
Salinity Stress Alters the Secondary Metabolic Profile of M. sativa, M. arborea and Their Hybrid (Alborea)

 ,
,
Abstract
:1. Introduction
2. Results
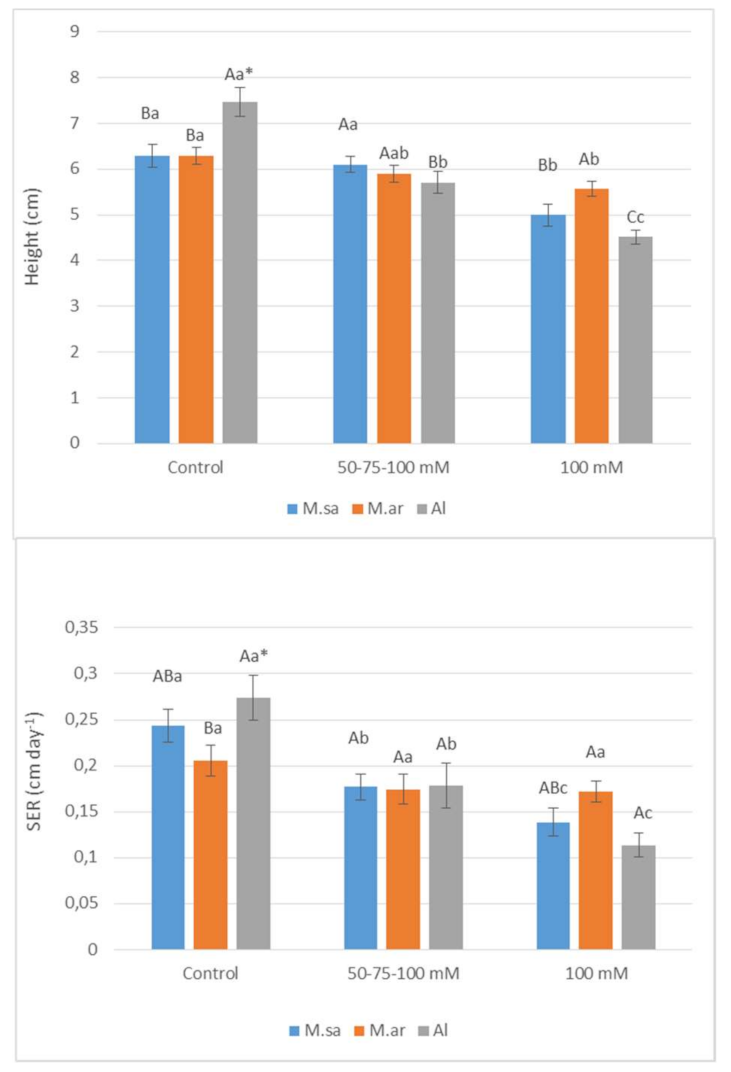
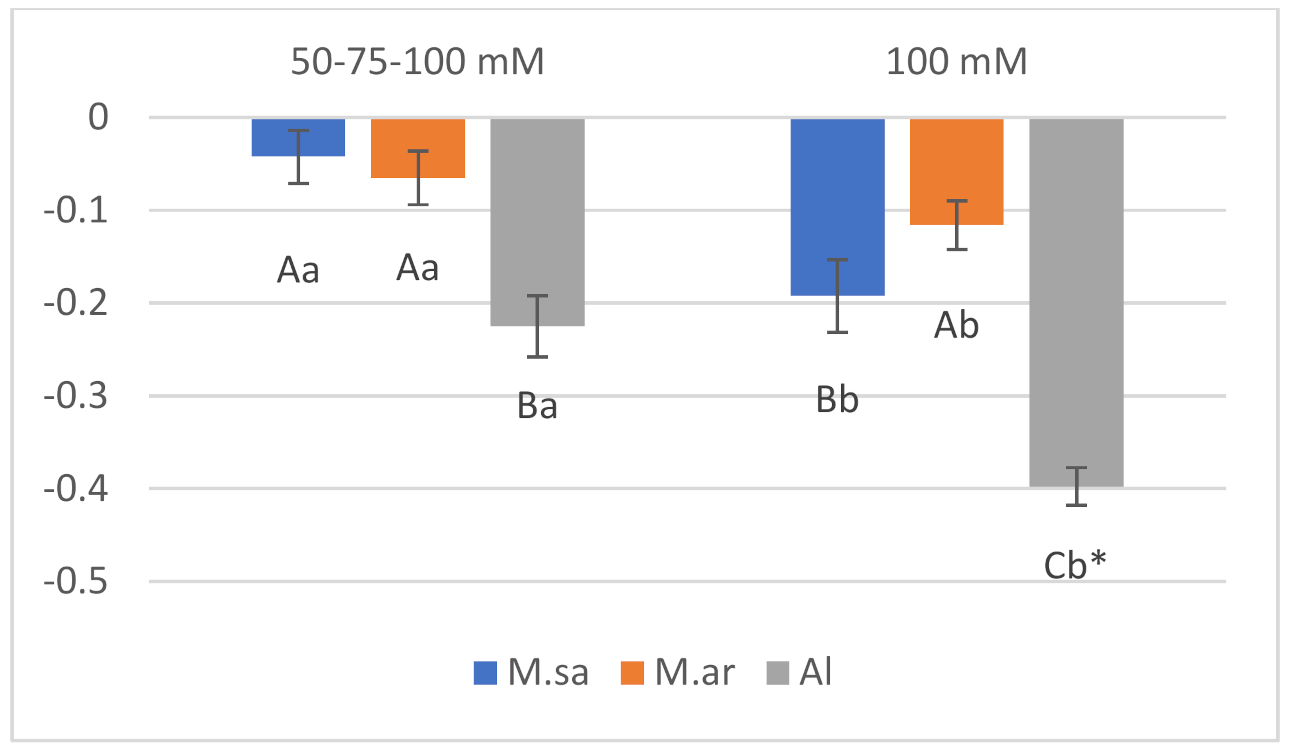
2.1. Growth Parameters and Salinity Sensitivity Index
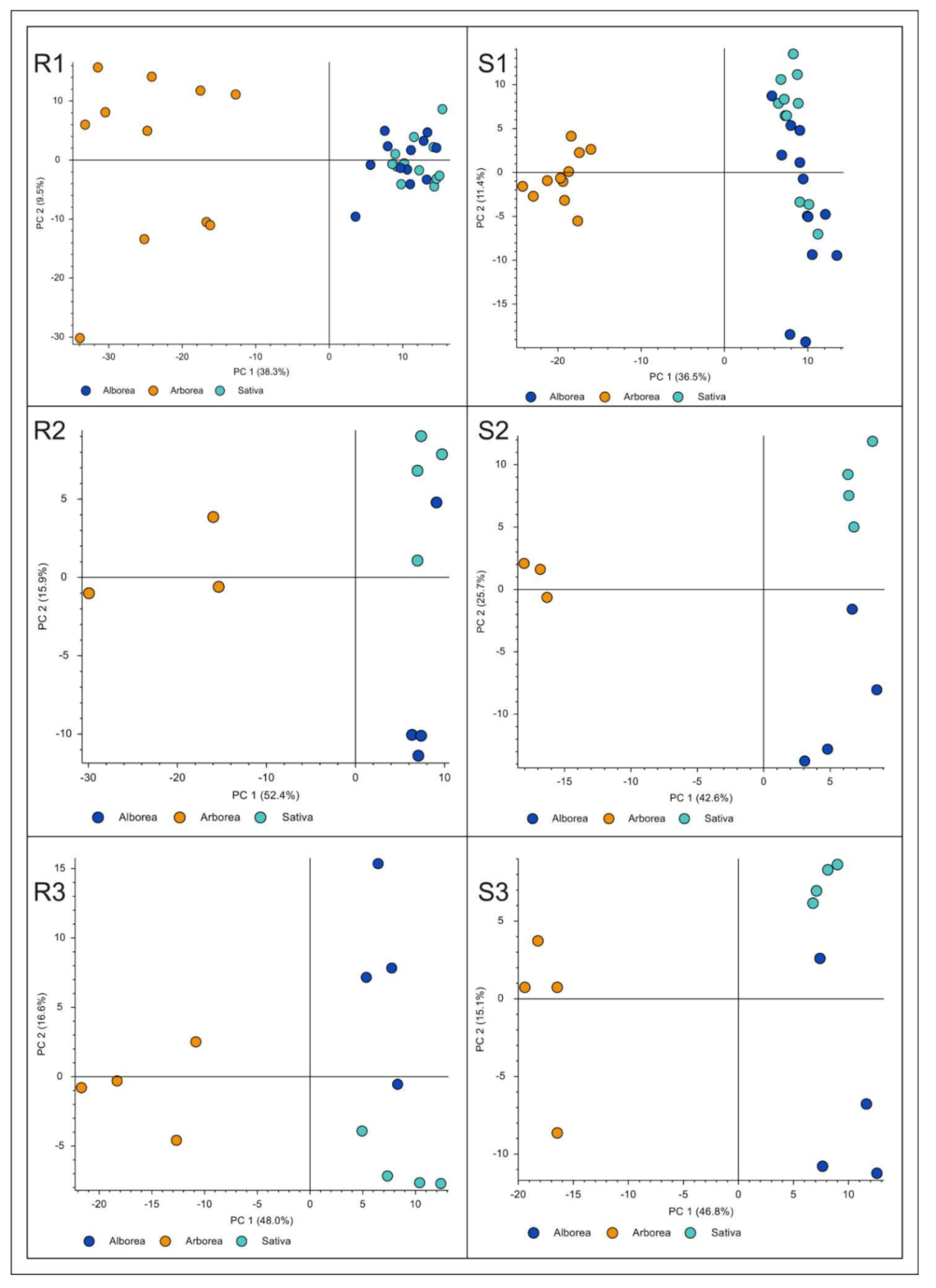
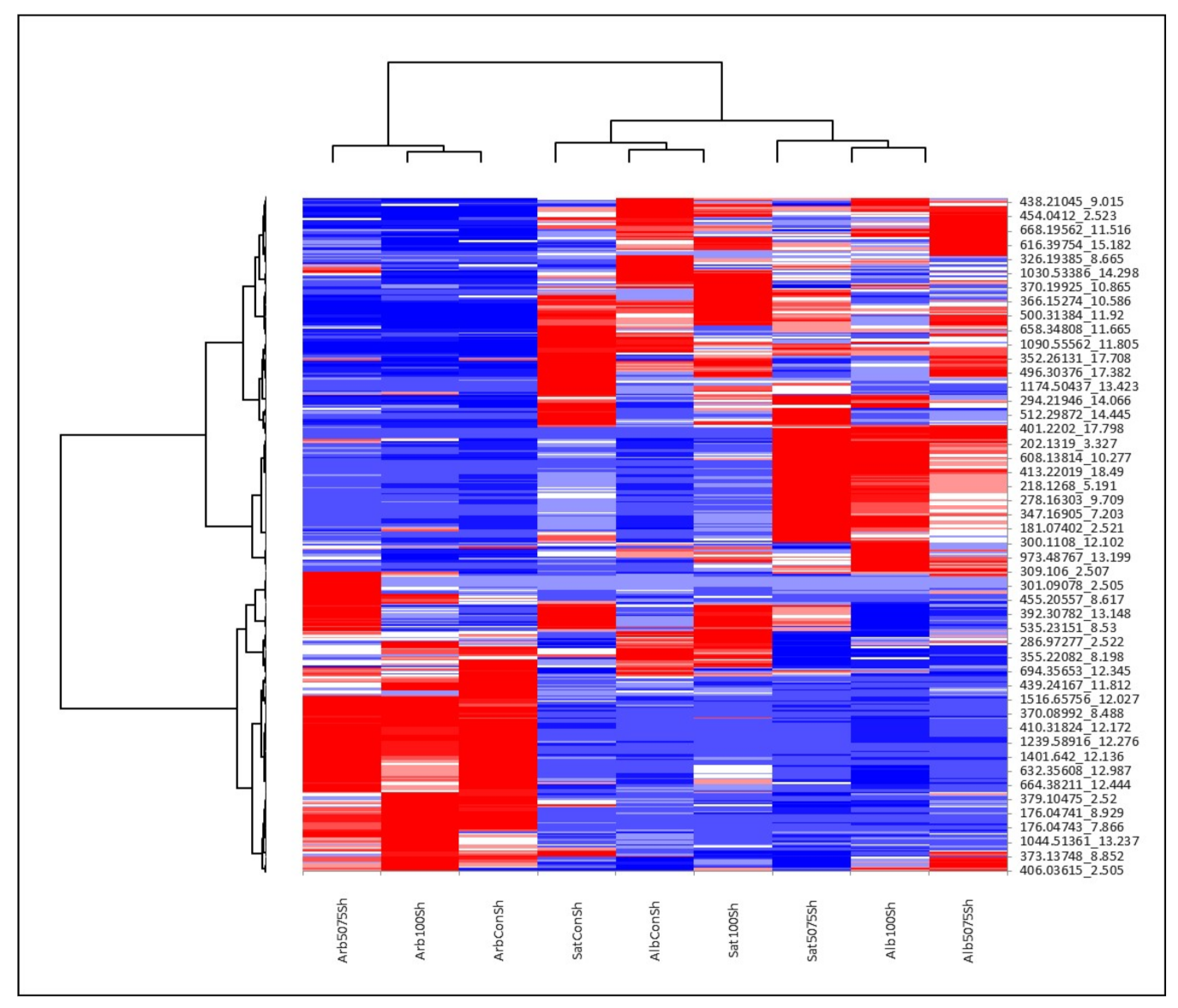
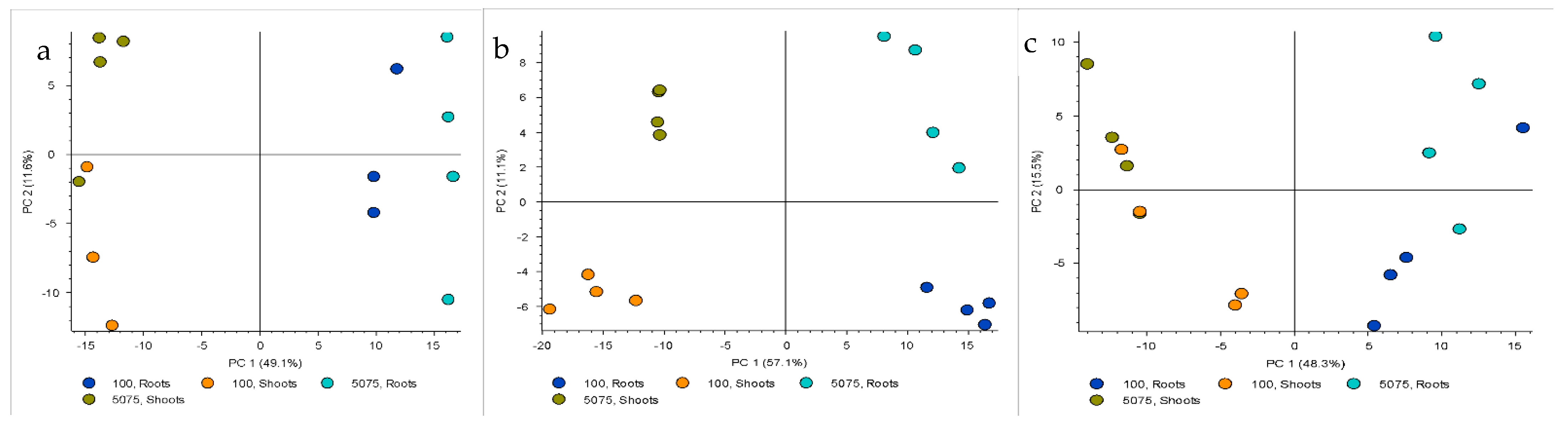
2.2. Metabolomics Analysis
2.2.1. Comparison among the Entries
2.2.2. Comparison between Roots and Shoots
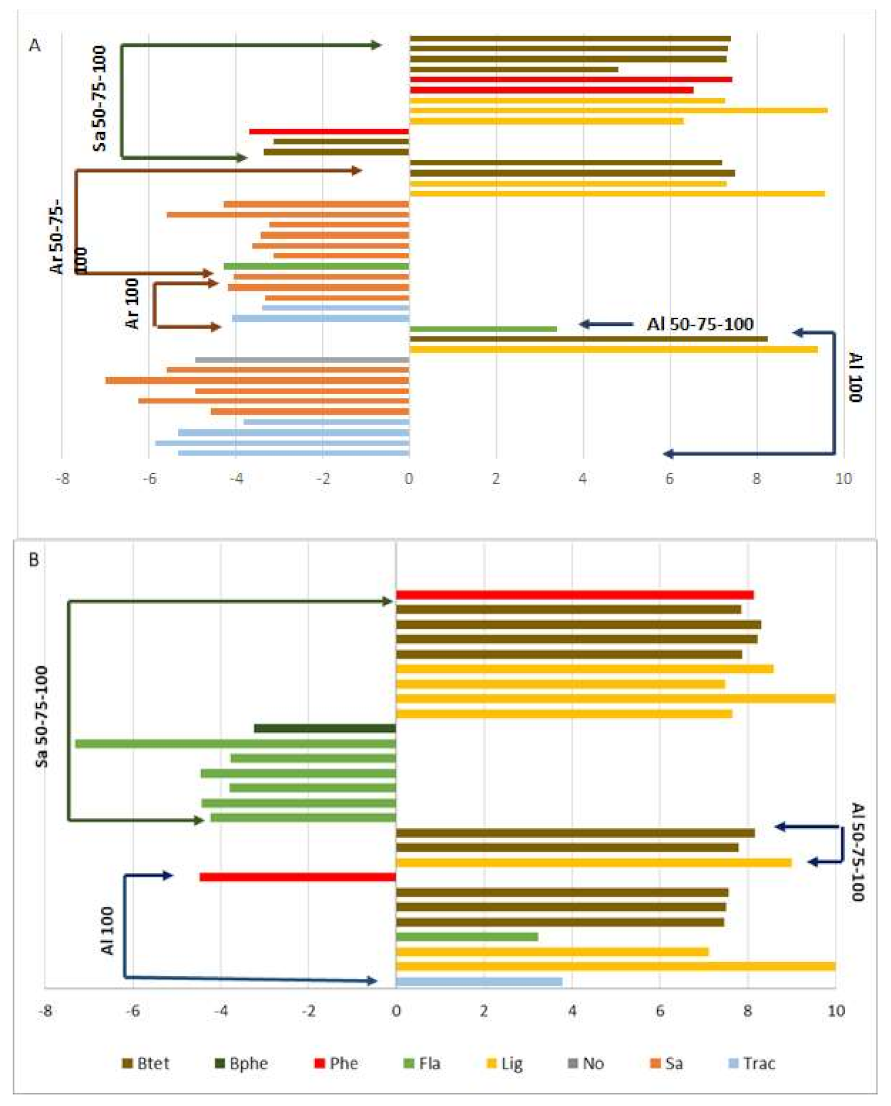
2.3. Secondary Metabolomic Changes in Response to Salt Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Seed Pretreatment
4.3. Growth Conditions
4.4. Salt Stress Treatments
4.5. Growth Characteristics Measurements
4.6. Metabolites Extraction
4.7. Metabolomic Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2004; pp. 3–20. [Google Scholar]
- FAO. State of The Worlds Forests; FAO: Rome, Italy, 1997. [Google Scholar]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Chandna, R.; Azooz, M.; Ahmad, P. Recent advances of metabolomics to reveal plant response during salt stress. Salt Stress Plants Signal. Omics Adapt. 2014, 1–14. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yang, Y. Germination, growth, osmotic adjustment and ionic balance of wheat in response to saline and alkaline stresses. Soil Sci. Plant. Nutr. 2009, 55, 667–679. [Google Scholar] [CrossRef]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucini, L.; Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Bernardi, J.; Colla, G. Mild potassium chloride stress alters the mineral composition, hormone network, and phenolic profile in artichoke leaves. Front. Plant Sci. 2016, 7, 948. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, A.; Kudapa, H.; Pazhamala, L.T.; Weckwerth, W.; Varshney, R.K. Proteomics and Metabolomics: Two emerging areas for legume improvement. Front. Plant Sci 2015, 6, 1116. [Google Scholar] [CrossRef] [Green Version]
- Borgognone, D.; Cardarelli, M.; Rea, E.; Lucini, L.; Colla, G. Salinity source-induced changes in yield, mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon grown in floating system. J. Sci. Food Agric. 2014, 94, 1231–1237. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant. Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Waskiewicz, A.; Muzolf-Panek, M.; Golinski, P. Phenolic Content Changes in Plants under Salt Stress. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013; pp. 283–314. [Google Scholar]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.L.; Aktas, L. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant. Physiol. 2007, 34, 67–78. [Google Scholar]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant. Signal. Behav 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.; Iqbal, S.; Qi, Z.; Saddiq, M.; Jacobsen, S.-E. Saponin seed priming improves salt tolerance in quinoa. J. Agron. Crop. Sci. 2017, 204. [Google Scholar] [CrossRef]
