
In Vitro Characterization of Neutralizing Hen Antibodies to Coxsackievirus A16

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
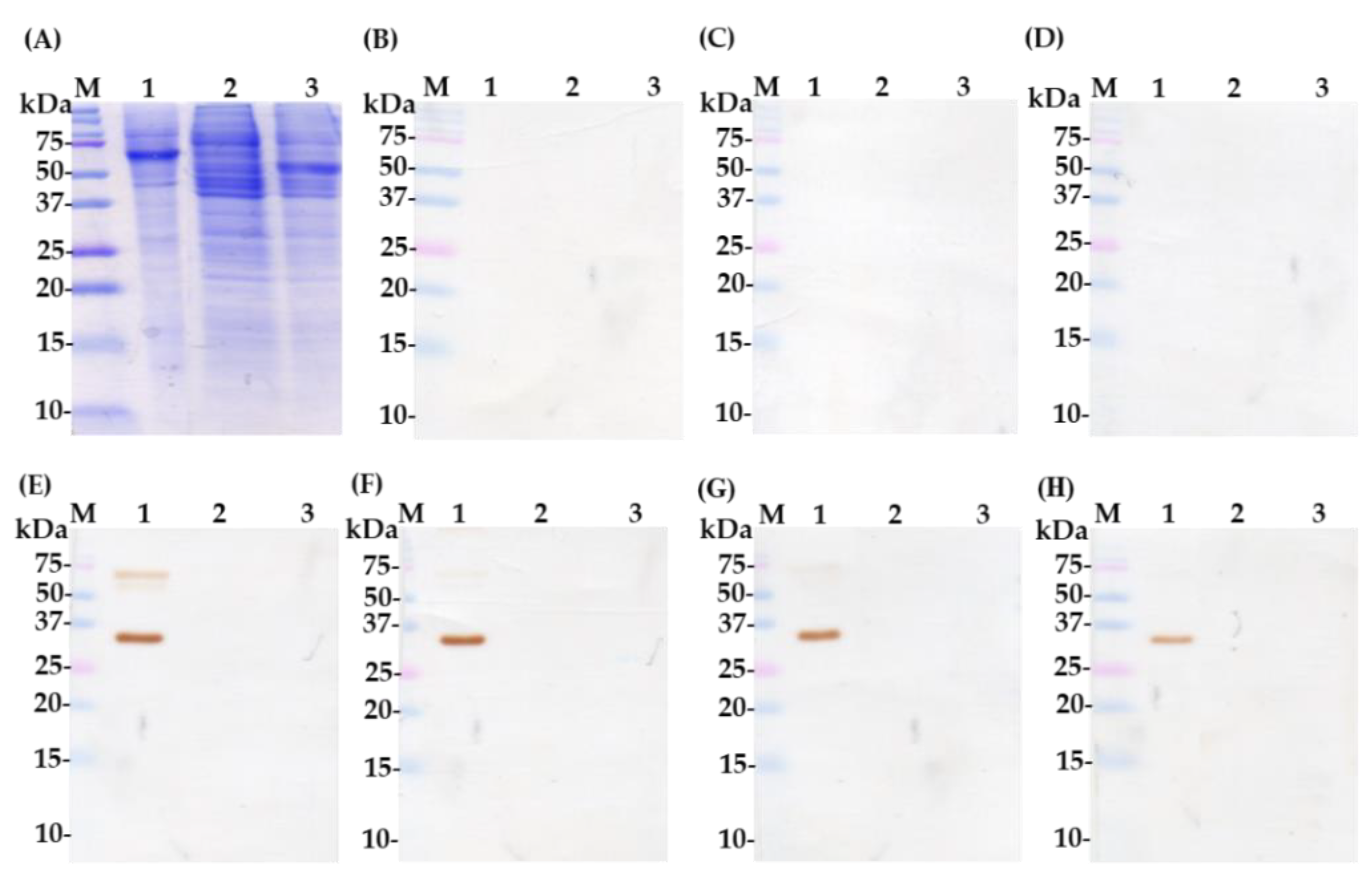
2.1. Characterization of Polyclonal Immunoglobulin Yolk (IgY)-Binding Protein Antibodies
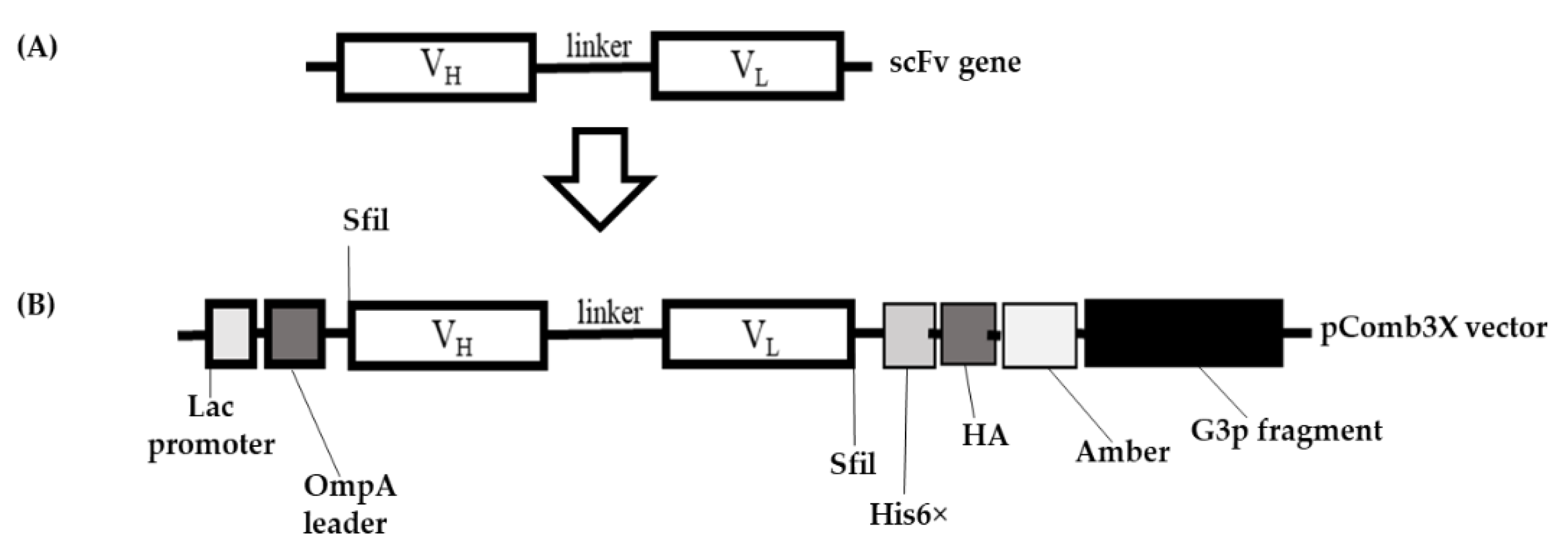
2.2. Cloning of Single-Chain Variable Fragment (scFv) Gene
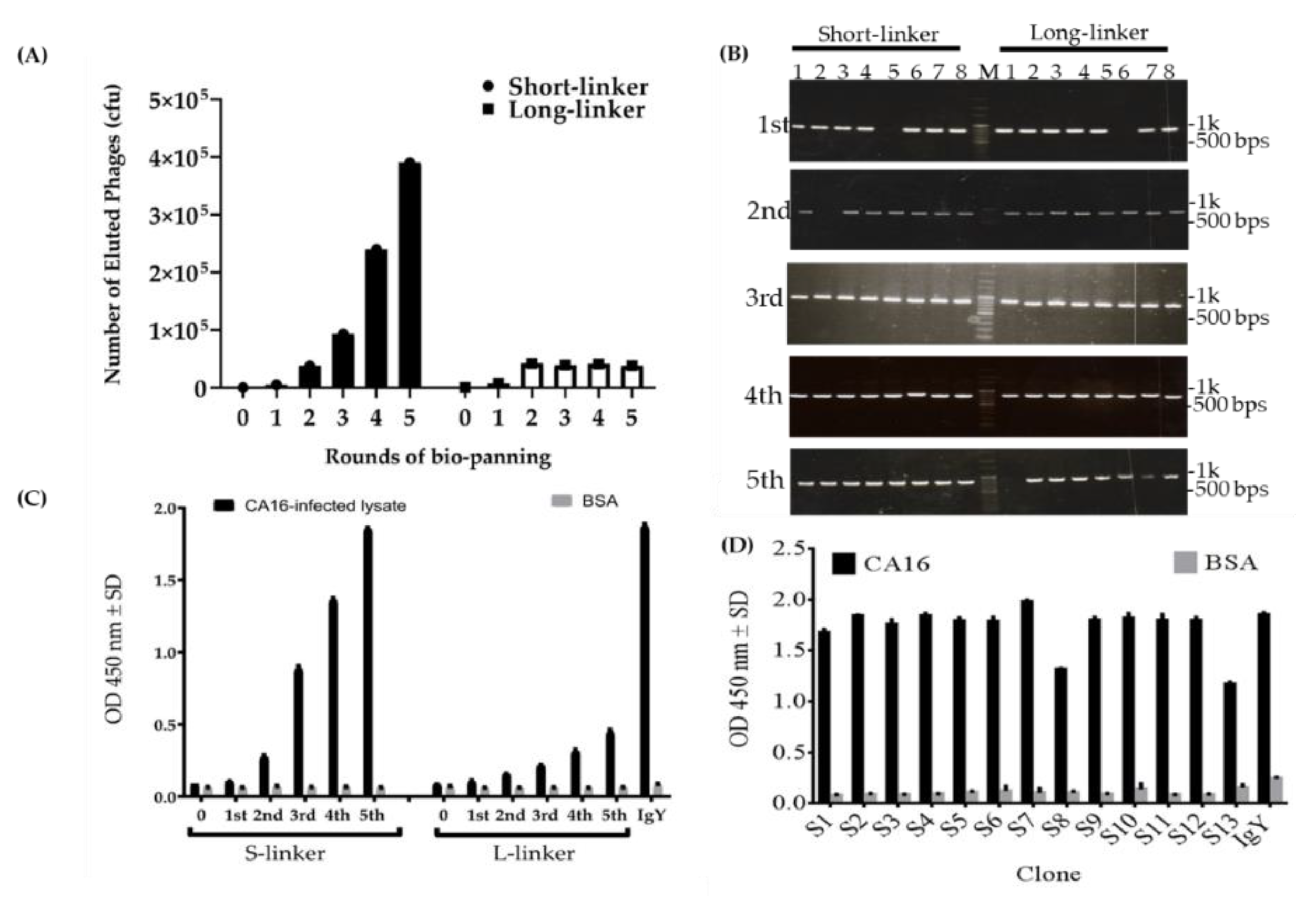
2.3. Characterization of Phage-Displaying scFv Libraries and Selection of scFv-Binding Protein Clones
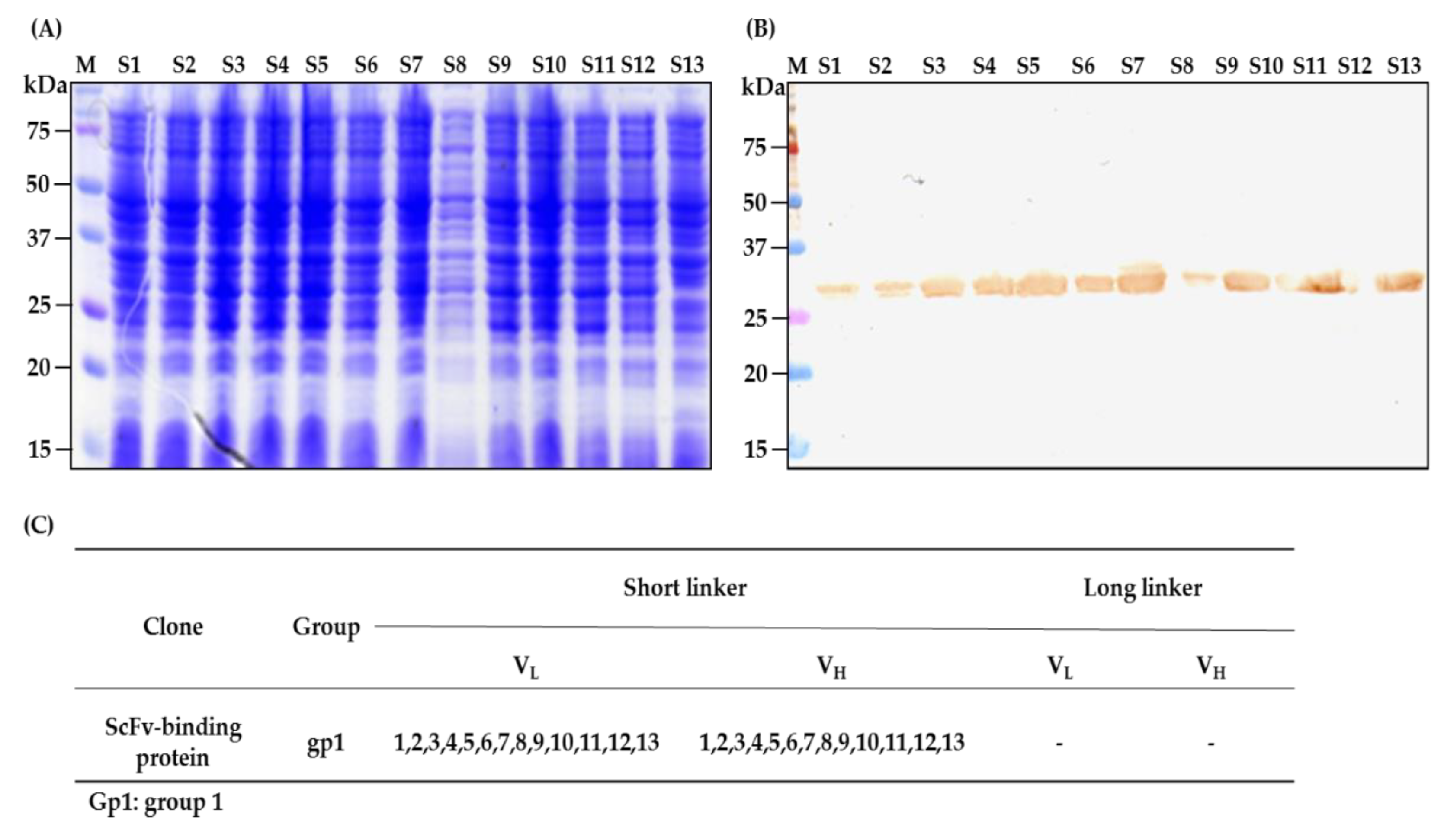
2.4. Characterization of scFv Antibody after Fifth Cycle of Bio-Panning
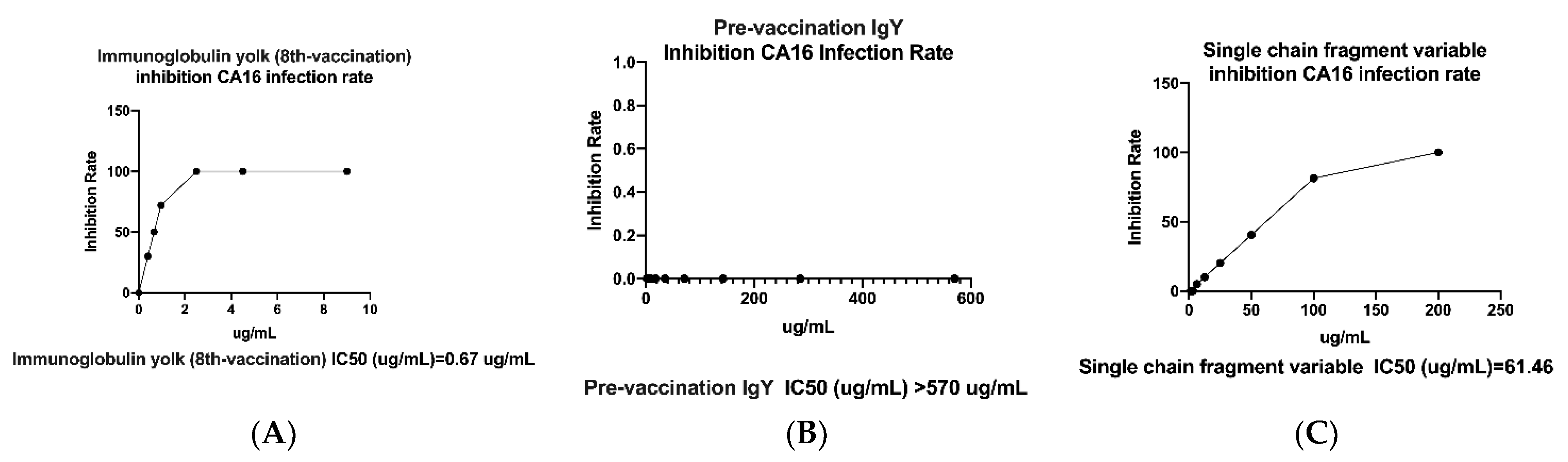
2.5. Neutralization Effect of IgY- and scFv-Binding Protein Antibodies
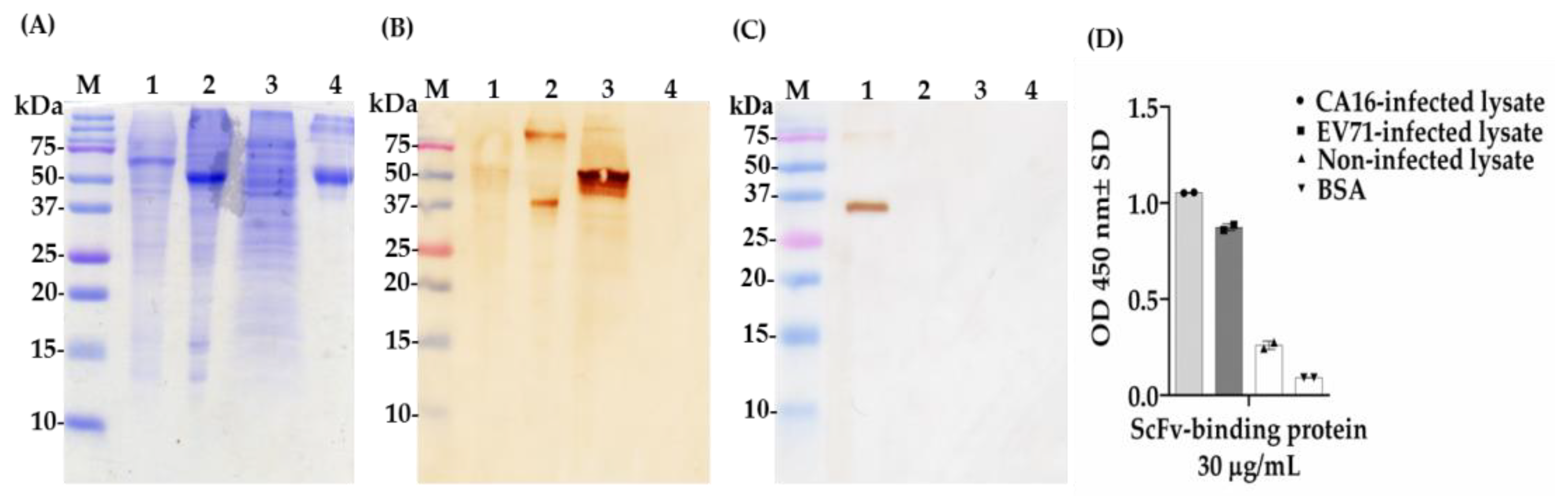
2.6. Determination of Cross-Reactivity of scFv-Binding Protein S1 and IgY against CA16- and EV71-Infected Lysate and BSA
3. Discussion
4. Materials and Methods
4.1. Ethics Declaration
4.2. Cells, Viruses, and Bacteria
4.3. Preparation of CA16-Infected Lysate
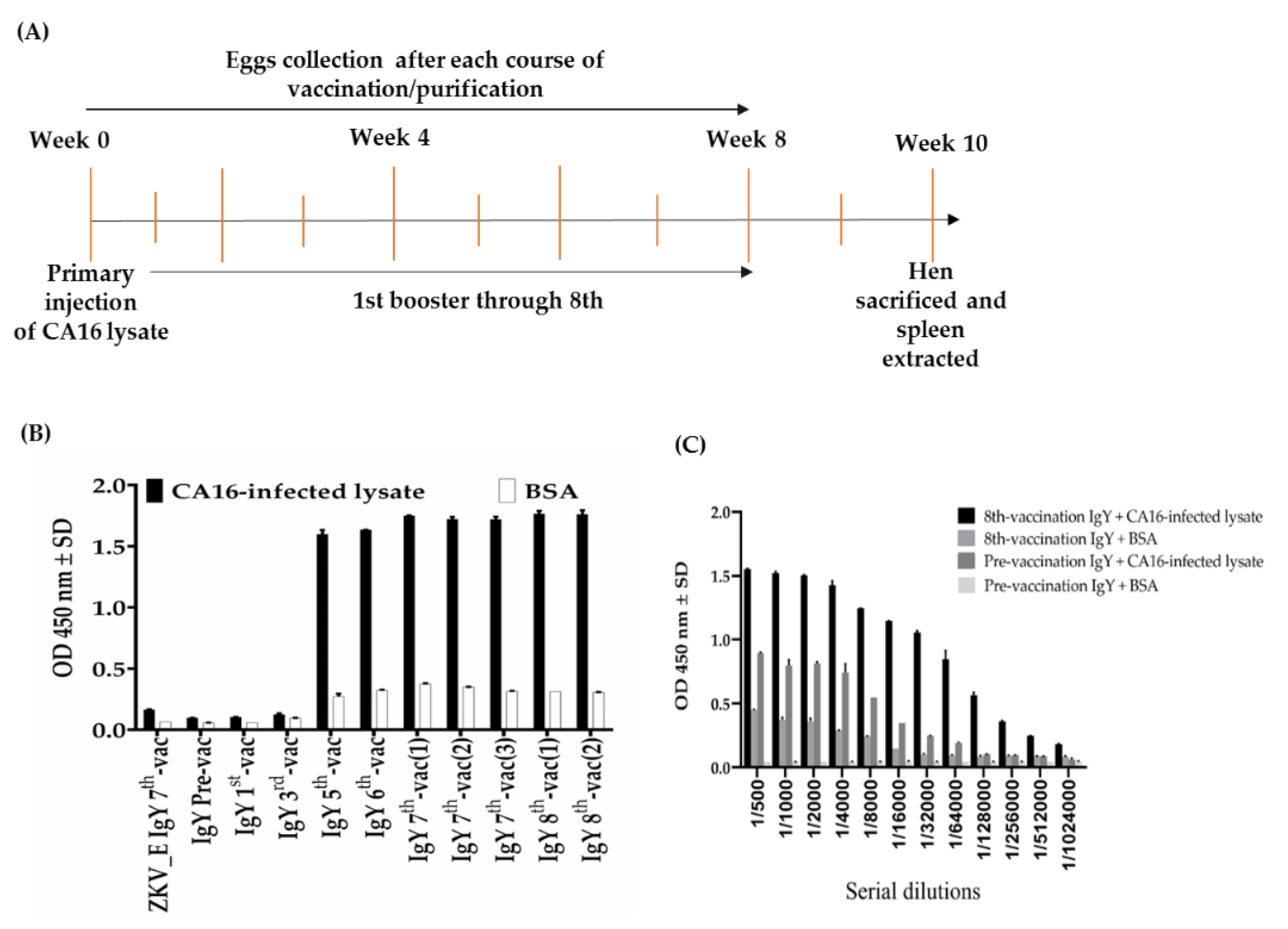
4.4. Hen Vaccination
4.5. Hen IgY Purification from Egg Yolk
4.6. Titration of IgY Antibodies
4.7. RNA Extraction and cDNA Synthesis
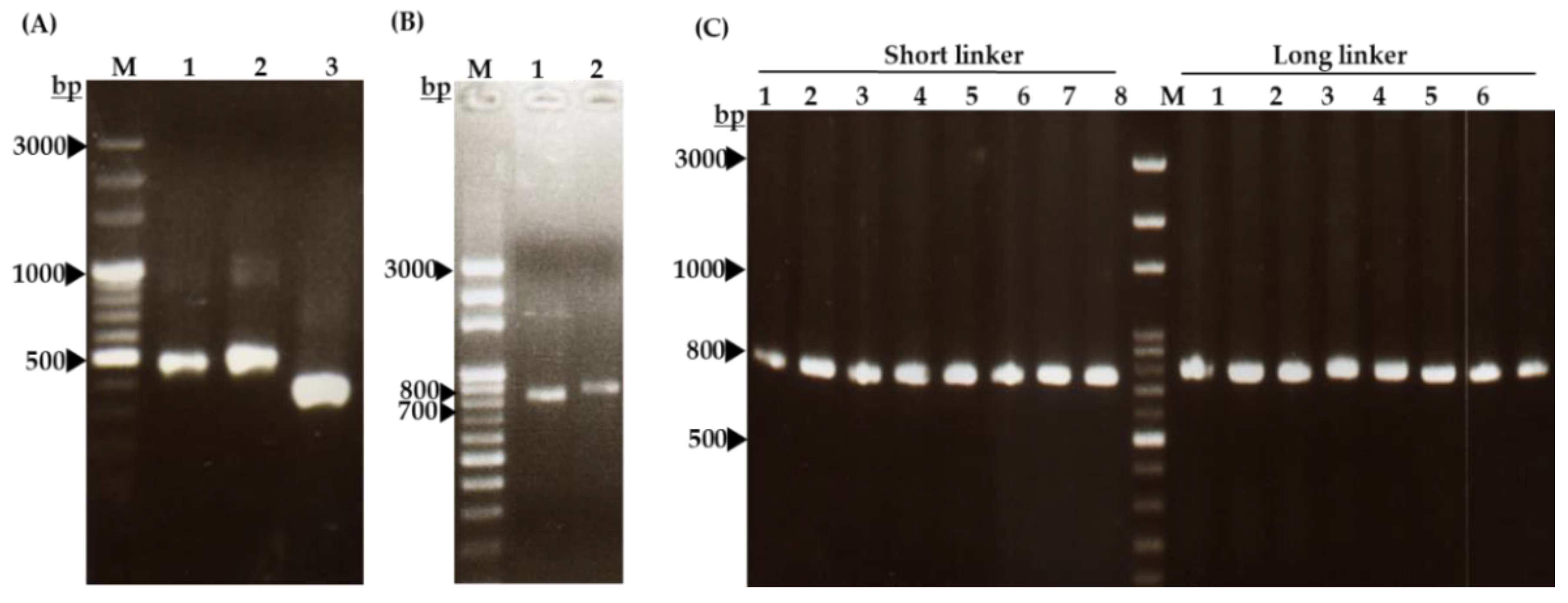
4.8. Preparation of Hen scFv Gene
4.9. Cloning of Hen scFv Gene into pComb3X DNA Vector
4.10. Construction of Phage-Displaying scFv Libraries
4.11. Bio-Panning of scFv-Binding Protein Antibodies and Selection
4.12. SDS-PAGE and Immunoblotting Techniques
4.13. Enzyme-Linked Immunosorbent Assay (ELISA)
4.14. In Vitro scFv-Binding Protein Neutralization Assay
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SCFV | Single-chain variable fragment |
| TMB | 3,3′, 5,5′-tetramethylbenzidene |
| PFU | Plaque-forming unit |
| SD | Standard deviation |
| OD | Optical density |
