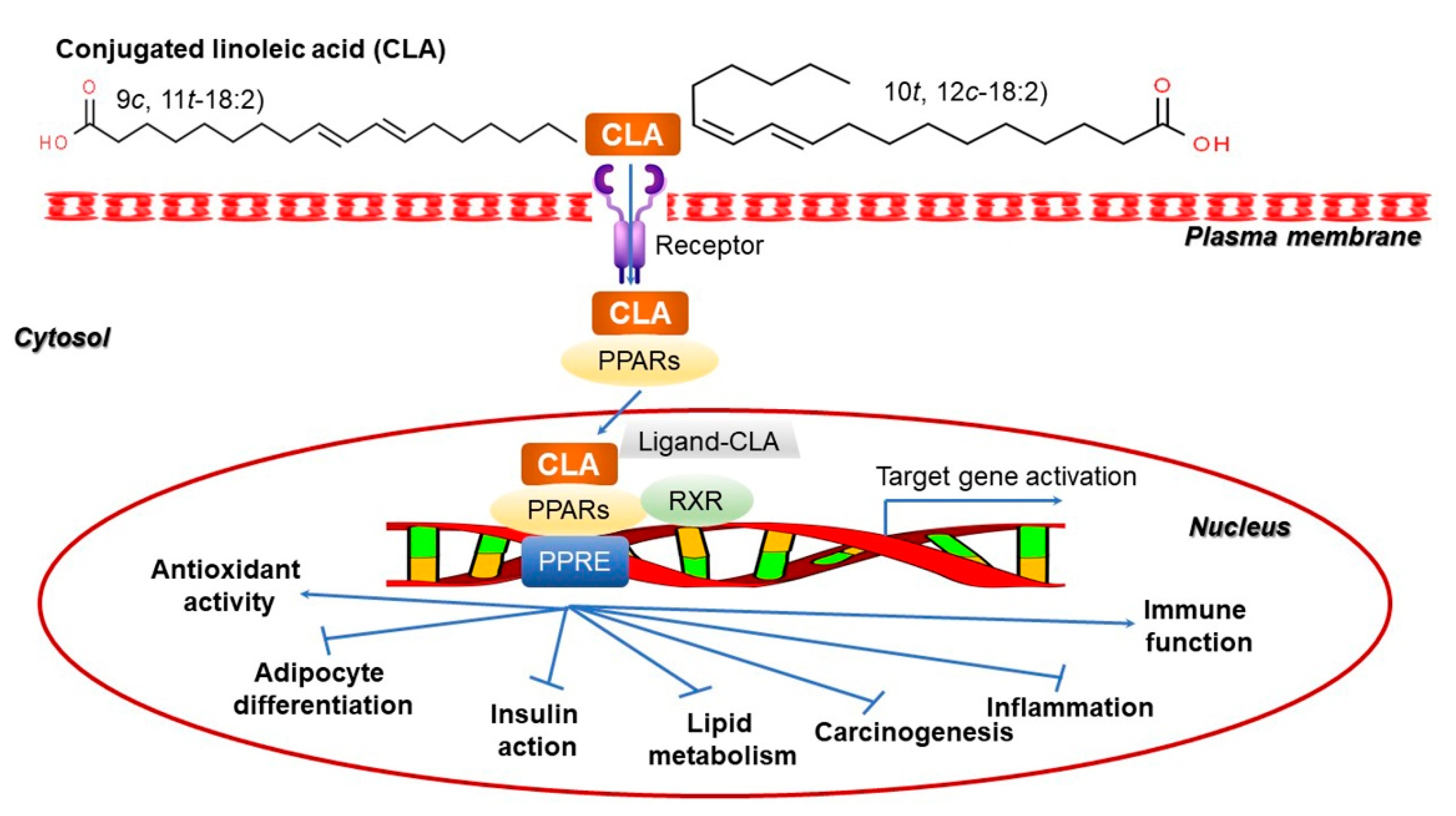
10. Conjugated Linoleic Acid
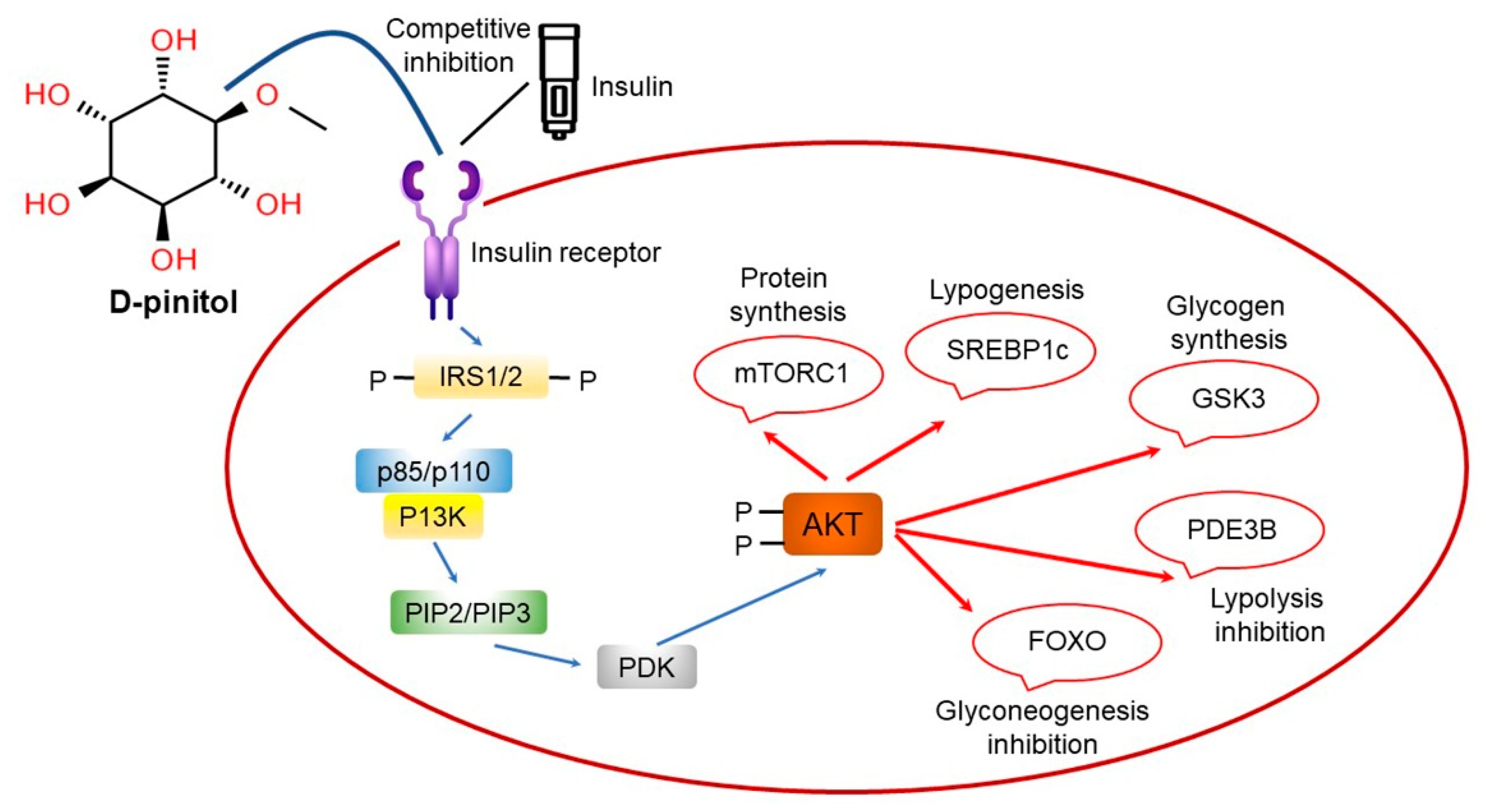
11. Pinitol
12. Conclusions
13. Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Chen, K.I.; Erh, M.H.; Su, N.W.; Liu, W.H.; Chou, C.C.; Cheng, K.C. Soyfoods and soybean products: From traditional use to modern applications. Appl. Microbiol. Biotechnol. 2012, 96, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.W.; Chung, Y.S.; Kwak, C.S.; Kwon, Y.H. Doenjang, a Korean traditional fermented soybean paste, ameliorates neuroinflammation and neurodegeneration in mice fed a high-fat diet. Nutrients 2019, 11, 1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Park, J.M.; Hwang, J.P. Analysis of iodine content in salts and Korean sauces for low-iodine diet education in Korean patients with thyroid cancer preparing for radioiodine therapy. Nucl. Med. Mol. Imaging 2018, 52, 229–233. [Google Scholar] [CrossRef]
- Rizzo, G.; Baroni, L. Soy, soy foods and their role in vegetarian diets. Nutrients 2018, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [Green Version]
- Messina, M. Soy and health update: Evaluation of the clinical and epidemiologic literature. Nutrients 2016, 8, 754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocrinol. 2010, 31, 400–419. [Google Scholar] [CrossRef] [Green Version]
- Akhlaghi, M.; Zare, M.; Nouripour, F. Effect of soy and soy isoflavones on obesity-related anthropometric measures: A systematic review and meta-analysis of randomized controlled clinical trials. Adv. Nutr. 2017, 8, 705–717. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Bhathena, S.J. Role of dietary soy protein in obesity. Int. J. Med. Sci. 2007, 4, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Thangavel, P.; Puga-Olguin, A.; Rodriguez-Landa, J.F.; Zepeda, R.C. Genistein as potential therapeutic candidate for menopausal symptoms and other related diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of fermentation in improving nutritional quality of soybean meal—A review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Wong, W.T.; Wu, R.; Lai, W.F. Biochemistry and use of soybean isoflavones in functional food development. Crit. Rev. Food Sci. Nutr. 2020, 60, 2098–2112. [Google Scholar] [CrossRef] [PubMed]
- Rollán, G.C.; Gerez, C.L.; LeBlanc, J.G. Lactic fermentation as a strategy to improve the nutritional and functional values of pseudocereals. Front. Nutr. 2019, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.Y. Bioactivity of soy-based fermented foods: A review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Roman, G.C.; Jackson, R.E.; Gadhia, R.; Roman, A.N.; Reis, J. Mediterranean diet: The role of long-chain omega-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. A critical review on polyphenols and health benefits of black soybeans. Nutrients 2017, 9, 455. [Google Scholar] [CrossRef] [Green Version]
- Sekikawa, A.; Ihara, M.; Lopez, O.; Kakuta, C.; Lopresti, B.; Higashiyama, A.; Aizenstein, H.; Chang, Y.F.; Mathis, C.; Miyamoto, Y.; et al. Effect of S-equol and soy isoflavones on heart and brain. Curr. Cardiol. Rev. 2019, 15, 114–135. [Google Scholar] [CrossRef]
- Zhou, T.; Meng, C.; He, P. Soy isoflavones and their effects on xenobiotic metabolism. Curr. Drug Metab. 2019, 20, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Udenigwe, C.C. Role of food-derived opioid peptides in the central nervous and gastrointestinal systems. J. Food Biochem. 2019, 43, e12629. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Lee, S.K.; Chun, O.K. Soy isoflavones and osteoporotic bone loss: A review with an emphasis on modulation of bone remodeling. J. Med. Food 2016, 19, 1–14. [Google Scholar] [CrossRef]
- Messina, M.; Messina, V. Soyfoods, soybean isoflavones, and bone health: A brief overview. J. Ren. Nutr. 2000, 10, 63–68. [Google Scholar] [CrossRef]
- Chang, Y.L.; Liu, T.; Tsai, M.L. Selective isolation of trypsin inhibitor and lectin from soybean whey by chitosan/tripolyphosphate/genipin co-crosslinked beads. Int. J. Mol. Sci. 2014, 15, 9979–9990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, U.K.; Lavignac, N.; Rahman, A.M.; Nielsen, B.V. Purification of lectin and Kunitz trypsin inhibitor from soya seeds. J. Chromatogr. Sci. 2018, 56, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Hellgren, L.I.; Olsen, E.; Skrede, A. Lipids rich in phosphatidylethanolamine from natural gas-utilizing bacteria reduce plasma cholesterol and classes of phospholipids: A comparison with soybean oil. Lipids 2004, 39, 833–841. [Google Scholar] [CrossRef]
- Jazi, V.; Mohebodini, H.; Ashayerizadeh, A.; Shabani, A.; Barekatain, R. Fermented soybean meal ameliorates Salmonella typhimurium infection in young broiler chickens. Poult. Sci. 2019, 98, 5648–5660. [Google Scholar] [CrossRef]
- Rajani, J.; Dastar, B.; Samadi, F.; Karimi Torshizi, M.A.; Abdulkhani, A.; Esfandyarpour, S. Effect of extracted galactoglucomannan oligosaccharides from pine wood (Pinus brutia) on Salmonella typhimurium colonisation, growth performance and intestinal morphology in broiler chicks. Br. Poult. Sci. 2016, 57, 682–692. [Google Scholar] [PubMed]
- Juritsch, A.F.; Moreau, R. Role of soybean-derived bioactive compounds in inflammatory bowel disease. Nutr. Rev. 2018, 76, 618–638. [Google Scholar] [CrossRef]
- Krol-Grzymala, A.; Amarowicz, R. Phenolic compounds of soybean seeds from two european countries and their antioxidant properties. Molecules 2020, 25, 2075. [Google Scholar] [CrossRef]