- Fukushima, A.; Kusano, M. Recent progress in the development of metabolome databases for plant systems biology. Front. Plant Sci 2013, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Chen, H.; Chen, J.; Yang, R.; Zou, L.; Wang, C.; Chen, J.; Tan, M.; Mei, Y.; Wei, L.; et al. Metabolomics characterizes the metabolic changes of Lonicerae Japonicae Flos under different salt stresses. PLoS ONE 2020, 15, e0243111. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Jha, R.; Parida, S.K. Salinity stress response and ‘omics’ approaches for improving salinity stress tolerance in major grain legumes. Plant. Cell Rep. 2019, 38, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Baligar, V.C.; Bailey, B.A. Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant. Anal. 2005, 36, 2733–2757. [Google Scholar] [CrossRef]
- Rafińska, K.; Pomastowski, P.; Wrona, O.; Górecki, R.; Buszewski, B. Medicago sativa as a source of secondary metabolites for agriculture and pharmaceutical industry. Phytochem. Lett. 2017, 20, 520–539. [Google Scholar] [CrossRef]
- Lefi, E.; Conesa, M.À.; Cifre, J.; Gulías, J.; Medrano, H. Dry matter allocation in Medicago arborea and Medicago citrina in response to drought and defoliation. Crop. Pasture Sci. 2012, 63, 179–189. [Google Scholar] [CrossRef]
- Tani, E.; Sarri, E.; Goufa, M.; Asimakopoulou, G.; Psychogiou, M.; Bingham, E.; Skaracis, G.N.; Abraham, E.M. Seedling growth and transcriptional responses to salt shock and stress in Medicago sativa L., Medicago arborea L., and their hybrid (Alborea). Agronomy 2018, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- Bingham, E.; Armour, D.; Irwin, J. The hybridization barrier between herbaceous Medicago sativa and woody M. arborea is weakened by selection of seed parents. Plants 2013, 2, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Barsch, A.; Carvalho, H.G.; Cullimore, J.V.; Niehaus, K. GC-MS based metabolite profiling implies three interdependent ways of ammonium assimilation in Medicago truncatula root nodules. J. Biotechnol. 2006, 127, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Kera, K.; Fine, D.D.; Wherritt, D.J.; Nagashima, Y.; Shimada, N.; Ara, T.; Ogata, Y.; Sumner, L.W.; Suzuki, H. Pathway-specific metabolome analysis with (18)O(2)-labeled Medicago truncatula via a mass spectrometry-based approach. Metabolomics 2018, 14, 71. