
MicroRNA Profiles of Maternal and Neonatal Endothelial Progenitor Cells in Preeclampsia

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Demographic Characteristics
2.2. ECFC Isolation and Characterization
2.3. MiRNA Profiles in the Comparison Groups
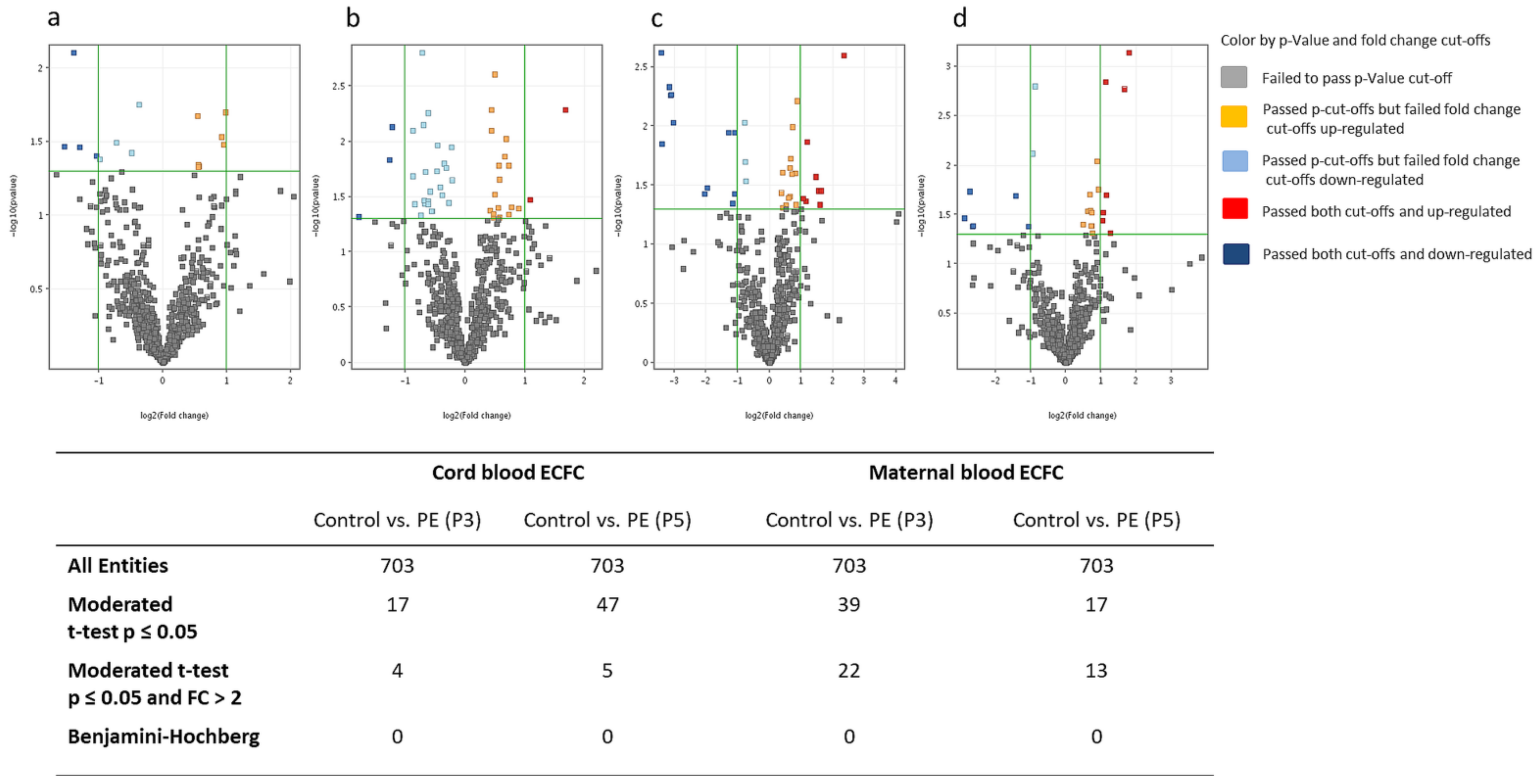
2.3.1. MiRNA Differences between Preeclampsia and Healthy Pregnancies
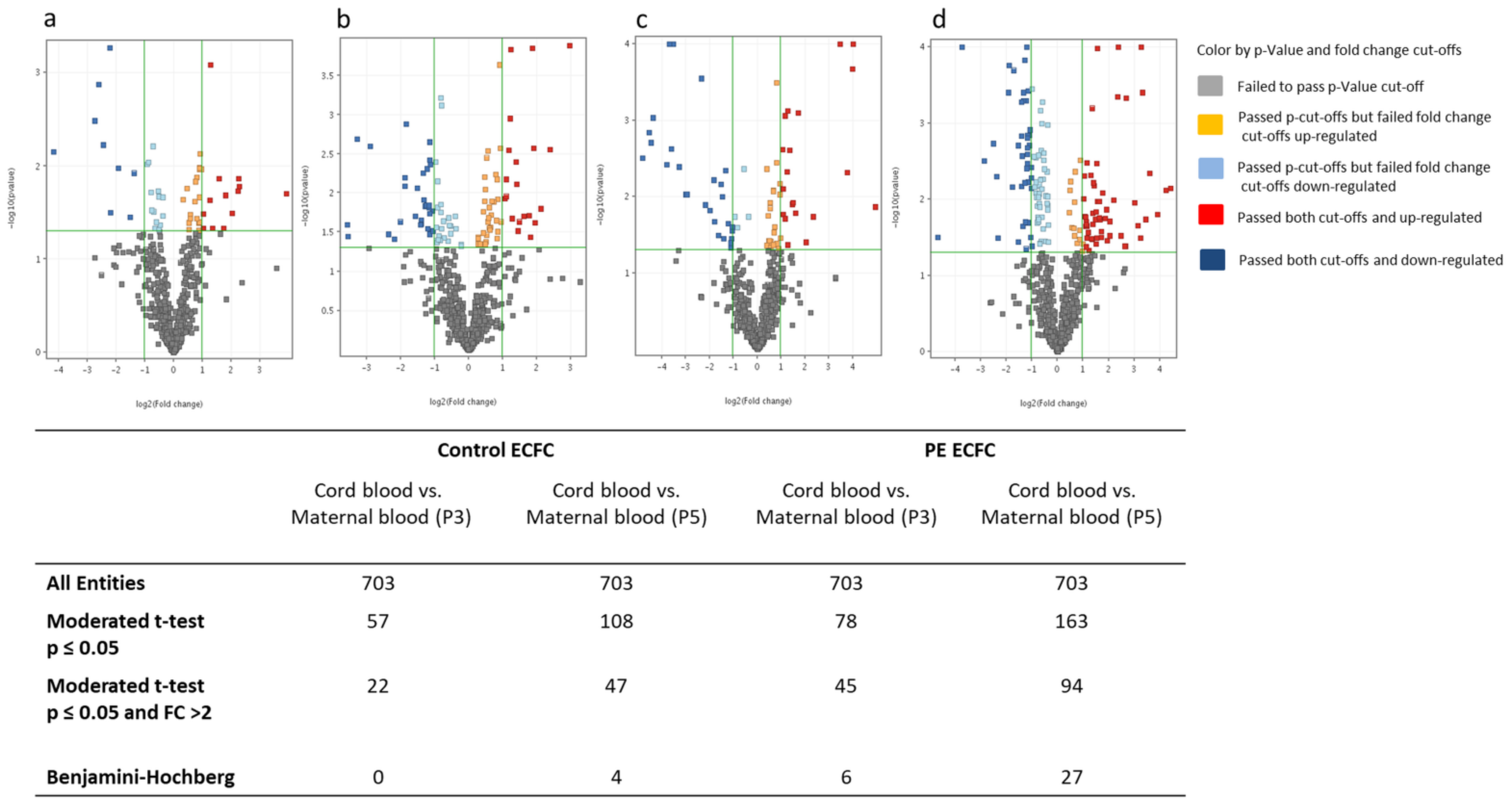
2.3.2. MiRNA Differences between Maternal and Cord Blood
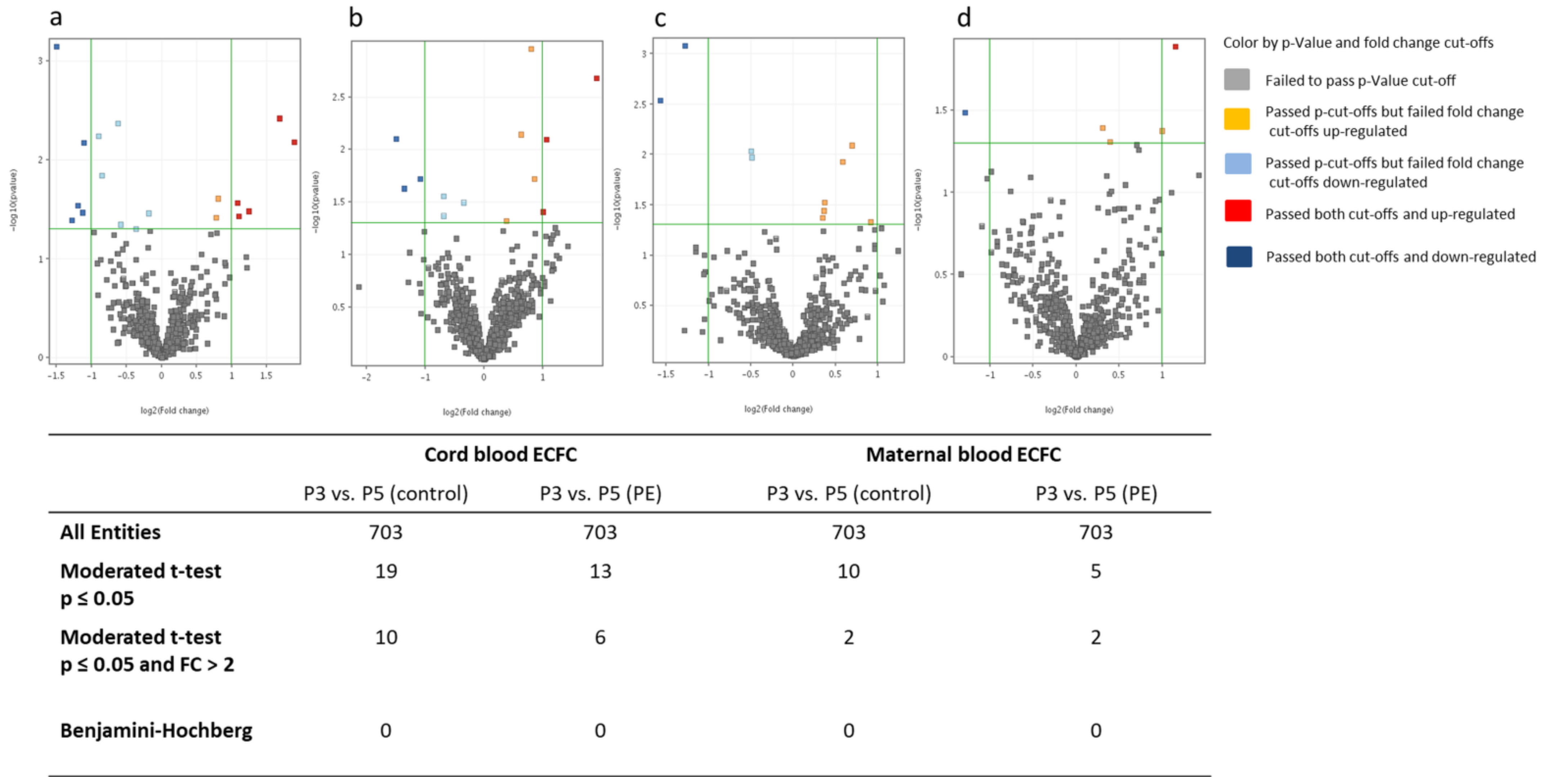
2.3.3. MiRNA Differences between Cell Culture Passages 3 and 5
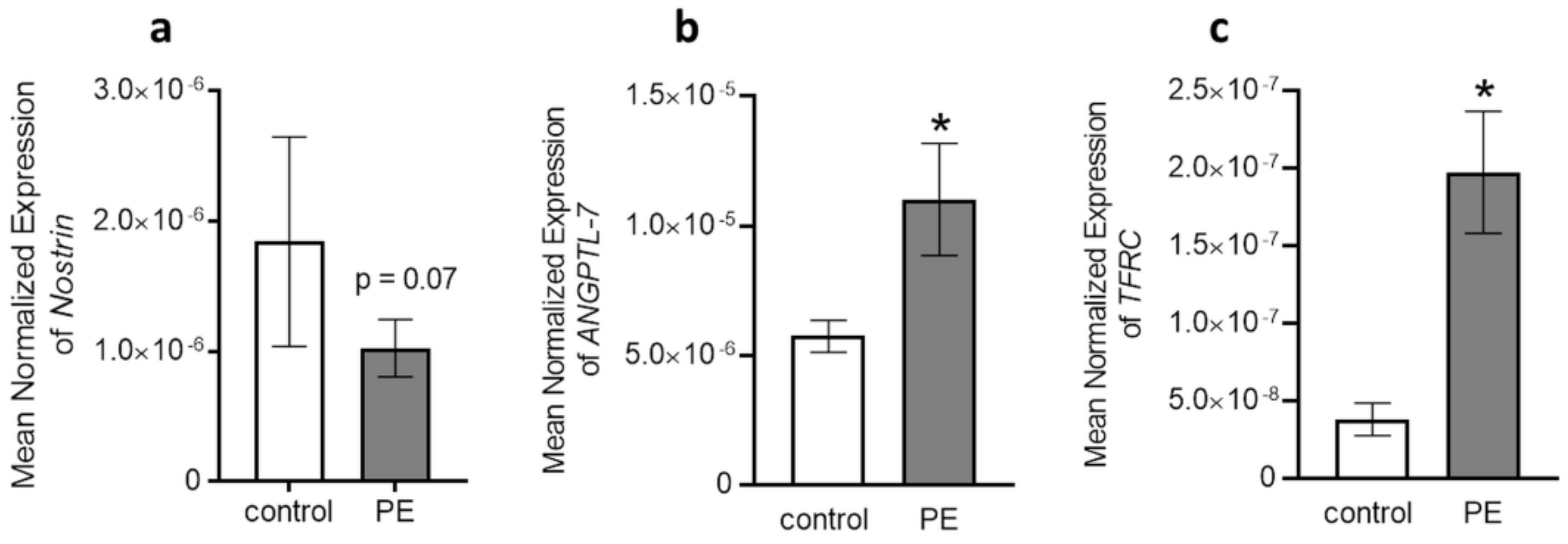
2.4. Quantitative Real-Time PCR Validation and Putative Target Gene Levels
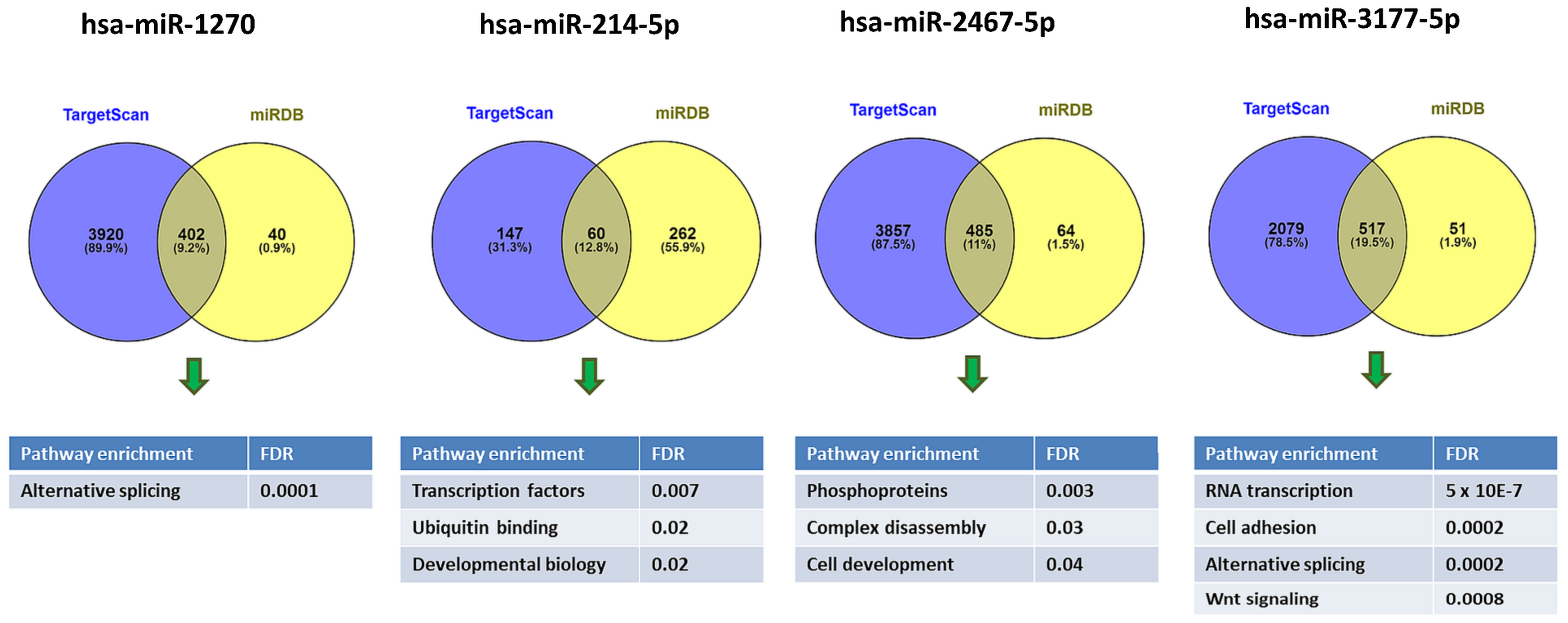
2.5. Pathway Enrichment Analysis
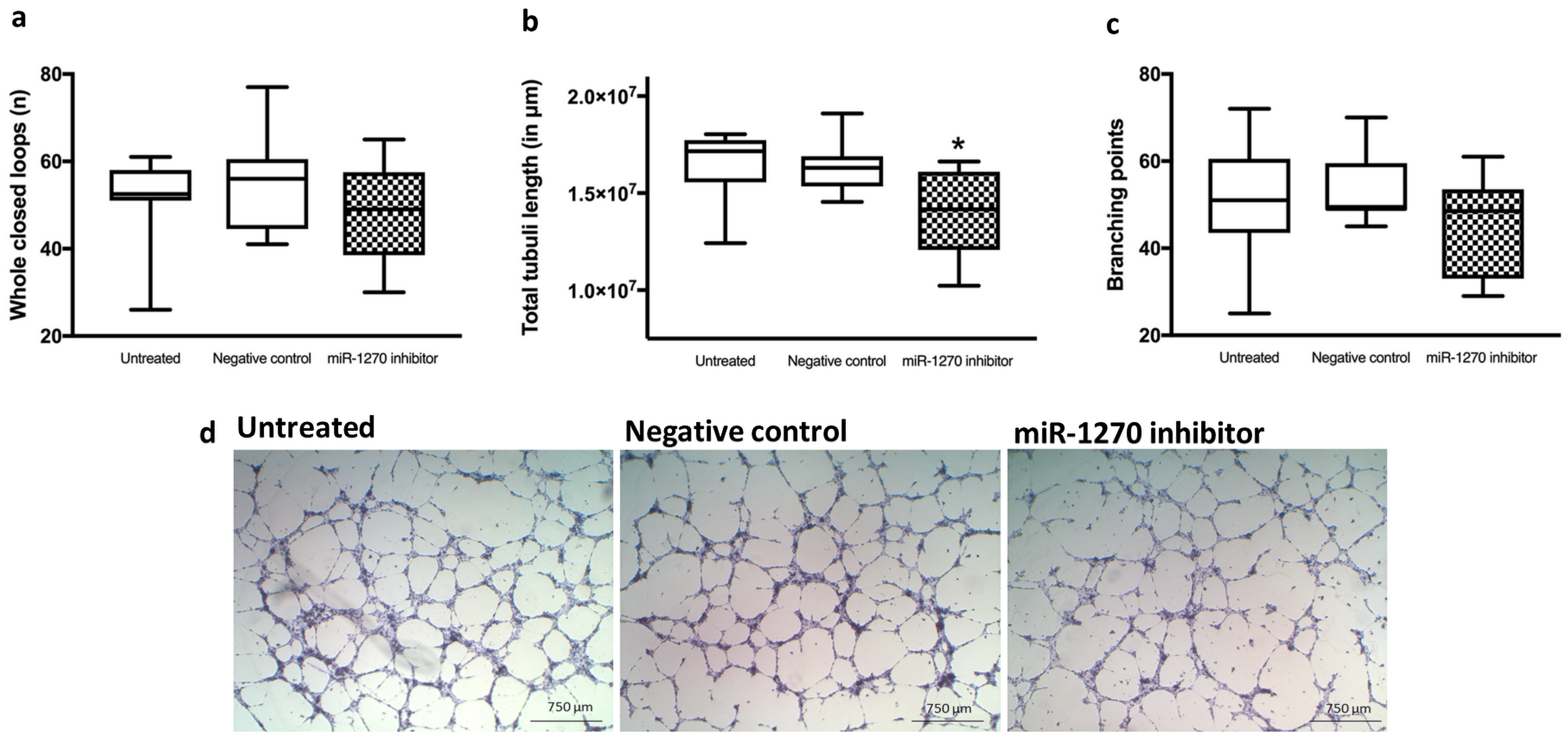
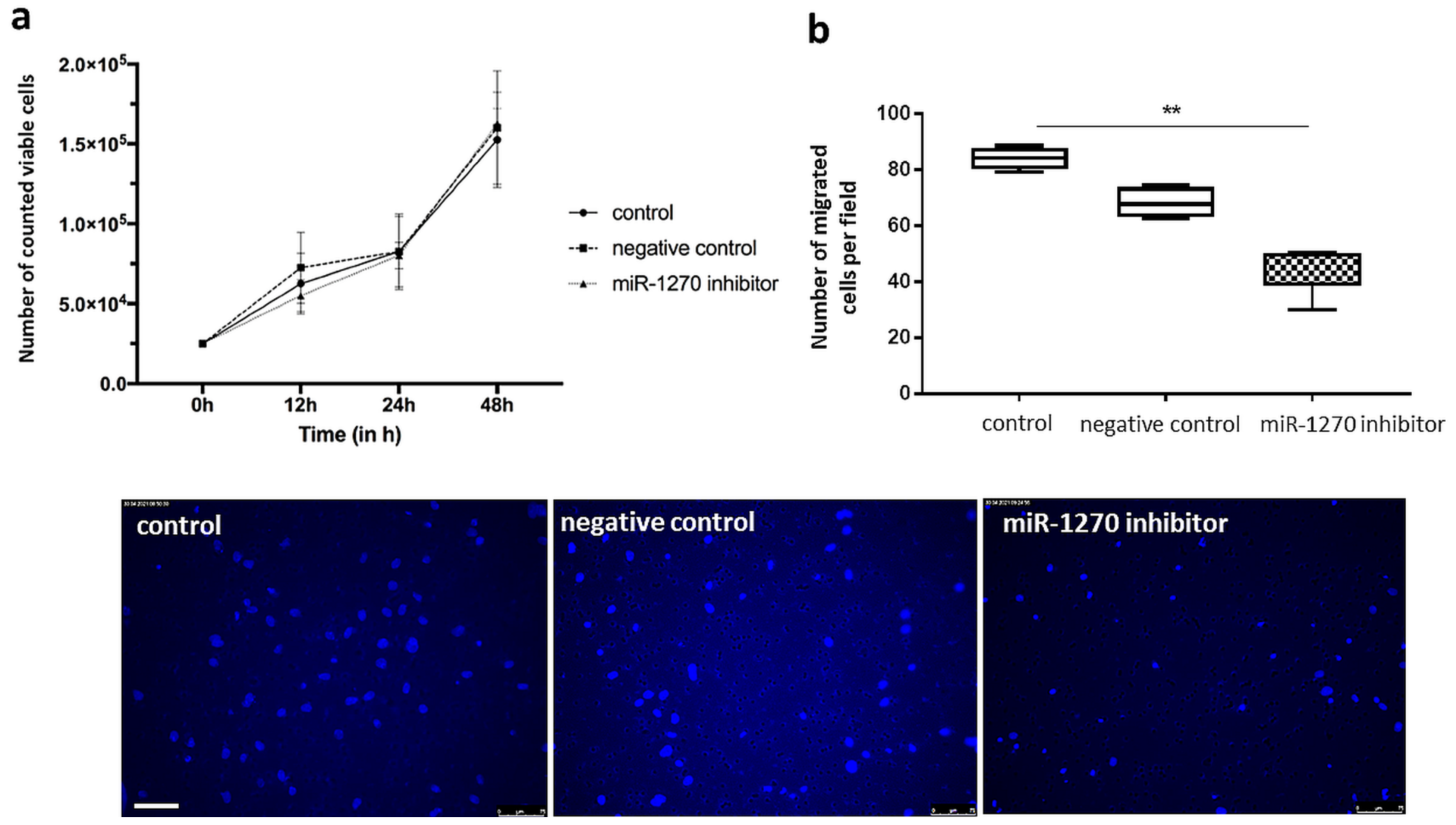
2.6. Hsa-miR-1270 Dependent Tube Formation, Proliferation and Chemotactic Motility
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. ECFC Isolation and Characterization
4.3. Small RNA Sequencing
4.4. qRT-PCR for miRNA Validation and Putative Target Gene Expression
4.5. Transfection of ECFC with miRNA Inhibitor
4.6. Tube Formation Assay
4.7. Cell Proliferation Assay
4.8. Chemotaxis Assay
4.9. Statistics and Bioinformatics Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stojanovska, V.; Scherjon, S.A.; Plosch, T. Preeclampsia as modulator of offspring health. Biol. Reprod. 2016, 94, 53. [Google Scholar] [CrossRef]
- Davis, E.F.; Lazdam, M.; Lewandowski, A.J.; Worton, S.A.; Kelly, B.; Kenworthy, Y.; Adwani, S.; Wilkinson, A.R.; McCormick, K.; Sargent, I.; et al. Cardiovascular risk factors in children and young adults born to preeclamptic pregnancies: A systematic review. Pediatrics 2012, 129, e1552–e1561. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.; Nelson, S.M.; Macdonald-Wallis, C.; Sattar, N.; Lawlor, D.A. Hypertensive disorders of pregnancy and cardiometabolic health in adolescent offspring. Hypertension 2013, 62, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.C.; Best, K.E.; Pearce, M.S.; Waugh, J.; Robson, S.C.; Bell, R. Cardiovascular disease risk in women with pre-eclampsia: Systematic review and meta-analysis. Eur. J. Epidemiol. 