
Application of microRNA in Human Osteoporosis and Fragility Fracture: A Systemic Review of Literatures

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials
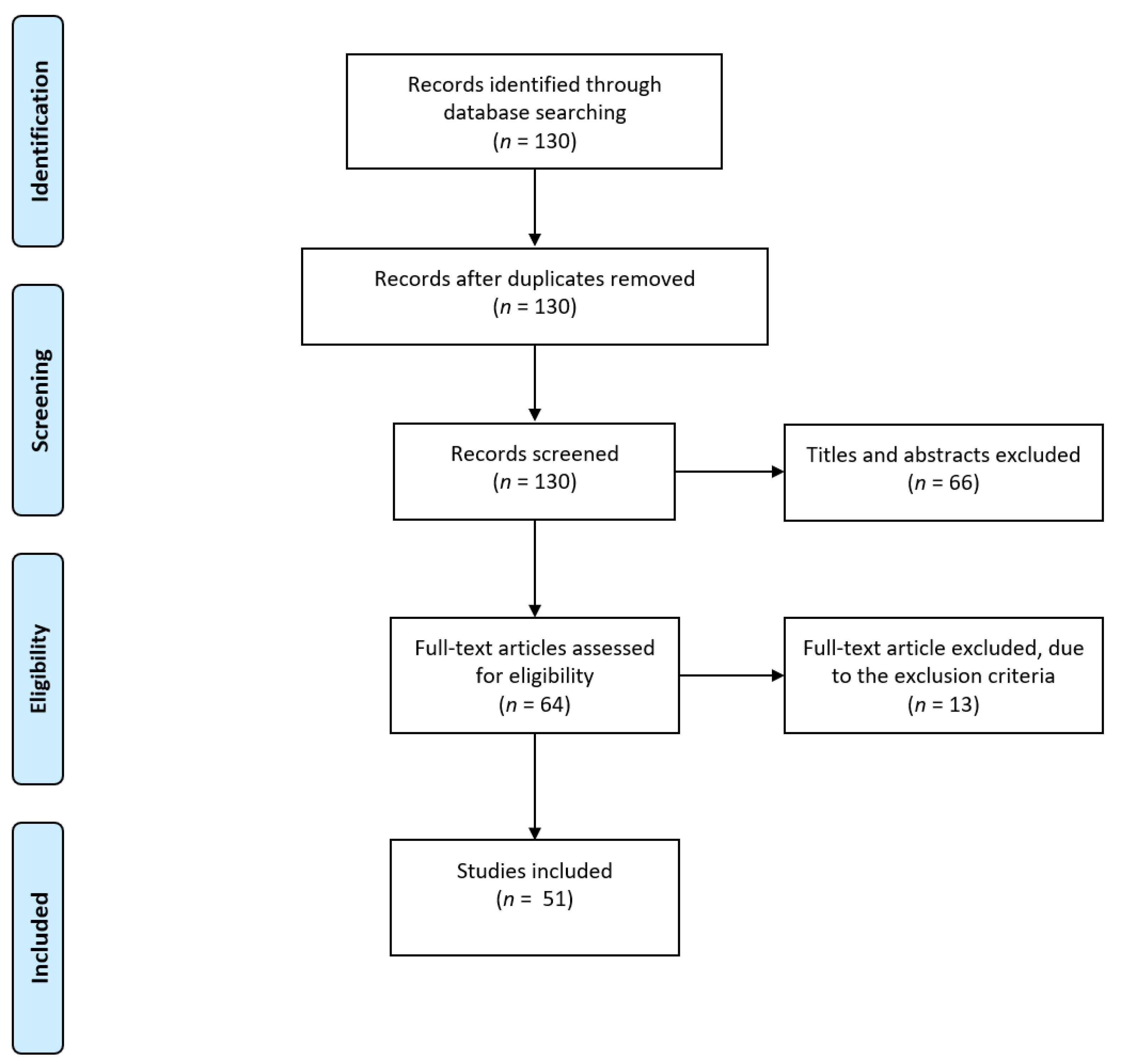
2.1. Searching Strategy
2.2. Inclusion and Exclusion Criteria
2.3. Study Selection
2.4. Study Characteristics
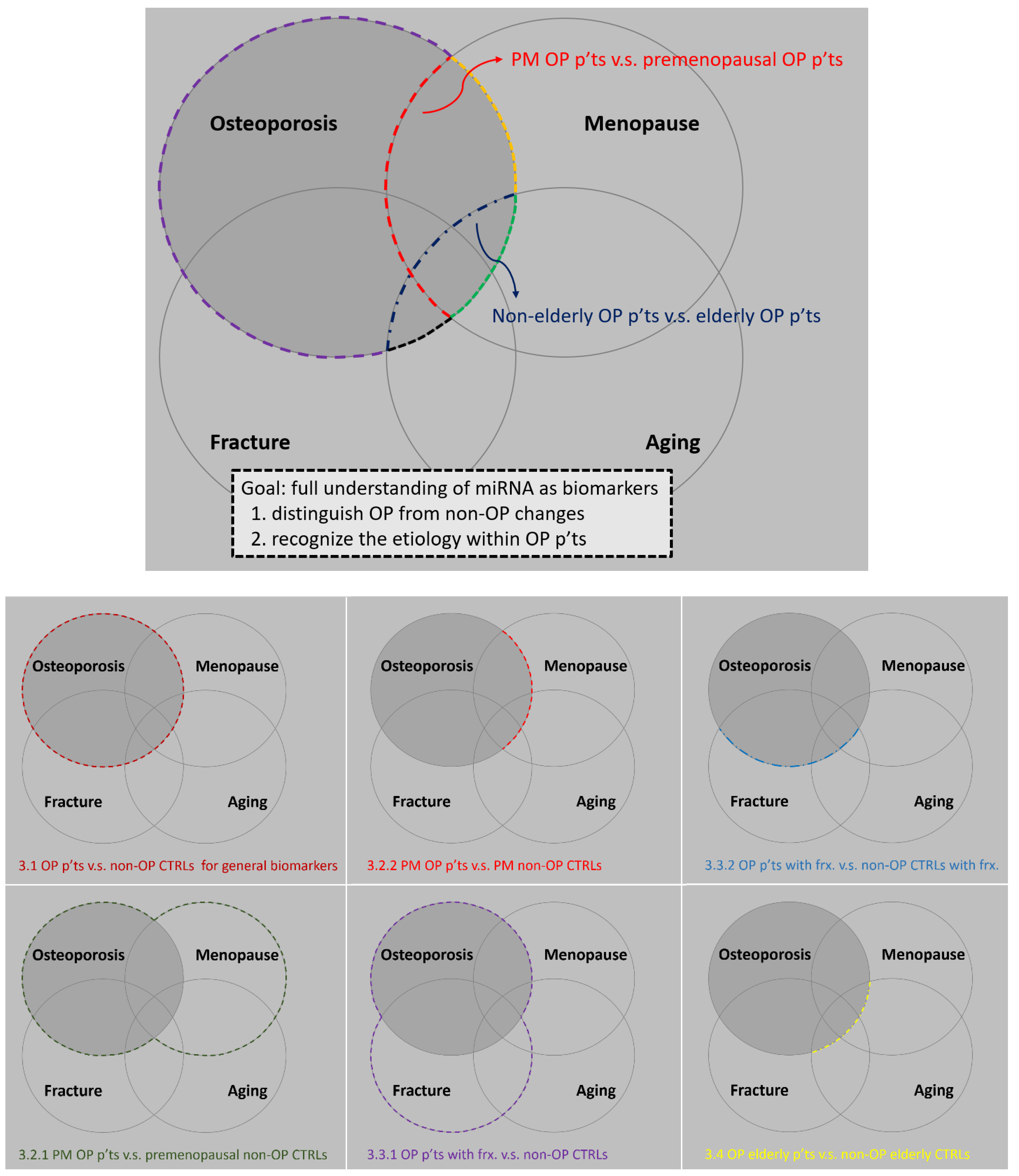
3. Serum miRNAs as Clinical Potential Biomarkers for Human Osteoporosis
3.1. General Biomarkers for Distinguishing Osteoporotic Patients from Non-Osteoporotic Controls
3.2. miRNAs That Are Potentially Associated with Estrogen
3.2.1. Studies That Aim to Accentuate the Role of Estrogen
3.2.2. Studies That Aim to Attenuate the Influence of Estrogen Itself
3.3. miRNAs That Are Potentially Associated with Fracture Healing
3.3.1. Study That Aims to Accentuate the Role of Sustained Fractures
- Comparison between osteoporotic patients with sustained low-traumatic fractures and controls [19]
3.3.2. Studies That Aim to Attenuate the Osteogenic Effect After a Recent Fracture
3.4. miRNAs That Potentially Associate with Osteoporosis in Elderly People
- Comparison between elderly osteoporotic patients and elderly controls [26].