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, E.; Rusilowicz, M.J.; Dickinson, M.; Charlton, A.J.; Bechtold, U.; Mullineaux, P.M.; Wilson, J. Integrating transcriptomic techniques and k-means clustering in metabolomics to identify markers of abiotic and biotic stress in Medicago truncatula. Metabolomics 2018, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Essemine, J.; Shang, C.; Zhang, H.; Zhu, X.; Yu, J.; Chen, G.; Qu, M.; Sun, D. Combined proteomics and metabolism analysis unravels prominent roles of antioxidant system in the prevention of Alfalfa (Medicago sativa L.) against salt stress. Int. J. Mol. Sci. 2020, 21, 909. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Long, R.; Kang, J.; Wang, Z.; Zhang, T.; Sun, H.; Li, X.; Yang, Q. Comparative proteomic analysis reveals that antioxidant system and soluble sugar metabolism contribute to salt tolerance in Alfalfa (Medicago sativa L.) leaves. J. Proteome Res. 2019, 18, 191–203. [Google Scholar] [CrossRef]
- Kang, Y.; Torres-Jerez, I.; An, Z.; Greve, V.; Huhman, D.; Krom, N.; Cui, Y.; Udvardi, M. Genome-wide association analysis of salinity responsive traits in Medicago truncatula. Plant Cell Environ. 2019, 42, 1513–1531. [Google Scholar] [CrossRef] [Green Version]
- Boughalleb, F.; Denden, M.; Tiba, B.B. Photosystem II photochemistry and physiological parameters of three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea under salt stress. Acta Physiol. Plant. 2009, 31, 463–476. [Google Scholar] [CrossRef]
- Boughalleb, F.; Denden, M.; Tiba, B.B. Anatomical changes induced by increasing NaCl salinity in three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea. Acta Physiol. Plant. 2009, 31, 947–960. [Google Scholar] [CrossRef]
- Aissat, A.; Mehdadi, Z.; Leogrande, R.; Stellacci, A.M. Characterization of Medicago arborea L. response to water and salt stress. J. Stress Physiol. Biochem. 2019, 15, 103–113. [Google Scholar]
- Bennett, R.; Wallsgrove, R. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Carelli, M.; Biazzi, E.; Tava, A.; Losini, I.; Abbruscato, P.; Depedro, C.; Scotti, C. Sapogenin content variation in Medicago inter-specific hybrid derivatives highlights some aspects of saponin synthesis and control. New Phytol. 2015, 206, 303–314. [Google Scholar] [CrossRef]
- Faizal, A.; Geelen, D. Saponins and their role in biological processes in plants. Phytochem. Rev. 2013, 12, 877–893. [Google Scholar] [CrossRef]