2013, 28, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Powers, R.W.; Catov, J.M.; Bodnar, L.M.; Gallaher, M.J.; Lain, K.Y.; Roberts, J.M. Evidence of endothelial dysfunction in preeclampsia and risk of adverse pregnancy outcome. Reprod. Sci. 2008, 15, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Lucke, C.; Rossig, L.; Fichtlscherer, S.; Vasa, M.; Britten, M.; Kamper, U.; Dimmeler, S.; Zeiher, A.M. Reduced number of circulating endothelial progenitor cells predicts future cardiovascular events: Proof of concept for the clinical importance of endogenous vascular repair. Circulation 2005, 111, 2981–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, N.; Kosiol, S.; Schiegl, T.; Ahlers, P.; Walenta, K.; Link, A.; Bohm, M.; Nickenig, G. Circulating endothelial progenitor cells and cardiovascular outcomes. N. Engl. J. Med. 2005, 353, 999–1007. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef]
- King, T.F.; McDermott, J.H. Endothelial progenitor cells and cardiovascular disease. J. Stem Cells 2014, 9, 93–106. [Google Scholar]
- Luppi, P.; Powers, R.W.; Verma, V.; Edmunds, L.; Plymire, D.; Hubel, C.A. Maternal circulating CD34+VEGFR-2+ and CD133+VEGFR-2+ progenitor cells increase during normal pregnancy but are reduced in women with preeclampsia. Reprod. Sci. 2010, 17, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Rajakumar, A.; Plymire, D.A.; Verma, V.; Markovic, N.; Hubel, C.A. Maternal endothelial progenitor colony-forming units with macrophage characteristics are reduced in preeclampsia. Am. J. Hypertens. 2009, 22, 1014–1019. [Google Scholar] [CrossRef]
- Von Versen-Hoynck, F.; Brodowski, L.; Dechend, R.; Myerski, A.C.; Hubel, C.A. Vitamin D antagonizes negative effects of preeclampsia on fetal endothelial colony forming cell number and function. PLoS ONE 2014, 9, e98990. [Google Scholar] [CrossRef]
- Brodowski, L.; von Hardenberg, S.; Schröder-Heurich, B.; von Kaisenberg, C.S.; Hubel, C.A.; Dörk, T.; von Versen-Höynck, F. Preeclampsia-associated alteration of dna methylation in fetal endothelial progenitor cells. Front. Cell. Dev. Biol. 2019, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Rosenfeld, N.; Aharonov, R.; Meiri, E.; Rosenwald, S.; Spector, Y.; Zepeniuk, M.; Benjamin, H.; Shabes, N.; Tabak, S.; Levy, A.; et al. MicroRNAs accurately identify cancer tissue origin. Nat. Biotechnol. 2008, 26, 462–469. [Google Scholar] [CrossRef]