| µg/mL | Microgram per milliliter |
| % | Percentage |
| M | Molar |
| MgCl2 | Magnesium chloride |
| mg/mL | Milligram per milliliter |
| w/v | Weight per volume |
| pmole | Picomole |
| µL | Microliter |
| ng | Nanogram |
| °C | Degree Celsius |
| PVDF | Polyvinylidene fluoride |
References
- Yu, W.; Xu, H.; Yin, C. Molecular epidemiology of human coxsackievirus A16 strains. Biomed. Rep. 2016, 4, 761–764. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Wang, Y.; Yao, X.; Bian, L.; Wu, X.; Xu, M.; Liang, Z. Coxsackievirus A16: Epidemiology, diagnosis, and vaccine. Hum. Vaccin. Immunother. 2014, 10, 360–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Liu, C.-F.; Yan, L.; Li, J.-J.; Wang, L.-J.; Qi, Y.; Cheng, R.-B.; Xiong, X.-Y. Distribution of enteroviruses in hospitalized children with hand, foot and mouth disease and relationship between pathogens and nervous system complications. Virol. J. 2012, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-Y.; Lu, F.L.; Wu, M.-H.; Lee, C.-Y.; Huang, L.-M. Fatal coxsackievirus A16 infection. Pediatr. Infect. Dis. J. 2004, 23, 275–276. [Google Scholar] [CrossRef]
- Bendig, J.; Fleming, D. Epidemiological, virological, and clinical features of an epidemic of hand, foot, and mouth disease in England and Wales. Commun. Dis. Rep. CDR Rev. 1996, 6, R81–R86. [Google Scholar]
- Chia, M.-Y.; Chiang, P.-S.; Chung, W.-Y.; Luo, S.-T.; Lee, M.-S. Epidemiology of enterovirus 71 infections in Taiwan. Pediatr. Neonatol. 2014, 55, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Han, X.; Wang, G.; Hu, W.; Zhang, W.; Yu, X.-F. Circulating coxsackievirus A16 identified as recombinant type A human enterovirus, China. Emerg. Infect. Dis. 2011, 17, 1537. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Z.; Yang, W.; Ren, J.; Tan, X.; Wang, Y.; Mao, N.; Xu, S.; Zhu, S.; Cui, A. An emerging recombinant human enterovirus 71 responsible for the 2008 outbreak of hand foot and mouth disease in Fuyang city of China. Virol. J. 2010, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.-y.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV71 vaccine, a new tool to control outbreaks of hand, foot and mouth disease (HFMD). Expert Rev. Vaccines 2016, 15, 599–606. [Google Scholar] [CrossRef]
- Luo, S.; Wu, F.; Ye, X.; Fu, T.; Tao, J.; Luo, W.; Wang, Y.; Jia, J.; Lou, L. Safety comparison of two enterovirus 71 (EV71) inactivated vaccines in Yiwu, China. J. Trop. Pediatr. 2019, 65, 547–551. [Google Scholar] [CrossRef]
- Tan, Y.; Chu, J. Sinovac EV71 vaccine: The silver bullet for hand, foot and mouth disease—Or not. J. Public Health Emerg. 2017, 1. [Google Scholar] [CrossRef]
- Marston, H.D.; Paules, C.I.; Fauci, A.S. Monoclonal antibodies for emerging infectious diseases—Borrowing from history. N. Engl. J. Med. 2018, 378, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, H.; Schroten, H.; Tenenbaum, T. Enterovirus Infections of the Central Nervous System in Children: An Update. Pediatr. Infect. Dis. J. 2016, 35, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Niwa, S.; Sekizuka, T.; Tsukagoshi, H.; Yokoyama, M.; Ryo, A.; Sato, H.; Kiyota, N.; Noda, M.; Kozawa, K. Molecular evolution of the VP1, VP2, and VP3 genes in human rhinovirus species C. Sci. Rep. 2015, 5, 8185. [Google Scholar] [CrossRef]
- Lewis, J.K.; Bothner, B.; Smith, T.J.; Siuzdak, G. Antiviral agent blocks breathing of the common cold virus. Proc. Nat. Acad. Sci. USA 1998, 95, 6774–6778. [Google Scholar] [CrossRef] [Green Version]
- Strauss, M.; Filman, D.J.; Belnap, D.M.; Cheng, N.; Noel, R.T.; Hogle, J.M. Nectin-like interactions between poliovirus and its receptor trigger conformational changes associated with cell entry. J. Virol. 2015, 89, 4143–4157. [Google Scholar] [CrossRef] [Green Version]
