
Increased Expression of 11β-Hydroxysteroid Dehydrogenase Type 1 Contributes to Epidermal Permeability Barrier Dysfunction in Aged Skin

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Stratum Corneum (SC) and Oral Epithelium Cortisol Levels Are Higher in the Aged Participant
2.2. Increased 11β-HSD1 Expression in the Skin of Aged Hairless Mice
2.3. Deterioration of Skin Barrier Function in Aged Hairless Mice
2.4. Increased Serum Inflammatory Cytokine Levels in Aged Hairless Mice
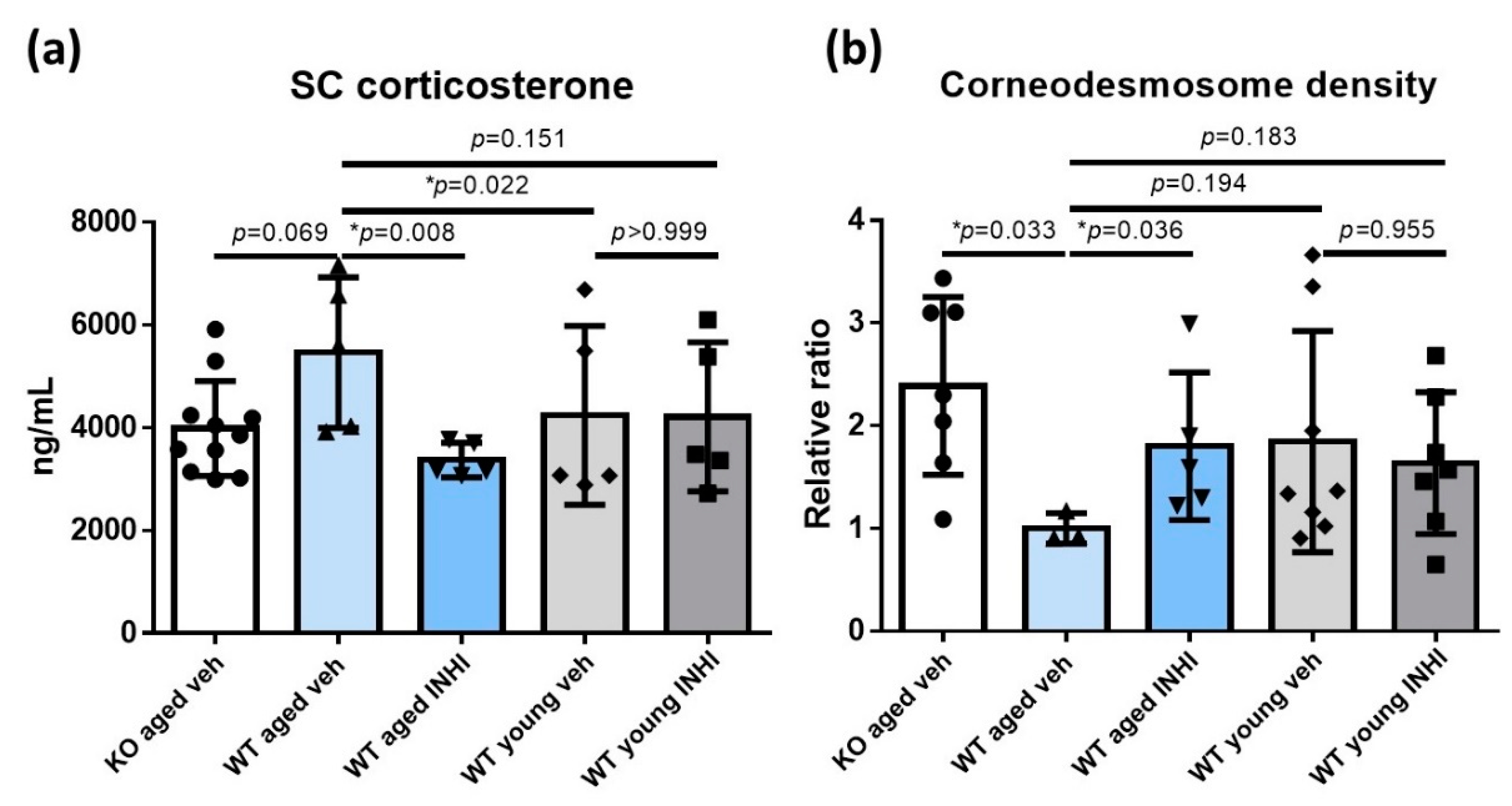
2.5. Reduced SC Corticosterone Concentration in Aged 11β-HSD1 (HSD11B1) Knockout (KO) Mice and Aged 11β-HSD1 Inhibitor (INHI)-Treated Wild-Type (WT) Mice
2.6. Decreased Corneodesmosome (CD) Density in WT Aged Mice Compared to That in Aged HSD11B1 KO Mice and Aged INHI-Treated WT Mice
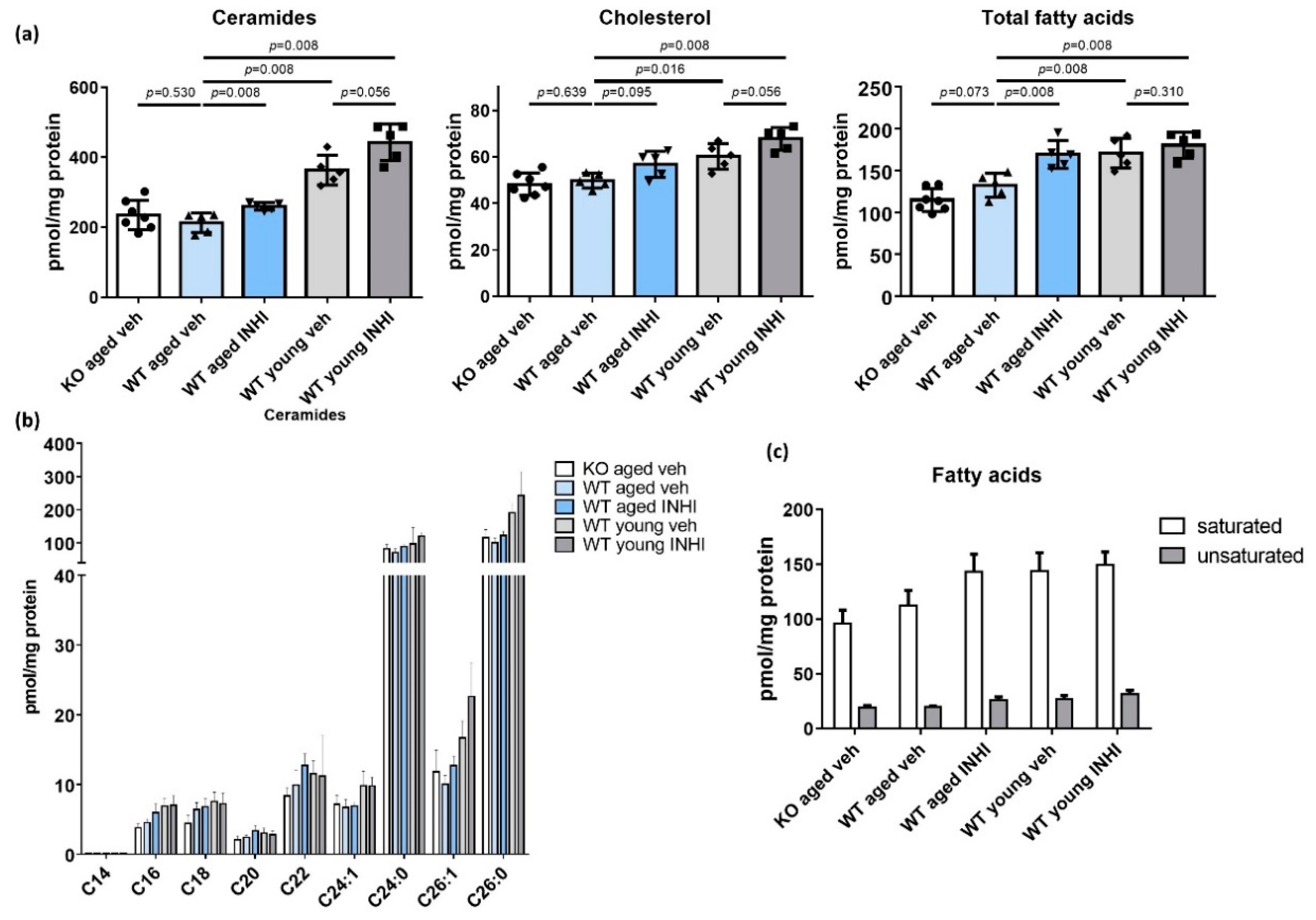
2.7. Recovery of the SC Lipid Levels in WT Aged Mice upon the Topical INHI Treatment
3. Discussion
4. Materials and Methods
4.1. The Human Experiment
4.2. The Animal Experiment
4.3. Preparation of 11β-HSD1 KO Mice
4.4. Quantification of Cortisol and Corticosterone by ELISA
4.5. Measurement of Skin Barrier Function
4.6. IHC Staining and Semi-Quantitative Analysis of 11β-HSD1
4.7. Serum Cytokine Level Assays
4.8. Quantitative EM
4.9. Lipid Analysis in the SC
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cerimele, D.; Celleno, L.; Serri, F. Physiological changes in ageing skin. Br. J. Dermatol. 1990, 122 (Suppl. 35), 13–20. [Google Scholar] [CrossRef]
- Tiganescu, A.; Tahrani, A.A.; Morgan, S.A.; Otranto, M.; Desmouliere, A.; Abrahams, L.; Hassan-Smith, Z.; Walker, E.A.; Rabbitt, E.H.; Cooper, M.S.; et al. 11beta-hydroxysteroid dehydrogenase blockade prevents age-induced skin structure and function defects. J. Clin. Investig. 2013, 123, 3051–3060. [Google Scholar] [CrossRef]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Keyes, B.E.; Liu, S.; Asare, A.; Naik, S.; Levorse, J.; Polak, L.; Lu, C.P.; Nikolova, M.; Pasolli, H.A.; Fuchs, E. Impaired epidermal to dendritic t cell signaling slows wound repair in aged skin. Cell 2016, 167, 1323–1338.e1314. [Google Scholar] [CrossRef] [Green Version]
- Ghadially, R.; Brown, B.E.; Hanley, K.; Reed, J.T.; Feingold, K.R.; Elias, P.M. Decreased epidermal lipid synthesis accounts for altered barrier function in aged mice. J. Investig. Dermatol. 1996, 106, 1064–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelova-Fischer, I.; Fischer, T.W.; Abels, C.; Zillikens, D. Accelerated barrier recovery and enhancement of the barrier integrity and properties by topical application of a ph 4 vs. A ph 5.8 water-in-oil emulsion in aged skin. Br. J. Dermatol. 