- Santulli, G. microRNAs and endothelial (Dys) function. J. Cell Physiol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Colpaert, R.M.W.; Calore, M. MicroRNAs in cardiac diseases. Cells 2019, 8, 737. [Google Scholar] [CrossRef] [Green Version]
- Mellis, D.; Caporali, A. MicroRNA-based therapeutics in cardiovascular disease: Screening and delivery to the target. Biochem. Soc. Trans. 2018, 46, 11–21. [Google Scholar] [CrossRef]
- Takahashi, Y.; Satoh, M.; Minami, Y.; Tabuchi, T.; Itoh, T.; Nakamura, M. Expression of miR-146a/b is associated with the Toll-like receptor 4 signal in coronary artery disease: Effect of renin-angiotensin system blockade and statins on miRNA-146a/b and Toll-like receptor 4 levels. Clin. Sci. 2010, 119, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Xiang, W.; Chen, H.; Xu, X.; Zhang, M.; Jiang, R. Expression of endothelial nitric oxide synthase traffic inducer in the placentas of women with pre-eclampsia. Int. J. Gynaecol. Obstet. 2005, 89, 103–107. [Google Scholar] [CrossRef]
- Lv, Y.; Lu, C.; Ji, X.; Miao, Z.; Long, W.; Ding, H.; Lv, M. Roles of microRNAs in preeclampsia. J. Cell Physiol. 2019, 234, 1052–1061. [Google Scholar] [CrossRef]
- Apicella, C.; Ruano, C.S.M.; Méhats, C.; Miralles, F.; Vaiman, D. The role of epigenetics in placental development and the etiology of preeclampsia. Int. J. Mol. Sci. 2019, 20, 2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hromadnikova, I.; Kotlabova, K.; Doucha, J.; Dlouha, K.; Krofta, L. Absolute and relative quantification of placenta-specific micrornas in maternal circulation with placental insufficiency-related complications. J. Mol. Diagn. 2012, 14, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Hromadnikova, I.; Kotlabova, K.; Ondrackova, M.; Kestlerova, A.; Novotna, V.; Hympanova, L.; Doucha, J.; Krofta, L. Circulating C19MC microRNAs in preeclampsia, gestational hypertension, and fetal growth restriction. Mediat. Inflamm. 2013, 2013, 186041. [Google Scholar] [CrossRef]
