
Detecting Variants in the NBN Gene While Testing for Hereditary Breast Cancer: What to Do Next?

 ,
,
Abstract
:1. Introduction
2. Results
2.1. NBN Sequence Variants
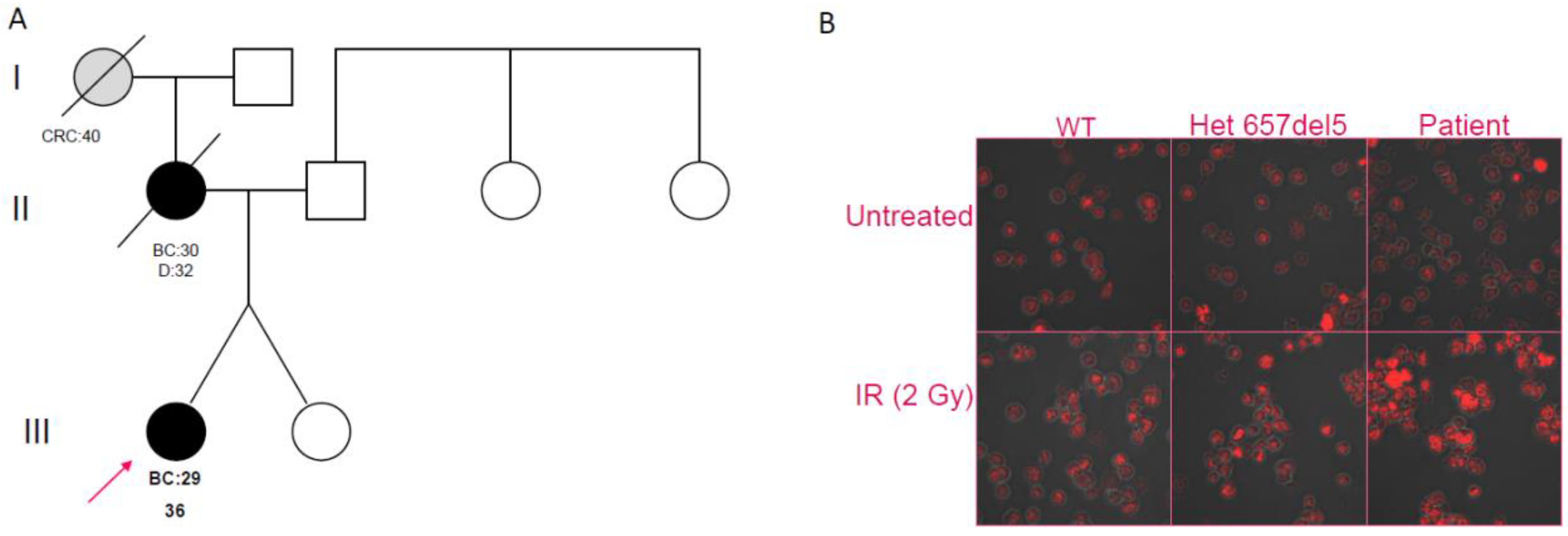
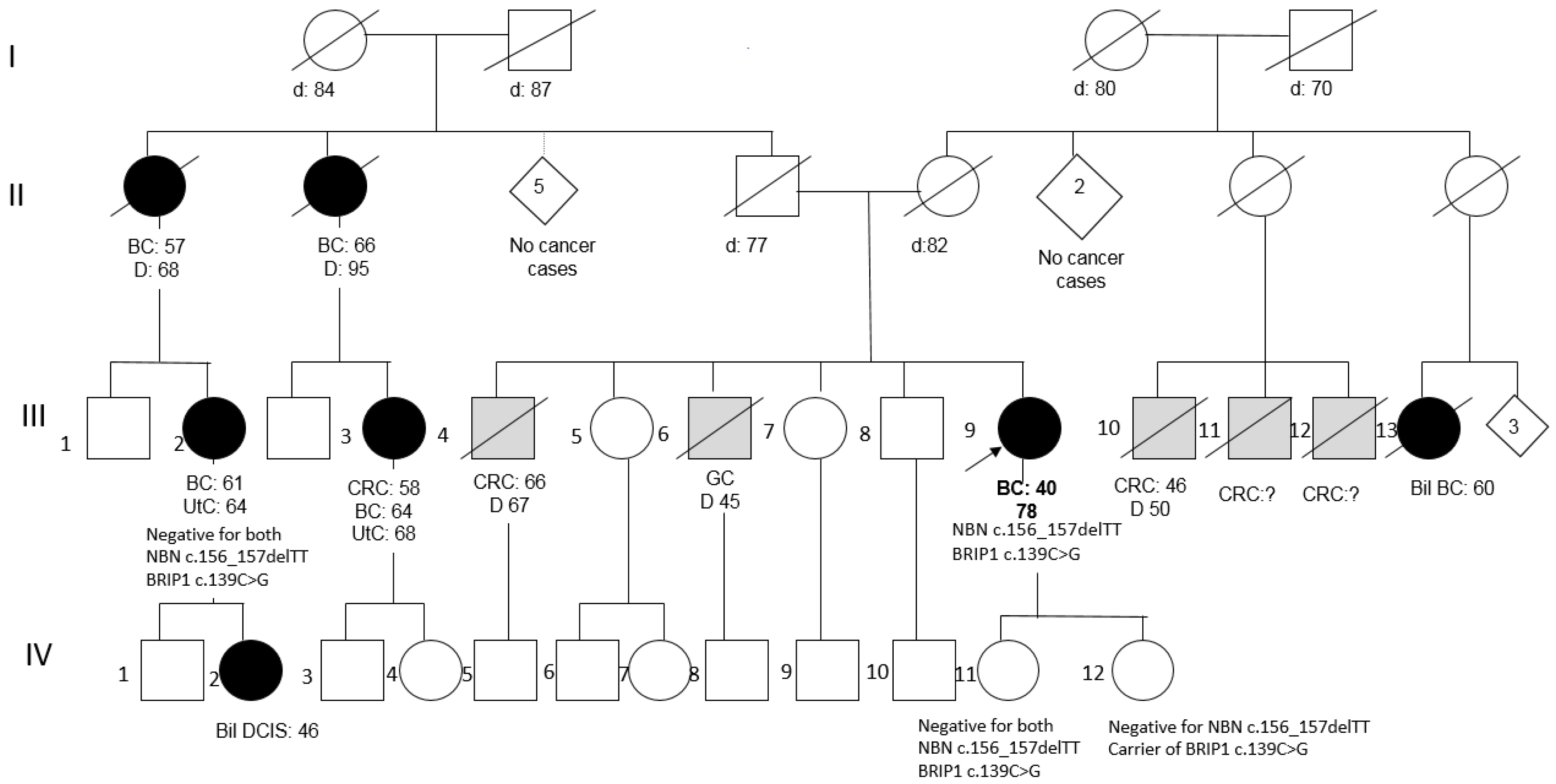
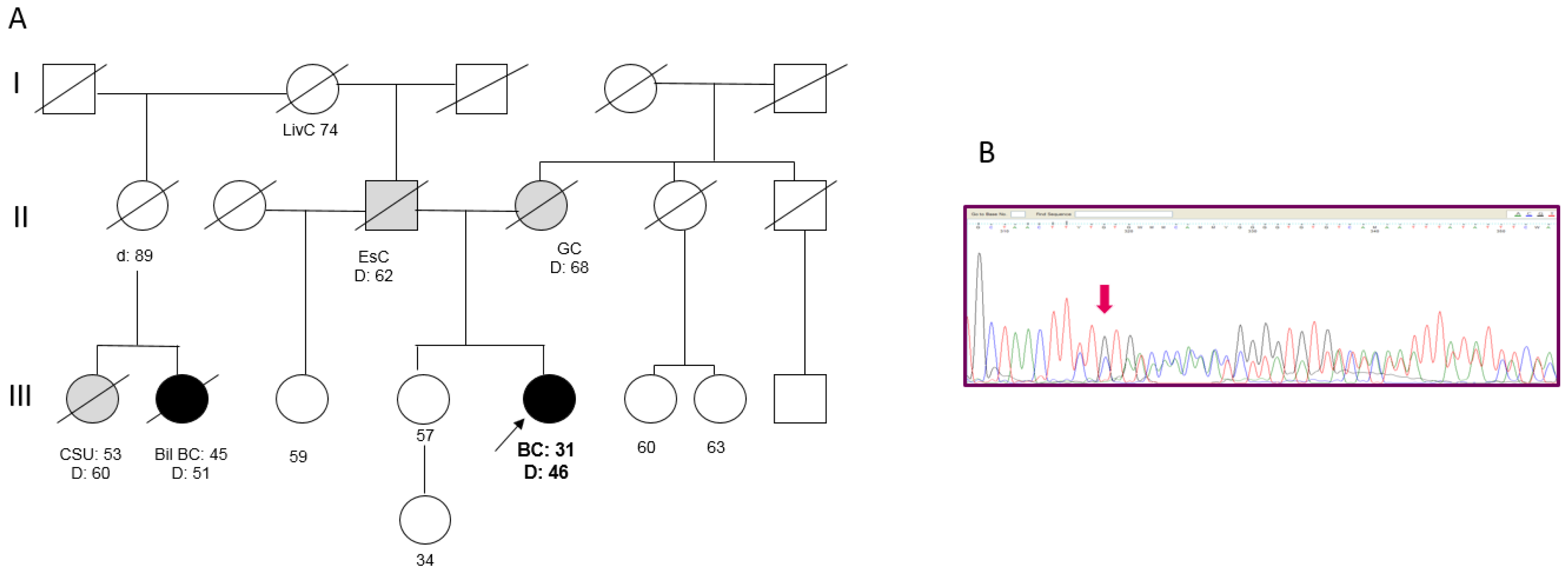
2.2. Clinical and Family History of Carriers of NBN Variants
2.3. In Silico Assessment of the Variants
2.4. Genotyping of the Independent Set of Cases and Controls
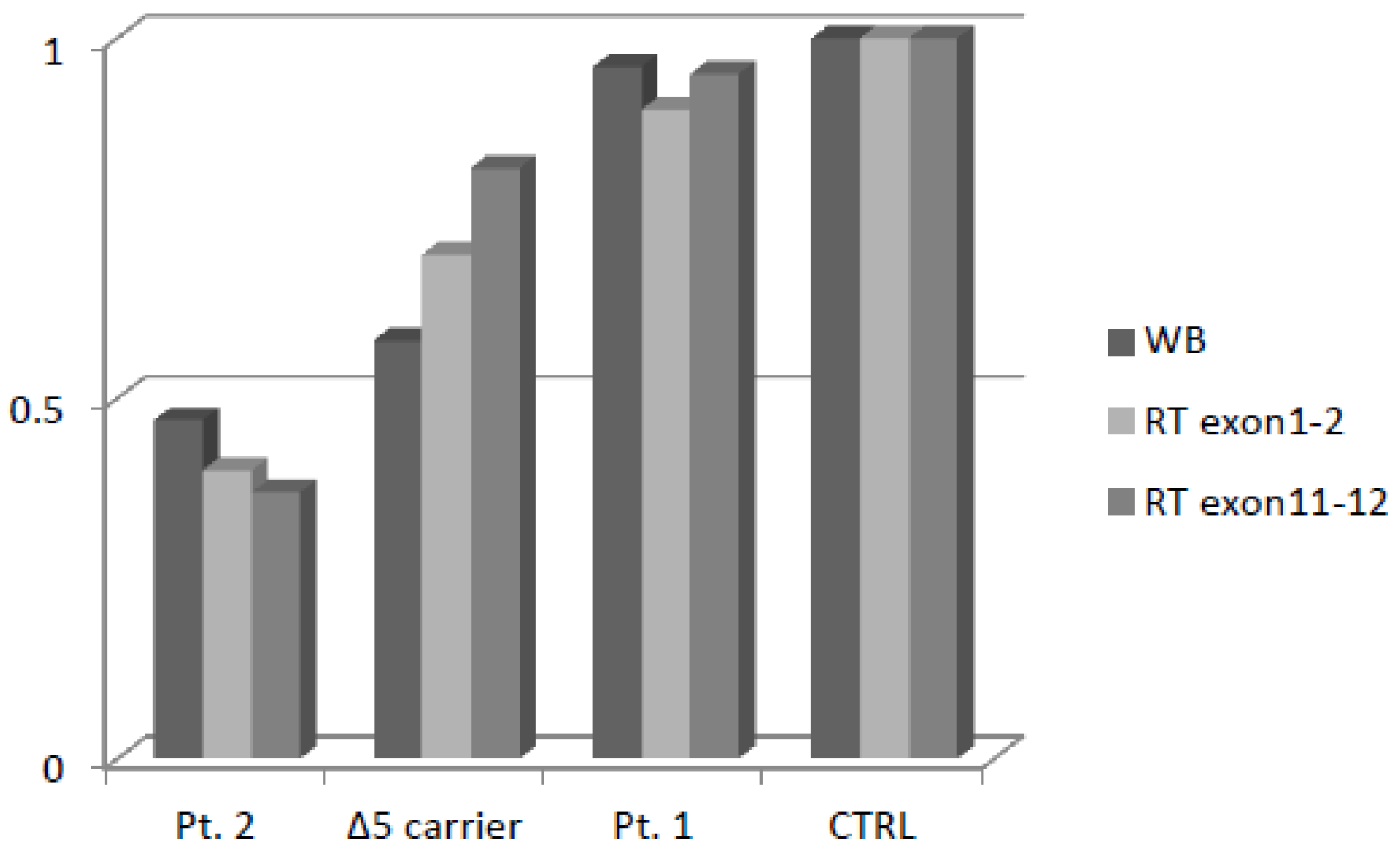
2.5. Expression of Variant Nibrin
2.6. Double-Strand Break Repair Assessment
2.7. NBN Haplotype Patterns
2.8. Whole Exome Sequencing Analysis
2.9. Review of the Literature
3. Discussion
4. Patients and Methods
4.1. Patient Recruitment
4.2. NBN Sequence Analysis
4.3. In Silico Variant Assessment
4.4. Genotyping of Independent Sets of Cases and Controls
4.5. Gene Expression Analyses
4.5.1. Real-Time PCR
4.5.2. Western Blotting
4.5.3. Double-Strand Break Repair Assessment
4.5.4. Haplotype Reconstruction and Evolutionary Analyses
4.5.5. Whole Exome Sequencing
4.5.6. Review of the Literature
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varon, R.; Vissinga, C.; Platzer, M.; Cerosaletti, K.M.; Chrzanowska, K.H.; Saar, K.; Beckmann, G.; Seemanová, E.; Cooper, P.R.; Nowak, N.J.; et al. Nibrin, a novel DNA double-strand break repair protein, is mutated in Nijmegen breakage syndrome. Cell 1998, 93, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Bogdanova, N.; Feshchenko, S.; Schürmann, P.; Waltes, R.; Wieland, B.; Hillemanns, P.; Rogov, Y.I.; Dammann, O.; Bremer, M.; Karstens, J.H.; et al. Nijmegen Breakage Syndrome mutations and risk of breast cancer. Int. J. Cancer 2008, 122, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, S.; Beauparlant, J.C.; Labrie, Y.; Ouellette, G.; Durocher, F.; INHERIT BRCAs. Variations in the NBN/NBS1 gene and the risk of breast cancer in non-BRCA1/2 French Canadian families with high risk of breast cancer. BMC Cancer 2009, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wei, Q.; Bondy, M.L.; Li, D.; Brewster, A.; Shete, S.; Yu, T.