- Easwar Rao, D.; Viswanatha Chaitanya, K. Changes in the antioxidant intensities of seven different soybean (Glycine max (L.) Merr.) cultivars during drought. J. Food Biochem. 2020, 44, e13118. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Bao, B.; Ahmad, A.; Sarkar, F.H. Induction of cancer cell death by isoflavone: The role of multiple signaling pathways. Nutrients 2011, 3, 877–896. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.D.; Ziegler, V.; Schwanz Goebel, J.T.; Hoffmann, J.F.; Carvalho, I.R.; Chaves, F.C.; de Oliveira, M. Changes in phenolic acid and isoflavone contents during soybean drying and storage. J. Agric. Food Chem. 2019, 67, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Meenu, M.; Xu, B.; Yu, H. Impact of processing technologies on isoflavones, phenolic acids, and antioxidant capacities of soymilk prepared from 15 soybean varieties. Food Chem. 2021, 345, 128612. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Jin, H.; Yu, Y.; Sun, J.; Zheng, H.; Zhang, Y.; Xu, J.; Zhu, X. The improvement of nanoemulsion stability and antioxidation via protein-chlorogenic acid-dextran conjugates as emulsifiers. Nanomaterials 2020, 10, 1094. [Google Scholar] [CrossRef] [PubMed]
- How, J.S.L.; Morr, C.V. Removal of phenolic compounds from soy protein extracts using activated carbon. J. Food Sci. 1982, 47, 933–940. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, S.; Wang, M.; Feng, X.Y. Characterization of free, conjugated, and bound phenolic acids in seven commonly consumed vegetables. Molecules 2017, 22, 1878. [Google Scholar] [CrossRef] [Green Version]
- Benkerroum, N. Aflatoxins: Producing-molds, structure, health issues and incidence in Southeast Asian and Sub-Saharan African countries. Int. J. Environ. Res. Public Health 2020, 17, 1215. [Google Scholar] [CrossRef] [Green Version]
- Kiokias, S.; Proestos, C.; Oreopoulou, V. Phenolic acids of plant origin-A review on their antioxidant activity in vitro (O/W emulsion systems) along with their in vivo health biochemical properties. Foods 2020, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Liu, Y.B.; Peng, J.; Wang, J.H.; Liu, X. Changes of isoflavone profile in the hypocotyls and cotyledons of soybeans during dry heating and germination. J. Agric. Food Chem. 2009, 57, 9002–9010. [Google Scholar] [CrossRef]
- Swart, A.C.; Johannes, I.D.; Sathyapalan, T.; Atkin, S.L. The effect of soy isoflavones on steroid metabolism. Front. Endocrinol. 2019, 10, 229. [Google Scholar] [CrossRef]
- Nakai, S.; Fujita, M.; Kamei, Y. Health promotion effects of soy isoflavones. J. Nutr. Sci. Vitaminol. 2020, 66, 502–507. [Google Scholar] [CrossRef]
- Ajdzanovic, V.; Filipovic, B.; Miljic, D.; Mijatovic, S.; Maksimovic-Ivanic, D.; Miler, M.; Zivanovic, J.; Milosevic, V. Prostate cancer metastasis and soy isoflavones: A dogfight over a bone. EXCLI J. 2019, 18, 106–126. [Google Scholar] [PubMed]
- Sivonova, M.K.; Kaplan, P.; Tatarkova, Z.; Lichardusova, L.; Dusenka, R.; Jurecekova, J. Androgen receptor and soy isoflavones in prostate cancer. Mol. Clin. Oncol. 2019, 10, 191–204. [Google Scholar]
- Wang, P.P.; Qin, X.S.; Yang, Q.Y.; Luo, Z.G.; Xiao, Z.G.; Peng, X.C. Comparative structural characterization of spiral dextrin inclusion complexes with vitamin E or soy isoflavone. J. Agric. Food Chem. 2017, 65, 8744–8753. [Google Scholar] [CrossRef]
- De Jesus, L.C.L.; Soares, R.P.; Moreira, V.R.; Pontes, R.L.; Castelo-Branco, P.V.; Pereira, S.R.F. Genistein and ascorbic acid reduce oxidative stress-derived DNA damage induced by the antileishmanial meglumine antimoniate. Antimicrob. Agents Chemother. 2018, 62, e00456-18. [Google Scholar] [CrossRef] [Green Version]
- Aichinger, G.; Pahlke, G.; Nagel, L.J.; Berger, W.; Marko, D. Bilberry extract, its major polyphenolic compounds, and the soy isoflavone genistein antagonize the cytostatic drug erlotinib in human epithelial cells. Food Funct. 2016, 7, 3628–3636. [Google Scholar] [CrossRef] [PubMed]
- Moorehead, R.A. Rodent models assessing mammary tumor prevention by soy or soy isoflavones. Genes 2019, 10, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Poveda, B.; Torres-Vargas, J.A.; Ocana, M.D.C.; Garcia-Caballero, M.; Medina, M.A.; Quesada, A.R. The mediterranean diet, a rich source of angiopreventive compounds in cancer. Nutrients 2019, 11, 2036. [Google Scholar] [CrossRef] [Green Version]
- Ziaei, S.; Halaby, R. Dietary isoflavones and breast cancer risk. Medicines 2017, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, M.; Mallick, A.K. The effect of soy isoflavones on the menopause rating scale scoring in perimenopausal and postmenopausal women: A pilot study. J. Clin. Diagn. Res. 2017, 11, FC13–FC16. [Google Scholar] [CrossRef]
- Taku, K.; Umegaki, K.; Sato, Y.; Taki, Y.; Endoh, K.; Watanabe, S. Soy isoflavones lower serum total and LDL cholesterol in humans: A meta-analysis of 11 randomized controlled trials. Am. J. Clin. Nutr. 2007, 85, 1148–1156. [Google Scholar] [CrossRef]
- Koury, O.H.; Scheede-Bergdahl, C.; Bergdahl, A. The role of casein in the development of hypercholesterolemia. J. Physiol. Biochem. 2014, 70, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Ramdath, D.D.; Padhi, E.M.; Sarfaraz, S.; Renwick, S.; Duncan, A.M. Beyond the cholesterol-lowering effect of soy protein: A review of the effects of dietary soy and its constituents on risk factors for cardiovascular disease. Nutrients 2017, 9, 324. [Google Scholar] [CrossRef] [Green Version]
- Palacios, S.; Stevenson, J.C.; Schaudig, K.; Lukasiewicz, M.; Graziottin, A. Hormone therapy for first-line management of menopausal symptoms: Practical recommendations. Womens Health 2019, 15, 1745506519864009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall, G.V.; Britt, K.L. Estrogen effects on the mammary gland in early and late life and breast cancer risk. Front. Oncol. 2017, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Sunyecz, J.A. The use of calcium and vitamin D in the management of osteoporosis. Ther. Clin. Risk Manag. 2008, 4, 827–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Bi, X.; Yu, B.; Chen, D. Isoflavones: Anti-inflammatory benefit and possible caveats. Nutrients 2016, 8, 361. [Google Scholar] [CrossRef] [Green Version]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular mechanisms of action of genistein in cancer: Recent advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Kligman, L.; Younus, J. Management of hot flashes in women with breast cancer. Curr. Oncol. 2010, 17, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Akhlaghi, M.; Ghasemi Nasab, M.; Riasatian, M.; Sadeghi, F. Soy isoflavones prevent bone resorption and loss, a systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 2327–2341. [Google Scholar] [CrossRef]
