
Apex Resection in Zebrafish (Danio rerio) as a Model of Heart Regeneration: A Video-Assisted Guide

 and
and 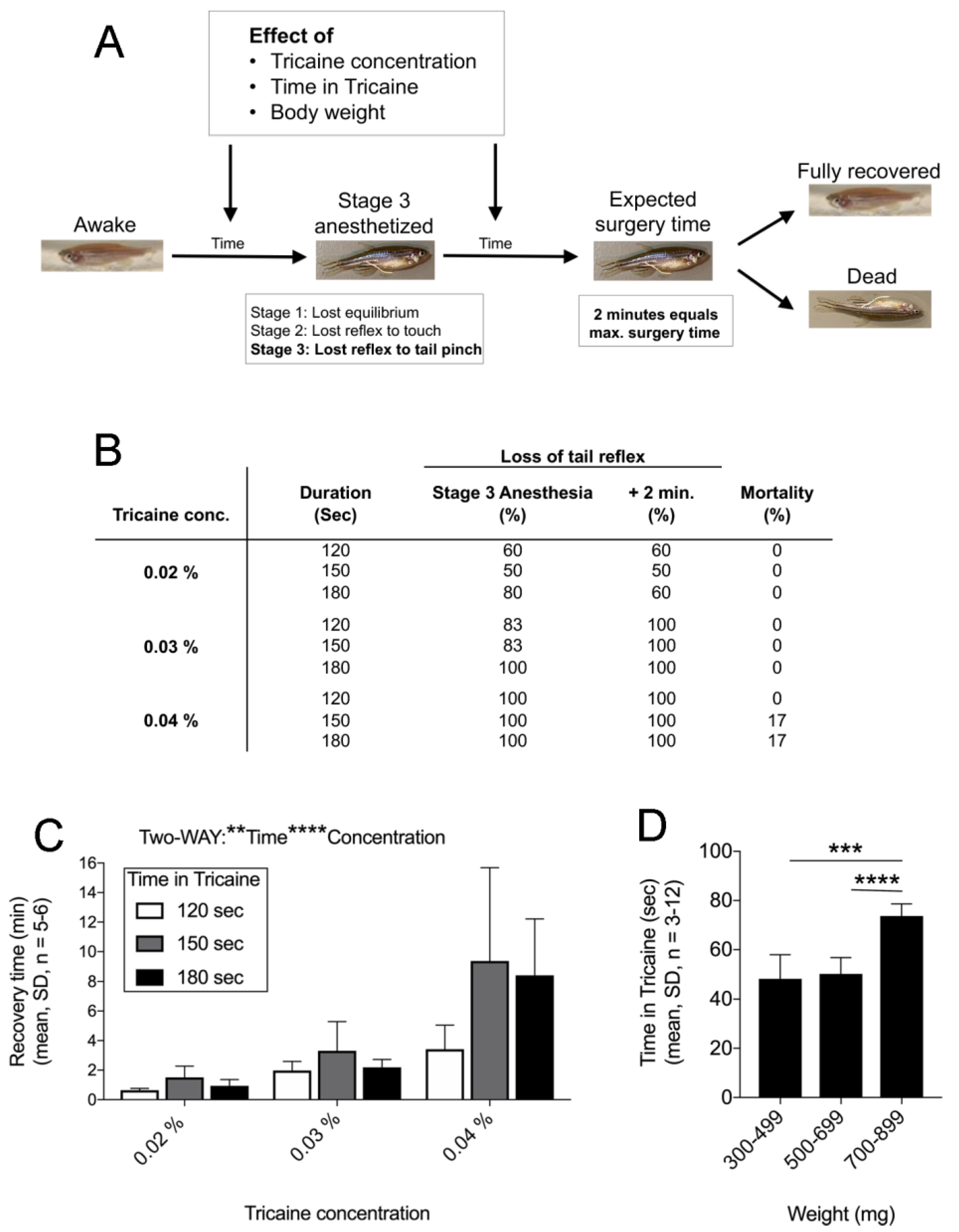{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Finetuning Anaesthesia in Zebrafish
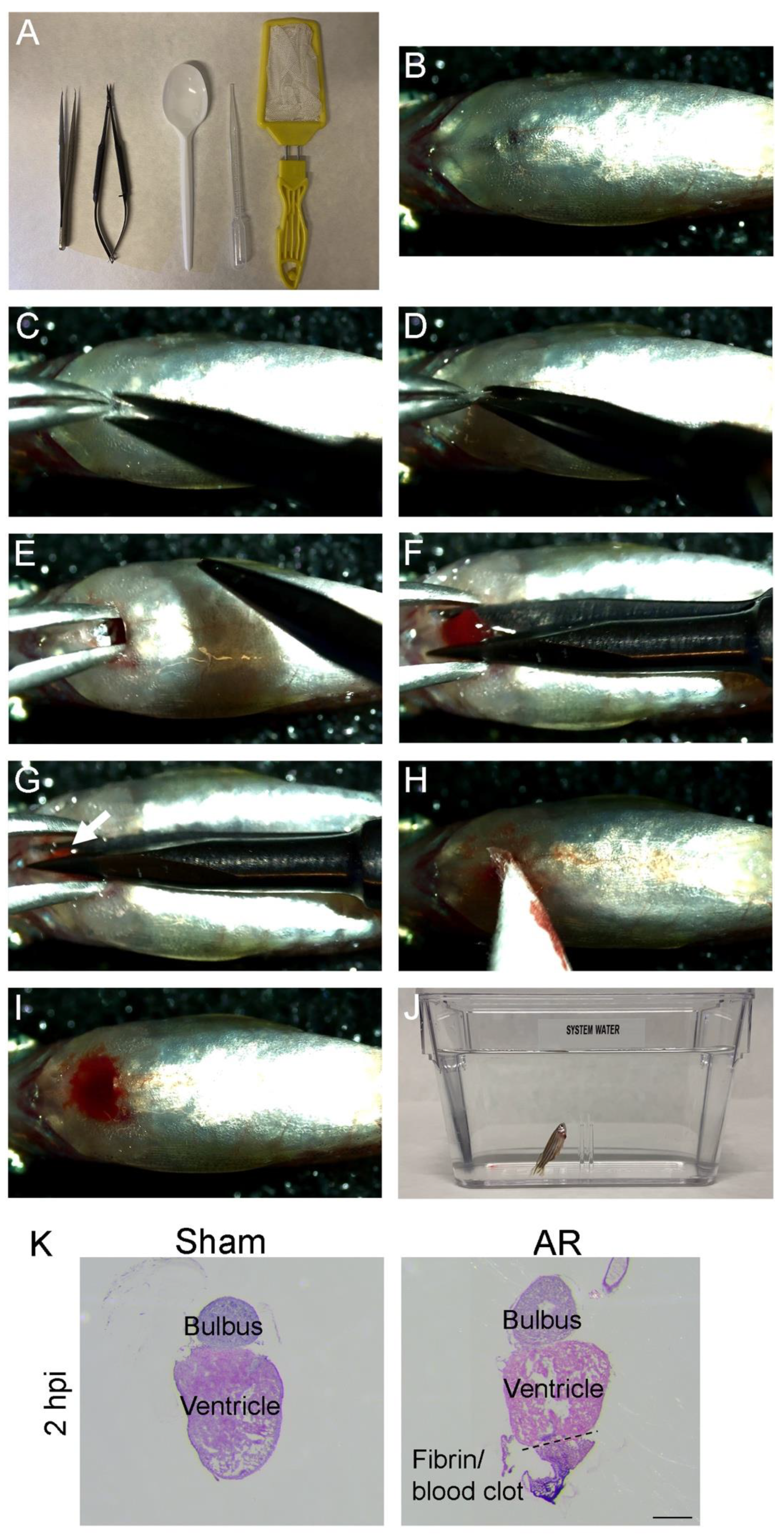
2.2. Apex Resection (AR) in Zebrafish Step-by-Step
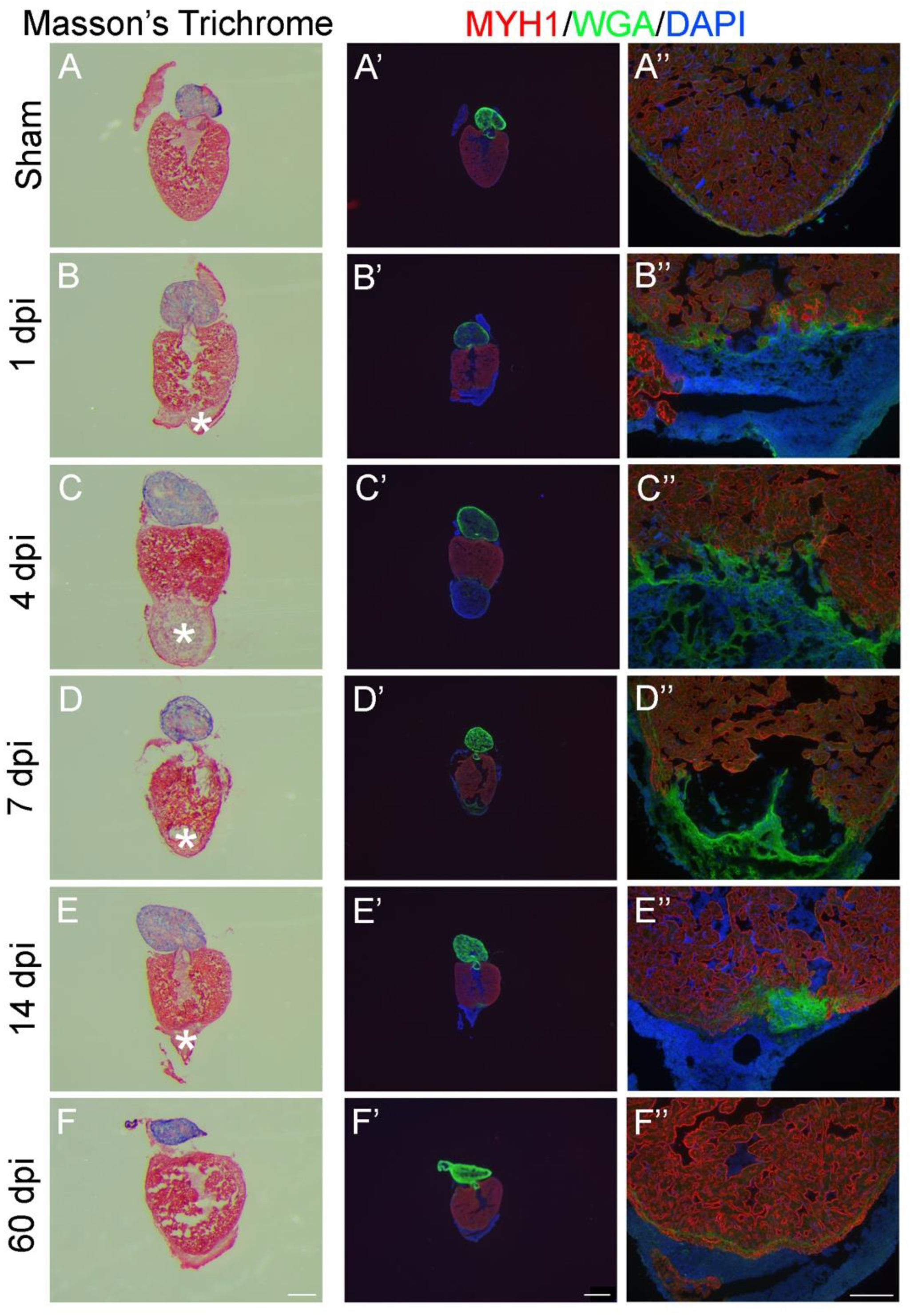
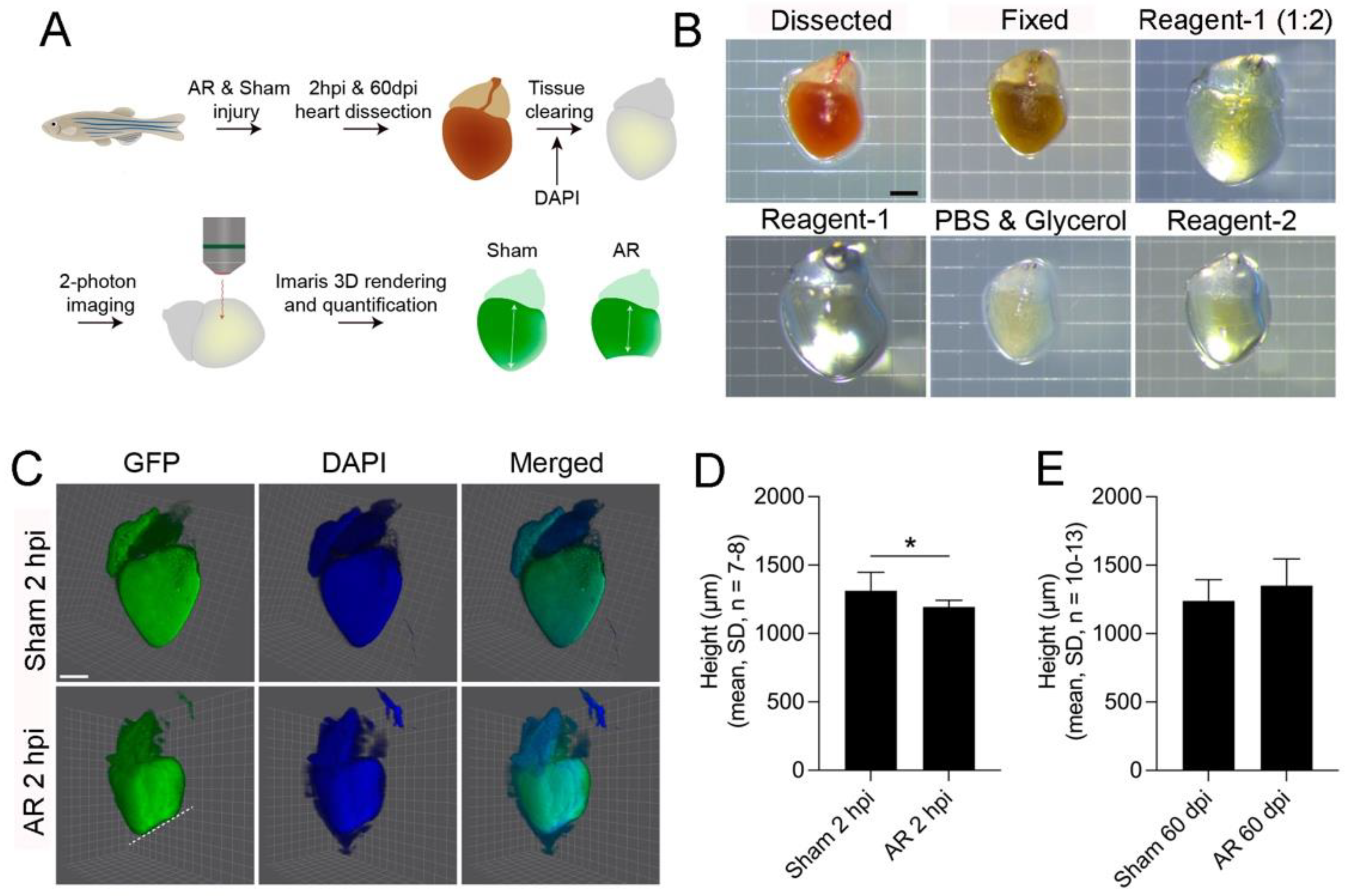
2.3. 2D- and 3D-Analysis for Heart Regeneration
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Zebrafish
4.3. Materials Required for Apex Resection (AR) in Zebrafish
- 2 small aquariums/tanks for sedation and recovery
- Small fishing net
- Microscope (e.g., Leica, S9i)
- Large plastic spoon
- Pasteur pipette
- Fine curved forceps (S&T, Agnthos, Lidingo, Sweden, cat. no. JFCL-3P TC)
- Micro scissors (LAWTON, Fridingen an der Donau, Germany, cat. no. 05-0050-C)
- Sponge with pre-cut slit big enough to hold the zebrafish
- Pro-ophta, pre-cut on the middle in a 30° angle/cotton swab
- Tissues for removing (zebrafish) scales from surgical instruments
- Tricaine solutions in water from your system/aquarium for anaesthesia
- Water from your system/aquarium
4.4. Tricaine Stock Solution
4.5. Anaesthesia of Zebrafish
4.6. Surgical Procedures and Recovery
4.7. Tissue Processing for 2D Analysis
4.8. Haematoxylin and Eosin Staining
4.9. Masson’s Trichrome Staining