3.5. miRNAs with High Diagnostic Value for Osteoporosis
- Index 1 = −0.394 + (0.105 × miR-30c-2-3p) + (−1.022 × miR-199a-5p) + (−0.078 × miR-424-5 p) + (− 0.046 × miR-497-5p) + (0.089 × miR-877-3p) (AUC: 0.86)
- Index 2 = (miR-30c-2-3p + miR-877-3p) − (miR-199a-5p + miR-424-5p) (AUC: 0.77)
4. Mechanisms of the Identified miRNAs
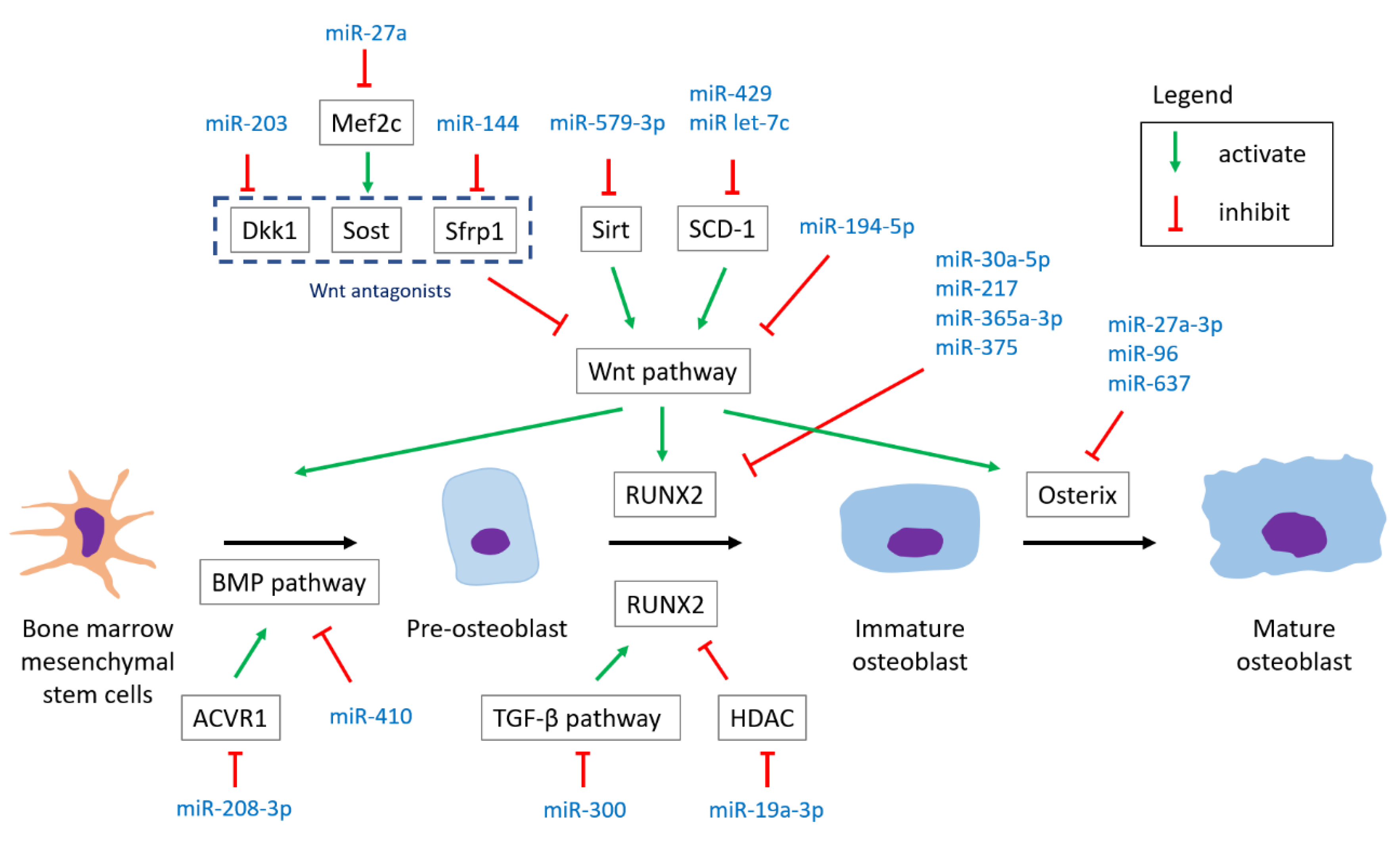
4.1. miRNAs Involved in Osteoblastogenesis
4.1.1. Wnt Pathway
- MiR-194-5p targets Wnt 5a, suppressing osteoblast differentiation. An in vivo study by micro-CT analysis disclosed that adult mice injected with miR-194-5p over femoral bone marrow had significantly decreased bone parameters (BMD and BV/TV) over the femur compared to controls three months later [53].
- MiR-144 and miR-203 target Wnt antagonists (Sfrp1 and DKK1, respectively), promoting osteoblast differentiation [28,43]. Besides, ovariectomized rats injected with antagomir-203 had decreased BMD over tibia and bone volume parameters compared to the control group injected with mutant antagomiR-203 six weeks after the injection [28].
- MiR-23b-3p targets MRC2 and is found to suppress Wnt signaling, inhibiting osteoblast differentiation. Although a relationship between MRC2 and Wnt signaling remained unclear, an in vivo study showed that OVX mice injected with the lenti-miR-23b-3p inhibitor had improved bone parameters [69].
4.1.2. TGF-β Pathway
- MiR-300 targets Smad3, inhibiting osteogenic differentiation. Moreover, micro-CT for evaluation in a rat model showed that miR-300 injections led to lower bone parameters (BMD and BV/TV) compared to sham and negative control groups [52].
4.1.3. BMP Pathway
- MiR-410 targets BMP-2, inhibiting osteoblast differentiation. Besides, the up-regulation of miR-410 was found in both postmenopausal osteoporotic patients compared to healthy controls and in the OVX mice group compared to the sham group [46].
4.1.4. Common Transcriptional Pathway
Runt-Related Transcription Factor 2 (RUNX2)
Osterix
- MiR-27a-3p, miR-96, and miR-637 target osterix, inhibiting osteogenic differentiation [26,59]. The experiment of a mice model showed that repetitively agomiR-96-injected young mice had significantly decreased BMD compared with vehicle-treated mice, and aged mice treated with antagomir-96 had higher bone strength compared to controls [26]. In addition, the signal transducer and activator of transcription 3 (Stat3) was found to be a pseudo-target of miR-637 by biological experiments [70].
4.1.5. Other Reactions in Osteogenic Differentiation
Histone Deacetylase (HDAC)
- MiR-19a-3p targets HDAC4, promoting osteogenic differentiation [32].
Phosphatase and Tensin Homolog (PTEN)/Phosphoinositide 3-Kinases (PI3K)/AKT Signaling Pathway
- MiR-19b targets PTEN, promoting osteogenic differentiation. Moreover, bone parameters including BMD, bone volume, and trabecular number were significantly higher in ovariectomized mice injected with agomiR-19b than those of the negative control group [57].
ATF3
- MiR-27a-3p targets ATF3, promoting osteogenic differentiation [35].
Glutaminase (GLS)
- MiR-200a-3p targets glutaminase, inhibiting osteogenic differentiation [41]
Activin A Receptor Type I (ACVR1)
- MiR-208a-3p targets ACVR1, inhibiting osteogenic differentiation. An in vivo study revealed that hip-limb-unloading (HLU) mice treated with antagomiR-208a-3p had higher parameters in bone formation and trabecular microarchitecture compared to the HLU control group [71].
Proprotein Convertase Subtilisin/Kexin Type 5 (PCSK5)
- MiR-338-3p targets PCSK5, inhibiting osteogenic differentiation [72].