- Bird, R.E.; Hardman, K.D.; Jacobson, J.W.; Johnson, S.; Kaufman, B.M.; Lee, S.-M.; Lee, T.; Pope, S.H.; Riordan, G.S.; Whitlow, M. Single-chain antigen-binding proteins. Science 1988, 242, 423–426. [Google Scholar] [CrossRef]
- Huston, J.S.; Levinson, D.; Mudgett-Hunter, M.; Tai, M.-S.; Novotný, J.; Margolies, M.N.; Ridge, R.J.; Bruccoleri, R.E.; Haber, E.; Crea, R. Protein engineering of antibody binding sites: Recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc. Nat. Acad. Sci. USA 1988, 85, 5879–5883. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, R.; Chen, X. Application of a single-chain fragment variable (scFv) antibody for the confirmatory diagnosis of hydatid disease in non-endemic areas. Electron. J. Biotechnol. 2017, 29, 57–62. [Google Scholar] [CrossRef]
- Krag, D.N.; Shukla, G.S.; Shen, G.-P.; Pero, S.; Ashikaga, T.; Fuller, S.; Weaver, D.L.; Burdette-Radoux, S.; Thomas, C. Selection of tumor-binding ligands in cancer patients with phage display libraries. Cancer Res. 2006, 66, 7724–7733. [Google Scholar] [CrossRef] [Green Version]
- Unkauf, T.; Miethe, S.; Fühner, V.; Schirrmann, T.; Frenzel, A.; Hust, M. Generation of recombinant antibodies against toxins and viruses by phage display for diagnostics and therapy. In Protein Targeting Compounds; Springer: New York, NY, USA, 2016; pp. 55–76. [Google Scholar]
- Corti, D.; Voss, J.; Gamblin, S.J.; Codoni, G.; Macagno, A.; Jarrossay, D.; Vachieri, S.G.; Pinna, D.; Minola, A.; Vanzetta, F. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science 2011, 333, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Milenic, D.E.; Whitlow, M.; Schlom, J. Rapid tumor penetration of a single-chain Fv and comparison with other immunoglobulin forms. Cancer Res. 1992, 52, 3402–3408. [Google Scholar] [PubMed]