- Mazahery-Laghab, H.; Yazdi-Samadi, B.; Bagheri, M.; Bagheri, A.R. Alfalfa (Medicago sativa L.) shoot saponins: Identification and bio-activity by the assessment of aphid feeding. Br. J. Nutr. 2011, 105, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [Green Version]
- Basyuni, M.; Baba, S.; Kinjo, Y.; Putri, L.A.; Hakim, L.; Oku, H. Salt-dependent increase in triterpenoids is reversible upon transfer to fresh water in mangrove plants Kandelia candel and Bruguiera gymnorrhiza. J. Plant. Physiol. 2012, 169, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pando, L.; Álvarez-Castro, R.; Eguiluz de la Barra, A. Short Communication: Effect of salt stress on Peruvian germplasm of Chenopodium quinoa Willd.: A promising crop. J. Agron. Crop. Sci. 2010, 196, 391–396. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristics of quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Oku, H.; Baba, S.; Koga, H.; Takara, K.; Iwasaki, H. Lipid composition of mangrove and its relevance to salt tolerance. J. Plant Res. 2003, 116, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guo, R.; Jiao, Y.; Jin, X.; Zhang, H.; Shi, L. Comparison of salt tolerance in soja based on metabolomics of seedling roots. Front. Plant Sci. 2017, 8, 1101. [Google Scholar] [CrossRef] [PubMed]
- Widodo; Patterson, J.H.; Newbigin, E.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 4089–4103. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, D.; Li, M.; Shi, L. Metabolic profiles reveal changes in wild and cultivated soybean seedling leaves under salt stress. PLoS ONE 2016, 11, e0159622. [Google Scholar] [CrossRef] [Green Version]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Placines, C.; Castañeda-Loaiza, V.; João Rodrigues, M.; Pereira, C.G.; Stefanucci, A.; Mollica, A.; Zengin, G.; Llorent-Martínez, E.J.; Castilho, P.C.; Custódio, A.L. Phenolic profile, toxicity, enzyme inhibition, in silico studies, and antioxidant properties of Cakile maritima Scop. (Brassicaceae) from Southern Portugal. Plants 2020, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Hossain, M.S.; Mahmud, J.A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced salt stress tolerance in rice seedlings: Regulation of ion homeostasis, antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2016, 22, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; De, B. Differential responses of cell wall bound phenolic compounds in sensitive and tolerant varieties of rice in response to salinity. Plant Signal. Behav. 2017, 12, e1379643. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, S.; Taniguchi, E. Synthesis and insecticidal activity of lignan analogs (II). Biosci. Biotechnol. Biochem. 