-K.; Sahin, A.; Meric-Bernstam, F.; Hunt, K.K.; et al. Polymorphisms andhaplotypes of the NBS1 gene are associated with risk of sporadic breast cancer in non-Hispanic white women <or = 55 years. Carcinogenesis 2006, 27, 2209–2216. [Google Scholar] [PubMed] [Green Version]
- Hsu, H.-M.; Wang, H.-C.; Chen, S.-T.; Hsu, G.C.; Shen, C.Y.; Yu, J.C. Breast Cancer Risk Is Associated with the Genes Encoding the DNA Double-Strand Break RepairMre11/Rad50/Nbs1 Complex. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2024–2032. [Google Scholar] [CrossRef] [Green Version]
- Roznowski, K.; Januszkiewicz-Lewandowska, D.; Mosor, M.; Pernak, M.; Litwiniuk, M.; Nowak, J. I171V germline mutation in the NBS1 gene significantly increases risk of breast cancer. BreastCancer Res. Treat. 2008, 110, 343–348. [Google Scholar] [CrossRef]
- Górski, B.; Debniak, T.; Masojc, B.; Mierzejewski, M.; Mdrek, K.; Cybulski, C.; Jakubowska, A.; Kurzawski, G.; Chosia, M.; Scott, R.; et al. Germline 657del5 mutation in theNBS1 gene in breast cancer patients. Int. J. Cancer 2003, 106, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, K.; Rapakko, K.; Karppinen, S.-M.; Erkko, H.; Knuutila, S.; Lundán, T.; Mannermaa, A.; Børresen-Dale, A.-L.; Borg, Å.; Barkardottir, R.B.; et al. RAD50 and NBS1 are breast cancer susceptibility genes associated with genomic instability. Carcinogenesis 2006, 27, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zeng, Y.; Liu, Z.; Wei, W. Significant association between Nijmegen breakage syndrome 1 657del5 polymorphism and breast cancer risk. Tumour Biol. 2013, 34, 2753. [Google Scholar] [CrossRef] [PubMed]
