
Assessment of TSPAN Expression Profile and Their Role in the VSCC Prognosis

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Case Selection and Clinical Features of the Study Population
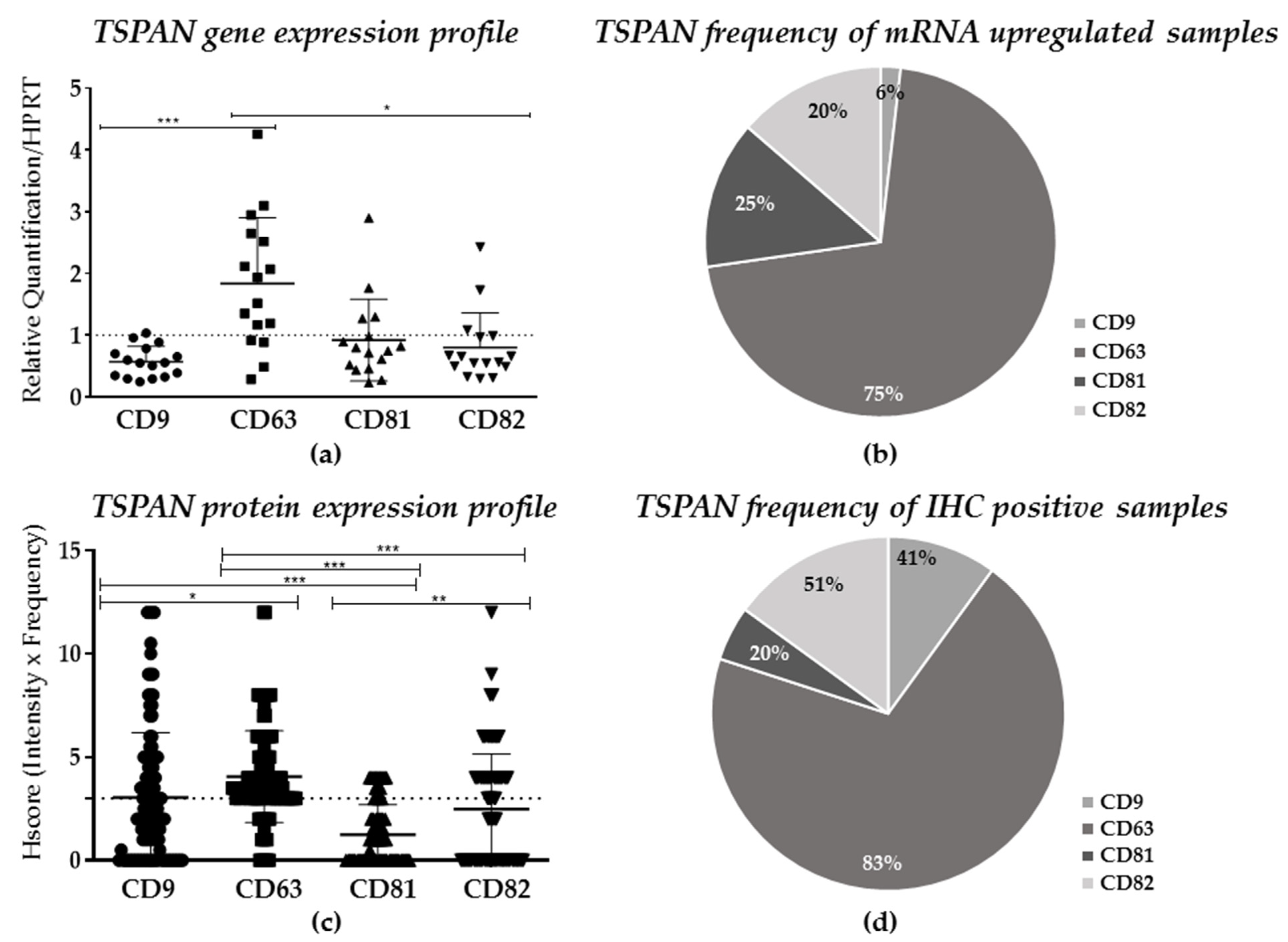
2.2. TSPANs Expression in Patients’ Samples
2.3. Clinical Implication of TSPANs Expression for VSCC Patients
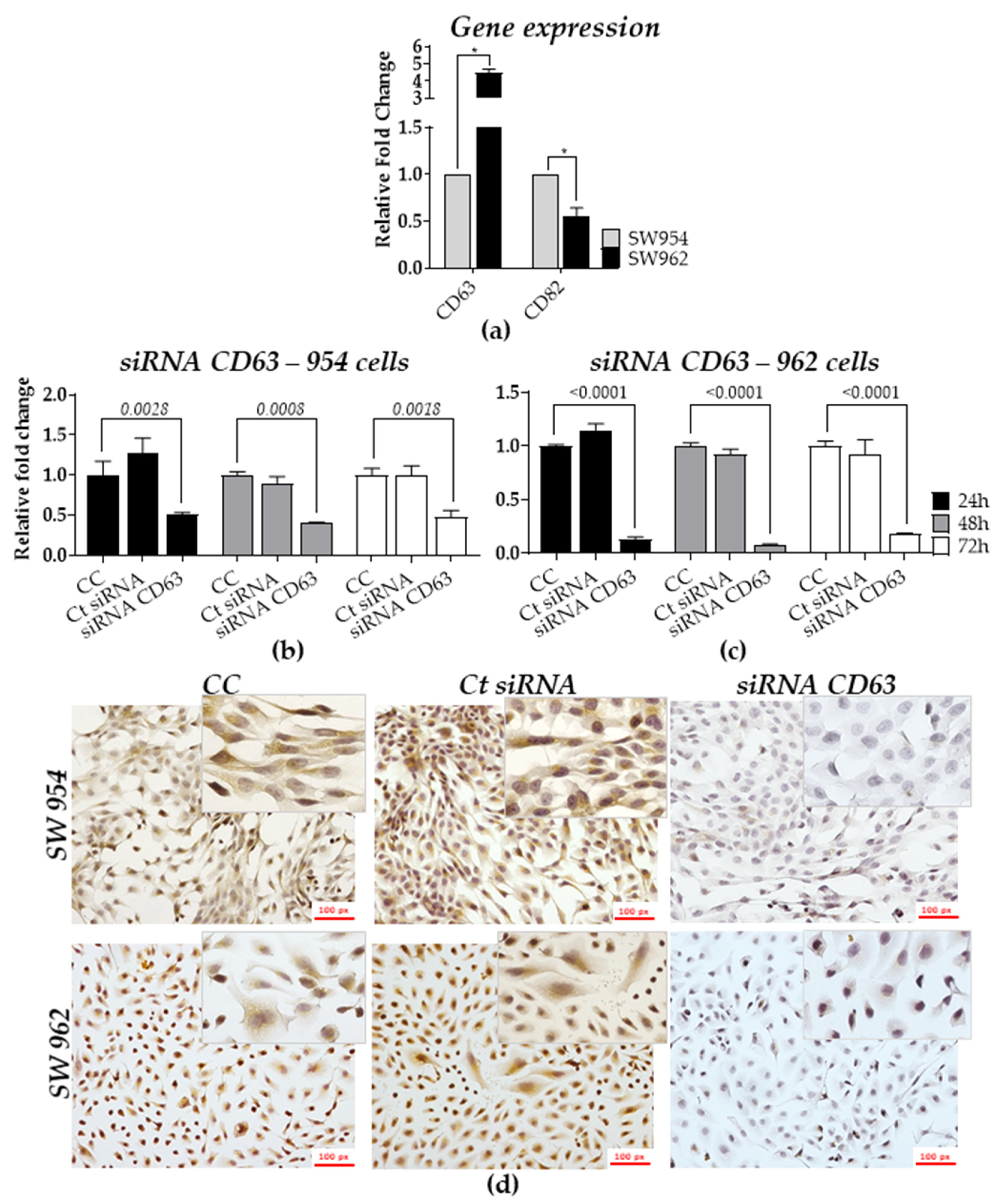
2.4. Role of TSPANs in Metastatic and Non-Metastatic Cell Lines
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. Cell Lines, Culture Conditions, and Authentication
4.3. Total RNA Isolation, Complementary DNA (cDNA) Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
4.4. TMA and Immunohistochemistry (IHC) Analysis
4.5. DNA Isolation and HPV Genotyping
4.6. CD63 Silencing-siRNA Transfection
4.7. Immunocytochemistry (ICC)
4.8. Cell Viability Assay
4.9. Wound Healing Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giulia, M.; Fragomeni, S.M.; Inzani, F.; Fagotti, A.; Della Corte, L.; Gentileschi, S.; Tagliaferri, L.; Zannoni, G.F.; Scambia, G.; Garganese, G. Molecular pathways in vulvar squamous cell carcinoma: Implications for target therapeutic strategies. J. Cancer Res. Clin. Oncol. 2020, 146, 1647–1658. [Google Scholar]
- American Cancer Society Home-Cancer Statistics Center. Available online: https://cancerstatisticscenter.cancer.org/#!/ (accessed on 13 July 2020).