1992, 56, 412–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Farid, I.B.; Marghany, M.R.; Rowezek, M.M.; Sheded, M.G. Effect of salinity stress on growth and metabolomic profiling of Cucumis sativus and Solanum lycopersicum. Plants 2020, 9, 1626. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Ali, Q. Response of two genetically diverse wheat cultivars to salt stress at different growth stages: Leaf lipid peroxidation and phenolic contents. Pak. J. Bot. 2010, 42, 559–565. [Google Scholar]
- Haghighi, Z.; Karimi, N.; Modarresi, M.; Mollaei, S. Enhancement of compatible solute and secondary metabolites production in Plantago ovata Forsk. by salinity stress. J. Med. Plants Res. 2012, 6, 3495–3500. [Google Scholar]
- Mohamad, A.H.; Marina, M.F.; Francisco José, R.S.; Oscar, V.; Monica, B. Effects of salt and water stress on plant growth and on accumulation of osmolytes and antioxidant compounds in cherry tomato. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43. [Google Scholar] [CrossRef]
- Irwin, J.; Sewell, J.; Woodfield, D.; Bingham, E. Restructuring Lucerne (Medicago sativa) through introgression of the Medicago arborea genome. Agric. Sci. 2015, 28, 40–46. [Google Scholar]
- Hamrouni, L.; Abdallah, F.B.; Abdelly, C.; Ghorbel, A. In vitro culture: A simple and efficient way for salt-tolerant grapevine genotype selection. C. R. Biol. 2008, 331, 152–163. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Height | SER | Salinity Sensitivity Index |
|---|---|---|---|
| Salt (A) | p ˂ 0.05 | p ˂ 0.05 | p ˂ 0.05 |
| Species (B) | ns | ns | p ˂ 0.05 |
| Dates (C) | p ˂ 0.05 | p ˂ 0.05 | p ˂ 0.05 |
| AXB (Interaction) | p ˂ 0.05 | p ˂ 0.05 | p ˂ 0.05 |
| AXC (Interaction) | p ˂ 0.05 | ns | ns |
| BXC (Interaction) | ns | p ˂ 0.05 | p ˂ 0.05 |
| AXBXC (Interaction) | ns * | p ˂ 0.05 | ns |
| a. For Aerial Parts (Shoots) | ||||||
|---|---|---|---|---|---|---|
| RT [min] | Molecular Weight | Formula | Category | Log2 Fold Change | Upregulated | VIP Score |
| 7.78 | 356.0742 | C15H16O10 | Caffeic acid derivative | 8.01 | Arb | 1.08 |
| 8.28 | 194.0579 | C10H10O4 | Ferulic acid | 6.93 | Arb | 1.52 |
| 8.31 | 370.0899 | C16H18O10 | Ferulic acid derivative | 7.8 | Arb | 1.07 |
| 9.39 | 798.1487 | C33H34O23 | Flavonoid (apigenin derivative) | 10.08 | Arb | 7.59 |
| 9.55 | 828.1593 | C34H36O24 | Flavonoid | 7.61 | Arb | 1.08 |
| 10.32 | 828.1746 | C38H36O21 | Flavonoid | 9.36 | Arb | 2.39 |
| 12.12 | 1090.5189 | C52H82O24 | Saponin | 4.82 | Arb | 3.80 |
| 12.89 | 1206.5670 | C57H90O27 | Saponin | 6.34 | Arb | 2.07 |
| 13.44 | 1190.5720 | C57H90O26 | Saponin | 4.77 | Arb | 2.07 |
| 15.05 | 1014.5392 | C51H82O20 | Triterpenic saponin | 7.22 | Arb | 1.52 |