- Silva, E.O.; Bracarense, A.P. Phytic acid: From antinutritional to multiple protection factor of organic systems. J. Food Sci. 2016, 81, R1357–R1362. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, M.; Talsma, E.F.; Taleon, V.; Londono, L.; Brychkova, G.; Gallego, S.; Raatz, B.; Spillane, C. Iron, zinc and phytic acid retention of biofortified, low phytic acid, and conventional bean varieties when preparing common household recipes. Nutrients 2020, 12, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marolt, G.; Kolar, M. Analytical methods for determination of phytic acid and other inositol phosphates: A review. Molecules 2020, 26, 174. [Google Scholar] [CrossRef] [PubMed]
- Masum Akond, A.; Crawford, H.; Berthold, J.; Talukder, Z.I.; Hossain, K. Minerals (Zn, Fe, Ca and Mg) and antinutrient (phytic Acid) constituents in common bean. Am. J. Food Technol. 2011, 6, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Popova, A.; Mihaylova, D. Antinutrients in plant-based foods: A review. Open Biotechnol. J. 2019, 13, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [Green Version]
- Abdulwaliyu, I.; Arekemase, S.O.; Adudu, J.A.; Batari, M.L.; Egbule, M.N.; Okoduwa, S.I.R. Investigation of the medicinal significance of phytic acid as an indispensable anti-nutrient in diseases. Clin. Nutr. Exp. 2019, 28, 42–61. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, B.; Raigond, P.; Sahu, C.; Mishra, U.N.; Sharma, S.; Lal, M.K. Phytic acid: Blessing in disguise, a prime compound required for both plant and human nutrition. Food Res. Int. 2021, 142, 110193. [Google Scholar] [CrossRef]
- Zhao, Z. Iron and oxidizing species in oxidative stress and Alzheimer’s disease. Aging Med. 2019, 2, 82–87. [Google Scholar] [CrossRef]
- Vucenik, I.; Druzijanic, A.; Druzijanic, N. Inositol hexaphosphate (IP6) and colon cancer: From concepts and first experiments to clinical application. Molecules 2020, 25, 5931. [Google Scholar] [CrossRef] [PubMed]
- Khatiwada, J.; Verghese, M.; Davis, S.; Williams, L.L. Green tea, phytic acid, and inositol in combination reduced the incidence of azoxymethane-induced colon tumors in Fisher 344 male rats. J. Med. Food 2011, 14, 1313–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A. The inositol pyrophosphate pathway in health and diseases. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1203–1227. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Chen, Z. Plant protease inhibitors in therapeutics-focus on cancer therapy. Front. Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellinger, R.; Gruber, C.W. Peptide-based protease inhibitors from plants. Drug Discov. Today 2019, 24, 1877–1889. [Google Scholar] [CrossRef] [PubMed]
- Gitlin-Domagalska, A.; Maciejewska, A.; Debowski, D. Bowman-Birk inhibitors: Insights into family of multifunctional proteins and peptides with potential therapeutical applications. Pharmaceuticals 2020, 13, 421. [Google Scholar] [CrossRef]
- Clemente, A.; Arques Mdel, C. Bowman-Birk inhibitors from legumes as colorectal chemopreventive agents. World J. Gastroenterol. 2014, 20, 10305–10315. [Google Scholar] [CrossRef]
- Kakizoe, T. Chemoprevention of cancer—Focusing on clinical trials. Jpn. J. Clin. Oncol. 2003, 33, 421–442. [Google Scholar] [CrossRef] [Green Version]
- Cristina Oliveira de Lima, V.; Piuvezam, G.; Leal Lima Maciel, B.; Heloneida de Araujo Morais, A. Trypsin inhibitors: Promising candidate satietogenic proteins as complementary treatment for obesity and metabolic disorders? J. Enzyme Inhib. Med. Chem. 2019, 34, 405–419. [Google Scholar] [CrossRef]
- Behr, M.; Sergeant, K.; Leclercq, C.C.; Planchon, S.; Guignard, C.; Lenouvel, A.; Renaut, J.; Hausman, J.F.; Lutts, S.; Guerriero, G. Insights into the molecular regulation of monolignol-derived product biosynthesis in the growing hemp hypocotyl. BMC Plant. Biol. 2018, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Frezza, C.; Venditti, A.; Toniolo, C.; De Vita, D.; Franceschin, M.; Ventrone, A.; Tomassini, L.; Foddai, S.; Guiso, M.; Nicoletti, M.; et al. Nor-lignans: Occurrence in plants and biological activities—A review. Molecules 2020, 25, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, K.D.; Brown, N.M.; Zimmer-Nechemias, L.; Wolfe, B.; Jha, P.; Heubi, J.E. Metabolism of secoisolariciresinol-diglycoside the dietary precursor to the intestinally derived lignan enterolactone in humans. Food Funct. 2014, 5, 491–501. [Google Scholar] [CrossRef]
- Meagher, L.P.; Beecher, G.R. Assessment of data on the lignan content of foods. J. Food Compos. Anal. 2000, 13, 935–947. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Toledo, E.; Delgado-Rodriguez, M.; Gaforio, J.J. Naturally lignan-rich foods: A dietary tool for health promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.; Dwyer, J.; Adlercreutz, H.; Scalbert, A.; Jacques, P.; McCullough, M.L. Dietary lignans: Physiology and potential for cardiovascular disease risk reduction. Nutr. Rev. 2010, 68, 571–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinardell, M.P.; Mitjans, M. Lignins and their derivatives with beneficial effects on human health. Int. J. Mol. Sci. 2017, 18, 1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.A.; Griffiths, K.; Morton, M.S. Inhibition of 5 alpha-reductase in genital skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids. J. Endocrinol. 1995, 147, 295–302. [Google Scholar] [CrossRef]