4.10. Fluorescence staining with Myosin Heavy Chain Antibody (MYH1), Wheat Germ Agglutinin (WGA) and 4′,6′-diamidino-2-phenylindole (DAPI)
4.11. CUBIC Tissue Clearing and 2-Photon Imaging
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Amerongen, M.J.; Engel, F.B. Features of Cardiomyocyte Proliferation and its Potential for Cardiac Regeneration. J. Cell. Mol. Med. 2008, 12, 2233–2244. [Google Scholar] [CrossRef] [Green Version]
- Burke, A.P.; Virmani, R. Pathophysiology of Acute Myocardial Infarction. Med. Clin. N. Am. 2007, 91, 553–572. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart Regeneration in Zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Raya, A.; Koth, C.M.; Buscher, D.; Kawakami, Y.; Itoh, T.; Raya, R.M.; Sternik, G.; Tsai, H.-J.; Rodríguez-Esteban, C.; Izpisúa-Belmonte, J.C. Activation of Notch Signaling Pathway Precedes Heart Regeneration in Zebrafish. Proc. Natl. Acad. Sci. USA 2003, 100, 11889–11895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rosa, J.M.; Mercader, N. Cryoinjury as a Myocardial Infarction Model for the Study of Cardiac Regeneration in the Zebrafish. Nat. Protoc. 2012, 7, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Poss, K.D. Methodologies for Inducing Cardiac Injury and Assaying Regeneration in Adult Zebrafish. Methods Mol. Biol. 2016, 1451, 225–235. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Lepilina, A.; Coon, A.N.; Kikuchi, K.; Holdway, J.E.; Roberts, R.W.; Burns, C.G.; Poss, K.D. A Dynamic Epicardial Injury Response Supports Progenitor Cell Activity during Zebrafish Heart Regeneration. Cell 2006, 127, 607–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, K.; Holdway, J.E.; Major, R.J.; Blum, N.; Dahn, R.D.; Begemann, G.; Poss, K.D. Retinoic Acid Production by Endocardium and Epicardium is an Injury Response Essential for Zebrafish Heart Regeneration. Dev. Cell 2011, 20, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Cao, J.; Dickson, A.L.; Poss, K.D. Epicardial Regeneration is Guided by Cardiac Outflow Tract and Hedgehog Signalling. Nat. Cell Biol. 2015, 522, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Jopling, C.; Sleep, E.; Raya, M.; Marti, M.; Raya, A.; Izpisua Belmonte, J.C. Zebrafish Heart Regeneration Occurs by Cardi-omyocyte Dedifferentiation and Proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Holdway, J.E.; Werdich, A.A.; Anderson, R.M.; Fang, Y.; Egnaczyk, G.F.; Evans, T.; MacRae, C.A.; Stainier, D.; Poss, K.D. Primary Contribution to Zebrafish Heart Regeneration by gata4+ Cardiomyocytes. Nat. Cell Biol. 2010, 464, 601–605. [Google Scholar] [CrossRef]
- Kikuchi, K.; Gupta, V.; Wang, J.; Holdway, J.E.; Wills, A.A.; Fang, Y.; Poss, K.D. tcf21+ Epicardial Cells Adopt Non-Myocardial Fates during Zebrafish Heart Development and Regeneration. Development 2011, 138, 2895–2902. [Google Scholar] [CrossRef] [Green Version]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.