- Pyo, H.-M.; Kim, I.-J.; Kim, S.-H.; Kim, H.-S.; Cho, S.-D.; Cho, I.-S.; Hyun, B.-H. Escherichia coli expressing single-chain Fv on the cell surface as a potential prophylactic of porcine epidemic diarrhea virus. Vaccine 2009, 27, 2030–2036. [Google Scholar] [CrossRef]
- Harmsen, M.; Fijten, H.; Engel, B.; Dekker, A.; Eblé, P. Passive immunization with llama single-domain antibody fragments reduces foot-and-mouth disease transmission between pigs. Vaccine 2009, 27, 904–1911. [Google Scholar] [CrossRef]
- Bird, C.R.; Thorpe, R. Purification of immunoglobulin Y (IgY) from chicken eggs. In The Protein Protocols Handbook; Springer: New York, NY, USA, 2009; pp. 1779–1781. [Google Scholar]
- Deng, X.K.; Nesbit, L.A.; Morrow, K.J. Recombinant Single-Chain Variable Fragment Antibodies Directed against Clostridium difficile Toxin B Produced by Use of an Optimized Phage Display System. Clin. Diagn. Lab. Immunol. 2003, 10, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Fehrsen, J.; Wemmer, S.; van Wyngaardt, W. Construction of Chicken Antibody Libraries. In Phage Display: Methods and Protocols; Hust, M., Lim, T.S., Eds.; Springer: New York, NY, USA, 2018; pp. 189–203. [Google Scholar]
- Yang, Q.; Ding, J.; Cao, J.; Huang, Q.; Hong, C.; Yang, B. Epidemiological and etiological characteristics of hand, foot, and mouth disease in Wuhan, China from 2012 to 2013: Outbreaks of coxsackieviruses A10. J. Med. Virol. 2015, 87, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Nilvebrant, J.; Sidhu, S.S. Construction of synthetic antibody phage-display libraries. In Phage Display; Springer: New York, NY, USA, 2018; pp. 45–60. [Google Scholar]