- Rusak, B.; Kluźniak, W.; Wokołorczyk, D.; Stempa, K.; Kashyap, A.; Rudnicka, H.; Gronwald, J.; Huzarski, T.; Dębniak, T.; Jakubowska, A.; et al. Polish Hereditary Breast Cancer Consortium. Allelic modification of breast cancer risk in women with an NBN mutation. Breast Cancer Res. Treat. 2019, 178, 427–431. [Google Scholar] [CrossRef]
- Castéra, L.; Krieger, S.; Rousselin, A.; Legros, A.; Baumann, J.J.; Bruet, O.; Brault, B.; Fouillet, R.; Goardon, N.; Letac, O.; et al. Next-generation sequencing for the diagnosis of hereditary breast and ovarian cancer using genomic capture targeting multiple candidate genes. Eur. J. Hum. Genet. 2014, 22, 1305–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurian, A.W.; Hare, E.E.; Mills, M.A.; Kingham, K.E.; McPherson, L.; Whittemore, A.S.; McGuire, V.; Ladabaum, U.; Kobayashi, Y.; Lincoln, S.E.; et al. Clinical evaluation of a multiple-gene sequencing panel for hereditary cancer risk assessment. J. Clin. Oncol. 2014, 32, 2001–2009. [Google Scholar] [CrossRef] [Green Version]
- Domagala, P.; Jakubowska, A.; Jaworska-Bieniek, K.; Kaczmarek, K.; Durda, K.; Kurlapska, A.; Cybulski, C.; Lubinski, J. Prevalence of Germline Mutations in Genes Engaged in DNA Damage Repair by Homologous Recombination in Patients with Triple-Negative and Hereditary Non-Triple-Negative Breast Cancers. PLoS ONE 2015, 10, e0130393. [Google Scholar] [CrossRef]
- Desmond, A.; Kurian, A.W.; Gabree, M.; Mills, M.A.; Anderson, M.J.; Kobayashi, Y.; Horick, N.; Yang, S.; Shannon, K.M.; Tung, N.; et al. Clinical Actionability of Multigene Panel Testing for Hereditary Breast and Ovarian Cancer Risk Assessment. JAMA Oncol. 2015, 1, 943–951. [Google Scholar] [CrossRef]
- Maxwell, K.N.; Wubbenhorst, B.; Andrea, K.; Garman, B.; Long, J.M.; Powers, J.; Rathbun, K.; Stopfer, J.E.; Zhu, J.; Bradbury, A.R.; et al. Prevalence of mutations in a panel of breast cancer susceptibility genes in BRCA1/2-negative patients with early-onset breast cancer. Genet. Med. 2015, 17, 