- Bizzarri, N.; Vellone, V.G.; Parodi, L.; Fraccono, L.C.; Ghirardi, V.; Costantini, S.; Menada, M.V.; Sala, P. Cutaneous metastasis from vulvar squamous cell carcinoma: A rare occurrence that should not be forgotten. J. Obstet. Gynaecol. 2017, 37, 975–981. [Google Scholar] [CrossRef]
- Rogers, L.J.; Cuello, M.A. Cancer of the vulva. Int. J. Gynecol. Obstet. 2018, 143, 4–13. [Google Scholar] [CrossRef]
- Proctor, L.; Hoang, L.; Moore, J.; Thompson, E.; Leung, S.; Natesan, D.; Chino, J.; Gilks, B.; McAlpine, J.N. Association of human papilloma virus status and response to radiotherapy in vulvar squamous cell carcinoma. Int. J. Gynecol. Cancer 2020, 30, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, C.L.; Sand, F.L.; Hoffmann Frederiksen, M.; Kaae Andersen, K.; Kjaer, S.K. Does HPV status influence survival after vulvar cancer? Int. J. Cancer 2018, 142, 1158–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Grootenhuis, N.C.; Pouwer, A.F.W.; de Bock, G.H.; Hollema, H.; Bulten, J.; van der Zee, A.G.J.; de Hullu, J.A.; Oonk, M.H.M. Prognostic factors for local recurrence of squamous cell carcinoma of the vulva: A systematic review. Gynecol. Oncol. 2018, 148, 622–631. [Google Scholar] [CrossRef]
- Rakislova, N.; Alemany, L.; Clavero, O.; Del Pino, M.; Saco, A.; Marimon, L.; Quirós, B.; Lloveras, B.; Ribera-Cortada, I.; Alejo, M.; et al. HPV-independent precursors mimicking high-grade squamous intraepithelial lesions (HSIL) of the vulva. Am. J. Surg. Pathol. 2020, 44, 1506–1514. [Google Scholar] [CrossRef]
- Baiocchi, G.; Silva Cestari, F.M.; Rocha, R.M.; Lavorato-Rocha, A.; Maia, B.M.; Cestari, L.A.; Kumagai, L.Y.; Faloppa, C.C.; Fukazawa, E.M.; Badiglian-Filho, L.; et al. Prognostic value of the number and laterality of metastatic inguinal lymph nodes in vulvar cancer: Revisiting the FIGO staging system. Eur. J. Surg. Oncol. 2013, 39, 780–785. [Google Scholar] [CrossRef]
- Baiocchi, G.; Mantoan, H.; De Brot, L.; Badiglian-Filho, L.; Kumagai, L.Y.; Faloppa, C.C.; Da Costa, A.A.B.A. How important is the pathological margin distance in vulvar cancer? Eur. J. Surg. Oncol. 2015, 41, 1653–1658. [Google Scholar] [CrossRef]
- Woelber, L.; Jaeger, A.; Prieske, K. New treatment standards for vulvar cancer 2020. Curr. Opin. Obstet. Gynecol. 2020, 32, 9–14. [Google Scholar] [CrossRef]
- Kramer, F.; Hertel, H.; Hillemanns, P. Use of the sentinel lymph node technique compared to complete inguino-femoral lymph node removal in patients with invasive vulvar cancer in Germany. Geburtshilfe Frauenheilkd. 2013, 73, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Palisoul, M.L.; Mullen, M.M.; Feldman, R.; Thaker, P.H. Identification of molecular targets in vulvar cancers. Gynecol. Oncol. 2017, 146, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Maia, B.M.; Rodrigues, I.S.; Akagi, E.M.; do Amaral, N.S.; Ling, H.; Monroig, P.; Soares, F.A.; Calin, G.A.; Rocha, R.M. MiR-223-5p works as an oncomiR in vulvar carcinoma by TP63 suppression. Oncotarget 2016, 7, 49217–49231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavorato-Rocha, A.M.; Rodrigues, I.S.A.; Maia, B.M.; Stiepcich, M.M.Á.; Baiocchi, G.; Carvalho, K.C.; Soares, F.A.; Vassallo, J.; Rocha, R.M. Cell cycle suppressor proteins are not related to HPV status or clinical outcome in patients with vulvar carcinoma. Tumor. Biol. 2013, 34, 3713–3720. [Google Scholar] [CrossRef] [PubMed]
- Tessier-Cloutier, B.; Pors, J.; Thompson, E.; Ho, J.; Prentice, L.; McConechy, M.; Aguirre-Hernandez, R.; Miller, R.; Leung, S.; Proctor, L.; et al. Molecular characterization of invasive and in situ squamous neoplasia of the vulva and implications for morphologic diagnosis and outcome. Mod. Pathol. 2020, 34, 508–518. [Google Scholar] [CrossRef]