2018, 179, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Biniek, K.; Kaczvinsky, J.; Matts, P.; Dauskardt, R.H. Understanding age-induced alterations to the biomechanical barrier function of human stratum corneum. J. Dermatol. Sci. 2015, 80, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.S.; Fluhr, J.W.; Man, M.Q.; Fowler, A.J.; Hachem, J.P.; Crumrine, D.; Ahn, S.K.; Brown, B.E.; Elias, P.M.; Feingold, K.R. Short-term glucocorticoid treatment compromises both permeability barrier homeostasis and stratum corneum integrity: Inhibition of epidermal lipid synthesis accounts for functional abnormalities. J. Investig. Dermatol. 2003, 120, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denda, M.; Tsuchiya, T.; Elias, P.M.; Feingold, K.R. Stress alters cutaneous permeability barrier homeostasis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R367–R372. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Brown, B.E.; Crumrine, D.; Chang, S.; Man, M.Q.; Elias, P.M.; Feingold, K.R. Mechanisms by which psychologic stress alters cutaneous permeability barrier homeostasis and stratum corneum integrity. J. Investig. Dermatol. 2005, 124, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Jozic, I.; Stojadinovic, O.; Kirsner, R.S.; Tomic-Canic, M. Stressing the steroids in skin: Paradox or fine-tuning? J. Investig. Dermatol. 2014, 134, 2869–2872. [Google Scholar] [CrossRef] [Green Version]
- Tampa, M.; Sarbu, M.I.; Mitran, M.I.; Mitran, C.I.; Matei, C.; Georgescu, S.R. The pathophysiological mechanisms and the quest for biomarkers in psoriasis, a stress-related skin disease. Dis. Markers 2018, 2018, 5823684. [Google Scholar] [CrossRef] [Green Version]
- Tiganescu, A.; Walker, E.A.; Hardy, R.S.; Mayes, A.E.; Stewart, P.M. Localization, age- and site-dependent expression, and regulation of 11beta-hydroxysteroid dehydrogenase type 1 in skin. J. Investig. Dermatol. 2011, 131, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, M.; Katayama, I. Local cortisol/corticosterone activation in skin physiology and pathology. J. Dermatol. Sci. 2016, 84, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Kinn, P.M.; Holdren, G.O.; Westermeyer, B.A.; Abuissa, M.; Fischer, C.L.; Fairley, J.A.; Brogden, K.A.; Brogden, N.K. Age-dependent variation in cytokines, chemokines, and biologic analytes rinsed from the surface of healthy human skin. Sci. Rep. 2015, 5, 10472. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Tuckey, R.C.; Paus, R. Differential expression of hpa axis homolog in the skin. Mol. Cell Endocrinol. 2007, 265–266, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine aspects of skin aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jozic, I.; Stojadinovic, O.; Kirsner, R.S.F.; Tomic-Canic, M. Skin under the (spot)-light: Cross-talk with the central hypothalamic-pituitary-adrenal (hpa) axis. J. Investig. Dermatol. 2015, 135, 1469–1471. [Google Scholar] [CrossRef] [Green Version]
- Wierzbicka, J.M.; Zmijewski, M.A.; Antoniewicz, J.; Sobjanek, M.; Slominski, A.T. Differentiation of keratinocytes modulates skin hpa analog. J. Cell Physiol. 2017, 232, 154–166. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Zbytek, B.; Tobin, D.J.; Theoharides, T.C.; Rivier, J. Key role of crf in the skin stress response system. Endocr. Rev. 2013, 34, 827–884. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zjawiony, J.; Wortsman, J.; Semak, I.; Stewart, J.; Pisarchik, A.; Sweatman, T.; Marcos, J.; Dunbar, C.; Turkey, R.C. A novel pathway for sequential transformation of 7-dehydrocholesterol and expression of the p450scc system in mammalian skin. Eur. J. Biochem. 2004, 271, 4178–4188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, R.M.; Tuckey, R.C.; Manna, P.R.; Jetten, A.M.; Postlethwaite, A.; Raman, C.; Slominski, A.T. Extra-adrenal glucocorticoid biosynthesis: Implications for autoimmune and inflammatory disorders. Genes Immun. 2020, 21, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.W.; Walker, E.A.; Bujalska, I.J.; Draper, N.; Lavery, G.G.; Cooper, M.S.; Hewison, M.; Stewart, P.M. 11beta-hydroxysteroid dehydrogenase type 1: A tissue-specific regulator of glucocorticoid response. Endocr. Rev. 2004, 25, 831–866. [Google Scholar] [CrossRef]
- Loerz, C.; Maser, E. The cortisol-activating enzyme 11beta-hydroxysteroid dehydrogenase type 1 in skeletal muscle in the pathogenesis of the metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2017, 174, 65–71. [Google Scholar] [CrossRef]
- Dammann, C.; Stapelfeld, C.; Maser, E. Expression and activity of the cortisol-activating enzyme 11beta-hydroxysteroid dehydrogenase type 1 is tissue and species-specific. Chem. Biol. Interact. 2019, 303, 57–61. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Yang, Q.; Shao, S. 11beta-hydroxysteroid dehydrogenase type 1 in obese subjects with type 2 diabetes mellitus. Am. J. Med Sci. 2017, 354, 408–414. [Google Scholar] [CrossRef]