| 15.33 | 912.5081 | C47H76O17 | Soyasaponin | 4.84 | Arb | 1.98 |
| 16.21 | 910.4925 | C47H74O17 | Triterpenic saponin | 6.15 | Arb | |
| 8.50 | 432.1267 | C18H24O12 | Iridoid | −6.49 | Sat, Alb | 8.68 |
| 9.04 | 622.1170 | C27H26O17 | Flavonoid | −6.98 | Sat, Alb | 3.58 |
| 9.59 | 974.1958 | C43H42O26 | Flavonoid | −9.21 | Alb | 5.02 |
| 11.07 | 798.1640 | C37H34O20 | Flavonoid | −6.71 | Sat, Alb | 5.69 |
| 11.19 | 1106.5505 | C53H86O24 | Triterpenic saponin | −7.84 | Sat | 1.01 |
| 11.43 | 768.1534 | C36H32O19 | Flavonoid | −6.89 | Sat | 1.34 |
| 11.63 | 1090.5555 | C53H86O23 | Triterpenic saponin | −8.71 | Sat, Alb | 2.47 |
| 11.92 | 1414.6249 | C64H102O34 | Triterpenic saponin | −6.62 | Sat, Alb | 1.20 |
| 11.53 | 1118.5510 | C54H86O24 | Triterpenic saponin | −7.21 | Alb | 1.41 |
| 11.74 | 1546.6690 | C62H114O43 | Triterpenic saponin | −7.06 | Sat, Alb | 1.78 |
| 11.75 | 1414.6250 | C57H106O39 | Triterpenic saponin | −5.72 | Alb | 1.04 |
| 14.98 | 912.5081 | C47H76O17 | Triterpenic saponin | −7.9 | Sat, Alb | 1.98 |
| 15.39 | 1028.5190 | C51H80O21 | Triterpenic saponin | −7.32 | Sat, Alb | 1.29 |
| 15.79 | 488.2197 | C30H32O6 | Triterpenic acid | −10.57 | Sat | 4.04 |
| b. Roots | ||||||
| RT [min] | Molecular Weight | Formula | Category | Log2 Fold Change | Upregulated in | VIP Score |
| 12.38 | 664.3829 | C36H56O11 | Phytolaccoside type triterpenic saponin | 8.04 | Arb | 2.21 |
| 12.70 | 644.3829 | C36H56O11 | Phytolaccoside type triterpenic saponin | 5.61 | Arb | 2.22 |
| 12.82 | 648.3879 | C36H56O10 | Triterpenic saponin | 6.06 | Arb | 1.14 |
| 12.83 | 1220.5478 | C57H88O28 | Triterpenic saponin | 7.89 | Arb | 2.18 |
| 12.83 | 1088.5044 | C52H80O24 | Triterpenic saponin | 8.4 | Arb | 2.63 |
| 12.83 | 956.4618 | C47H72O20 | Triterpenic saponin | 7.57 | Arb | 1.01 |
| 12.97 | 678.3620 | C36H54O12 | Saponin (monoglycosilated) | 6.23 | Arb | 2.15 |
| 12.97 | 956.4619 | C47H72O20 | Triterpenic saponin | 6.86 | Arb | 1.45 |
| 13.40 | 620.3930 | C35H56O9 | Steroidal Saponin | 7.95 | Arb | 1.51 |
| 13.41 | 782.4457 | C41H66O14 | Steroidal Saponin | 7.11 | Arb | 1.31 |
| 13.78 | 706.3936 | C38H58O12 | Saponin | 6.97 | Arb | 1.14 |
| 14.32 | 1176.5935 | C57H92O25 | Saponin | 4.03 | Arb | 1.66 |
| 15.33 | 1072.5455 | C53H84O22 | Saponin | 4.55 | Arb | 2.67 |
| 15.84 | 926.4881 | C47H74O18 | Triterpenic saponin | 5.38 | Arb | 1.65 |
| 10.72 | 504.1271 | C24H24O12 | Flavonoid | −6.05 | Alb | 1.15 |
| 11.18 | 516.0907 | C24H20O13 | Dibenzofuran | −4.7 | Sat | 2.10 |
| 11.54 | 1118.5513 | C54H86O24 | Saponin | −7.84 | Sat | 1.12 |