- Jones, E.L.; Wee, J.L.; Demaria, M.C.; Blakeley, J.; Ho, P.K.; Vega-Ramos, J.; Villadangos, J.A.; van Spriel, A.B.; Hickey, M.J.; Hämmerling, G.J.; et al. Dendritic cell migration and antigen presentation are coordinated by the opposing functions of the tetraspanins CD82 and CD37. J. Immunol. 2016, 196, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Hemler, M.E. Tetraspanin functions and associated microdomains. Nat. Rev. Molecul. Cell Biol. 2005, 6, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Termini, C.M.; Gillette, J.M. Tetraspanins function as regulators of cellular signaling. Front. Cell Dev. Biol. 2017, 5, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, T.; Hochheimer, N. Tetraspanins. Curr. Biol. 2020, 30, 204–206. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, X.; Geradts, J.; Wei, Q.; Hochwald, S.; Xu, H.; Huang, H. Expression of tetraspanins NET-6 and CD151 in breast cancer as a potential tumor biomarker. Clin. Exp. Med. 2019, 19, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.M.H.; Amoussou, N.G.; Mai, H.L.; Logé, C.; Brouard, S. Tetraspanins: Useful multifunction proteins for the possible design and development of small-molecule therapeutic tools. Drug Discov. Today 2020, 26, 56–68. [Google Scholar] [CrossRef]
- Van Spriel, A.B.; Figdor, C.G. The role of tetraspanins in the pathogenesis of infectious diseases. Microbes Infect. 2010, 12, 106–112. [Google Scholar] [CrossRef]
- Protzel, C.; Kakies, C.; Kleist, B.; Poetsch, M.; Giebel, J. Down-regulation of the metastasis suppressor protein KAI1/CD82 correlates with occurrence of metastasis, prognosis and presence of HPV DNA in human penile squamous cell carcinoma. Virchows Arch. 2008, 452, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Spoden, G.; Freitag, K.; Husmann, M.; Boller, K.; Sapp, M.; Lambert, C.; Florin, L. Clathrin- and caveolin-independent entry of human papillomavirus type 16-Involvement of tetraspanin-enriched microdomains (TEMs). PLoS ONE 2008, 3, e3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thali, M. The roles of tetraspanins in HIV-1 replication. Curr. Top. Microbiol. Immunol. 2009, 339, 85–102. [Google Scholar] [PubMed] [Green Version]
- Thali, M. Tetraspanin functions during HIV-1 and influenza virus replication. Biochem. Soc. Trans. 2011, 39, 529–531. [Google Scholar] [CrossRef]
- Fast, L.A.; Mikuličić, S.; Fritzen, A.; Schwickert, J.; Boukhallouk, F.; Hochdorfer, D.; Sinzger, C.; Suarez, H.; Monk, P.N.; Yáñez-Mó, M.; et al. Inhibition of tetraspanin functions impairs human papillomavirus and cytomegalovirus infections. Int. J. Mol. Sci. 2018, 19, 3007. [Google Scholar] [CrossRef] [Green Version]
- Earnest, J.T.; Hantak, M.P.; Li, K.; McCray, P.B.; Perlman, S.; Gallagher, T. The tetraspanin CD9 facilitates MERS-coronavirus entry by scaffolding host cell receptors and proteases. PLoS Pathog. 2017, 13, e1006546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hantak, M.P.; Qing, E.; Earnest, J.T.; Gallagher, T. Tetraspanins: Architects of viral entry and exit platforms. J. Virol. 2019, 93, e01429-17. [Google Scholar] [CrossRef] [Green Version]
- Kaprio, T.; Hagström, J.; Andersson, L.C.; Haglund, C. Tetraspanin CD63 independently predicts poor prognosis in colorectal cancer. Histol. Histopathol. 2020, 35, 887–892. [Google Scholar]
- Ridnour, L.A.; Barasch, K.M.; Windhausen, A.N.; Dorsey, T.H.; Lizardo, M.M.; Yfantis, H.G.; Lee, D.H.; Switzer, C.H.; Cheng, R.Y.S.; Heinecke, J.L.; et al. Nitric Oxide Synthase and breast cancer: Role of TIMP-1 in NO-mediated Akt activation. PLoS ONE 2012, 7, e44081. [Google Scholar]
- Park, C.S.; Kim, T.K.; Kim, H.G.; Kim, Y.J.; Jeoung, M.H.; Lee, W.R.; Go, N.K.; Heo, K.; Lee, S. Therapeutic targeting of tetraspanin8 in epithelial ovarian cancer invasion and metastasis. Oncogene 2016, 35, 4540–4548. [Google Scholar] [CrossRef] [PubMed]
- Hiroshima, K.; Shiiba, M.; Oka, N.; Hayashi, F.; Ishida, S.; Fukushima, R.; Koike, K.; Iyoda, M.; Nakashima, D.; Tanzawa, H.; et al. Tspan15 plays a crucial role in metastasis in oral squamous cell carcinoma. Exp. Cell Res. 2019, 384, 111622. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Ju, J.; Zhang, S.W.; Shen, Z.Y.; Liang, L.; Yang, X.M.; Ma, C.; Ni, Q.W.; Sun, M.Y. p12CDK2-AP1 interacts with CD82 to regulate the proliferation and survival of human oral squamous cell carcinoma cells. Oncol. Rep. 2016, 36, 737–744. [Google Scholar] [CrossRef] [Green Version]
- Huan, J.; Gao, Y.; Xu, J.; Sheng, W.; Zhu, W.; Zhang, S.; Cao, J.; Ji, J.; Zhang, L.; Tian, Y. Overexpression of CD9 correlates with tumor stage and lymph node metastasis in esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 3054–3061. [Google Scholar] [PubMed]
- Zeng, T.D.; Zheng, B.; Zheng, W.; Chen, C. CD82/KAI1 inhibits invasion and metastasis of esophageal squamous cell carcinoma via TGF-β1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5928–5937. [Google Scholar]