630–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Meeks, H.; Feng, B.J.; Healey, S.; Thorne, H.; Makunin, I.; Ellis, J.; kConFab Investigators; Campbell, I.; Southey, M.; et al. Targeted massively parallel sequencing of a panel of putative breast cancer susceptibility genes in a large cohort of multiple-case breast and ovarian cancer families. Med. Genet. 2016, 53, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Tung, N.; Lin, N.U.; Kidd, J., II; Allen, B.A.; Singh, N.; Wenstrup, R.J.; Hartman, A.R.; Winer, E.P.; Garber, J.E. Frequency of Germline Mutations in 25 Cancer Susceptibility Genes in a Sequential Series of Patients With Breast Cancer. J. Clin. Oncol. 2016, 34, 1460–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.; Mandell, J.B.; Norquist, B.M.; Casadei, S.; Gulsuner, S.; Lee, M.K.; King, M.C. Genetic Predisposition to Breast Cancer Due to Mutations Other Than BRCA1 and BRCA2 Founder Alleles Among Ashkenazi Jewish Women. JAMA Oncol. 2017, 3, 1647–1653. [Google Scholar] [CrossRef]
- Kraus, C.; Hoyer, J.; Vasileiou, G.; Wunderle, M.; Lux, M.P.; Fasching, P.A.; Krumbiegel, M.; Uebe, S.; Reuter, M.; Beckmann, M.W.; et al. Gene panel sequencing in familial breast/ovarian cancer patients identifies multiple novel mutations also in genes others than BRCA1/2. Int. J. Cancer 2017, 140, 95–102. [Google Scholar] [CrossRef]
- Jian, W.; Shao, K.; Qin, Q.; Xiaohong, X.; Song, S.; Wang, X. Clinical and genetic characterization of hereditary breast cancer in a Chinese population. Hered. Cancer Clin. Pract. 2017, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Hauke, J.; Horvath, J.; Groß, E.; Gehrig, A.; Honisch, E.; Hackmann, K.; Schmidt, G.; Arnold, N.; Faust, U.; Sutter, C.; et al. Gene panel testing of 5589 BRCA1/2-negative index patients with breast cancer in a routine diagnostic setting: Results of the German Consortium for Hereditary Breast and Ovarian Cancer. Cancer Med. 2018, 7, 1349–1358. [Google Scholar] [CrossRef]