- Gunel, T.; Zeybek, Y.G.; Akcakaya, P.; Kalelioglu, I.; Benian, A.; Ermis, H.; Aydinli, K. Serum microRNA expression in pregnancies with preeclampsia. Genet. Mol. Res. 2011, 10, 4034–4040. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Lu, J.; Wang, S.; Li, H.; Ge, Q.; Lu, Z. Application of next-generation sequencing technology to profile the circulating microRNAs in the serum of preeclampsia versus normal pregnant women. Clin. Chim. Acta Int. J. Clin. Chem. 2011, 412, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, M.; Xue, G.; Zhou, Q.; Jia, Y.; Li, L.; Xin, H.; Sun, S. Elevated levels of hypoxia-inducible microRNA-210 in pre-eclampsia: New insights into molecular mechanisms for the disease. J. Cell. Mol. Med. 2012, 16, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Enquobahrie, D.A.; Abetew, D.F.; Sorensen, T.K.; Willoughby, D.; Chidambaram, K.; Williams, M.A. Placental microRNA expression in pregnancies complicated by preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, 178.e12–178.e21. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhou, X.; Chen, S.; Miao, H.; Fan, H.; Wang, Z.; Hu, Y.; Hou, Y. Differential expression of microRNAs in decidua-derived mesenchymal stem cells from patients with pre-eclampsia. J. Biomed. Sci. 2014, 21, 81. [Google Scholar] [CrossRef] [Green Version]
- Khatun, R.; Wu, Y.; Kanenishi, K.; Ueno, M.; Tanaka, S.; Hata, T.; Sakamoto, H. Immunohistochemical study of transferrin receptor expression in the placenta of pre-eclamptic pregnancy. Placenta 2003, 24, 870–876. [Google Scholar] [CrossRef]
- Yi, T.; Zhou, X.; Sang, K.; Zhou, J.; Ge, L. MicroRNA-1270 modulates papillary thyroid cancer cell development by regulating SCAI. Biomed. Pharmacother. 2019, 109, 2357–2364. [Google Scholar] [CrossRef]
- Wei, L.; Li, P.; Zhao, C.; Wang, N.; Wei, N. Upregulation of microRNA-1270 suppressed human glioblastoma cancer cell proliferation migration and tumorigenesis by acting through WT1. OncoTargets Ther. 2019, 12, 4839–4848. [Google Scholar] [CrossRef] [Green Version]
- Comes, N.; Buie, L.K.; Borrás, T. Evidence for a role of angiopoietin-like 7 (ANGPTL7) in extracellular matrix formation of the human trabecular meshwork: Implications for glaucoma. Genes Cells 2011, 16, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Palmer, K.R.; Tong, S.; Kaitu’u-Lino, T.J. Placental-specific sFLT-1: Role in pre-eclamptic pathophysiology and its translational possibilities for clinical prediction and diagnosis. Mol. Hum. Reprod. 2017, 23, 69–78. [Google Scholar] [CrossRef]