- Tiganescu, A.; Hupe, M.; Jiang, Y.J.; Celli, A.; Uchida, Y.; Mauro, T.M.; Bikle, D.D.; Elias, P.M.; Holleran, W.M. Uvb induces epidermal 11beta-hydroxysteroid dehydrogenase type 1 activity in vivo. Exp. Dermatol. 2015, 24, 370–376. [Google Scholar] [CrossRef]
- Terao, M.; Murota, H.; Kimura, A.; Kato, A.; Ishikawa, A.; Igawa, K.; Miyoshi, E.; Katayama, I. 11beta-hydroxysteroid dehydrogenase-1 is a novel regulator of skin homeostasis and a candidate target for promoting tissue repair. PLoS ONE 2011, 6, e25039. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Murota, H.; Terao, M.; Katayama, I. Attenuated activation of homeostatic glucocorticoid in keratinocytes induces alloknesis via aberrant artemin production. J. Investig. Dermatol. 2018, 138, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Boudon, S.; Heidl, M.; Vuorinen, A.; Wandeler, E.; Campiche, R.; Odermatt, A.; Jackson, E. Design, synthesis, and biological evaluation of novel selective peptide inhibitors of 11beta-hydroxysteroid dehydrogenase 1. Bioorg. Med. Chem. 2018, 26, 5128–5139. [Google Scholar] [CrossRef]
- Tiganescu, A.; Hupe, M.; Uchida, Y.; Mauro, T.; Elias, P.M.; Holleran, W.M. Increased glucocorticoid activation during mouse skin wound healing. J. Endocrinol. 2014, 221, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Williamson, S.; Merritt, J.; De Benedetto, A. Atopic dermatitis in the elderly: A review of clinical and pathophysiological hallmarks. Br. J. Dermatol. 2020, 182, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Norman, R.A. Xerosis and pruritus in the elderly: Recognition and management. Dermatol. Ther. 2003, 16, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Jafferany, M.; Huynh, T.V.; Silverman, M.A.; Zaidi, Z. Geriatric dermatoses: A clinical review of skin diseases in an aging population. Int. J. Dermatol. 2012, 51, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.R.; Kim, B.J.; Lee, C.H.; Lee, Y.B.; Lee, S.; Hwang, H.J.; Kim, E.; Kim, S.H.; Lee, M.G.; Lee, S.E.; et al. Role of 11beta-hydroxysteroid dehydrogenase type 1 in the development of atopic dermatitis. Sci. Rep. 2020, 10, 20237. [Google Scholar] [CrossRef] [PubMed]
- Boudon, S.M.; Vuorinen, A.; Geotti-Bianchini, P.; Wandeler, E.; Kratschmar, D.V.; Heidl, M.; Campiche, R.; Jackson, E.; Odermatt, A. Novel 11beta-hydroxysteroid dehydrogenase 1 inhibitors reduce cortisol levels in keratinocytes and improve dermal collagen content in human ex vivo skin after exposure to cortisone and uv. PLoS ONE 2017, 12, e0171079. [Google Scholar] [CrossRef] [Green Version]