- Brooks, J.D.; Thompson, L.U. Mammalian lignans and genistein decrease the activities of aromatase and 17beta-hydroxysteroid dehydrogenase in MCF-7 cells. J. Steroid Biochem. Mol. Biol. 2005, 94, 461–467. [Google Scholar] [CrossRef]
- Cotterchio, M.; Boucher, B.A.; Manno, M.; Gallinger, S.; Okey, A.; Harper, P. Dietary phytoestrogen intake is associated with reduced colorectal cancer risk. J. Nutr. 2006, 136, 3046–3053. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Yao, S.; Tritchler, D.; Hullar, M.A.; Lampe, J.W.; Thompson, L.U.; McCann, S.E. Genetic variation in steroid and xenobiotic metabolizing pathways and enterolactone excretion before and after flaxseed intervention in African American and European American women. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 265–274. [Google Scholar] [CrossRef] [Green Version]
- De Silva, S.F.; Alcorn, J. Flaxseed lignans as important dietary polyphenols for cancer prevention and treatment: Chemistry, pharmacokinetics, and molecular targets. Pharmaceuticals 2019, 12, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezzat, S.M.; Shouman, S.A.; Elkhoely, A.; Attia, Y.M.; Elsesy, M.S.; El Senousy, A.S.; Choucry, M.A.; El Gayed, S.H.; El Sayed, A.A.; Sattar, E.A.; et al. Anticancer potentiality of lignan rich fraction of six flaxseed cultivars. Sci. Rep. 2018, 8, 544. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Arunasalam, K.; Yeung, D.; Kakuda, Y.; Mittal, G.; Jiang, Y. Saponins from edible legumes: Chemistry, processing, and health benefits. J. Med. Food 2004, 7, 67–78. [Google Scholar] [CrossRef]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [Green Version]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef]
- Kamo, S.; Suzuki, S.; Sato, T. The content of soyasaponin and soyasapogenol in soy foods and their estimated intake in the Japanese. Food Sci. Nutr. 2014, 2, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Neacsu, M.; Raikos, V.; Benavides-Paz, Y.; Duncan, S.H.; Duncan, G.J.; Christie, J.S.; Johnstone, A.M.; Russell, W.R. Sapogenol is a major microbial metabolite in human plasma associated with high protein soy-based diets: The relevance for functional food formulations. Foods 2020, 9, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, T.K. Glycyrrhiza glabra. Edible Medicinal and Non-Medicinal Plants; Lim, T.K., Ed.; Springer: New York, NY, USA, 2016; Volume 10, pp. 354–457. [Google Scholar] [CrossRef]
- Berhow, M.A.; Kong, S.B.; Vermillion, K.E.; Duval, S.M. Complete quantification of group A and group B soyasaponins in soybeans. J. Agric. Food Chem. 2006, 54, 2035–2044. [Google Scholar] [CrossRef]
- Guajardo-Flores, D.; Garcia-Patino, M.; Serna-Guerrero, D.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Characterization and quantification of saponins and flavonoids in sprouts, seed coats and cotyledons of germinated black beans. Food Chem. 2012, 134, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [Green Version]
- Dawid, C.; Hofmann, T. Quantitation and bitter taste contribution of saponins in fresh and cooked white asparagus (Asparagus officinalis L.). Food Chem. 2014, 145, 427–436. [Google Scholar] [CrossRef]
- Marrelli, M.; Conforti, F.; Araniti, F.; Statti, G.A. Effects of saponins on lipid metabolism: A review of potential health benefits in the treatment of obesity. Molecules 2016, 21, 1404. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.N.; Dilger, R.N. Immunomodulatory potential of dietary soybean-derived isoflavones and saponins in pigs. J. Anim. Sci. 2018, 96, 1288–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, Y.S.; Contador, C.A.; Ng, M.S.; Yu, J.; Chung, G.; Lam, H.M. The effects of domestication on secondary metabolite composition in legumes. Front. Genet. 2020, 11, 581357. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Sharif, Y.; Abbas, S.; Afzal, M.Z.; Qasim, M.; Khalofah, A.; Ansari, M.J.; Khan, K.A.; Tao, L.; Li, S. Saponin toxicity as key player in plant defense against pathogens. Toxicon 2021, 193, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wang, K.; Wang, X.; Wen, H.; Zhou, H.; Liu, C.; Mai, K.; He, G. Soybean saponin modulates nutrient sensing pathways and metabolism in zebrafish. Gen. Comp. Endocrinol. 2018, 257, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, Q.N.; Kim, M.J.; Kim, T.G.; Pan, J.H.; Kim, J.H.; Park, S.J.; Lee, J.H.; Kim, Y.J. Saponin-based nanoemulsification improves the antioxidant properties of vitamin A and E in AML-12 cells. Int. J. Mol. Sci. 2016, 17, 1406. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. A century of cholesterol and coronaries: From plaques to genes to statins. Cell 2015, 161, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Yamanashi, Y.; Takada, T.; Kurauchi, R.; Tanaka, Y.; Komine, T.; Suzuki, H. Transporters for the intestinal absorption of cholesterol, vitamin E, and vitamin K. J. Atheroscler. Thromb. 2017, 24, 347–359. [Google Scholar] [CrossRef] [Green Version]