- Ackerman, W.; Salomonis, N.; Zhao, G.; Summerfield, T.; Buhimschi, C.S.; Buhimschi, I.A. 477: Preeclampsia (PE) and global dysregulation of placental alternative RNA splicing events. Am. J. Obstet. Gynecol. 2016, 214, S261–S262. [Google Scholar] [CrossRef]
- Beavers, K.R.; Nelson, C.E.; Duvall, C.L. MiRNA inhibition in tissue engineering and regenerative medicine. Adv. Drug Deliv. Rev. 2015, 88, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Brennan, G.P.; Vitsios, D.M.; Casey, S.; Looney, A.M.; Hallberg, B.; Henshall, D.C.; Boylan, G.B.; Murray, D.M.; Mooney, C. RNA-sequencing analysis of umbilical cord plasma microRNAs from healthy newborns. PLoS ONE 2018, 13, e0207952. [Google Scholar] [CrossRef]
- Khoo, C.P.; Roubelakis, M.G.; Schrader, J.B.; Tsaknakis, G.; Konietzny, R.; Kessler, B.; Harris, A.L.; Watt, S.M. miR-193a-3p interaction with HMGB1 downregulates human endothelial cell proliferation and migration. Sci. Rep. 2017, 7, 44137. [Google Scholar] [CrossRef] [Green Version]
- Van Craenenbroeck, A.H.; Van Craenenbroeck, E.M. Endothelial progenitor cells and cardiovascular risk: Does ageing trump all other factors? Ann. Transl. Med. 2016, 4, 553. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932–936. [Google Scholar] [CrossRef]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Kuehbacher, A.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Role of dicer and drosha for endothelial microRNA expression and angiogenesis. Circ. Res. 2007, 101, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, Y.; Fernandez-Hernando, C.; Pober, J.S.; Sessa, W.C. Dicer dependent microRNAs regulate gene expression and functions in human endothelial cells. Circ. Res. 2007, 100, 1164–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, Y.; Fernandez-Hernando, C.; Yu, J.; Gerber, S.A.; Harrison, K.D.; Pober, J.S.; Iruela-Arispe, M.L.; Merkenschlager, M.; Sessa, W.C. Dicer-dependent endothelial microRNAs are necessary for postnatal angiogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 14082–14087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tranquilli, A.; Dekker, G.; Magee, L.; Roberts, J.; Sibai, S.; Steyn, W.; Zeeman, G.; Brown, M. The classification, diagnosis and management of the hypertensive disorders of pregnancy: A revised statement from the ISSHP. Pregnancy Hypertens. 