- Yang, Y.G.; Sari, I.N.; Zia, M.F.; Lee, S.R.; Song, S.J.; Kwon, H.Y. Tetraspanins: Spanning from solid tumors to hematologic malignancies. Exp. Hematol. 2016, 44, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Heo, K.; Lee, S. TSPAN8 as a novel emerging therapeutic target in cancer for monoclonal antibody therapy. Biomolecules 2020, 10, 388. [Google Scholar] [CrossRef] [Green Version]
- Pols, M.S.; Klumperman, J. Trafficking and function of the tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef]
- Kudo, K.; Yoneda, A.; Sakiyama, D.; Kojima, K.; Miyaji, T.; Yamazaki, M.; Yaita, S.; Hyodo, T.; Satow, R.; Fukami, K. Cell surface CD63 increased by up-regulated polylactosamine modification sensitizes human melanoma cells to the BRAF inhibitor PLX4032. FASEB J. 2019, 33, 3851–3869. [Google Scholar] [CrossRef]
- Garcia-Mayea, Y.; Mir, C.; Carballo, L.; Castellvi, J.; Temprana-Salvador, J.; Lorente, J.; Benavente, S.; García-Pedrero, J.M.; Allonca, E.; Rodrigo, J.P.; et al. TSPAN1: A Novel Protein Involved in Head and Neck Squamous Cell Carcinoma Chemoresistance. Cancers (Basel) 2020, 12, 3269. [Google Scholar] [CrossRef]
- An, H.J.; Kim, M.H.; Kim, S.H.; Lee, G.W.; Song, D.H. Prognostic role of extracellular vesicles in squamous cell carcinoma of the lung. Thorac. Cancer 2020, 11, 1989–1995. [Google Scholar] [CrossRef]
- Lavorato-Rocha, A.M.; Maia, B.M.; Rodrigues, I.S.A.; Stiepcich, M.M.Á.; Baiocchi, G.; Cestari, F.M.D.S.; Carvalho, K.C.; Soares, F.A.; Rocha, R.M. Prognostication of vulvar cancer based on p14ARF status: Molecular assessment of transcript and protein. Ann. Surg. Oncol. 2012, 20, 31–39. [Google Scholar] [CrossRef]
- Akagi, E.M.; Lavorato-Rocha, A.M.; Maia, B.M.; Rodrigues, I.S.; Carvalho, K.C.; Stiepcich, M.M.; Baiocchi, G.; Sato-Kuwabara, Y.; Rogatto, S.R.; Soares, F.A.; et al. ROCK1 as a novel prognostic marker in vulvar cancer. BMC Cancer 2014, 14, 822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, B.M.; Fontes, A.M.; Lavorato-Rocha, A.M.; Rodrigues, I.S.A.; De Brot, L.; Baiocchi, G.; Stiepcich, M.M.; Soares, F.A.; Rocha, R.M. EGFR expression in vulvar cancer: Clinical implications and tumor heterogeneity. Hum. Pathol. 2014, 45, 917–925. [Google Scholar] [CrossRef]
- Lavorato-Rocha, A.M.; Anjos, L.G.; Cunha, I.W.; Vassallo, J.; Soares, F.A.; Rocha, R.M. Immunohistochemical assessment of PTEN in vulvar cancer: Best practices for tissue staining, evaluation, and clinical association. Methods 2015, 77, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Lavorato-Rocha, A.M.; Akagi, E.M.; Maia, B.M.; Rodrigues, I.S.; Botelho, M.C.S.; Marchi, F.A.; Fernandes, G.; Baiocchi, G.; Soares, F.A.; Rogatto, S.R.; et al. An integrative approach uncovers biomarkers that associate with clinically relevant disease outcomes in vulvar carcinoma. Mol. Cancer Res. 2016, 14, 720–729. [Google Scholar] [CrossRef] [Green Version]
- Maia, B.M.; Lavorato-Rocha, A.M.; Rodrigues, I.S.; Baiocchi, G.; Cestari, F.M.; Stiepcich, M.M.; Chinen, L.T.D.; Carvalho, K.C.; Soares, F.A.; Rocha, R.M. Prognostic significance of c-KIT in vulvar cancer: Bringing this molecular marker from bench to bedside. J. Transl. Med. 2012, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Maia, B.M.; Cestari, F.M.; Lavorato-Rocha, A.M.; Rodrigues, I.S.A.; Baiocchi, G.; Guimarães, G.C.; Vassallo, J.; Stiepcich, M.; Soares, F.A.; Rocha, R.M. Characterization of sociodemographic and clinicopathological features in brazilian patients with vulvar squamous cell carcinoma. Gynecol. Obst. Investig. 2013, 75, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Baiocchi, G.; Silva Cestari, F.M.; Rocha, R.M.; Faloppa, C.C.; Kumagai, L.Y.; Fukazawa, E.M.; Badiglian-Filho, L.; Cestari, L.A.; Sant’Ana Rodrigues, I.; Lavorato-Rocha, A.; et al. Does the count after inguinofemoral lymphadenectomy in vulvar cancer correlate with outcome? Eur. J. Surg. Oncol. 2013, 39, 339–343. [Google Scholar] [CrossRef]
- Chen, J.; LN, H. A review of prognostic factors in squamous cell carcinoma of the vulva: Evidence from the last decade. Semin. Diagn. Pathol. 2020, 38, 37–49. [Google Scholar]