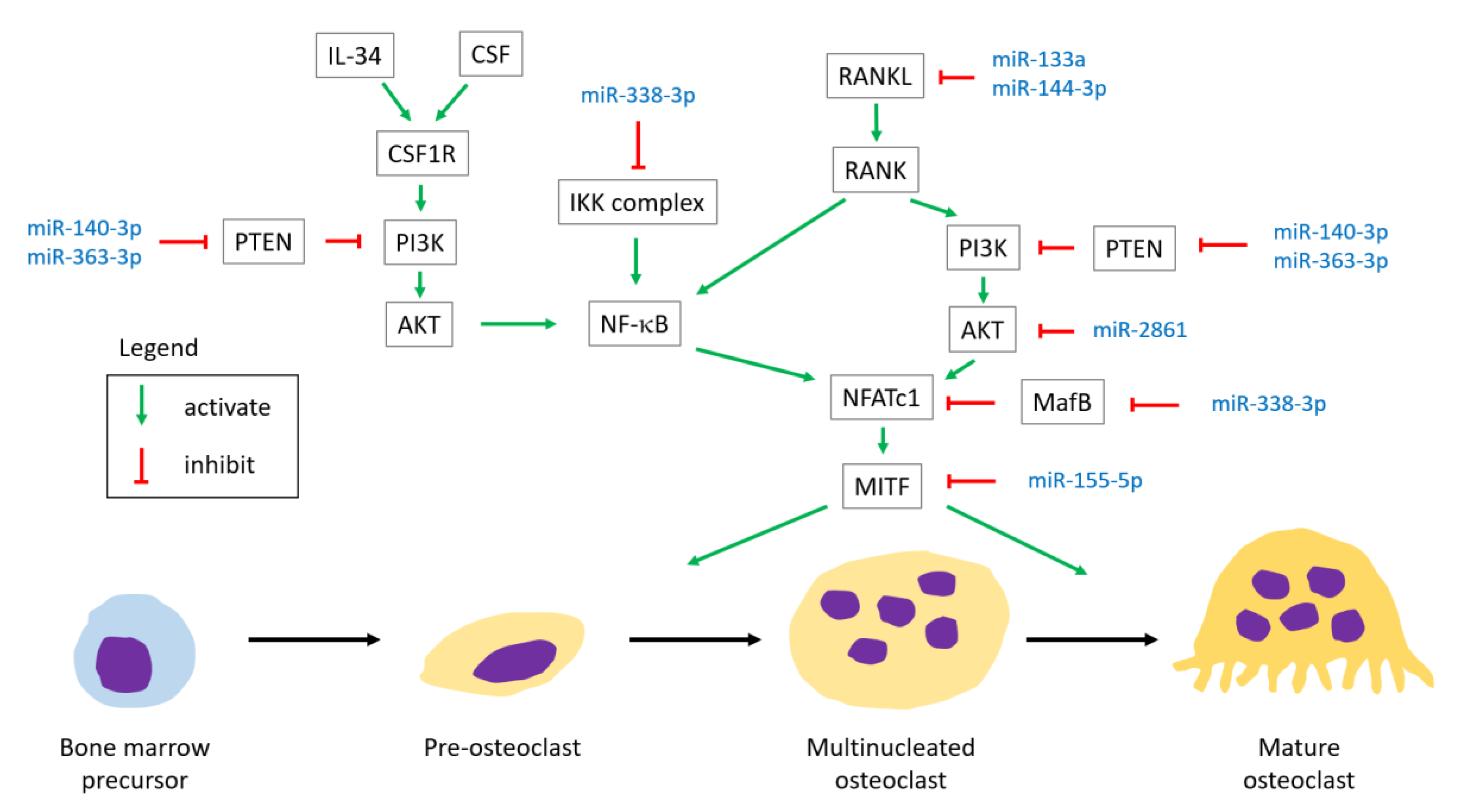
4.2. miRNAs Involved in Osteoclastogenesis
4.2.1. RANK
- MiR-144-3p targets RANK, inhibiting osteoclast differentiation [30].
- MiR-133a overexpression promotes RANKL-induced osteoclast differentiation. An in vivo study using a rat model showed that a significant decrease of osteoclastogenesis-related factors (M-CSF, RANKL, TNF-α, IL-1α, and CTX-I) was observed in OVX rats with a miR-133a knockdown compared to the controls [25].
- MiR-21 was found to correlate positively with RANKL level and the RANKL/OPG ratio, and correlate negatively with TGF-β1 and OPG by analysis of blood samples [42].
- MiR-338-3p targets MafB, promoting osteoclast differentiation [86].
4.2.2. Colony Stimulating Factor-1 Receptor (CSF1R)
4.2.3. Phosphatase and Tensin Homolog (PTEN)/Phosphoinositide 3-Kinases (PI3K)/AKT Signaling Pathway
- MiR-2861 targets AKT2, suppressing osteoclast differentiation [49].
4.2.4. Others
Microphthalmia Associated Transcription Factor (MITF)
- MiR-155-5p targets MITF, inhibiting osteoclast differentiation [87].
Inhibitor of Nuclear Factor Kappa-κ Kinase Subunit β (IKKβ) Gene
- MiR-338-3p targets IKKβ gene, inhibiting osteoclast differentiation [87].
5. miRNAs with Both Documented Diagnostic Values as Serum Biomarker for Osteoporosis and Identified Underlying Mechanisms
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Office of the Surgeon General. Reports of the surgeon general. In Bone Health and Osteoporosis: A Report of the Surgeon General; Office of the Surgeon General: Rockville, MD, USA, 2004. [Google Scholar]
- Burge, R.; Dawson-Hughes, B.; Solomon, D.H.; Wong, J.B.; King, A.; Tosteson, A. Incidence and economic burden of osteoporosis-related fractures in the united states, 2005–2025. J. Bone Min. Res. Off. J. Am. Soc. Bone Miner. Res. 2007, 22, 465–475. [Google Scholar] [CrossRef]
- Bottani, M.; Banfi, G.; Lombardi, G. Perspectives on mirnas as epigenetic markers in osteoporosis and bone fracture risk: A step forward in personalized diagnosis. Front. Genet. 2019, 10, 1044. [Google Scholar] [CrossRef]
- Bellavia, D.; De Luca, A.; Carina, V.; Costa, V.; Raimondi, L.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated mirnas in bone health: Epigenetic roles in osteoporosis. Bone 2019, 122, 52–75. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. Microrna. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microrna biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Huntzinger, E.; Izaurralde, E. Gene silencing by micrornas: Contributions of translational repression and mrna decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandão, B.B.; Kahn, C.R. Extracellular mirnas: From biomarkers to mediators of physiology and disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef]
- Backes, C.; Meese, E.; Keller, A. Specific mirna disease biomarkers in blood, serum and plasma: Challenges and prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef]
- Chen, G.; Deng, C.; Li, Y.P. Tgf-β and bmp signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Pala, E.; Denkçeken, T. Differentially expressed circulating mirnas in postmenopausal osteoporosis: A meta-analysis. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, Z.; Fu, Q.; Zhang, J. Plasma mirna levels correlate with sensitivity to bone mineral density in postmenopausal osteoporosis patients. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2014, 19, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; van Griensven, M. Five freely circulating mirnas and bone tissue mirnas are associated with osteoporotic fractures. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2014, 29, 1718–1728. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhang, D.; Pan, N.; Sun, N.; Wang, Q.; Fan, J.; Zhou, P.; Zhu, W.; Jiang, L. Identification of mir-194-5p as a potential biomarker for postmenopausal osteoporosis. PeerJ 2015, 3, e971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilner, S.; Skalicky, S.; Salzer, B.; Keider, V.; Wagner, M.; Hildner, F.; Gabriel, C.; Dovjak, P.; Pietschmann, P.; Grillari-Voglauer, R.; et al. Differentially circulating mirnas after recent osteoporotic fractures can influence osteogenic differentiation. Bone 2015, 79, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Bedene, A.; Mencej Bedrač, S.; Ješe, L.; Marc, J.; Vrtačnik, P.; Preželj, J.; Kocjan, T.; Kranjc, T.; Ostanek, B. Mir-148a the epigenetic regulator of bone homeostasis is increased in plasma of osteoporotic postmenopausal women. Wiener Klinische Wochenschrift 2016, 128, 519–526. [Google Scholar] [CrossRef]
- Chen, J.; Li, K.; Pang, Q.; Yang, C.; Zhang, H.; Wu, F.; Cao, H.; Liu, H.; Wan, Y.; Xia, W.; et al. Identification of suitable reference gene and biomarkers of serum mirnas for osteoporosis. Sci. Rep. 2016, 6, 36347. [Google Scholar] [CrossRef]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating microrna signatures in patients with idiopathic and postmenopausal osteoporosis and fragility fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef]
- Sun, W.; Zhao, C.; Li, Y.; Wang, L.; Nie, G.; Peng, J.; Wang, A.; Zhang, P.; Tian, W.; Li, Q.; et al. Osteoclast-derived microrna-containing exosomes selectively inhibit osteoblast activity. Cell Discov. 2016, 2, 16015. [Google Scholar] [CrossRef]
- You, L.; Pan, L.; Chen, L.; Gu, W.; Chen, J. Mir-27a is essential for the shift from osteogenic differentiation to adipogenic differentiation of mesenchymal stem cells in postmenopausal osteoporosis. Cell. Physiol. Biochem. 2016, 39, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Kelch, S.; Balmayor, E.R.; Seeliger, C.; Vester, H.; Kirschke, J.S.; van Griensven, M. Mirnas in bone tissue correlate to bone mineral density and circulating mirnas are gender independent in osteoporotic patients. Sci. Rep. 2017, 7, 15861. [Google Scholar] [CrossRef] [PubMed]
- Yavropoulou, M.P.; Anastasilakis, A.D.; Makras, P.; Tsalikakis, D.G.; Grammatiki, M.; Yovos, J.G. Expression of micrornas that regulate bone turnover in the serum of postmenopausal women with low bone mass and vertebral fractures. Eur. J. Endocrinol. 2017, 176, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Liao, X.; Chen, F.; Wang, B.; Huang, J.; Jian, G.; Huang, Z.; Yin, G.; Liu, H.; Jin, D. Circulating micrornas, mir-10b-5p, mir-328-3p, mir-100 and let-7, are associated with osteoblast differentiation in osteoporosis. Int. J. Clin. Exp. Pathol. 2018, 11, 1383–1390. [Google Scholar] [PubMed]
- Li, Z.; Zhang, W.; Huang, Y. Mirna-133a is involved in the regulation of postmenopausal osteoporosis through promoting osteoclast differentiation. Acta Biochim. Biophys. Sin. 2018, 50, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, Q.; Wu, X.P.; He, H.B.; Fu, L. Mir-96 regulates bone metabolism by targeting osterix. Clin. Exp. Pharm. Physiol. 2018, 45, 602–613. [Google Scholar] [CrossRef]
- Mandourah, A.Y.; Ranganath, L.; Barraclough, R.; Vinjamuri, S.; Hof, R.V.; Hamill, S.; Czanner, G.; Dera, A.A.; Wang, D.; Barraclough, D.L. Circulating micrornas as potential diagnostic biomarkers for osteoporosis. Sci. Rep. 2018, 8, 8421. [Google Scholar] [CrossRef]
- Qiao, L.; Liu, D.; Li, C.G.; Wang, Y.J. Mir-203 is essential for the shift from osteogenic differentiation to adipogenic differentiation of mesenchymal stem cells in postmenopausal osteoporosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5804–5814. [Google Scholar]