-D.; Guerquin-Kern, J.-L.; Lechene, C.P.; Lee, R.T. Mammalian Heart Renewal by Pre-Existing Cardiomyocytes. Nat. Cell Biol. 2013, 493, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bersell, K.; Arab, S.; Haring, B.; Kühn, B. Neuregulin1/ErbB4 Signaling Induces Cardiomyocyte Proliferation and Repair of Heart Injury. Cell 2009, 138, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargehr, J.; Ong, L.P.; Colzani, M.; Davaapil, H.; Hofsteen, P.; Bhandari, S.; Gambardella, L.; Le Novère, N.; Iyer, D.; Sampaziotis, F.; et al. Epicardial Cells Derived from Human Embryonic Stem Cells Augment Cardiomyocyte-Driven Heart Regeneration. Nat. Biotechnol. 2019, 37, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Thatcher, J.E.; McAnally, J.; Kong, Y.; Qi, X.; Tan, W.; DiMaio, J.M.; Amatruda, J.F.; Gerard, R.D.; Hill, J.A.; et al. C/EBP Transcription Factors Mediate Epicardial Activation During Heart Development and Injury. Science 2012, 338, 1599–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, N.; Bollini, S.; Dubé, K.N.; Vieira, J.M.; Zhou, B.; Davidson, S.; Yellon, D.; Riegler, J.; Price, A.N.; Lythgoe, M.; et al. De novo Cardiomyocytes from within the Activated Adult Heart after Injury. Nat. Cell Biol. 2011, 474, 640–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belling, H.J.; Hofmeister, W.; Andersen, D.C. A Systematic Exposition of Methods used for Quantification of Heart Regeneration after Apex Resection in Zebrafish. Cells 2020, 9, 548. [Google Scholar] [CrossRef] [Green Version]