- Schubert, S.; van Luttikhuizen, J.L.; Auber, B.; Schmidt, G.; Hofmann, W.; Penkert, J.; Davenport, C.F.; Hille-Betz, U.; Wendeburg, L.; Bublitz, J.; et al. The identification of pathogenic variants in BRCA1/2 negative, high risk, hereditary breast and/or ovarian cancer patients: High frequency of FANCM pathogenic variants. Int. J. Cancer 2019, 144, 2683–2694. [Google Scholar] [CrossRef]
- Kurian, A.W.; Ward, K.C.; Howlader, N.; Deapen, D.; Hamilton, A.S.; Mariotto, A.; Miller, D.; Penberthy, L.S.; Katz, S.J. Genetic Testing and Results in a Population-Based Cohort of Breast Cancer Patients and Ovarian Cancer Patients. J. Clin. Oncol. 2019, 37, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Bernstein-Molho, R.; Barnes-Kedar, I.; Ludman, M.D.; Reznik, G.; Feldman, H.B.; Samra, N.N.; Eilat, A.; Peretz, T.; Peretz, L.P.; Shapira, T.; et al. The yield of full BRCA1/2 genotyping in Israeli Arab high-risk breast/ovarian cancer patients. Breast Cancer Res. Treat. 2019, 178, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Scarpitta, R.; Zanna, I.; Aretini, P.; Gambino, G.; Scatena, C.; Mei, B.; Ghilli, M.; Rossetti, E.; Roncella, M.; Congregati, C.; et al. Germline investigation in male breast cancer of DNA repair genes by next-generation sequencing. Breast Cancer Res. Treat. 2019, 178, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Guo, X.; Wen, W.; Shi, J.; Long, J.; Cai, Q.; Shu, X.O.; Xiang, Y.; Zheng, W. Association Between One-carbon Metabolism-related Vitamins and Risk of Breast Cancer: A Systematic Review and Meta-analysis of Prospective Studies. Clin. Breast Cancer. 2020, 20, e469–e480. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Yang, L.S.; Huang, F.; Hao, J.H.; Su, P.Y.; Sun, Y.H. Current evidence on the relationship between two polymorphisms in the NBS1 gene and breast cancer risk: A meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 5375–5379. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.S.; Mclellan, J.; Verbakel, J.Y.; Aronson, J.K.; Lasserson, D.S.; Pidduck, N.; Roberts, N.; Fleming, S.; O’Callaghan, C.A.; Bankhead, C.R.; et al. Effects of antihypertensives, lipid-modifying drugs, glycaemic control drugs and sodium bicarbonate on the progression of stages 3 and 4 chronic kidney disease in adults: A systematic review and meta-analysis. BMJ Open 2019, 9, e030596. [Google Scholar] [CrossRef]
- Moretta, J.; Berthet, P.; Bonadona, V.; Caron, O.; Cohen-Haguenauer, O.; Colas, C.; Corsini, C.; Cusin, V.; De Pauw, A.; Delnatte, C.; et al. Recommandations françaises du Groupe Génétique et Cancer pour l’analyse en panel de gènes dans les prédispositions héréditaires au cancer du sein ou de l’ovaire. Bull. Cancer 2018, 105, 907–917. [Google Scholar] [CrossRef]
- Hu, C.; Hart, S.N.; Gnanaolivu, R.; Huang, H.; Lee, K.Y.; Na, J.; Gao, C.; Lilyquist, J.; Yadav, S.; Boddicker, N.J.; et al. A Population-Based Study of Genes Previously Implicated in Breast Cancer. N. Engl. J. Med. 2021, 384, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Breast Cancer Association Consortium. Breast Cancer Risk Genes-Association Analysis in More than 113,000 Women. N. Engl. J. Med. 2021, 384, 428–439. [Google Scholar] [CrossRef]
- Easton, D.F.; Pharoah, P.D.; Antoniou, A.C.; Tischkowitz, M.; Tavtigian, S.V.; Nathanson, K.L.; Devilee, P.; Meindl, A.; Couch, F.J.; Southey, M.; et al. Gene-panel sequencing and the prediction of breast-cancer risk. N. Engl. J. Med. 2015, 372, 2243–2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Servizio Sanità Pubblica, Regione Emilia-Romagna: Contributo n. 91/2016: “Protocollo Assistenziale Nelle Donne a Rischio Ereditario di Tumore Della Mammella e/o ovaio”. 2016. ISSN 2464-9252. N°91-2016. Available online: https://salute.regione.emilia-romagna.it/normativa-e-documentazione/rapporti/contributi (accessed on 24 May 2021).