- Da Silva, M.C.; Schaefer, R.; Gava, D.; Souza, C.K.; da Silva Vaz, I., Jr.; Bastos, A.P.; Venancio, E.J. Production and application of anti-nucleoprotein IgY antibodies for influenza A virus detection in swine. J. Immunol. Methods 2018, 461, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Leu, S.-J.; Lee, Y.-C.; Liu, C.-I.; Lin, L.-T.; Mwale, P.F.; Chiang, J.-R.; Tsai, B.-Y.; Chen, C.-C.; Hung, C.-S. Characterization of chicken-derived single chain antibody fragments against venom of Naja naja atra. Toxins 2018, 10, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Liu, Q.; Huang, X.; Li, D.; Ku, Z.; Zhang, Y.; Huang, Z. Active immunization with a Coxsackievirus A16 experimental inactivated vaccine induces neutralizing antibodies and protects mice against lethal infection. Vaccine 2013, 31, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, Q.; Cai, Y.; Leng, Q.; Huang, Z. Construction and characterization of an infectious clone of coxsackievirus A16. Virol. J. 2011, 8, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.-W.; Cheng, S.-F.; Tseng, Y.-T.; Yang, Y.-C.; Liu, W.-C.; Wang, S.-C.; Chou, M.-J.; Lin, Y.-J.; Wang, Y.; Hsiao, P.-W. Development of single-chain variable fragments (scFv) against influenza virus targeting hemagglutinin subunit 2 (HA2). Arch. Virol. 2016, 161, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Mao, Q.; Hu, Y.; Lang, S.; Sun, S.; Li, K.; Gao, F.; Bian, L.; Yang, C.; Cui, B.; et al. A potential therapeutic neutralization monoclonal antibody specifically against multi-coxsackievirus A16 strains challenge. Hum. Vaccines Immunother. 2019, 15, 2343–2350. [Google Scholar] [CrossRef]