- Tiganescu, A.; Hupe, M.; Uchida, Y.; Mauro, T.; Elias, P.M.; Holleran, W.M. Topical 11beta-hydroxysteroid dehydrogenase type 1 inhibition corrects cutaneous features of systemic glucocorticoid excess in female mice. Endocrinology 2018, 159, 547–556. [Google Scholar] [CrossRef]
- Terao, M.; Tani, M.; Itoi, S.; Yoshimura, T.; Hamasaki, T.; Murota, H.; Katayama, I. 11beta-hydroxysteroid dehydrogenase 1 specific inhibitor increased dermal collagen content and promotes fibroblast proliferation. PLoS ONE 2014, 9, e93051. [Google Scholar] [CrossRef] [Green Version]
- Sheu, H.M.; Tai, C.L.; Kuo, K.W.; Yu, H.S.; Chai, C.Y. Modulation of epidermal terminal differentiation in patients after long-term topical corticosteroids. J. Dermatol. 1991, 18, 454–464. [Google Scholar] [CrossRef]
- Sheu, H.M.; Lee, J.Y.; Kuo, K.W.; Tsai, J.C. Permeability barrier abnormality of hairless mouse epidermis after topical corticosteroid: Characterization of stratum corneum lipids by ruthenium tetroxide staining and high-performance thin-layer chromatography. J. Dermatol. 1998, 25, 281–289. [Google Scholar] [CrossRef]
- Ali, S.M.; Yosipovitch, G. Skin ph: From basic science to basic skin care. Acta Derm. Venereol. 2013, 93, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, G.; Mauro, T.M.; Kim, P.L.; Hupe, M.; Zhai, Y.; Sun, R.; Crumrine, D.; Cheung, C.; Nuno-Gonzalez, A.; Elias, P.M.; et al. Topical hesperidin prevents glucocorticoid-induced abnormalities in epidermal barrier function in murine skin. Exp. Dermatol. 2014, 23, 645–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, S.J.; Kim, D.; Kim, E.J.; Ahn, J.S.; Choi, E.J.; Son, E.D.; Lee, T.R.; Choi, E.H. Psychological stress deteriorates skin barrier function by activating 11beta-hydroxysteroid dehydrogenase 1 and the hpa axis. Sci. Rep. 2018, 8, 6334. [Google Scholar] [CrossRef]
- Choi, E.H. Aging of the skin barrier. Clin. Dermatol. 2019, 37, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Man, M.Q.; Xu, P.; Xin, S.; Liu, Z.; Crumrine, D.A.; Jiang, Y.J.; Fluhr, J.W.; Feingold, K.R.; Elias, P.M.; et al. Stratum corneum acidification is impaired in moderately aged human and murine skin. J. Investig. Dermatol. 2007, 127, 2847–2856. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Mauro, T.M.; Dang, E.; Man, G.; Zhang, J.; Lee, D.; Wang, G.; Feingold, K.R.; Elias, P.M.; Man, M.Q. Epidermal dysfunction leads to an age-associated increase in levels of serum inflammatory cytokines. J. Investig. Dermatol. 2017, 137, 1277–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, Y.; Ogo, M.; Denda, M. Epidermal interleukin-1 alpha generation is amplified at low humidity: Implications for the pathogenesis of inflammatory dermatoses. Br. J. Dermatol. 2001, 144, 238–243. [Google Scholar] [CrossRef]