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. 1000 Genomes Project Consortium. Nature 2015, 526, 68–74. [Google Scholar] [PubMed] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, M.; Scheet, P. Accounting for decay of linkage disequilibrium in haplotype inference and missing-data imputation. Am. J. Hum. Genet. 2005, 76, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Cox, M.P. Accuracy of molecular dating with the rho statistic: Deviations from coalescent expectations under a range of demographic models. Hum. Biol. 2008, 80, 335–357. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Country | Study Population | Testing Type | N Patients | N with NBN Variants (%) | Specific Variants (N) | Proband Cancer History | Variant Frequency in the Study | Population Frequency (gnomAD) § | p Value | Current Classification (ClinVar/LOVD) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| [11] | France | Pts with suspected hereditary BC | NGS panel | 708 | 8 (1.13%) | c.1142delC (1) | ND | 0.14% | 0.005% 1 | 0.01 | C4/C5 |
| c.156_157delTT (1) | 0.14% | 0.004% 1,2 | 0.01 | C4/C5 | |||||||
| c.37+5G>A (3) | 0.42% | 0.56% 3 | 0.82 | C1/C2 | |||||||
| c.38-10T>A (1) | 0.14% | 0.03% 3 | 0.50 | C3 | |||||||
| c.657_661delACAAA (1) | 0.14% | 0.04% 1,2,4,5 | 0.69 | C5 | |||||||
| c.788T>C (1) | 0.14% | 0.04% 1,2,6 | 0.68 | C3 | |||||||
| [12] | USA | Pts previously tested for BRCA mut | NGS panel (42 genes) | 198 | 2 (1.01%) | c.643C>T (2) | (1) BC age 50; FH: BC, Mel (2) no cancer; FH: BC, CRC, OC | 1.01% | 0.49% 3 | 0.59 | C3 |
| [13] | Poland | Pts with TNBC (158) or suspected hereditary, NTNBC (44) | Screening of 36 mutations in 8 genes | 202 | 7 (3.47%) | c.657_661delACAAA (1) | NTNBC age 63 | 0.49% | 0.04% | 0.14 | C5 |
| c.511A>G (6 *) | 4: TNBC age 44–81 | ||||||||||
| 2: NTNBC age 49, 59 | 2.97% | 0.30% 3 | <0.0001 | C1 | |||||||
| [14] | USA | Pts referred for BRCA and tested negative | Commercial NGS panels (25–29 genes) | 1046 | 2 (0.19%) only deleterious variants reported | c.657_661delACAAA (1) | BC age 54, PC; FH: BC, PrC, Mel, PC, LC, others BC age 49; FH: BC | 0.10% | 0.04% | 0.91 | C5 |
| c.1142delC (1) | 0.10% | 0.005% | 0.07 | C4/C5 | |||||||
| [15] | USA | Pts with early-onset BC (<40) BRCA-negative | NGS panel (22 genes) | 278 | 1 (0.36%) | c.664T>C | BC age 37, Leu 39; FH: PrC, Mel | 0.36% | 0.004% 1 | <0.0001 | C3 |
| [16] | Australia | Familial BC pts | NGS panel (19 genes) | 684 | 1 (0.15%) | c.698_701delAACA (1) | BC age 42; FH: PrC | 0.15% | 0.004% 1,6 | 0.01 | C5 |
| [17] | USA | BC pts | NGS panel (25 genes) | 488 | 1 (0.2%) | c.127C>T (1) | TNBC age 56; FH: LC, CNS, PC | 0.2% | 0.006% 1,4,6 | 0.01 | C5 |
| [18] | USA | BC pts with Ashkenazi ancestry | NGS panel (23 genes) | 1007 | 1 (0.1%) | c.1903A>T (1) | BC age 42 | 0.1% | 0.15% 7 | 0.96 | C5 |