- Zhang, W.; Dai, W.; Zhang, C.; Zhou, Y.; Xiong, P.; Wang, S.; Ye, X.; Liu, Q.; Zhou, D.; Huang, Z. A virus-like particle-based tetravalent vaccine for hand, foot, and mouth disease elicits broad and balanced protective immunity. Emerg. Microbes Infect. 2018, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, X.; Li, X.; Ma, Z.; Feng, R. Construction, expression, and characterization of a single-chain variable fragment (ScFv) antibody targeting to the encephalomyocarditis virus. J. Med. Virol. 2018, 90, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Mwale, P.F.; Lee, C.-H.; Leu, S.-J.; Lee, Y.-C.; Wu, H.-H.; Lin, L.-T.; Lin, T.E.; Huang, Y.-J.; Yang, Y.-Y. Antigenic epitopes on the outer membrane protein A of Escherichia coli identified with single-chain variable fragment (scFv) antibodies. Appl. Microbiol. Biotechnol. 2019, 103, 5285–5299. [Google Scholar] [CrossRef]
- Leu, S.-J.; Lee, Y.-C.; Shih, N.-Y.; Huang, I.-J.; Liu, K.-J.; Lu, H.-F.; Huang, S.-Y.; Yang, Y.-Y. Generation and characterization of anti-α-enolase single-chain antibodies in chicken. Vet. Immunol. Immunopathol. 2010, 137, 251–260. [Google Scholar] [CrossRef]
- Terpe, K. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2006, 72, 211. [Google Scholar] [CrossRef]
- Tikunova, N.; Morozova, V. Phage display on the base of filamentous bacteriophages: Application for recombinant antibodies selection. Acta Nat. 2009, 1, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Backman, L.; Persson, K. The No-Nonsens SDS-PAGE. Methods Mol. Biol. 2018, 1721, 89–94. [Google Scholar]
- Engvall, E.; Perlmann, P. Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Immunochemistry 1971, 8, 871–874. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amount of Antigen (μg, CA16-Infected Lysate) | Volume of 1× PBS (μL) | Adjuvants (500 μL) | |
|---|---|---|---|
| 1st-vaccination | 100 | 500 | Complete freund’s adjuvant |