- Huang, C.L.; Taki, T.; Adachi, M.; Yagita, M.; Sawada, S.; Takabayashi, A.; Inufusa, H.; Yoshie, O.; Miyake, M. MRP-1/CD9 and KAI1/CD82 expression in normal and various cancer tissues. Int. J. Oncol. 1997, 11, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Jang, N.; Choi, J.E.; Kim, J.R.; Bae, Y.K. CD9 expression in tumor cells is associated with poor prognosis in patients with invasive lobular carcinoma. J. Breast Cancer 2019, 22, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Nakano, K.; Itoi, S.; Koh, T.; Taki, T. Motility-related Protein-1 (MRP-1/CD9) reduction as a factor of poor prognosis in breast cancer. Cancer Res. 1996, 56, 1244–1249. [Google Scholar]
- Huang, C.I.; Kohno, N.; Ogawa, E.; Adachi, M.; Taki, T.; Miyake, M. Correlation of reduction in MRP-1/CD9 and KAI1/CD82 expression with recurrences in breast cancer patients. Am. J. Pathol. 1998, 153, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Uchida, S.; Shimada, Y.; Watanabe, G.; Li, Z.G.; Hong, T.; Miyake, M.; Imamura, M. Motility-related protein (MRP-1/CD9) and KAI1/CD82 expression inversely correlate with lymph node metastasis in oesophageal squamous cell carcinoma. Br. J. Cancer 1999, 79, 1168–1173. [Google Scholar] [CrossRef] [Green Version]
- Kusukawa, J.; Ryu, F.; Kameyama, T.; Mekada, E. Reduced expression of CD9 in oral squamous cell carcinoma: CD9 expression inversely related to high prevalence of lymph node metastasis. J. Oral. Pathol. Med. 2001, 30, 73–79. [Google Scholar] [CrossRef]
- Buim, M.E.C.; Lourenço, S.V.; Carvalho, K.C.; Cardim, R.; Pereira, C.; Carvalho, A.L.; Fregnani, J.H.; Soares, F.A. Downregulation of CD9 protein expression is associated with aggressive behavior of oral squamous cell carcinoma. Oral. Oncol. 2010, 46, 166–171. [Google Scholar] [CrossRef]
- Hori, H.; Yano, S.; Koufuji, K.; Takeda, J.; Shirouzu, K. CD9 expression in gastric cancer and its significance. J. Surg. Res. 2004, 117, 208–215. [Google Scholar] [CrossRef]
- Soyuer, S.; Soyuer, I.; Unal, D.; Ucar, K.; Yildiz, O.G.; Orhan, O. Prognostic significance of CD9 expression in locally advanced gastric cancer treated with surgery and adjuvant chemoradiotherapy. Pathol. Res. Pract. 2010, 206, 607–610. [Google Scholar] [CrossRef]
- Kwon, H.J.; Min, S.Y.; Park, M.J.; Lee, C.; Park, J.H.; Chae, J.Y.; Moon, K.C. Expression of CD9 and CD82 in clear cell renal cell carcinoma and its clinical significance. Pathol. Res. Pract. 2014, 210, 285–290. [Google Scholar] [CrossRef]
- Sho, M.; Adachi, M.; Taki, T.; Hashida, H.; Konishi, T.; Huang, C.; Ikeda, N.; Nakajima, Y.; Kanehiro, H.; Hisanaga, M.; et al. Transmembrane 4 superfamily as a prognostic factor in pancreatic cancer. Int. J. Cancer 1998, 79, 509–516. [Google Scholar] [CrossRef]
- Saito, Y.; Tachibana, I.; Takeda, Y.; Yamane, H.; He, P.; Suzuki, M.; Minami, S.; Kijima, T.; Yoshida, M.; Kumagai, T.; et al. Absence of CD9 enhances adhesion-dependent morphologic differentiation, survival, and matrix metalloproteinase-2 production in small cell lung cancer cells. Cancer Res. 2006, 66, 9557–9565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeyama, S.; Koyama, M.; Yamaoko, M.; Sasada, R.; Miyake, M. Suppression of cell motility and metastasis by transfection with human motility-related protein (MRP-1/CD9) DNA. J. Exp. Med. 1993, 177, 1231–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, I.K.; Jin, Y.J.; Byun, H.J.; Jeoung, D.I.; Kim, Y.M.; Lee, H. Homophilic interactions of tetraspanin CD151 up-regulate motility and matrix metalloproteinase-9 expression of human melanoma cells through adhesion-dependent c-Jun activation signaling pathways. J. Biol. Chem. 2006, 281, 24279–24292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidone, L.R.M. Imunoexpressão CD9 No Carcinoma Epidermóide Do Pênis. Master’s Thesis, (MsC) FAP-Hospital A C Camargo, São Paulo, SP, Brazil, 2010. [Google Scholar]
- Zhang, X.A.; Lane, W.S.; Charrin, S.; Rubinstein, E.; Liu, L. EWI2/PGRL Associates with the Metastasis SuppressorSS KAI1/CD82 and Inhibits the Migration of Prostate Cancer Cells. Cancer Res. 2003, 63, 2665–2674. [Google Scholar]
- Malik, F.A.; Sanders, A.J.; Jiang, W.G. KAI-1/CD82, the molecule and clinical implication in cancer and cancer metastasis. Histol. Histopathol. 2009, 24, 519–530. [Google Scholar]