- Ramírez-Salazar, E.G.; Carrillo-Patiño, S.; Hidalgo-Bravo, A.; Rivera-Paredez, B.; Quiterio, M.; Ramírez-Palacios, P.; Patiño, N.; Valdés-Flores, M.; Salmerón, J.; Velázquez-Cruz, R. Serum mirnas mir-140-3p and mir-23b-3p as potential biomarkers for osteoporosis and osteoporotic fracture in postmenopausal mexican-mestizo women. Gene 2018, 679, 19–27. [Google Scholar] [CrossRef]
- Wang, C.; He, H.; Wang, L.; Jiang, Y.; Xu, Y. Reduced mir-144-3p expression in serum and bone mediates osteoporosis pathogenesis by targeting rank. Biochem. Cell Biol. 2018, 96, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.L.; Wang, Y.; Sun, Q.D.; Du, X.F. Mir-203 is involved in osteoporosis by regulating dkk1 and inhibiting osteogenic differentiation of mscs. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5098–5105. [Google Scholar]
- Chen, R.; Qiu, H.; Tong, Y.; Liao, F.; Hu, X.; Qiu, Y.; Liao, Y. Mirna-19a-3p alleviates the progression of osteoporosis by targeting hdac4 to promote the osteogenic differentiation of hmscs. Biochem. Biophys. Res. Commun. 2019, 516, 666–672. [Google Scholar] [CrossRef]
- Chen, Z.; Bemben, M.G.; Bemben, D.A. Bone and muscle specific circulating micrornas in postmenopausal women based on osteoporosis and sarcopenia status. Bone 2019, 120, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Yang, M.M.; Yang, R.H. Mirna-365a-3p promotes the progression of osteoporosis by inhibiting osteogenic differentiation via targeting runx2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7766–7774. [Google Scholar]
- Fu, Y.C.; Zhao, S.R.; Zhu, B.H.; Guo, S.S.; Wang, X.X. Mirna-27a-3p promotes osteogenic differentiation of human mesenchymal stem cells through targeting atf3. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 73–80. [Google Scholar]
- Lei, N.B.; Liang, X.; Wang, P.; Liu, Q.; Wang, W.G. Teriparatide alleviates osteoporosis by promoting osteogenic differentiation of hmscs via mir-375/runx2 axis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 11043–11050. [Google Scholar]
- Li, L.Y.; Wang, X.L.; Wang, G.S.; Zhao, H.Y. Mir-373 promotes the osteogenic differentiation of bmscs from the estrogen deficiency induced osteoporosis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7247–7255. [Google Scholar]
- Li, M.; Luo, R.; Yang, W.; Zhou, Z.; Li, C. Mir-363-3p is activated by myb and regulates osteoporosis pathogenesis via pten/pi3k/akt signaling pathway. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 376–386. [Google Scholar] [CrossRef]
- Lin, C.; Yu, S.; Jin, R.; Xiao, Y.; Pan, M.; Pei, F.; Zhu, X.; Huang, H.; Zhang, Z.; Chen, S.; et al. Circulating mir-338 cluster activities on osteoblast differentiation: Potential diagnostic and therapeutic targets for postmenopausal osteoporosis. Theranostics 2019, 9, 3780–3797. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Yang, J.F.; Wang, Y.H.; Qu, G.B.; Hao, P.D.; Zeng, Z.J.; Yuan, J.; Yang, R.; Yuan, Y. Microrna-579-3p promotes the progression of osteoporosis by inhibiting osteogenic differentiation of mesenchymal stem cells through regulating sirt1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6791–6799. [Google Scholar] [PubMed]
- Lv, R.; Pan, X.; Song, L.; Sun, Q.; Guo, C.; Zou, S.; Zhou, Q. Microrna-200a-3p accelerates the progression of osteoporosis by targeting glutaminase to inhibit osteogenic differentiation of bone marrow mesenchymal stem cells. Biomed. Pharm. 2019, 116, 108960. [Google Scholar] [CrossRef]
- Suarjana, I.N.; Isbagio, H.; Soewondo, P.; Rachman, I.A.; Sadikin, M.; Prihartono, J.; Malik, S.G.; Soeroso, J. The role of serum expression levels of microrna-21 on bone mineral density in hypostrogenic postmenopausal women with osteoporosis: Study on level of rankl, opg, tgfβ-1, sclerostin, rankl/opg ratio, and physical activity. Acta Med. Indones. 2019, 51, 245–252. [Google Scholar] [PubMed]
- Tang, L.; Lu, W.; Huang, J.; Tang, X.; Zhang, H.; Liu, S. Mir-144 promotes the proliferation and differentiation of bone mesenchymal stem cells by downregulating the expression of sfrp1. Mol. Med. Rep. 2019, 20, 270–280. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, Z.; Kang, N.; Yang, J.C.; Wei, X.; Hai, Y. Circ-vangl1 promotes the progression of osteoporosis by absorbing mirna-217 to regulate runx2 expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 949–957. [Google Scholar] [PubMed]
- Zhang, H.L.; Du, X.Y.; Dong, Q.R. Lncrna xixt promotes osteogenic differentiation of bone mesenchymal stem cells and alleviates osteoporosis progression by targeting mirna-30a-5p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8721–8729. [Google Scholar]
- Zhang, H.; Ding, W.; Ji, F.; Wu, D. Microrna-410 participates in the pathological process of postmenopausal osteoporosis by downregulating bone morphogenetic protein-2. Exp. Ther. Med. 2019, 18, 3659–3666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, X.; Zou, D.; Lian, Y.; Tian, S.; Dou, Z. Expression of microrna-21 in osteoporotic patients and its involvement in the regulation of osteogenic differentiation. Exp. Ther. Med. 2019, 17, 709–714. [Google Scholar] [CrossRef]
- Zhou, Z.; Lu, Y.; Wang, Y.; Du, L.; Zhang, Y.; Tao, J. Let-7c regulates proliferation and osteodifferentiation of human adipose-derived mesenchymal stem cells under oxidative stress by targeting scd-1. Am. J. Physiol. Cell Physiol. 2019, 316, C57–C69. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Wu, B.; Fan, S.; Liu, Y.; Ma, X.; Fu, X. Snhg14 induces osteogenic differentiation of human stromal (mesenchymal) stem cells in vitro by downregulating mir-2861. BMC Musculoskelet. Disord. 2020, 21, 525. [Google Scholar] [CrossRef]
- Gao, G.C.; Yang, D.W.; Liu, W. Lncrna terc alleviates the progression of osteoporosis by absorbing mirna-217 to upregulate runx2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 526–534. [Google Scholar]
- Ismail, S.M.; El Boghdady, N.A.; Hamoud, H.S.; Shabayek, M.I. Evaluation of circulating mirna-208a-3p, mirna-155-5p and mirna-637 as potential non-invasive biomarkers and the possible mechanistic insights into pre- and postmenopausal osteoporotic females. Arch. Biochem. Biophys. 2020, 684, 108331. [Google Scholar] [CrossRef] [PubMed]
- Kaur, T.; John, A.A.; Sharma, C.; Vashisht, N.K.; Singh, D.; Kapila, R.; Kapila, S. Mir300 intervenes smad3/β-catenin/runx2 crosstalk for therapy with an alternate function as indicative biomarker in osteoporosis. Bone 2021, 143, 115603. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.; Long, L.; Xie, K.; Liu, J.; Zhou, L.; Pan, S.; Liang, J.; Tu, Z.; Gao, Z.; Tang, Y. Mirna-429 suppresses osteogenic differentiation of human adipose-derived mesenchymal stem cells under oxidative stress via targeting scd-1. Exp. Ther. Med. 2020, 19, 696–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Chen, S.; Cai, P.; Chen, K.; Li, L.; Yang, X.; Yi, J.; Luo, X.; Du, Y.; Zheng, H. Mirna-483-5p is involved in the pathogenesis of osteoporosis by promoting osteoclast differentiation. Mol. Cell. Probes. 2020, 49, 101479. [Google Scholar] [CrossRef]
- Mi, B.; Yan, C.; Xue, H.; Chen, L.; Panayi, A.C.; Hu, L.; Hu, Y.; Cao, F.; Sun, Y.; Zhou, W.; et al. Inhibition of circulating mir-194-5p reverses osteoporosis through wnt5a/β-catenin-dependent induction of osteogenic differentiation. Mol. Ther. Nucleic Acids 2020, 21, 814–823. [Google Scholar] [CrossRef]
- Shuai, Y.; Liao, L.; Su, X.; Sha, N.; Li, X.; Wu, Y.; Jing, H.; Kuang, H.; Deng, Z.; Li, Y.; et al. Circulating micrornas in serum as novel biomarkers for osteoporosis: A case-control study. Ther. Adv. Musculoskelet. Dis. 2020, 12. [Google Scholar] [CrossRef]