| [19] | Germany | BC/OC pts | NGS panel (14 genes) | 581 | 6 (1.03%) | c.1397+1delG (1) | NTNBC age 35; FH: BC NTNBC age 43; FH: BC NTNBC age 52; FH: BC (1)BC age 53; FH: BC (2) BC age 49; FH: BC (3)TNBC/OC age 56/66; FH: BC, LC, Leu | 0.17% | 0.0008% 2 | <0.0001 | C5 |
| c.2028delT(1) | 0.17% | ND | - | ND (C4) | |||||||
| c.2097dupT (1) | 0.17% | ND | - | ND (C4) | |||||||
| c.657_661delACAAA (3) | 0.52% | 0.04% | <0.0001 | C5 | |||||||
| c | |||||||||||
| [20] | China | Women with personal or familial history of BC | NGS panel (27 genes) | 240 | 1 (0.42%) | c.2140C>T (1) | BC age 33 | 0.42% | 0.005% 1,7 | <0.0001 | C5 |
| [21] | Germany | BC pts tested negative for BRCA | NGS panels (8 genes selected) | 5589 | 12 (0.21%) only truncating variants reported | c.123delC (1) | ND | 0.02% | 0.0008% 1 | 0.15 | C5 |
| c.211_212insGA (1) | 0.02% | 0.0008% 1 | 0.15 | C5 | |||||||
| c.657_661delACAAA (7) | 0.13% | 0.04% | 0.008 | C5 | |||||||
| c.1141del (1) | 0.02% | ND | - | ND (C4) | |||||||
| c.1396del (1) | 0.02% | ND | - | ND (C4) | |||||||
| c.1651dup (1) | 0.02% | 0.002% 6 | 0.29 | C5 | |||||||
| [22] | Germany | BC pts tested negative for BRCA | NGS panel (94 genes) | 237 | 1 (0.4%) VUS excluded | c.657_661delACAAA | BC age 32 | 0.4% | 0.04% | 0.19 | C5 |
| [23] | US | BC/OC pts | NGS panels (11 genes selected) | 5436 | ND (0.35%) path ND (2.5%) VUS | ND | |||||
| [24] | Israel | BC pts tested negative for founder BRCA variants | Commercial NGS panels (30–83 genes) | 144 | 1 (0.7%) VUS excluded | c.966C>G | ND (BRCAPRO prob: 0.6%) | 0.7% | ND | - | ND (C4) |
| [25] | Italy | Male BC pts | NGS panel (24 genes) | 81 | 1 (1.2%) | c.547G>A | Male BC age 49; FH: BC | 1.2% | 0.003% 1,7 | <0.0001 | C3 |
| [26] | China | BC pts | Virtual panel from WES | 831 | 1 (0.1%) | c.127C>T | ND | 0.1% | 0.006% 1,4,6 | 0.07 | C5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuntini, R.; Bonora, E.; Pradella, L.M.; Amato, L.B.; Vidone, M.; De Fanti, S.; Catucci, I.; Cortesi, L.; Medici, V.; Ferrari, S.; et al. Detecting Variants in the NBN Gene While Testing for Hereditary Breast Cancer: What to Do Next? Int. J. Mol. Sci. 2021, 22, 5832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115832
Zuntini R, Bonora E, Pradella LM, Amato LB, Vidone M, De Fanti S, Catucci I, Cortesi L, Medici V, Ferrari S, et al. Detecting Variants in the NBN Gene While Testing for Hereditary Breast Cancer: What to Do Next? International Journal of Molecular Sciences. 2021; 22(11):5832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115832
Chicago/Turabian StyleZuntini, Roberta, Elena Bonora, Laura Maria Pradella, Laura Benedetta Amato, Michele Vidone, Sara De Fanti, Irene Catucci, Laura Cortesi, Veronica Medici, Simona Ferrari, and et al. 2021. "Detecting Variants in the NBN Gene While Testing for Hereditary Breast Cancer: What to Do Next?" International Journal of Molecular Sciences 22, no. 11: 5832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115832