- Zheng, Z.; Tian, R.; Wang, P. Roles of KAI1 and nm23 in lymphangiogenesis and lymph metastasis of laryngeal squamous cell carcinoma. World J. Surg. Oncol. 2017, 15, 211. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wei, L.L.; Tang, C.; Slack, R.; Mueller, S.; Lippman, M.E. Overexpression of KAI1 Suppresses in vitro invasiveness and in vivo metastasis in breast cancer cells. Cancer Res. 2001, 61, 5284–5288. [Google Scholar]
- Kussaibi, H.; Alkharsah, K.R. Investigations of a possible role of SNPs in KAI1 gene on its down-regulation in breast cancer. Asian Pac. J. Cancer Prev. 2020, 21, 2549–2553. [Google Scholar] [CrossRef] [PubMed]
- Jee, B.K.; Park, K.M.; Surendran, S.; Lee, W.K.; Han, C.W.; Kim, Y.S.; Lim, Y. KAI1/CD82 suppresses tumor invasion by MMP9 inactivation via TIMP1 up-regulation in the H1299 human lung carcinoma cell line. Biochem. Biophys. Res. Commun. 2006, 342, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Du, L.; Ju, J.; Ma, C.; Shen, Z.; Yang, X.; Liang, L.; Ni, Q.; Sun, M. Overexpression of KAI1/CD82 suppresses in vitro cell growth, migration, invasion and xenograft growth in oral cancer. Mol. Med. Rep. 2017, 15, 1527–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Gao, Y.; Liu, W.; Zheng, L.; Jin, B.; Duan, B.; Xie, H.; Guo, P.; Zeng, J.; Wang, K.; et al. CD82 Suppresses ADAM17-Dependent E-Cadherin cleavage and cell migration in prostate cancer. Dis. Markers 2020, 2020, 8899924. [Google Scholar] [CrossRef]
- Tavsan, Z.; Kayalı, H.A. EpCAM-claudin-tetraspanin-modulated ovarian cancer progression and drug resistance. Cell Adh. Migr. 2020, 14, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Mizoshiri, N.; Shirai, T.; Terauchi, R.; Tsuchida, S.; Mori, Y.; Hayashi, D.; Kishida, T.; Arai, Y.; Mazda, O.; Nakanishi, T.; et al. The tetraspanin CD81 mediates the growth and metastases of human osteosarcoma. Cell Oncol. 2019, 42, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Vences-Catalán, F.; Duault, C.; Kuo, C.C.; Rajapaksa, R.; Levy, R.; Levy, S. CD81 as a tumor target. Biochem. Soc. Trans. 2017, 45, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ou, X.; Wu, X. Proteomics profiling of plasma exosomes in epithelial ovarian cancer: A potential role in the coagulation cascade, diagnosis and prognosis. Int. J. Oncol. 2019, 54, 1719–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Qian, H.; Xu, A.; Yang, G. Increased expression of CD81 is associated with poor prognosis of prostate cancer and increases the progression of prostate cancer cells in vitro. Exp. Ther. Med. 2019, 19, 755–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carloni, V.; Mazzocca, A.; Mello, T.; Galli, A.; Capaccioli, S. Cell fusion promotes chemoresistance in metastatic colon carcinoma. Oncogene 2013, 32, 2649–2660. [Google Scholar] [CrossRef] [Green Version]
- Quagliano, A.; Gopalakrishnapillai, A.; Anders Kolb, E.; Barwe, S.P. CD81 knockout promotes chemosensitivity and disrupts in vivo homing and engraftment in acute lymphoblastic leukemia. Blood Adv. 2020, 4, 4393–4405. [Google Scholar] [CrossRef] [PubMed]
- American Type Culture Collection Search ATCC STR Database. Available online: https://www.atcc.org/STR_Database.aspx (accessed on 19 October 2018).
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, K.C.; Cunha, I.W.; Rocha, R.M.; Buim, M.E.; Maschietto, M.; Kuwabara, Y.; Reis, L.F.L.; Soares, F.A. Tetraspanin expression profile in soft tissue sarcomas. J. Dis. Markers 2015, 2, 1024. [Google Scholar]
- Rodrigues, I.S.; Lavorato-Rocha, A.M.; Maia, B.M.; Stiepcich, M.M.A.; Carvalho, F.M.; Baiocchi, G.; Soares, F.A.; Rocha, R.M. Epithelial-mesenchymal transition-like events in vulvar cancer and its relation with HPV. Br. J. Cancer 2013, 109, 184–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, B.C.; dos Anjos, L.G.; Uno, M.; Cunha, I.W.; Soares, F.A.; Baiocchi, G.; Baracat, E.C.; Carvalho, K.C. Let-7 miRNA’s expression profile and its potential prognostic role in uterine leiomyosarcoma. Cells 2019, 8, 1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Nie, J.; Chen, H.-S.; Ma, H.; Stewart, R.; Li, X.; Lu, M.-Z.; Taylor, W.M.; Wei, H. Evaluation of gene association methods for coexpression network construction and biological knowledge discovery. PLoS ONE 2012, 7, e50411. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Categories | Overall Population by Medical Record/Percentage Frequencies |