- Galmes, S.; Serra, F.; Palou, A. Vitamin E metabolic effects and genetic variants: A challenge for precision nutrition in obesity and associated disturbances. Nutrients 2018, 10, 1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tin, M.M.; Cho, C.H.; Chan, K.; James, A.E.; Ko, J.K. Astragalus saponins induce growth inhibition and apoptosis in human colon cancer cells and tumor xenograft. Carcinogenesis 2007, 28, 1347–1355. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Zhao, H.; Jiang, G.; Feng, X.; Sui, W.; Liu, H. Soyasapogenol B exhibits anti-growth and anti-metastatic activities in clear cell renal cell carcinoma. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, H.; Niesler, N.; Trautner, A.; Sama, S.; Jerz, G.; Panjideh, H.; Weng, A. Glycosylated triterpenoids as endosomal escape enhancers in targeted tumor therapies. Biomedicines 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.H.; Li, T.; Fong, C.M.; Chen, X.; Chen, X.J.; Wang, Y.T.; Huang, M.Q.; Lu, J.J. Saponins from Chinese medicines as anticancer agents. Molecules 2016, 21, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, X.; Huang, N.; Liu, R.; Sun, R. A comprehensive review and perspectives on pharmacology and toxicology of saikosaponins. Phytomedicine 2018, 50, 73–87. [Google Scholar] [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Markowiak-Kopec, P.; Slizewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Bae, S.H. Diets for constipation. Pediatr. Gastroenterol. Hepatol. Nutr. 2014, 17, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Jõudu, I.; Bhat, R. Dietary fiber from underutilized plant resources—A positive approach for valorization of fruit and vegetable wastes. Sustainability 2020, 12, 5401. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, C.H.; Saps, M. The role of fiber in the treatment of functional gastrointestinal disorders in children. Nutrients 2018, 10, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wang, Y.; Li, W.; Qin, F.; Chen, J. Effects of oligosaccharides and soy soluble polysaccharide on the rheological and textural properties of calcium sulfate-induced soy protein gels. Food Bioproc. Technol. 2016, 10, 556–567. [Google Scholar] [CrossRef]
- Salarbashi, D.; Bazeli, J.; Tafaghodi, M. Environment-friendly green composites based on soluble soybean polysaccharide: A review. Int. J. Biol. Macromol. 2019, 122, 216–223. [Google Scholar] [CrossRef]
- Pan, L.; Farouk, M.H.; Qin, G.; Zhao, Y.; Bao, N. The influences of soybean agglutinin and functional oligosaccharides on the intestinal tract of monogastric animals. Int. J. Mol. Sci. 2018, 19, 554. [Google Scholar] [CrossRef] [Green Version]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, T.F.; Casarotti, S.N.; de Oliveira, G.L.V.; Penna, A.L.B. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit. Rev. Food Sci. Nutr. 2021, 61, 337–355. [Google Scholar] [CrossRef]
- Saez-Lara, M.J.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: A systematic review of randomized human clinical trials. Biomed. Res. Int. 2015, 2015, 505878. [Google Scholar] [CrossRef]
- Pique, N.; Berlanga, M.; Minana-Galbis, D. Health benefits of heat-killed (tyndallized) probiotics: An overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Yan, Q.; Liu, J.; Wu, X.; Jiang, Z. Can functional oligosaccharides reduce the risk of diabetes mellitus? FASEB J. 2019, 33, 11655–11667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, G.; Mian, R.; Vuong, T.; Pantalone, V.; Song, Q.; Chen, P.; Shannon, G.J.; Carter, T.C.; Nguyen, H.T. Molecular mapping and genomics of soybean seed protein: A review and perspective for the future. Theor. Appl. Genet. 2017, 130, 1975–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Kumar, R.; Sabapathy, S.N.; Bawa, A.S. Functional and edible uses of soy protein products. Compr. Rev. Food Sci. Food Saf. 2008, 7, 14–28. [Google Scholar] [CrossRef]
- Nachvak, S.M.; Moradi, S.; Anjom-Shoae, J.; Rahmani, J.; Nasiri, M.; Maleki, V.; Sadeghi, O. Soy, soy isoflavones, and protein intake in relation to mortality from all causes, cancers, and cardiovascular diseases: A systematic review and dose-response meta-analysis of prospective cohort studies. J. Acad. Nutr. Diet. 2019, 119, 1483–1500. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Daneshzad, E.; Azadbakht, L. The effects of isolated soy protein, isolated soy isoflavones and soy protein containing isoflavones on serum lipids in postmenopausal women: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2020, 60, 3414–3428. [Google Scholar] [CrossRef] [PubMed]
- George, K.S.; Munoz, J.; Akhavan, N.S.; Foley, E.M.; Siebert, S.C.; Tenenbaum, G.; Khalil, D.A.; Chai, S.C.; Arjmandi, B.H. Is soy protein effective in reducing cholesterol and improving bone health? Food Funct. 2020, 11, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Monte Singer, W.; Zhang, B.; Rouf Mian, M.A.; Huang, H. Soybean amino acids in health, genetics, and evaluation. Soybean for Human Consumption and Animal Feed; Sudaric, A., Ed.; IntechOpen: London, UK, 2020. [Google Scholar]
- Kearney, J. Food consumption trends and drivers. Philos. Trans. R Soc. B Biol. Sci. 2010, 365, 2793–2807. [Google Scholar] [CrossRef]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial role of phytochemicals on oxidative stress and age-related diseases. Biomed. Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [Green Version]
- Imai, S. Soybean and processed soy foods ingredients, and their role in cardiometabolic risk prevention. Recent Pat. Food Nutr. Agric. 2015, 7, 75–82. [Google Scholar] [CrossRef]
- Montgomery, K.S. Soy protein. J. Perinat. Educ. 2003, 12, 42–45. [Google Scholar] [CrossRef]
- Richter, C.K.; Skulas-Ray, A.C.; Fleming, J.A.; Link, C.J.; Mukherjea, R.; Krul, E.S.; Kris-Etherton, P.M. Effects of isoflavone-containing soya protein on ex vivo cholesterol efflux, vascular function and blood markers of CVD risk in adults with moderately elevated blood pressure: A dose-response randomised controlled trial. Br. J. Nutr. 2017, 117, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yang, J.; Lei, L.; Wang, L.; Wang, X.; Ma, K.Y.; Yang, X.; Chen, Z.Y. Isoflavones enhance the plasma cholesterol-lowering activity of 7S protein in hypercholesterolemic hamsters. Food Funct. 2019, 10, 7378–7386. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.D. Enzymatic hydrolysis of soy proteins and the hydrolysates utilisation. Int. J. Food Sci. Technol. 2011, 46, 2447–2459. [Google Scholar] [CrossRef]