- Sheng, D.Z.; Zheng, D.; Kikuchi, K. Cardiac Resection Injury in Zebrafish. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin, Germany, 2020; pp. 63–69. [Google Scholar]
- Collymore, C.; Tolwani, A.; Lieggi, C.; Rasmussen, S. Efficacy and Safety of 5 Anesthetics in Adult Zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 198–203. [Google Scholar]
- Carter, K.M.; Woodley, C.M.; Brown, R.S. A Review of Tricaine Methanesulfonate for Anesthesia of Fish. Rev. Fish Biol. Fish. 2011, 21, 51–59. [Google Scholar] [CrossRef]
- Valentim, A.M.; Félix, L.M.; Carvalho, L.; Diniz, E.; Antunes, L.M. A New Anaesthetic Protocol for Adult Zebrafish (Danio rerio): Propofol Combined with Lidocaine. PLoS ONE 2016, 11, e0147747. [Google Scholar] [CrossRef]
- Ehrlich, O.; Karamalakis, A.; Krylov, A.J.; Dudczig, S.; Hassell, K.L.; Jusuf, P.R. Clove Oil and AQUI-S Efficacy for Zebrafish Embryo, Larva, and Adult Anesthesia. Zebrafish 2019, 16, 451–459. [Google Scholar] [CrossRef]
- Narumanchi, S.; Kalervo, K.; Perttunen, S.; Wang, H.; Immonen, K.; Kosonen, R.; Laine, M.; Ruskoaho, H.; Tikkanen, I.; Lakkisto, P.; et al. Inhibition of let-7c Regulates Cardiac Regeneration after Cryoinjury in Adult Zebrafish. J. Cardiovasc. Dev. Dis. 