2014, 4, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, M.; Haidar, M.; Placzko, S.; Niendorf, R.; Darashchonak, N.; Hubel, C.A.; von Versen-Höynck, F. Vitamin D improves the angiogenic properties of endothelial progenitor cells. Am. J. Physiol. Cell Physiol. 2012, 303, C954–C962. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.D.; Balasubramaniam, V.; Mourani, P.M.; Sontag, M.K.; Black, C.P.; Ryan, S.L.; Abman, S.H. Cord blood angiogenic progenitor cells are decreased in bronchopulmonary dysplasia. Eur. Respir. J. 2012, 40, 1516–1522. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef]
- Schröder-Heurich, B.; von Hardenberg, S.; Brodowski, L.; Kipke, B.; Meyer, N.; Borns, K.; von Kaisenberg, C.S.; Brinkmann, H.; Claus, P.; von Versen-Höynck, F. Vitamin D improves endothelial barrier integrity and counteracts inflammatory effects on endothelial progenitor cells. FASEB J. 2019, 33, 9142–9153. [Google Scholar] [CrossRef]
- Liu, K.; Ma, L.; Zhou, F.; Yang, Y.; Hu, H.B.; Wang, L.; Zhong, L. Identification of microRNAs related to myocardial ischemic reperfusion injury. J. Cell. Physiol. 2019, 234, 11380–11390. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. Gene ontology consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cord-Blood-Derived ECFC | Maternal-Blood-Derived ECFC | |||||
|---|---|---|---|---|---|---|
| Healthy Pregnancy (n = 6) | Preeclamptic Pregnancy (n = 6) | p-Value | Healthy Pregnancy (n = 6) | Preeclamptic Pregnancy (n = 6) | p-Value | |
| Maternal age at delivery (years) | 32.8 ± 5.2 | 31.5 ± 3.7 | 0.65 | 31.7 ± 7.4 | 30.8 ± 5.5 | 0.85 |
| Gestational age at delivery (weeks) | 38.6 ± 0.5 | 36.8 ± 1.1 | 0.03 | 38.3 ± 0.7 | 33.1 ± 3.9 | 0.01 |
| Multiparous n (%) | 5 (83%) | 4 (67%) | 1.00 | 6 (100%) | 4 (67%) | 0.45 |
| Maternal pre-pregnancy BMI (kg/m2) | 26.2 ± 6.3 | 29.7 ± 12.9 | 0.80 | 27.2 ± 4.5 | 26.9 ± 6.1 | 0.90 |
| Gestational SBP, pre-delivery (mmHg) | 121 ± 8 | 164 ± 18 | <0.001 | 121 ± 8 | 163 ± 11 | <0.001 |