- Sun, M.; Hu, L.; Wang, S.; Huang, T.; Zhang, M.; Yang, M.; Zhen, W.; Yang, D.; Lu, W.; Guan, M.; et al. Circulating microrna-19b identified from osteoporotic vertebral compression fracture patients increases bone formation. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2020, 35, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Bai, Y.; Zhang, Z.; Lu, J. A validated mirna signature for the diagnosis of osteoporosis related fractures using svm algorithm classification. Exp. Ther. Med. 2020, 20, 2209–2217. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, D.; Zhu, Z.; Li, L.; Jin, Y.; Ma, C.; Zhang, W. Mir-27a-3p negatively regulates osteogenic differentiation of mc3t3-e1 preosteoblasts by targeting osterix. Mol. Med. Rep. 2020, 22, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Jiang, J.; Deng, H.; Wang, Z.; Gu, R.; Wang, F. Mir-140-3p aggregates osteoporosis by targeting pten and activating pten/pi3k/akt signaling pathway. Hum. Cell 2020, 33, 569–581. [Google Scholar] [CrossRef]
- Yu, Y.; Yao, P.; Wang, Z.; Xie, W. Down-regulation of ftx promotes the differentiation of osteoclasts in osteoporosis through the notch1 signaling pathway by targeting mir-137. BMC Musculoskelet. Disord. 2020, 21, 456. [Google Scholar] [CrossRef] [PubMed]
- Zarecki, P.; Hackl, M.; Grillari, J.; Debono, M.; Eastell, R. Serum micrornas as novel biomarkers for osteoporotic vertebral fractures. Bone 2020, 130, 115105. [Google Scholar] [CrossRef]
- Zhou, J.G.; Hua, Y.; Liu, S.W.; Hu, W.Q.; Qian, R.; Xiong, L. Microrna-1286 inhibits osteogenic differentiation of mesenchymal stem cells to promote the progression of osteoporosis via regulating fzd4 expression. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1–10. [Google Scholar]
- Vrtačnik, P.; Ostanek, B.; Mencej-Bedrač, S.; Marc, J. The many faces of estrogen signaling. Biochem. Med. 2014, 24, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Nugent, M. Micrornas and fracture healing. Calcif. Tissue Int. 2017, 101, 355–361. [Google Scholar] [CrossRef]
- Kumar, S.; Vijayan, M.; Bhatti, J.S.; Reddy, P.H. Micrornas as peripheral biomarkers in aging and age-related diseases. Prog. Mol. Biol. Transl. Sci. 2017, 146, 47–94. [Google Scholar]
- Wagner, K.H.; Cameron-Smith, D.; Wessner, B.; Franzke, B. Biomarkers of aging: From function to molecular biology. Nutrients 2016, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, A.; Srinaath, N.; Rohini, M.; Selvamurugan, N. Regulation of runx2 by micrornas in osteoblast differentiation. Life Sci. 2019, 232, 116676. [Google Scholar] [CrossRef]
- Li, R.; Ruan, Q.; Yin, F.; Zhao, K. Mir-23b-3p promotes postmenopausal osteoporosis by targeting mrc2 and regulating the wnt/β-catenin signaling pathway. J. Pharmacol. Sci. 2021, 145, 69–78. [Google Scholar] [CrossRef]
- Zhang, J.F.; Fu, W.M.; He, M.L.; Wang, H.; Wang, W.M.; Yu, S.C.; Bian, X.W.; Zhou, J.; Lin, M.C.; Lu, G.; et al. Mir-637 maintains the balance between adipocytes and osteoblasts by directly targeting osterix. Mol. Biol. Cell 2011, 22, 3955–3961. [Google Scholar] [CrossRef] [Green Version]
- Arfat, Y.; Basra, M.A.R.; Shahzad, M.; Majeed, K.; Mahmood, N.; Munir, H. Mir-208a-3p suppresses osteoblast differentiation and inhibits bone formation by targeting acvr1. Mol. Ther. Nucleic Acids 2018, 11, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Zhang, M.; Li, X.; Ren, G. Microrna-338-3p regulates age-associated osteoporosis via targeting pcsk5. Mol. Med. Rep. 2021, 23, 136. [Google Scholar] [CrossRef] [PubMed]
- Karner, C.M.; Long, F. Wnt signaling and cellular metabolism in osteoblasts. Cell. Mol. Life Sci. 2017, 74, 1649–1657. [Google Scholar] [CrossRef]
- Sebastian, A.; Hum, N.R.; Morfin, C.; Murugesh, D.K.; Loots, G.G. Global gene expression analysis identifies mef2c as a potential player in wnt16-mediated transcriptional regulation. Gene 2018, 675, 312–321. [Google Scholar] [CrossRef]
- Simic, P.; Zainabadi, K.; Bell, E.; Sykes, D.B.; Saez, B.; Lotinun, S.; Baron, R.; Scadden, D.; Schipani, E.; Guarente, L. Sirt1 regulates differentiation of mesenchymal stem cells by deacetylating β-catenin. EMBO Mol. Med. 2013, 5, 430–440. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.P. Tgf-β and bmp signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Komori, T. Roles of runx2 in skeletal development. Adv. Exp. Med. Biol. 2017, 962, 83–93. [Google Scholar]
- Sinha, K.M.; Zhou, X. Genetic and molecular control of osterix in skeletal formation. J. Cell. Biochem. 2013, 114, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatani, T.; Chen, T.; Johnson, J.; Westendorf, J.J.; Partridge, N.C. The deletion of hdac4 in mouse osteoblasts influences both catabolic and anabolic effects in bone. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2018, 33, 1362–1375. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, T.M.; Westendorf, J.J. Histone deacetylase inhibitors promote osteoblast maturation. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2005, 20, 2254–2263. [Google Scholar] [CrossRef]
- Nielsen-Preiss, S.M.; Silva, S.R.; Gillette, J.M. Role of pten and akt in the regulation of growth and apoptosis in human osteoblastic cells. J. Cell. Biochem. 2003, 90, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.C. Atf3 mediates the inhibitory action of tnf-α on osteoblast differentiation through the jnk signaling pathway. Biochem. Biophys. Res. Commun. 2018, 499, 696–701. [Google Scholar] [CrossRef]
- Kamiya, N.; Kaartinen, V.M.; Mishina, Y. Loss-of-function of acvr1 in osteoblasts increases bone mass and activates canonical wnt signaling through suppression of wnt inhibitors sost and dkk1. Biochem. Biophys. Res. Commun. 2011, 414, 326–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoac, B.; Susan-Resiga, D.; Essalmani, R.; Marcinkiweicz, E.; Seidah, N.G.; McKee, M.D. Osteopontin as a novel substrate for the proprotein convertase 5/6 (pcsk5) in bone. Bone 2018, 107, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Szumska, D.; Pieles, G.; Essalmani, R.; Bilski, M.; Mesnard, D.; Kaur, K.; Franklyn, A.; El Omari, K.; Jefferis, J.; Bentham, J.; et al. Vacterl/caudal regression/currarino syndrome-like malformations in mice with mutation in the proprotein convertase pcsk5. Genes Dev. 2008, 22, 1465–1477. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhang, B.; Zhu, W.; Wei, W.; Ma, J.; Tay, F.R. A potential therapeutic target for regulating osteoporosis via suppression of osteoclast differentiation. J. Dent. 2019, 82, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.; Roy, M.; Bisson, M.; Nguyen, H.D.; Scott, M.S.; Boire, G.; Bouchard, L.; Roux, S. Osteoclast signaling-targeting mir-146a-3p and mir-155-5p are downregulated in paget’s disease of bone. Biochimica Et Biophysica Acta Mol. Basis Dis. 2020, 1866, 165852. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Gong, Z.; Sun, X.; Yuan, J.; Zheng, T.; Wang, X.; Fan, X.; Mao, Y.; Liu, X.; Tang, B.; et al. Mir-338-3p regulates osteoclastogenesis via targeting ikkβ gene. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 243–251. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current understanding of rank signaling in osteoclast differentiation and maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar]