- McCarthy, A.; O’Callaghan, Y.; O’Brien, N. Protein hydrolysates from agricultural crops—bioactivity and potential for functional food development. Agriculture 2013, 3, 112–130. [Google Scholar] [CrossRef] [Green Version]
- Hanafi, M.A.; Hashim, S.N.; Chay, S.Y.; Ebrahimpour, A.; Zarei, M.; Muhammad, K.; Abdul-Hamid, A.; Saari, N. High angiotensin-I converting enzyme (ACE) inhibitory activity of Alcalase-digested green soybean (Glycine max) hydrolysates. Food Res. Int. 2018, 106, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S. Structure-function properties of hypolipidemic peptides. J. Food Chem. 2019, 43, e12539. [Google Scholar] [CrossRef] [Green Version]
- Robert, C.; Couedelo, L.; Vaysse, C.; Michalski, M.C. Vegetable lecithins: A review of their compositional diversity, impact on lipid metabolism and potential in cardiometabolic disease prevention. Biochimie 2020, 169, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Le, N.T.T.; Cao, V.D.; Nguyen, T.N.Q.; Le, T.T.H.; Tran, T.T.; Hoang Thi, T.T. Soy lecithin-derived liposomal delivery systems: Surface modification and current applications. Int. J. Mol. Sci. 2019, 20, 4706. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, B.; Ao, H.; Fu, J.; Wang, Y.; Feng, Y.; Guo, Y.; Wang, X. Soybean lecithin stabilizes disulfiram nanosuspensions with a high drug-loading content: Remarkably improved antitumor efficacy. J. Nanobiotechnol. 2020, 18, 4. [Google Scholar] [CrossRef]
- Van Hoogevest, P. Review—An update on the use of oral phospholipid excipients. Eur. J. Pharm. Sci. 2017, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kateh Shamshiri, M.; Momtazi-Borojeni, A.A.; Khodabandeh Shahraky, M.; Rahimi, F. Lecithin soybean phospholipid nano-transfersomes as potential carriers for transdermal delivery of the human growth hormone. J. Cell. Biochem. 2019, 120, 9023–9033. [Google Scholar] [CrossRef] [PubMed]
- Tai, K.; Liu, F.; He, X.; Ma, P.; Mao, L.; Gao, Y.; Yuan, F. The effect of sterol derivatives on properties of soybean and egg yolk lecithin liposomes: Stability, structure and membrane characteristics. Food Res. Int. 2018, 109, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Van Hoogevest, P.; Wendel, A. The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur. J. Lipid Sci. Technol. 2014, 116, 1088–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blesso, C.N. Egg phospholipids and cardiovascular health. Nutrients 2015, 7, 2731–2747. [Google Scholar] [CrossRef] [Green Version]
- Alhajj, M.J.; Montero, N.; Yarce, C.J.; Salamanca, C.H. Lecithins from vegetable, land, and marine animal sources and their potential applications for cosmetic, food, and pharmaceutical sectors. Cosmetics 2020, 7, 87. [Google Scholar] [CrossRef]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane lipid composition: Effect on membrane and organelle structure, function and compartmentalization and therapeutic avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovic, M.; Ben-Shabat, S.; Aponick, A.; Zimmermann, E.M.; Dahan, A. Lipids and lipid-processing pathways in drug delivery and therapeutics. Int. J. Mol. Sci. 2020, 21, 3248. [Google Scholar] [CrossRef]
- Hails, G.; Cerminati, S.; Paoletti, L.; Cabrera, R.; Peiru, S.; Aguirre, A.; Castelli, M.E.; Menzella, H.G. A novel lecithin:cholesterol acyltransferase for soybean oil refining provides higher yields and extra nutritional value with a cleaner process. Appl. Microbiol. Biotechnol. 2020, 104, 7521–7532. [Google Scholar] [CrossRef]
- Li, X.Z.; Park, B.K.; Hong, B.C.; Ahn, J.S.; Shin, J.S. Effect of soy lecithin on total cholesterol content, fatty acid composition and carcass characteristics in the Longissimus dorsi of Hanwoo steers (Korean native cattle). Anim. Sci. J. 2017, 88, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, M.; Couedelo, L.; Meugnier, E.; Loizon, E.; Plaisancie, P.; Durand, A.; Geloen, A.; Joffre, F.; Vaysse, C.; Michalski, M.C.; et al. Soybean polar lipids differently impact adipose tissue inflammation and the endotoxin transporters LBP and sCD14 in flaxseed vs. palm oil-rich diets. J. Nutr. Biochem. 2017, 43, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.A.; Meservey, C.M.; Nicolosi, R.J. Soy lecithin reduces plasma lipoprotein cholesterol and early atherogenesis in hypercholesterolemic monkeys and hamsters: Beyond linoleate. Atherosclerosis 1998, 140, 147–153. [Google Scholar] [CrossRef]
- Félix, R.; Valentão, P.; Andrade, P.B.; Félix, C.; Novais, S.C.; Lemos, M.F.L. Evaluating the in vitro potential of natural extracts to protect lipids from oxidative damage. Antioxidants 2020, 9, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- More, M.I.; Freitas, U.; Rutenberg, D. Positive effects of soy lecithin-derived phosphatidylserine plus phosphatidic acid on memory, cognition, daily functioning, and mood in elderly patients with Alzheimer’s disease and dementia. Adv. Ther. 2014, 31, 1247–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jager, R.; Purpura, M.; Kingsley, M. Phospholipids and sports performance. J. Int. Soc. Sports Nutr. 2007, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Yamatoya, H.; Sakai, M.; Kataoka, A.; Furushiro, M.; Kudo, S. Oral administration of soybean lecithin transphosphatidylated phosphatidylserine improves memory impairment in aged rats. J. Nutr. 2001, 131, 2951–2956. [Google Scholar] [CrossRef] [Green Version]
- Morganstern, I.; Ye, Z.; Liang, S.; Fagan, S.; Leibowitz, S.F. Involvement of cholinergic mechanisms in the behavioral effects of dietary fat consumption. Brain Res. 2012, 1470, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, M.S.; Oliveros, L.B.; Gomez, N.N. Nutritional deficiencies and phospholipid metabolism. Int. J. Mol. Sci. 2011, 12, 2408–2433. [Google Scholar] [CrossRef] [Green Version]