| 14.71 | 942.5189 | C48H78O18 | Soyasaponin I | −7.38 | Sat, Alb | 3.54 |
| 15.28 | 502.3295 | C30H46O6 | Medicagenic acid (aglycon) | −7.59 | Alb | 3.29 |
| 15.49 | 438.3132 | C29H42O3 | Phenol | −7.84 | Sat, Alb | 1.64 |
| 15.49 | 750.3823 | C39H58O14 | Saponin | −7.59 | Sat, Alb | 7.19 |
| 15.49 | 456.3240 | C29H44O4 | Triterpenic acid | −8.15 | Sat, Alb | 1.66 |
| 15.49 | 686.3662 | C38H54O11 | Triterpenic acid derivative | −7.67 | Sat, Alb | 1.16 |
| 15.79 | 386.1727 | C22H26O6 | Lignan | −11.99 | Alb (vs. Sat) | 13.72 |
| 16.27 | 1026.50357 | C51H78O21 | Steroidal saponin | −7.81 | Sat, Alb | 1.37 |
| a. For Aerial Parts (Shoots) | ||||||
|---|---|---|---|---|---|---|
| RT [min] | Formula | Molecular Weight | Category | Log2 Fold Change | Upregulated | VIP Score |
| 7.78 | C15H16O10 | 356.0741 | Caffeic acid derivative | 8.25 | Arb | 1.21 |
| 8.28 | C10H10O4 | 194.0579 | Ferulic acid | 7.84 | Arb | 1.65 |
| 8.31 | C16H18O10 | 370.0898 | Cinnamic acid derivative | 7.95 | Arb | 1.11 |
| 9.40 | C33H34O23 | 798.1483 | Flavonoid (e.g., apigenin glycoside) | 10.15 | Arb | 6.63 |
| 9.55 | C34H36O24 | 828.1591 | Flavonoid (e.g., diosmetin glycoside) | 8.23 | Arb | 1.67 |
| 10.46 | C40H40O23 | 888.1954 | Flavonoid (flavonol glycoside) | 10.21 | Arb | 1.73 |
| 10.68 | C39H38O22 | 858.1851 | Flavonoid (flavonol glycoside) | 8.38 | Arb | 1.92 |
| 12.90 | C57H90O27 | 1206.5664 | Medicagenic acid saponin | 7.52 | Arb | 1.74 |
| 8.40 | C33H34O23 | 798.1492 | Flavonoid (e.g., apigenin glycoside) | −7.03 | Alb, Sat | 5.15 |
| 8.51 | C18H24O12 | 432.1268 | Iridoid | −7.36 | Alb, Sat | 7.11 |
| 9.04 | C27H26O17 | 622.1173 | Flavonoid (e.g., apigenin glycoside) | −8.65 | Alb, Sat | 4.92 |
| 9.59 | C43H42O26 | 974.1961 | Flavonoid (e.g., apigenin glycoside) | −7.25 | Alb, Sat | 7.31 |
| 10.83 | C21H18O11 | 455.0850 | Flavonoid (e.g., flavone glycoside) | −5.03 | Sat | 1.14 |
| 11.07 | C37H34O20 | 798.1641 | Flavonoid (e.g., apigenin glycoside) | −5.53 | Sat, Alb | 5.73 |
| 11.28 | C17H14O6 | 314.0790 | Flavonoid | −7.69 | Alb (vs. Sat) | 2.21 |
| 11.63 | C53H86O23 | 1090.5560 | Triterpenic saponin | −6.85 | Alb, Sat | 2.67 |
| 11.91 | C64H102O34 | 1414.6255 | Saponin | −7.43 | Sat | 2.40 |
| 12.13 | C48H76O21 | 988.4877 | Triterpenic saponin | −6.89 | Alb, Sat | 1.24 |
| 12.48 | C69H110O37 | 1530.6740 | Saponin | −6.77 | Sat | 1.01 |
| 15.796 | C22H26O6 | 386.1727 | Lignan | −11.57 | Alb (vs. Sat) | 16.01 |
| b. Roots | ||||||
| RT [min] | Molecular Weight | Formula | Category | Log2 Fold Change | Upregulated | VIP Score |
| 12.03 | 1236.5425 | C57H88O29 | Zahnic acid saponin | 8.03 | Arb | 1.55 |