| Gestational SBP, before 20 week gestation (mmHg) | 112 ± 11 | 125 ± 21 | 0.20 | 116 ± 14 | 113 ± 16 | 0.70 |
| Gestational DBP, pre-delivery (mmHg) | 70 ± 5 | 98 ± 6 | <0.001 | 74 ± 9 | 95 ± 13 | 0.009 |
| Gestational DBP, before 20 week gestation (mmHg) | 69 ± 11 | 80 ± 10 | 0.10 | 68 ± 11 | 76 ± 9 | 0.18 |
| Birth weight (g) | 3487 ± 523 | 2741 ± 669 | 0.06 | 3314 ± 651 | 1628 ± 815 | 0.004 |
| Birth weight percentile | 55 ± 30 | 35 ± 35 | 0.32 | 51 ± 35 | 21 ± 16 | 0.11 |
| Birth weight percentile <10th n (%) | 0 (0%) | 1 (17%) | 1.00 | 1 (17%) | 2 (34%) | 1.00 |
| Caesarean delivery n (%) | 6 (100%) | 5 (83%) | 1.00 | 6 (100%) | 6 (100%) | 1.00 |
| Maternal race, White n (%) | 6 (100%) | 5 (83%) | 1.00 | 6 (100%) | 6 (100%) | 1.00 |
| Baby sex, Male n (%) | 3 (50%) | 1 (17%) | 0.55 | 1 (17%) | 2 (34%) | 1.00 |
| miRNA | Fold Change | p-Value |
|---|---|---|
| ECFC from Cord blood in P3 | ||
| hsa-miR-4726-5p | −2.93 | 0.03 |
| hsa-miR-1270 | −2.63 | 0.008 |
| hsa-miR-148a-5p | −2.48 | 0.03 |
| hsa-miR-1255a | −2.06 | 0.04 |
| ECFC from Cord blood in P5 | ||
| hsa-miR-2467-5p | 3.19 | 0.005 |
| hsa-miR-4687-3p | 2.11 | 0.03 |
| hsa-miR-148a-3p | −3.44 | 0.04 |
| hsa-miR-1226-5p | −2.40 | 0.02 |
| hsa-miR-3911 | −2.34 | 0.007 |
| ECFC from Maternal blood in P3 | ||
| hsa-miR-214-5p | −10.66 | 0.002 |
| hsa-miR-199a-5p | −10.45 | 0.01 |
| hsa-miR-214-3p | −8.89 | 0.005 |
| hsa-miR-199a-3p | −8.52 | 0.005 |
| hsa-miR-199b-3p | −8.52 | 0.005 |
| hsa-miR-139-3p | −8.25 | 0.009 |
| hsa-miR-139-5p | −4.13 | 0.04 |
| hsa-miR-551a | −3.91 | 0.03 |
| hsa-miR-4684-5p | −2.44 | 0.01 |
| hsa-miR-3115 | −2.25 | 0.04 |
| hsa-miR-3164 | −2.16 | 0.04 |
| hsa-miR-199b-5p | −2.14 | 0.01 |
| hsa-miR-4728-3 p | 5.06 | 0.003 |
| hsa-miR-1250 | 3.10 | 0.04 |
| hsa-miR-338-5p | 3.00 | 0.04 |
| hsa-miR-338-3p | 2.92 | 0.04 |
| hsa-miR-4485 | 2.75 | 0.03 |
| hsa-miR-3177-5p | 2.27 | 0.01 |
| hsa-miR-3128 | 2.21 | 0.04 |
| hsa-miR-503 | 2.08 | 0.04 |
| ECFC from Maternal blood in P5 | ||
| hsa-miR-214-5p | −7.34 | 0.03 |
| hsa-miR-214-3p | −6.70 | 0.02 |
| hsa-miR-199a-3p | −6.26 | 0.04 |
| hsa-miR-326 | −2.70 | 0.02 |
| hsa-miR-491-3p | −2.09 | 0.04 |
| hsa-miR-4511 | 3.45 | 0.007 |
| hsa-miR-3128 | 3.18 | 0.002 |