- Denda, M.; Sato, J.; Tsuchiya, T.; Elias, P.M.; Feingold, K.R. Low humidity stimulates epidermal DNA synthesis and amplifies the hyperproliferative response to barrier disruption: Implication for seasonal exacerbations of inflammatory dermatoses. J. Investig. Dermatol. 1998, 111, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Gan, Y.; He, C.; Chen, Z.; Zhou, C. The mechanism of skin lipids influencing skin status. J. Dermatol. Sci. 2018, 89, 112–119. [Google Scholar] [CrossRef]
- van Smeden, J.; Bouwstra, J.A. Stratum corneum lipids: Their role for the skin barrier function in healthy subjects and atopic dermatitis patients. Curr. Probl. Dermatol. 2016, 49, 8–26. [Google Scholar] [CrossRef]
- Zettersten, E.M.; Ghadially, R.; Feingold, K.R.; Crumrine, D.; Elias, P.M. Optimal ratios of topical stratum corneum lipids improve barrier recovery in chronologically aged skin. J. Am. Acad. Dermatol. 1997, 37, 403–408. [Google Scholar] [CrossRef]
- Ghadially, R.; Brown, B.E.; Sequeira-Martin, S.M.; Feingold, K.R.; Elias, P.M. The aged epidermal permeability barrier. Structural, functional, and lipid biochemical abnormalities in humans and a senescent murine model. J. Clin. Investig. 1995, 95, 2281–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Skobowiat, C.; Dowdy, J.C.; Sayre, R.M.; Tuckey, R.C.; Slominski, A. Cutaneous hypothalamic-pituitary-adrenal axis homolog: Regulation by ultraviolet radiation. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E484–E493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skobowiat, C.; Sayre, R.M.; Dowdy, J.C.; Slominski, A.T. Ultraviolet radiation regulates cortisol activity in a waveband-dependent manner in human skin ex vivo. Br. J. Dermatol. 2013, 168, 595–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How uv light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [Green Version]
- Skobowiat, C.; Postlethwaite, A.E.; Slominski, A.T. Skin exposure to ultraviolet b rapidly activates systemic neuroendocrine and immunosuppressive responses. Photochem. Photobiol. 2017, 93, 1008–1015. [Google Scholar] [CrossRef]