|---|---|---|
| Age | ≤50 | 12 (10.5%) |
| >50 | 102 (89.5%) | |
| Menarche | 11 to 14 years old | 41 (68.3%) |
| 15 to 19 years old | 19 (31.7%) | |
| Menopause | No | 3 (4.6%) |
| Yes | 62 (95.4%) | |
| Sexarche | 15 to 18 years old | 16 (51.6%) |
| ≥19 years old | 15 (48.4%) | |
| Number of partners | ≤2 | 36 (92.3%) |
| ≥3 | 3 (7.7%) | |
| Symptoms | No | 1 (1.2%) |
| Yes | 86 (98.8%) | |
| Contraceptive | No | 61 (92.4%) |
| Yes | 5 (7.6%) | |
| Alcoholism | No | 74 (98.6%) |
| Yes | 1 (1.4%) | |
| Smoking | No | 59 (77%) |
| Yes | 17 (23%) | |
| Tumor size (mm3) | ≤5 mm | 42 (37.5%) |
| 6 to 10 mm | 21 (18.8%) | |
| ≥10 mm | 49 (43.7%) | |
| Histological diagnosis | VSCC 1 | 44 (38%) |
| VSCC 2 | 47 (40.5%) | |
| VSCC 3 | 11 (9.5%) | |
| Basaloid carcinoma | 11 (9.5%) | |
| Verrucous carcinoma | 3 (2.5%) | |
| Neoadjuvant chemotherapy | No | 69 (87%) |
| Yes | 10 (13%) | |
| Adjuvant chemotherapy | No | 76 (91.5%) |
| Yes | 7 (8.5%) | |
| Adjuvant radiotherapy | No | 73 (89%) |
| Yes | 9 (11%) | |
| Pelvic lymphadenectomy | No | 69 (89.6%) |
| Yes | 8 (10.4%) | |
| Depth of invasion | Superficial dermis | 11 (9.8%) |
| Dermis | 70 (62.5%) | |
| Lower layer of the dermis | 20 (17.9%) | |
| Subcutaneous | 9 (8%) | |
| Rectal mucosa | 1 (0.9%) | |
| Dermis and subcutaneous | 1 (0.9%) | |
| Inflammatory infiltrate | 1+ Low | 37 (33%) |
| 2+ Moderate | 67 (60%) | |
| 3+ High | 8 (7%) | |
| Vascular invasion | No | 98 (83.8%) |
| Yes | 19 (16.2%) | |
| Perineural invasion | No | 102 (91%) |
| Yes | 10 (9%) | |
| FIGO 1 stage | I (A/B) and II | 43 (57.4%) |
| III A | 5 (6.7%) | |
| III B | 19 (25.3%) | |
| III C | 7 (9.3%) | |
| IV (A/B) | 1 (1.3%) | |
| Relapse | No | 41 (50.6%) |
| Yes | 40 (49.4%) | |
| Lymph node metastasis | No | 24 (60%) |
| Yes | 16 (40%) | |
| Status | Alive | 33 (38%) |
| Death | 54 (62%) | |
| HPV infection | No | 20 (39.2%) |
| Yes | 31 (60.8%) | |
| HPV co-infection | No | 20 (64.5%) |
| Yes | 11 (35.5%) | |
| HPV type | High-risk | 25 (80.6%) |
| Low-risk | 3 (9.7%) | |
| Both | 3 (9.7%) |
| Target/ Staining | CD9 n (%) | CD63 n (%) | CD81 n (%) | CD82 n (%) | Total n (%) | p |
|---|---|---|---|---|---|---|
| Negative | 66 (59) | 18 (17) | 84 (80) | 56 (52) | 223 (52) | <0.0001 |
| Positive | 45 (41) | 89 (83) | 21 (20) | 51 (48) | 206 (48) | |
| Total * | 111 (100) | 107 (100) | 105 (100) | 107 (100) | 429 (100) |
| TSPAN | Correlation Coefficient (r) | p | |
|---|---|---|---|
| CD9 | CD63 | 0.325 | 0.001 |
| CD81 | 0.423 | <0.0001 | |
| CD82 | 0.597 | <0.0001 | |
| CD63 | CD81 | 0.202 | 0.039 |
| CD82 | 0.422 | <0.0001 | |
| CD81 | CD82 | 0.356 | <0.0001 |
| Variable | HR (95% CI) | p |
|---|---|---|
| CD81 expression 1 | 0.41 (0.11–1.49) | 0.178 |
| Neoadjuvant chemotherapy 2 | 0.69 (0.19–2.46) | 0.576 |
| Treatment Group | Non-Metastatic Positive n (%) | n (%) | Metastatic Positive n (%) | n (%) | * p |
|---|---|---|---|---|---|
| Cell Control | 2002 (98) | 2043 (100) | 561 (98) | 571 (100) | 0.0005 |
| siRNA Control | 2318 (98.5) | 2354 (100) | 603 (98.7) | 611 (100) | |
| CD63 siRNA | 410 (21) | 1966 (100) | 67 (14) | 472 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, K.P.; de Almeida, B.C.; dos Anjos, L.G.; Baiocchi, G.; Soares, F.A.; Rocha, R.M.; Baracat, E.C.; Dobroff, A.S.; Carvalho, K.C. Assessment of TSPAN Expression Profile and Their Role in the VSCC Prognosis. Int. J. Mol. Sci. 2021, 22, 5015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095015
Ferreira KP, de Almeida BC, dos Anjos LG, Baiocchi G, Soares FA, Rocha RM, Baracat EC, Dobroff AS, Carvalho KC. Assessment of TSPAN Expression Profile and Their Role in the VSCC Prognosis. International Journal of Molecular Sciences. 2021; 22(9):5015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095015
Chicago/Turabian StyleFerreira, Kelly Pedrozo, Bruna Cristine de Almeida, Laura Gonzalez dos Anjos, Glauco Baiocchi, Fernando Augusto Soares, Rafael Malagoli Rocha, Edmund Chada Baracat, Andrey Senos Dobroff, and Katia Candido Carvalho. 2021. "Assessment of TSPAN Expression Profile and Their Role in the VSCC Prognosis" International Journal of Molecular Sciences 22, no. 9: 5015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095015