| hsa-miR-4421 | 2.41 | 0.04 |
| hsa-miR-2682-5p | 2.23 | 0.02 |
| hsa-miR-3177-5p | 2.19 | 0.001 |
| hsa-miR-5690 | 2.08 | 0.03 |
| hsa-miR-2277-3p | 2.06 | 0.04 |
| miRNA | Fold Change | p-Value |
|---|---|---|
| Cord blood ECFC from controls in P3 vs. P5 | ||
| hsa-miR-4725-3p | −2.83 | 0.0007 |
| hsa-miR-1293 | −2.44 | 0.04 |
| hsa-miR-3155a | −2.30 | 0.03 |
| hsa-miR-548s | −2.19 | 0.03 |
| hsa-miR-3911 | −2.16 | 0.007 |
| hsa-miR-451a | 3.70 | 0.007 |
| hsa-miR-122-5p | 3.20 | 0.004 |
| hsa-miR-5582-3p | 2.36 | 0.03 |
| hsa-miR-4726-5p | 2.14 | 0.04 |
| hsa-miR-2682-3p | 2.10 | 0.03 |
| Cord blood ECFC from preeclampsia in P3 vs. P5 | ||
| hsa-miR-1226-5p | −28.23 | 0.008 |
| hsa-miR-5001-5p | −25.60 | 0.02 |
| hsa-miR-3911 | −21.26 | 0.02 |
| hsa-miR-3149 | 20.86 | 0.008 |
| hsa-miR-4791 | 20.16 | 0.04 |
| hsa-miR-2467-5p | 3.77 | 0.002 |
| Maternal blood ECFC from controls in P3 vs. P5 | ||
| hsa-let-7g-3p | −24.30 | 0.008 |
| hsa-miR-4421 | −2.97 | 0.003 |
| Maternal blood ECFC from preeclampsia in P3 vs. P5 | ||
| hsa-miR-4677-5p | −2.45 | 0.03 |
| hsa-miR-4754 | 2.22 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brodowski, L.; Schröder-Heurich, B.; von Hardenberg, S.; Richter, K.; von Kaisenberg, C.S.; Dittrich-Breiholz, O.; Meyer, N.; Dörk, T.; von Versen-Höynck, F. MicroRNA Profiles of Maternal and Neonatal Endothelial Progenitor Cells in Preeclampsia. Int. J. Mol. Sci. 2021, 22, 5320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105320
Brodowski L, Schröder-Heurich B, von Hardenberg S, Richter K, von Kaisenberg CS, Dittrich-Breiholz O, Meyer N, Dörk T, von Versen-Höynck F. MicroRNA Profiles of Maternal and Neonatal Endothelial Progenitor Cells in Preeclampsia. International Journal of Molecular Sciences. 2021; 22(10):5320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105320
Chicago/Turabian StyleBrodowski, Lars, Bianca Schröder-Heurich, Sandra von Hardenberg, Katja Richter, Constantin S. von Kaisenberg, Oliver Dittrich-Breiholz, Nadia Meyer, Thilo Dörk, and Frauke von Versen-Höynck. 2021. "MicroRNA Profiles of Maternal and Neonatal Endothelial Progenitor Cells in Preeclampsia" International Journal of Molecular Sciences 22, no. 10: 5320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105320