- Boyce, B.F.; Xing, L. Functions of rankl/rank/opg in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Kim, J.H.; Lee, J.; Jin, H.M.; Kook, H.; Kim, K.K.; Lee, S.Y.; Kim, N. Mafb negatively regulates rankl-mediated osteoclast differentiation. Blood 2007, 109, 3253–3259. [Google Scholar] [CrossRef]
- Mun, S.H.; Park, P.S.U.; Park-Min, K.H. The m-csf receptor in osteoclasts and beyond. Exp. Mol. Med. 2020, 52, 1239–1254. [Google Scholar] [CrossRef]
- Hrdlicka, H.C.; Lee, S.K.; Delany, A.M. Micrornas are critical regulators of osteoclast differentiation. Curr. Mol. Biol. Rep. 2019, 5, 65–74. [Google Scholar] [CrossRef]
- Carracedo, A.; Pandolfi, P.P. The pten-pi3k pathway: Of feedbacks and cross-talks. Oncogene 2008, 27, 5527–5541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Zhang, W.; Wang, C.; Zhang, W.; Zhou, C.; Jiang, G.; Hong, J.; Yan, S.; Yan, W. Catalpol suppresses osteoclastogenesis and attenuates osteoclast-derived bone resorption by modulating pten activity. Biochem. Pharm. 2020, 171, 113715. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.Y.; Li, M.; Lin, Y.L. Mitf regulates osteoclastogenesis by modulating nfatc1 activity. Exp. Cell Res. 2014, 328, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, M.S.; Ghosh, S. Shared principles in nf-kappab signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Amer, Y. Nf-κb signaling and bone resorption. Osteoporos. Int. A. J. Establ. Result Coop. Between Eur. Found. Osteoporos. Natl. Osteoporos. Found. USA 2013, 24, 2377–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feurer, E.; Kan, C.; Croset, M.; Sornay-Rendu, E.; Chapurlat, R. Lack of association between select circulating mirnas and bone mass, turnover, and fractures: Data from the ofely cohort. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2019, 34, 1074–1085. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study/Reference | Main Studied miRNA | Sample Size | Patient Characteristics | Male: Female | Mean Age or Range of Age | Study Domain |
|---|---|---|---|---|---|---|
| Li et al. 2014 [13] | multiple | 120 | all PM female 40 OP p’ts v.s. 40 LBM p’ts v.s. 40 CTRLs | all female | 57.5 (OP), 56.7 (LBM), 56.5 (CTRL) | human blood sample |
| Seeliger et al. 2014 [14] | multiple | 63 | all have a hip frx 33 OP p’ts v.s. 30 non-OP CTRLs | 3: 60 | NA | human blood sample human bone tissue sample |
| Meng et al. 2015 [15] | miR-194-5p | 48 (discovery) 86 (validation) | 25 OP p’ts v.s. 23 LBM p’ts 24 OP p’ts v.s. 30 LBM p’ts v.s. 32 CTRLs | all female | 66.1 (OP), 64.7 (LBM) 64.0 (all 3 groups) | human blood sample |
| Weilner et al. 2015 [16] | multiple | 14 (discovery) 23 (validation) | 7 OP p’ts v.s. 7 CTRLs | all female | 72.4 (OP), 71.0 (CTRL) 77.8 (OP), 81.5 (CTRL) | human blood sample in vitro: human ASC |
| Bedene et al. 2016 [17] | miR-148a | 74 | all PM female 17 OP p’ts v.s. 57 CTRLs | all female | 62.0 (OP), 61.0 (CTRL) | human blood sample |
| Chen et al. 2016 [18] | multiple | 36 | all PM female 19 OP p’ts v.s. 7 LBM p’ts v.s. 10 CTRLs | all female | 77.4 (OP), 72.86 (LBM), 51.89 (CTRL) | human blood sample animal blood sample |
| Kocijan et al. 2016 [19] | miR-29b-3p | 75 | 36 p’ts with low-traumatic frx v.s. 39 CTRLs | 20: 16 (OP) 23: 16 (CTRL) | 46.6 (OP), 46.6 (CTRL) | human blood sample |
| Sun et al. 2016 [20] | miR-214 | 65 | 42 OP p’ts v.s. 23 CTRLs | NA | Men: 50–90 years old Woman: over 5 years of menopause | human blood sample in vitro: human PBMCs in vivo: mice model |
| You et al. 2016 [21] | miR-27a | 155 | 81 OP PM p’ts v.s. 74 premenopausal CTRLs | all female | 65.8 (OP), 43.3 (CTRL) | human blood sample in vitro: human MSC in vivo: mice model |
| Kelch et al. 2017 [22] | multiple | 28 | 7 female OP p’ts v.s. 7 female CTRLs v.s. 7 male OP p’ts v.s. 7 male CTRLs | 14: 14 | 81.9 (♀OP), 71.2 (♀CTRLs), 78.0 (♂OP), 68.6 (♂CTRLs) | human blood sample in vitro: osteoblast isolation & human PBMCs |
| Yavropoulou et al. 2017 [23] | miR-21-5p | 100 | all PM female 35 p’ts with LBM and vertebral frx v.s. 35 p’ts with LBM without frx v.s. 30 CTRLs | all female | 68 (frx.), 71 (no frx.), 68 (CTRL) | human blood sample |
| Chen et al. 2018 [24] | multiple | 18 | 9 OP p’ts v.s. 9 CTRLs | NA | 69.2 (OP), 67.1 (CTRL) | human blood sample in vitro: human & mice osteoblast cells |
| Li et al. 2018 [25] | miR-133a | 20 | all PM female 10 OP p’ts v.s. 10 CTRLs | all female | 59-80 (OP), 62–75 (CTRL) | human blood sample in vitro: human monocytic & murine macrophage cells in vivo: rats model |
| Liu et al. 2018 [26] | miR-96 | 80 | 20 PM OP p’t v.s. 20 premenopausal CTRLs; 20 elderly OP p’ts v.s. 20 elderly CTRLs | NA | 45–60 65–80 | human blood sample in vitro: human & mice MSC in vivo: mice model |
| Mandourah et al. 2018 [27] | miR-122-5p miR-4516 | 161 | 53 OP p’ts v.s. 78 LBM p’ts v.s. 30 CTRLs | 30: 131 | 69.1 (OP), 65.9 (LBM), 67.0 (CTRL) | human blood sample |
| Qiao et al. 2018 [28] | miR-203 | 100 | 60 PM OP p’ts v.s. 40 CTRLs | NA | 63.4 (OP), 59.3 (CTRL) | human blood sample in vitro: human MSC in vivo: rats model |
| Ramírez-Salazar et al. 2018 [29] | miR-140-3p miR-23b-3p | 40 (discovery) 97 (validation) | 20 OP p’ts v.s. 20 CTRLs 21 OP p’ts with frx. v.s. 26 OP p’ts without frx. v.s. 28 LBM p’ts v.s. 22 CTRLs | all female | 73.8 (OP), 71.1 (CTRL) 82.5 (OP with frx.), 68.9 (OP without frx.), 64.3 (LBM), 60.5 (CTRL) | human blood sample |
| Wang et al. 2018 [30] | miR-144-3p | 60 | all have a hip frx 45 OP p’ts v.s. 15 non-OP CTRLs | NA | NA | human blood sample in vitro: human PBMC |
| Xia et al. 2018 [31] | miR-203 | 120 | 60 OP p’ts v.s. 60 CTRLs | all female | NA | human blood sample in vitro: rat MSC |
| Chen et al. 2019 (a) [32] | miR-19a-3p | 84 | 42 OP p’ts v.s. 42 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Chen et al. 2019 (b) [33] | multiple | 75 | all PM female sacropenic p’t (1) v.s. sacropenic OP p’ts (15) v.s. OP p’t (46) v.s. CTRL (13) | all female | 85.8 (sacropenic), 68.9 (sacropenic OP), 69.6 (OP), 68.9 (CTRL) | human blood sample |
| Cheng et al. 2019 [34] | miR-365a-3p | 60 | 30 OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Fu et al. 2019 [35] | miR-27a-3p | 40 | 20 OP p’ts v.s. 20 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Lei et al. 2019 [36] | miR-375 | 60 | 30 OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Li et al. 2019 (a) [37] | miR-373 | 40 | 20 PM OP p’ts v.s. 20 CTRLs | NA | NA | human blood sample human bone tissue sample in vitro: rats PBMC in vivo: rats model |
| Li et al. 2019 (b) [38] | miR-363-3p | 12 | all p’ts have a frx 6 OP p’ts v.s. 6 non-OP CTRLs | NA | NA | human blood sample in vitro: human PBMC, C2C12 cells |
| Lin et al. 2019 [39] | miR-338 cluster | 30 | 15 PM OP p’ts v.s. 15 CTRLs | all female | 58–68 | human blood sample in vitro: mice PBMC in vivo: mice model |
| Luo et al. 2019 [40] | miR-579-3p | NA | OP p’ts v.s. CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Lv et al. 2019 [41] | miR-200a-3p | 60 | 30 OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Suarjana et al. 2019 [42] | miR-21 | 120 | all PM hypoestrogenic female 60 OP p’ts v.s. 60 non-OP CTRLs | all female | 62 (OP), 58.5 (CTRL) | human blood sample |
| Tang et al. 2019 [43] | miR-144 | 30 | all PM female 15 OP p’ts v.s. 15 CTRLs | all female | 54–64 | human blood sample in vitro: rats MSC |
| Yang et al. 2019 [44] | miR-217 | 30 | 15 OP p’ts v.s. 15 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Zhang et al. 2019 (a) [45] | miR-30a-5p | NA | OP p’ts v.s. CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Zhang et al. 2019 (b) [46] | miR-410 | 55 | 26 PM OP p’ts v.s. 29 CTRLs | all female | 55.6 (OP), 55.1 (CTRL) | human blood sample in vitro: human & mice PBMC |