| 12.08 | 1104.4988 | C52H80O25 | Zahnic acid saponin | 7.33 | Arb | 1.03 |
| 12.15 | 1222.5630 | C57H90O28 | Zahnic acid saponin | 7.21 | Arb | 1.14 |
| 12.83 | 1220.5476 | C57H88O28 | Medicagenic acid saponin | 6.81 | Arb | 2.13 |
| 12.83 | 1088.5040 | C52H80O24 | Medicagenic acid saponin | 8.16 | Arb | 3.04 |
| 12.83 | 956.4616 | C47H72O20 | Medicagenic acid saponin | 5.77 | Arb | 2.46 |
| 12.97 | 648.3876 | C36H56O10 | Medicagenic acid saponin | 6.11 | Arb | 1.58 |
| 12.97 | 1106.5337 | C49H86O27 | Medicagenic acid saponin | 5.34 | Arb | 1.087 |
| 12.97 | 648.3876 | C36H56O10 | Medicagenic acid saponin | 6.48 | Arb | 1.36 |
| 12.97 | 956.4616 | C47H72O20 | Medicagenic acid saponin | 5.77 | Arb | 2.15 |
| 12.97 | 632.3563 | C35H52O10 | Medicagenic acid saponin | 6.73 | Arb | 1.04 |
| 12.97 | 678.3618 | C36H54O12 | Medicagenic acid saponin | 5.82 | Arb | 2.64 |
| 12.97 | 824.4196 | C42H64O16 | Medicagenic acid saponin | 6.74 | Arb | 2.22 |
| 13.41 | 470.3398 | C30H46O4 | Triterpenic acid | 7.57 | Arb | 2.27 |
| 13.48 | C48H70O22 | Steroidic saponin | −8.27 | Sat | −2.87 | |
| 15.50 | 750.3825 | C39H58O14 | Steroidic saponin | −6.48 | Alb | 5.46 |
| 15.50 | 502.3296 | C30H46O6 | Triterpenic acid (e.g., medicagenic) | −7.56 | Sat, Alb | −3.06 |
| 15.82 | 284.1258 | C14H20O6 | Lignan | −11.08 | Alb | 1.97 |
| 15.82 | 386.1727 | C22H26O6 | Lignan | −11.99 | Alb | 11.93 |
| 15.82 | 266.1152 | C14H18O6 | Simple phenolic acid | −10.19 | Alb | 1.85 |
| 16.50 | 486.3350 | C30H46O5 | Triterpenic acid (e.g., quiallic) | −5.17 | Sat | 1.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarri, E.; Termentzi, A.; Abraham, E.M.; Papadopoulos, G.K.; Baira, E.; Machera, K.; Loukas, V.; Komaitis, F.; Tani, E. Salinity Stress Alters the Secondary Metabolic Profile of M. sativa, M. arborea and Their Hybrid (Alborea). Int. J. Mol. Sci. 2021, 22, 4882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094882
Sarri E, Termentzi A, Abraham EM, Papadopoulos GK, Baira E, Machera K, Loukas V, Komaitis F, Tani E. Salinity Stress Alters the Secondary Metabolic Profile of M. sativa, M. arborea and Their Hybrid (Alborea). International Journal of Molecular Sciences. 2021; 22(9):4882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094882
Chicago/Turabian StyleSarri, Efi, Aikaterini Termentzi, Eleni M. Abraham, George K. Papadopoulos, Eirini Baira, Kyriaki Machera, Vassilis Loukas, Fotios Komaitis, and Eleni Tani. 2021. "Salinity Stress Alters the Secondary Metabolic Profile of M. sativa, M. arborea and Their Hybrid (Alborea)" International Journal of Molecular Sciences 22, no. 9: 4882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094882