| 2nd-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 3rd-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 4th-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 5th-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 6th-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 7th-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| 8th-vaccination | 100 | 500 | Incomplete freund’s adjuvant |
| ScFv Clone | Region | Diff/totl (%) * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FR1 | FR2 | FR3 | FR4 | Total FRs | CDR1 | CDR2 | CDR3 | Total CDRs | ||
| ScFv-binding protein S1 | VL | 1/20 (5) | 1/16 (6.25) | 6/32 (18.75) | 0/10 (0) | 8/78 (10.25) | 7/13 (53.85) a | 2/7 (28.57) | 4/11 (36.36) | 13/31 (41.94) a |
| VH | 3/30 (10) | 0/14 (0) | 3/32 (9.38) | 0/11 (0) | 6/87 (6.90) | 2/5(40) | 8/17 (47.06) | 12/15 (80) a | 22/37 (59.46) a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwale, P.F.; Lee, C.-H.; Huang, P.-N.; Tseng, S.-N.; Shih, S.-R.; Huang, H.-Y.; Leu, S.-J.; Huang, Y.-J.; Chiang, L.-C.; Mao, Y.-C.; et al. In Vitro Characterization of Neutralizing Hen Antibodies to Coxsackievirus A16. Int. J. Mol. Sci. 2021, 22, 4146. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084146
Mwale PF, Lee C-H, Huang P-N, Tseng S-N, Shih S-R, Huang H-Y, Leu S-J, Huang Y-J, Chiang L-C, Mao Y-C, et al. In Vitro Characterization of Neutralizing Hen Antibodies to Coxsackievirus A16. International Journal of Molecular Sciences. 2021; 22(8):4146. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084146
Chicago/Turabian StyleMwale, Pharaoh Fellow, Chi-Hsin Lee, Peng-Nien Huang, Sung-Nien Tseng, Shin-Ru Shih, Hsin-Yuan Huang, Sy-Jye Leu, Yun-Ju Huang, Liao-Chun Chiang, Yan-Chiao Mao, and et al. 2021. "In Vitro Characterization of Neutralizing Hen Antibodies to Coxsackievirus A16" International Journal of Molecular Sciences 22, no. 8: 4146. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084146