| Zhao et al. 2019 [47] | miR-21 | 96 | 48 OP p’ts v.s. 48 CTRLs | NA | NA | human blood sample in vitro: rats MSC |
| Zhou et al. 2019 [48] | miR let-7c | 144 | 99 PM OP p’ts v.s. 45 premenopausal CTRLs | all female | 40–65 | human blood sample in vitro: human ASC |
| Du et al. 2020 [49] | miR-2861 | 40 | 20 OP p’ts v.s. 20 CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Gao et al. 2020 [50] | miR-217 | NA | OP p’ts v.s. CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Ismail et al. 2020 [51] | miR-208a-3p miR-155-5p miR-637 | 140 | 70 OP p’ts v.s. 70 CTRLs | all female | 61.3 (PM OP), 36.0 (premenopausal OP), 60.1 (PM CTRL), 34.0 (premenopausal CTRL) | human blood sample |
| Kaur et al. 2020 [52] | miR-300 | 60 | 30 OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample In vitro: human & rat osteoblast cells In vivo: rat model |
| Lan et al. 2020 [53] | miR-429 | 60 | 30 OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample in vitro: human ASC |
| Li et al. 2020 [54] | miR-483-5p | 72 | all have a hip frx 36 OP p’ts v.s. 36 non-OP CTRLs | all female | 62 (OP), 59 (CTRL) | human blood sample in vitro: human PBMC |
| Mi et al. 2020 [55] | miR-194-5p | 100 | 50 OP p’ts v.s. 50 non-OP CTRLs | NA | NA | human blood sample in vitro: mice MSC |
| Shuai et al. 2020 [56] | multiple | 25 (discovery) 288 (training) 160(validation) | 5 OP p’ts v.s. 10 LBM p’ts v.s. 10 CTRLs 86 OP p’ts v.s. 76 LBM p’ts v.s. 126 CTRLs 48 OP p’ts v.s. 56 LBM p’ts v.s. 56 CTRLs | NA 139: 149 73: 87 | 19–80 | human blood sample |
| Sun et al. 2020 [57] | miR-19b | 18 (discovery) 72 (validation) | 6 OP p’ts v.s. 6 LBM p’ts v.s. 6 CTRLs 24 OP p’ts v.s. 24 LBM p’ts v.s. 24 CTRLs | 3: 17 12: 60 | 73.1 (OP), 66.5 (LBM), 46.1 (CTRL) | human blood sample in vitro: human MSC and mice cell in vivo: mice model |
| Tang et al. 2020 [58] | multiple | 36 | all PM female 19 OP p’ts v.s. 17 CTRLs | all female | 64.7 (OP) v.s. 58.1 (CTRL) | human blood sample human bone tissue in vitro: human osteoblast |
| Xu et al. 2020 [59] | miR-27a-3p | 137 | 85 OP p’ts v.s. 52 CTRLs | all female | 50–90 | human blood sample In vitro: mice cell |
| Yin et al. 2020 [60] | miR-140-3p | 60 | 30 PM OP p’ts v.s. 30 CTRLs | NA | NA | human blood sample in vitro: human PBMC & C2C12 cell |
| Yu et al. 2020 [61] | miR-137 | 51 | 30 OP p’ts with frx. v.s. 21 CTRLs | 14: 37 | 60.8 (OP), 62 (CTRL) | human blood sample in vitro: human PBMC |
| Zarecki et al. 2020 [62] | multiple | 116 | all PM female 24 OP p’ts with frx. v.s. 17 OP p’ts with frx. under tx v.s. 35 LBM p’ts without frx. v.s. 40 CTRLs | all female | 69.6 (OP with frx.), 69.6 (OP with frx. under treatment), 67.9 (PM LBM without frx.), 68.8 (CTRL) | human blood sample |
| Zhou et al. 2020 [63] | miR-1286 | NA | OP p’ts v.s. CTRLs | NA | NA | human blood sample in vitro: human MSC |
| Osteoporotic Patients versus Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-10b-5p | [24] | miR-19a-3p miR-21 miR-22-3p miR-27a-3p miR-122-5p miR-133b miR-203 miR-328-3p miR-518 miR-2861 miR-4516 miR-let-7g-5p | [32] [47] [16,24] [35] [27] [24] [31] [16,24] [24] [24] [27] [16,24] | ||
| miR-21 | [24] | ||||
| miR-21-5p | linearly correlate with BMD | [22] | |||
| miR-23 | [24] | ||||
| miR-23b-3p | correlate with low BMD | [29] | |||
| miR-24-3p | linearly correlate with BMD | [22] | |||
| miR-27a-3p | [59] | ||||
| miR-30a-5p | may negatively correlate with XIXT | [45] | |||
| miR-93-5p | linearly correlate with BMD | [22] | |||
| miR-100 | [24] | ||||
| miR-100-5p | linearly correlate with BMD | [22] | |||
| miR-125b | [24] | ||||
| miR-125b-5p | linearly correlate with BMD | [22] | |||
| miR-137 | [61] | ||||
| miR-140-3p | correlate with low BMD | [29] | |||
| miR-155-5p | [51] | ||||
| miR-194-5p | [55] | ||||
| miR-200a-3p | [41] | ||||
| miR-208a-3p | [51] | ||||
| miR-214 | [20] | ||||
| miR-217 | negatively correlate with RUNX2 | [44] | |||
| may negatively correlate with TERC | [50] | ||||
| miR-300 | [52] | ||||
| miR-365a-3 | [34] | ||||
| miR-375 | [36] | ||||
| miR-429 | [53] | ||||
| miR-579-3p | [40] | ||||
| miR-637 | [51] | ||||
| miR-1286 | [63] | ||||
| Osteoporotic Patients versus Osteopenia Patients versus Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-30c-2-3p | [56] | miR-19b | [57] | ||
| miR-130b-3p | negatively correlate with BMD | [15] | miR-30b-5p | positively correlate with hip BMD | [18] |
| miR-151a-3p | negatively correlate with BMD | [15] | miR-103-3p | positively correlate with hip BMD | [18] |
| miR-151b | negatively correlate with BMD | [15] | miR-142-3p | positively correlate with hip BMD | [18] |
| miR-194-5p | negatively correlate with BMD | [15,56] | miR-199a-5p | [56] | |
| miR-497-5p | [56] | miR-328-3p | positively correlate with hip BMD | [18] | |
| miR-590-5p | [15] | miR-424-5p | [56] | ||
| miR-660-5p | [15] | ||||
| miR-877-3p | [56] | ||||
| Postmenopausal Osteoporotic Patients versus Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-133a | negatively correlate with lumbar spine BMD | [25] | miR-28 | [37] | |
| miR-101 | [37] | ||||
| miR-140-3p | negatively correlate with PTEN | [60] | miR-203 | [28] | |
| miR-338-3p | [39] | miR-373 | [37] | ||
| miR-410 | may negatively correlate with BMP-2 | [46] | |||
| miR-3065-5p | [39] | ||||
| Postmenopausal Osteoporotic Patients versus Premenopausal Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR let-7c | [48] | miR-27a | [21] | ||
| Postmenopausal Osteoporotic Patients versus Postmenopausal Non-Osteoporotic Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-21 | negatively correlate with BMD positively correlate with both RANKL and RANKL/OPG ratio | [42] | miR-21 | positively correlate with hip and spine BMDs | [13] |
| miR-21-5p | negatively correlate with lumbar spine aBMD | [33] | miR-125b-5p | positively correlate with age | [33] |
| miR-23a-3p | positively correlate with TRAP5b | [33] | miR-330-3p | [58] | |
| miR-133a | negatively correlate with hip and spine BMDs | [13] | |||
| miR-135a-5p | [58] | ||||
| miR-144 | positively correlate with Sfrp1 | [43] | |||
| miR-148a | [17] | ||||
| miR-181a-3p | [58] | ||||
| miR-188-3p | [58] | ||||
| miR-194-5p | [15] | ||||
| miR-576-3p | [58] | ||||
| miR-942 | [58] | ||||
| Postmenopausal Osteoporotic Patients with Fracture versus Postmenopausal Osteoporotic Patients without Fracture versus Postmenopausal Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-19b-3p | positively correlate with serum levels of osteocalcin, ALP, and CTX | [62] | miR-21-5p | not found to correlate with BMD | [23] |
| miR-21-5p | [62] | miR-23a-3p | not found to correlate with BMD | [23] | |
| miR-23a-3p | [62] | miR-29a-3p | not found to correlate with BMD | [23] | |
| miR-124-3p | not found to correlate with BMD | [23] | |||
| miR-152-3p | [62] | ||||
| miR-335-5p | [62] | ||||
| miR-375 | [62] | ||||
| miR-532-3p | positively correlate with ALP | [62] | |||
| miR-2861 | not found to correlate with BMD | [23] | |||
| Osteoporotic Patients with Low-Traumatic Fracture versus Controls | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-152–3p | [19] | miR-19a-3p | [19] | ||
| miR-335–5p | [19] | miR-19b-3p | correlated with lumbar spine aBMD | [19] | |
| miR-30e-5p | [19] | ||||
| miR-140–5p | [19] | ||||
| miR-324–3p | correlated with lumbar spine aBMD | [19] | |||
| miR-550a-3p | [19] | ||||