- Ridgway, N.D. The role of phosphatidylcholine and choline metabolites to cell proliferation and survival. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 20–38. [Google Scholar] [CrossRef]
- Poly, C.; Massaro, J.M.; Seshadri, S.; Wolf, P.A.; Cho, E.; Krall, E.; Jacques, P.F.; Au, R. The relation of dietary choline to cognitive performance and white-matter hyperintensity in the framingham offspring cohort. Am. J. Clin. Nutr. 2011, 94, 1584–1591. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, D.; Park, Y. Conjugated linoleic acid (CLA) promotes endurance capacity via peroxisome proliferator-activated receptor delta-mediated mechanism in mice. J. Nutr. Biochem. 2016, 38, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, C.E.; Kim, S.C.; Kim, D.H.; Lee, H.Y.; Suh, H.K.; Cho, K.M.; Lee, J.H. Enhancement of isoflavone aglycone, amino acid, and CLA contents in fermented soybean yogurts using different strains: Screening of antioxidant and digestive enzyme inhibition properties. Food Chem. 2021, 340, 128199. [Google Scholar] [CrossRef]
- Den Hartigh, L.J. Conjugated linoleic acid effects on cancer, obesity, and atherosclerosis: A review of pre-clinical and human trials with current perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, R.; van der Wielen, R.P.; Brink, E.J.; Hendriks, H.F.; Dorovska-Taran, V.N.; Mohede, I.C. Effects of cis-9, trans-11 and trans-10, cis-12 conjugated linoleic acid (CLA) isomers on immune function in healthy men. Eur. J. Clin. Nutr. 2003, 57, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Y.M.; Kadir, A.A.; Ahmad, Z.; Yaakub, H.; Zakaria, Z.A.; Abdullah, M.N. Free radical scavenging activity of conjugated linoleic acid as single or mixed isomers. Pharm. Biol. 2012, 50, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrugala, D.; Leatherwood, J.L.; Morris, E.F.; Dickson, E.C.; Latham, C.M.; Owen, R.N.; Beverly, M.M.; Kelley, S.F.; White-Springer, S.H. Dietary conjugated linoleic acid supplementation alters skeletal muscle mitochondria and antioxidant status in young horses. J. Anim. Sci. 2021, 99, skab037. [Google Scholar] [CrossRef]
- Bialek, A.; Czerwonka, M.; Bialek, M.; Lepionka, T.; Kaszperuk, K.; Banaszkiewicz, T.; Tokarz, A. Influence of pomegranate seed oil and grape seed oil on cholesterol content and fatty acids profile in livers of chickens. Acta Pol. Pharm. 2017, 74, 624–632. [Google Scholar]
- Gilbert, W.; Gadang, V.; Proctor, A.; Jain, V.; Devareddy, L. Trans-trans conjugated linoleic acid enriched soybean oil reduces fatty liver and lowers serum cholesterol in obese zucker rats. Lipids 2011, 46, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Bezan, P.N.; Holland, H.; de Castro, G.S.; Cardoso, J.F.R.; Ovidio, P.P.; Calder, P.C.; Jordao, A.A. High dose of a conjugated linoleic acid mixture increases insulin resistance in rats fed either a low fat or a high fat diet. Exp. Clin. Endocrinol. Diabetes 2018, 126, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Bauer, P.V.; Duca, F.A.; Waise, T.M.Z.; Dranse, H.J.; Rasmussen, B.A.; Puri, A.; Rasti, M.; O’Brien, C.A.; Lam, T.K.T. Lactobacillus gasseri in the upper small intestine impacts an ACSL3-dependent fatty acid-sensing pathway regulating whole-body glucose homeostasis. Cell Metab. 2018, 27, 572–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Rienzi, S.C.; Jacobson, J.; Kennedy, E.A.; Bell, M.E.; Shi, Q.; Waters, J.L.; Lawrence, P.; Brenna, J.T.; Britton, R.A.; Walter, J.; et al. Resilience of small intestinal beneficial bacteria to the toxicity of soybean oil fatty acids. Elife 2018, 7, e32581. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Xu, M.; Yu, F.; Fu, C.L.; Yu, X.; Cheng, M.; Gao, H.Q. Effects of D-pinitol on myocardial apoptosis and fibrosis in streptozocin-induced aging-accelerated mice. J. Food Biochem. 2021, e13669. [Google Scholar]
- Lin, Y.; Wu, Y.; Su, J.; Wang, M.; Wu, X.; Su, Z.; Yi, X.; Wei, L.; Cai, J.; Sun, Z. Therapeutic role of D-pinitol on experimental colitis via activating Nrf2/ARE and PPAR-gamma/NF-kappaB signaling pathways. Food Funct. 2021. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.S.; Kim, S.; Kim, M.; Hong, Y.A.; Shin, S.J.; Park, C.W.; Chang, Y.S.; Chung, S.; Kim, H.S. D-Pinitol alleviates cyclosporine-A induced renal tubulointerstitial fibrosis via activating Sirt1 and Nrf2 antioxidant pathways. Int. J. Mol. Med. 2018, 41, 1826–1834. [Google Scholar] [CrossRef]
- Dumschott, K.; Dechorgnat, J.; Merchant, A. Water deficit elicits a transcriptional response of genes governing d-pinitol biosynthesis in soybean (Glycine max). Int. J. Mol. Sci. 2019, 20, 2411. [Google Scholar] [CrossRef] [Green Version]
- Ignacio López-Sánchez, J.; Moreno, D.A.; García-Viguer, C. D-pinitol, a highly valuable product from carob pods: Health-promoting effects and metabolic pathways of this natural super-food ingredient and its derivatives. AIMS Agric. Food 2018, 3, 41–63. [Google Scholar] [CrossRef]
- Larner, J.; Brautigan, D.L.; Thorner, M.O. D-chiro-inositol glycans in insulin signaling and insulin resistance. Mol. Med. 2010, 16, 543–552. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Bizzarri, M. Inositols in insulin signaling and glucose metabolism. Int. J. Endocrinol. 2018, 2018, 1968450. [Google Scholar] [CrossRef] [Green Version]
- Mancini, M.; Andreassi, A.; Salvioni, M.; Pelliccione, F.; Mantellassi, G.; Banderali, G. Myoinositol and D-chiro inositol in improving insulin resistance in obese male children: Preliminary data. Int. J. Endocrinol. 2016, 2016, 8720342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, L.C.; Tang, T.; Yasmin, E.; Norman, R.J.; Balen, A.H. Insulin-sensitising drugs (metformin, rosiglitazone, pioglitazone, D-chiro-inositol) for women with polycystic ovary syndrome, oligo amenorrhoea and subfertility. Cochrane Database Syst. Rev. 2017, 11, CD003053. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hidalgo, M.; León-González, A.J.; Gálvez-Peralta, M.; González-Mauraza, N.H.; Martin-Cordero, C. D-Pinitol: A cyclitol with versatile biological and pharmacological activities. Phytochem. Rev. 2020. [Google Scholar] [CrossRef]