- Skobowiat, C.; Slominski, A.T. Uvb activates hypothalamic-pituitary-adrenal axis in c57bl/6 mice. J. Investig. Dermatol. 2015, 135, 1638–1648. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.S.; Monson, J.P.; Kaltsas, G.; Putignano, P.; Wood, P.J.; Sheppard, M.C.; Besser, G.M.; Taylor, N.F.; Stewart, P.M. Modulation of 11beta-hydroxysteroid dehydrogenase isozymes by growth hormone and insulin-like growth factor: In vivo and in vitro studies. J. Clin. Endocrinol. Metab. 1999, 84, 4172–4177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reschke-Hernandez, A.E.; Okerstrom, K.L.; Bowles Edwards, A.; Tranel, D. Sex and stress: Men and women show different cortisol responses to psychological stress induced by the trier social stress test and the iowa singing social stress test. J. Neurosci. Res. 2017, 95, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelfsema, F.; van Heemst, D.; Iranmanesh, A.; Takahashi, P.; Yang, R.; Veldhuis, J.D. Impact of age, sex and body mass index on cortisol secretion in 143 healthy adults. Endocr. Connect. 2017, 6, 500–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semjonous, N.M.; Sherlock, M.; Jeyasuria, P.; Parker, K.L.; Walker, E.A.; Stewart, P.M.; Lavery, G.G. Hexose-6-phosphate dehydrogenase contributes to skeletal muscle homeostasis independent of 11beta-hydroxysteroid dehydrogenase type 1. Endocrinology 2011, 152, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Oh, Y.; Jung, M.; Lee, S.; Jeon, H.; Cho, M.Y.; Lee, S.H.; Choi, E.H. Topical calcitriol restores the impairment of epidermal permeability and antimicrobial barriers induced by corticosteroids. Br. J. Dermatol. 2010, 162, 1251–1260. [Google Scholar] [CrossRef]
- Lee, H.J.; Yoon, N.Y.; Lee, N.R.; Jung, M.; Kim, D.H.; Choi, E.H. Topical acidic cream prevents the development of atopic dermatitis- and asthma-like lesions in murine model. Exp. Dermatol. 2014, 23, 736–741. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, N.R.; Kim, B.K.; Jung, M.; Kim, D.H.; Moniaga, C.S.; Kabashima, K.; Choi, E.H. Acidification of stratum corneum prevents the progression from atopic dermatitis to respiratory allergy. Exp. Dermatol. 2017, 26, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Menon, G.K.; Feingold, K.R.; Elias, P.M. Lamellar body secretory response to barrier disruption. J. Investig. Dermatol. 1992, 98, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.H.; Demerjian, M.; Crumrine, D.; Brown, B.E.; Mauro, T.; Elias, P.M.; Feingold, K.R. Glucocorticoid blockade reverses psychological stress-induced abnormalities in epidermal structure and function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1657–R1662. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Ikushiro, H.; Seo, H.S.; Shin, K.O.; Kim, Y.I.; Kim, J.Y.; Lee, Y.M.; Yano, T.; Holleran, W.M.; Elias, P.; et al. Er stress stimulates production of the key antimicrobial peptide, cathelicidin, by forming a previously unidentified intracellular s1p signaling complex. Proc. Natl. Acad. Sci. USA 2016, 113, E1334–E1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.O.; Lim, C.J.; Park, H.Y.; Kim, S.; Kim, B.; Lee, Y.; Chung, H.; Jeong, S.K.; Park, K.; Park, K. Activation of sirt1 enhances epidermal permeability barrier formation through ceramide synthase 2- and 3-dependent mechanisms. J. Investig. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, A.; Chitarrini, G.; Di Gangi, I.M.; Masuero, D.; Soini, E.; Mattivi, F.; Vrhovsek, U. A rapid lc-ms/ms method for quantitative profiling of fatty acids, sterols, glycerolipids, glycerophospholipids and sphingolipids in grapes. Talanta 2015, 140, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Perez-Navarro, J.; Da Ros, A.; Masuero, D.; Izquierdo-Canas, P.M.; Hermosin-Gutierrez, I.; Gomez-Alonso, S.; Mattivi, F.; Vrhovsek, U. Lc-ms/ms analysis of free fatty acid composition and other lipids in skins and seeds of vitis vinifera grape cultivars. Food Res. Int. 2019, 125, 108556. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Siegmund, W.; Oswald, S. Rapid lc-ms/ms method for the determination of 4-hydroxycholesterol/cholesterol ratio in serum as endogenous biomarker for cyp3a activity in human and foals. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1033–1034, 193–199. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.J.; Lee, N.R.; Lee, C.H.; Lee, Y.B.; Choe, S.J.; Lee, S.; Hwang, H.J.; Kim, E.; Lavery, G.G.; Shin, K.-O.; et al. Increased Expression of 11β-Hydroxysteroid Dehydrogenase Type 1 Contributes to Epidermal Permeability Barrier Dysfunction in Aged Skin. Int. J. Mol. Sci. 2021, 22, 5750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115750
Kim BJ, Lee NR, Lee CH, Lee YB, Choe SJ, Lee S, Hwang HJ, Kim E, Lavery GG, Shin K-O, et al. Increased Expression of 11β-Hydroxysteroid Dehydrogenase Type 1 Contributes to Epidermal Permeability Barrier Dysfunction in Aged Skin. International Journal of Molecular Sciences. 2021; 22(11):5750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115750
Chicago/Turabian StyleKim, Beom Jun, Noo Ri Lee, Chung Hyeok Lee, Young Bin Lee, Sung Jay Choe, Solam Lee, Hyun Jee Hwang, Eunjung Kim, Gareth G. Lavery, Kyong-Oh Shin, and et al. 2021. "Increased Expression of 11β-Hydroxysteroid Dehydrogenase Type 1 Contributes to Epidermal Permeability Barrier Dysfunction in Aged Skin" International Journal of Molecular Sciences 22, no. 11: 5750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115750