| Osteoporotic Patients with Fracture versus Non-Osteoporotic Controls with Fracture | |||||
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-21 | [14] | miR-144-3p | [37] | ||
| miR-23a | [14] | ||||
| miR-24 | [14] | ||||
| miR-24-3p | [30] | ||||
| miR-25 | [14] | ||||
| miR-27a-3p | [30] | ||||
| miR-93 | [14] | ||||
| miR-100 | [14,30] | ||||
| miR-122a | [14,30] | ||||
| miR-124a | [14] | ||||
| miR-125b | [14,30] | ||||
| miR-148a | [14] | ||||
| miR-363-3p | [38] | ||||
| miR-483-5p | may negatively correlate with IGF2 | [54] | |||
| Elderly Osteoporotic Patients versus Elderly Controls | |||||
|---|---|---|---|---|---|
| Up-Regulated | Down-Regulated | ||||
| MiRNA | Correlation | Ref. | MiRNA | Correlation | Ref. |
| miR-96 | [26] | ||||
| miR-107 | [26] | ||||
| Study Setting | MiRNA | Area under Curve (AUC) | Sensitivity | Specificity | Reference |
|---|---|---|---|---|---|
| Osteoporotic patients v.s. controls | miR-10b-5p | 0.87 | NA | NA | [24] |
| miR-23b-3p | 0.69 | NA | NA | [29] | |
| miR-100 | 0.89 | NA | NA | [24] | |
| miR-140-3p | 0.92 | NA | NA | [29] | |
| miR-300 | 0.969 | NA | NA | [52] | |
| miR-328-3p | 0.87 | NA | NA | [24] | |
| miR-4516 | 0.727 | 71% | 62% | [27] | |
| let-7g-5p | 0.89 | NA | NA | [24] | |
| Premenopausal osteoporotic patients v.s. controls | miR-155-5p | 0.9 | 94.29% | 77.14% | [51] |
| miR-208a-3p | 0.816 | 77.14% | 82.86% | [51] | |
| Postmenopausal osteoporotic patients v.s. controls | miR-135a-5p | 0.759 | NA | NA | [58] |
| miR-155-5p | 0.828 | 80% | 80% | [51] | |
| miR-181a-3p | 0.817 | NA | NA | [58] | |
| miR-188-3p | 0.889 | NA | NA | [58] | |
| miR-208a-3p | 0.851 | 80% | 82.86% | [51] | |
| miR-338-3p | 0.74 | NA | NA | [39] | |
| miR-576-3p | 0.751 | NA | NA | [58] | |
| miR-637 | 0.814 | 77.14% | 85.71% | [51] | |
| miR-942-3p | 0.678 | NA | NA | [58] | |
| miR-3065-5p | 0.87 | NA | NA | [39] | |
| Postmenopausal osteoporotic patients with fracture. v.s. | miR-21-5p | 0.66 | 66% | 71% | [23] |
| postmenopausal osteoporotic patients without fracture v.s. | |||||
| postmenopausal controls | |||||
| Osteoporotic patients with fracture v.s. non-osteoporotic controls with fracture | miR-122a | 0.77 | 74.14% | 72.14% | [14] |
| Involved Pathways | MiRNA | Target | Effect to Osteoblasto-Genesis | In Vitro Evidence | In Vivo Evidence | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|
| Regulation of Target Gene Confirmed by miRNA Mimics or Inhibitor Transfection | Effect of miRNA Altered by Overexpression, Knockdown or Silence of the Target Gene | By DXA | By micro-CT | ||||||
| Wnt | miR-23b-3p | MRC2 | inhibition | v (wild type v.s. mutant UTR) | v | Mice model | [69] | ||
| v | v | ||||||||
| miR-27a | Mef2c | promotion | v (wild type v.s. mutant UTR) | v | Mice model | [21] | |||
| v | v | ||||||||
| miR-144 | Sfrp1 | promotion | v (wild type v.s. mutant UTR) | v | [43] | ||||
| miR-194-5p | Wnt 5a | inhibition | v (wild type v.s. mutant UTR) | Mice model | [55] | ||||
| v | |||||||||
| miR-203 | DKK1 | promotion | v (wild type v.s. mutant UTR) | Rat model | [28,31] | ||||
| v | |||||||||
| miR-429 | SCD-1 | inhibition | v (wild type v.s. mutant UTR) | v | [53] | ||||
| miR-579-3p | Sirt | inhibition | v (wild type v.s. mutant UTR) | v | [40] | ||||
| miR let-7c | SCD-1 | inhibition | v (wild type v.s. mutant UTR) | v | [48] | ||||
| TGF-β | miR-300 | Smad | inhibition | v | Rat model | [52] | |||
| v | v | ||||||||
| BMP | miR-410 | BMP-2 | inhibition | v (wild type v.s. mutant UTR) | [46] | ||||
| Common regulatory factors | miR-30a-5p | RUNX2 | inhibition | v (wild type v.s. mutant UTR) | v | [45] | |||
| miR-217 | RUNX2 | v (wild type v.s. mutant UTR) | v | [50] [44] | |||||
| miR-365a-3p | RUNX2 | v (wild type v.s. mutant UTR) | v | [34] | |||||
| miR-375 | RUNX2 | v (wild type v.s. mutant UTR) | v | [36] | |||||
| miR-27a-3p * | osterix | inhibition | v (wild type v.s. mutant UTR) | v | [59] | ||||
| miR-96 | osterix | v (wild type v.s. mutant UTR) | v | Mice model | [26] | ||||
| miR-637 | osterix | v (wild type v.s. mutant UTR) | [70] | ||||||
| Others | miR-19a-3p | HDAC4 | promotion | v (wild type v.s. mutant UTR) | v | [32] | |||
| miR-19b | PTEN | promotion | v (wild type v.s. mutant UTR) | Mice model | [57] | ||||
| v | |||||||||
| miR-27a-3p * | ATF3 | promotion | v (wild type v.s. mutant UTR) | v | [35] | ||||
| miR-200a-3p | glutaminase | inhibition | v (wild type v.s. mutant UTR) | v | [41] | ||||
| miR-208a-3p | ACVR1 | inhibition | v (wild type v.s. mutant UTR) | Mice model | [71] | ||||
| v | |||||||||
| miR-338-3p | PCSK5 | inhibition | v (wild type v.s. mutant UTR) | v | [72] | ||||
| Involved Pathways | MiRNA | Target | Effect to Osteoclasto- Genesis | In Vitro Evidence | In Vivo Evidence | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| Regulation of Target Gene Confirmed by miRNA Mimics or Inhibitor Transfection | Effect of miRNA Altered by Overexpression, Knockdown or Silence of the Target Gene | By DXA | By Micro-CT | |||||
| RANK | miR-133a | promotion | v | Rat model | [25] | |||
| v | ||||||||
| miR-144-3p | SMAD4 RANK | inhibition | v (wild type v.s. mutant UTR) | [30] | ||||
| miR-338-3p * | MafB | promotion | v (wild type v.s. mutant UTR) | v | [86] | |||
| PTEN/PI3K/AKT signaling pathway | miR-140-3p | PTEN | promotion | v (wild type v.s. mutant UTR) | v | [60] | ||
| miR-363-3p | PTEN | v (wild type v.s. mutant UTR) | v | [38] | ||||
| miR-2861 | AKT2 | inhibition | v | [49] | ||||
| Others | miR-155-5p | MITF | inhibition | v | [87] | |||
| miR-338-3p * | IKKβ gene | inhibition | v (wild type v.s. mutant UTR) | [88] | ||||
| MiRNA | Clinical Setting | Mechanism (Target) | Conflicting Results |
|---|---|---|---|
| miR-23b-3p | OP p’ts v.s. CTRLs | Inhibition of osteoblastogenesis (MRC2) | |
| miR-140-3p | OP p’ts v.s. CTRLs | Promotion of osteoclastogenesis (PTEN) | |
| miR-300 | OP p’ts v.s. CTRLs | Inhibition of osteoblastogenesis (Smad) | |
| miR-155-5p | PM OP p’ts v.s. CTRLs | Inhibition of osteoclastogenesis (MITF) | |
| miR-208a-3p | PM OP p’ts v.s. CTRLs | Inhibition of osteoblastogenesis (ACVR1) | |
| miR-338-3p | PM OP p’ts v.s. CTRLs | Inhibition of osteoblastogenesis (PCSK5) Conflicting results on osteoclastogenesis (MafB, IKKβ) | v |
| miR-637 | PM OP p’ts v.s. CTRLs | Inhibition of osteoblastogenesis (osterix) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-Z.; Huang, H.-T.; Cheng, T.-L.; Lu, Y.-M.; Lin, S.-Y.; Ho, C.-J.; Lee, T.-C.; Hsu, C.-H.; Huang, P.-J.; Huang, H.H.; et al. Application of microRNA in Human Osteoporosis and Fragility Fracture: A Systemic Review of Literatures. Int. J. Mol. Sci. 2021, 22, 5232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105232
Wu Y-Z, Huang H-T, Cheng T-L, Lu Y-M, Lin S-Y, Ho C-J, Lee T-C, Hsu C-H, Huang P-J, Huang HH, et al. Application of microRNA in Human Osteoporosis and Fragility Fracture: A Systemic Review of Literatures. International Journal of Molecular Sciences. 2021; 22(10):5232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105232
Chicago/Turabian StyleWu, Yen-Zung, Hsuan-Ti Huang, Tsung-Lin Cheng, Yen-Mou Lu, Sung-Yen Lin, Cheng-Jung Ho, Tien-Ching Lee, Chia-Hao Hsu, Peng-Ju Huang, Han Hsiang Huang, and et al. 2021. "Application of microRNA in Human Osteoporosis and Fragility Fracture: A Systemic Review of Literatures" International Journal of Molecular Sciences 22, no. 10: 5232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105232