- Kang, M.J.; Kim, J.I.; Yoon, S.Y.; Kim, J.C.; Cha, I.J. Pinitol from soybeans reduces postprandial blood glucose in patients with type 2 diabetes mellitus. J. Med. Food 2006, 9, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Antonowski, T.; Osowski, A.; Lahuta, L.; Górecki, R.; Rynkiewicz, A.; Wojtkiewicz, J. Health-promoting properties of selected cyclitols for metabolic syndrome and diabetes. Nutrients 2019, 11, 2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichim, M.C. The Romanian experience and perspective on the commercial cultivation of genetically modified crops in Europe. Transg. Res. 2019, 28, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bøhn, T.; Millstone, E. The introduction of thousands of tonnes of glyphosate in the food chain-An evaluation of glyphosate tolerant soybean. Foods 2019, 8, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricroch, A.E.; Hénard-Damave, M.C. Next biotech plants: New traits, crops, developers and technologies for addressing global challenges. Crit. Rev. Biotechnol. 2016, 36, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Tyczewska, A.; Woźniak, E.; Gracz, J.; Kuczyński, J.; Twardowski, T. Towards food security: Current state and future prospects of agrobiotechnology. Trends Biotechnol. 2018, 36, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Tsatsakis, A.M.; Nawaz, M.A.; Tutelyan, V.A.; Golokhvast, K.S.; Kalantzi, O.I.; Chung, D.H.; Kang, S.J.; Coleman, M.D.; Tyshko, N.; Yang, S.H.; et al. Impact on environment, ecosystem, diversity and health from culturing and using GMOs as feed and food. Food Chem. Toxicol. 2017, 107, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Selb, R.; Wal, J.M.; Moreno, F.J.; Lovik, M.; Mills, C.; Hoffmann-Sommergruber, K.; Fernandez, A. Assessment of endogenous allergenicity of genetically modified plants exemplified by soybean—Where do we stand? Food Chem. Toxicol. 2017, 101, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Panchin, A.Y.; Tuzhikov, A.I. Published GMO studies find no evidence of harm when corrected for multiple comparisons. Crit. Rev. Biotechnol. 2017, 37, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crops Food 2017, 8, 13–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, W.A.; Chandanshive, S.U.; Bhatt, V.; Nadaf, A.B.; Vats, S.; Katara, J.L.; Sonah, H.; Deshmukh, R. Genome editing in cereals: Approaches, applications and challenges. Int. J. Mol. Sci. 2020, 21, 4040. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Nutritional Value (Per 100 g) | Component | Nutritional Value (Per 100 g) |
|---|---|---|---|
| Carbohydrates | 30.20 g | Glycine | 1.88 g |
| Sugars | 7.30 g | Proline | 2.38 g |
| Protein | 36.49 g | Serine | 2.36 g |
| Tryptophan | 0.59 g | Fat | 19.94 g |
| Threonine | 1.77 g | Saturated FA | 2.89 g |
| Isoleucine | 1.97 g | Monounsaturated FA | 4.40 g |
| Leucine | 3.31 g | Polyunsaturated FA | 11.26 g |
| Lysine | 2.71 g | Water | 8.54 g |
| Methionine | 0.55 g | Vitamin A | 0.001 g |
| Phenylalanine | 2.12 g | Vitamin B6 | 0.006 g |
| Tyrosine | 1.54 g | Vitamin C | 0.047 g |
| Valine | 2.03 g | Vitamin K | 0.277 g |
| Arginine | 3.15 g | Calcium | 1.57 g |
| Histidine | 1.10 g | Magnesium | 0.28 g |
| Alanine | 1.92 g | Phosphorous | 0.704 g |
| Aspartate | 5.12 g | Sodium | 1.797 g |
| Glutamate | 7.87 g | Zinc | 0.002 g |
| Total calories | 466 kcal | ||
| Peptide Sequences | Biological Effects |
|---|---|
| APP; IPP; AFH; PPYY; PPYY; YVVPK; IPPGVPYWT; LAIPVNKP; LPHF; VLIVP; SPYP; WL; NWGPLV; IVF; LLF; LNF; LSW; IAV; LEF; LEPP; FFYY; FVP; LHPDAQR; VNP; WNPR; WHP; VAHINVGK; YVWK; SY; GY | ACE inhibitor |
| ADPVLDNEGNPLENGGTYYI | ACE inhibitor and antioxidant |
| KNPQLR; EITPEKNPQLR; RKQEEDEEQQRE | Fatty acid synthase inhibitor |
| VRIRLLQRFNKRS | Appetite suppressant |
| HCQRPR; QRPR | Phagocytosis-stimulating peptide |
| VK; KA; SY | Lower triglyceride |
| ILL; LLL; VHVV | Lipolysis |
| HHL; PGTAVPK; YVVFK; IPPCVPYWT; PNNKPFQ; NWGPLV; TRRVF | Antihypertensive |
| PGTAVPK; HTSKALDMLKRLGK | Antimicrobial |
| RQRK; VIK | Anti-inflammatory |
| IQN | Adipogenesis inhibition |
| QRPR; HCQRPR | Immunomodulator |
| LPYP; LPYPR; WGAPSL; VAWWMY; FVVNATSN; IIAEK | Hypocholesterolemic |
| Vglycin | Antidiabetic |
| IAVPGEVA; IAVPTGVA; | Hypocholesterolemic and antidiabetic |
| LLPHH; RPLKPW | Antioxidative and antihypertensive |
| (X)MLPSWSPW; SLWQHQQDSCRLQLQGVNLFPCEL HIMELIQGRGDDDDDDD | Anticancer |
| Bowman-Birk inhibitor | Anticancer, protease inhibition, and chemoprevention |
| SKWQHQQDSCRKQKQGVNLTPCEKHIHEKIQGRGD DDDDDDDD | Antioxidative, anti-inflammatory, anticancer, and hypocholesterolemic |
| LPYPR; PGP | Antiobesity |
| YPFVV; YPFVVN; YPFVVNA | Antidiabetic, immunomodulator, lower triglyceride, and suppress feed and intestinal transit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.-S.; Kim, C.-H.; Yang, W.-S. Physiologically Active Molecules and Functional Properties of Soybeans in Human Health—A Current Perspective. Int. J. Mol. Sci. 2021, 22, 4054. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084054
Kim I-S, Kim C-H, Yang W-S. Physiologically Active Molecules and Functional Properties of Soybeans in Human Health—A Current Perspective. International Journal of Molecular Sciences. 2021; 22(8):4054. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084054
Chicago/Turabian StyleKim, Il-Sup, Cheorl-Ho Kim, and Woong-Suk Yang. 2021. "Physiologically Active Molecules and Functional Properties of Soybeans in Human Health—A Current Perspective" International Journal of Molecular Sciences 22, no. 8: 4054. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084054