2019, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salic, A.; Mitchison, T.J. A Chemical Method for Fast and Sensitive Detection of DNA Synthesis In vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 2415–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariel, P. A Beginner’s Guide to Tissue Clearing. Int. J. Biochem. Cell Biol. 2017, 84, 35–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susaki, A.E.; Tainaka, K.; Perrin, D.; Yukinaga, H.; Kuno, A.; Ueda, H. Advanced CUBIC Protocols for Whole-Brain and Whole-Body Clearing and Imaging. Nat. Protoc. 2015, 10, 1709–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, T.; Diniz, E.; Félix, L.M.; Antunes, L. Evaluation of Anaesthetic Protocols for Laboratory Adult Zebrafish (Danio rerio). PLoS ONE 2018, 13, e0197846. [Google Scholar] [CrossRef] [Green Version]
- Matthews, M.; Varga, Z.M. Anesthesia and Euthanasia in Zebrafish. ILAR J. 2012, 53, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Sande-Melón, M.; Marques, I.J.; Galardi-Castilla, M.; Langa, X.; Pérez-López, M.; Botos, M.A.; Sánchez-Iranzo, H.; Guz-mán-Martínez, G.; Ferreira Francisco, D.M.; Pavlinic, D.; et al. Adult sox10(+) Cardio-Myocytes Contribute to Myocardial Regeneration in the Zebrafish. Cell Rep. 2019, 29, 1041–1054.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rosa, J.M.; Burns, C.E. Zebrafish Heart Regeneration: 15 Years of Discoveries. Regeneration 2017, 4, 105–123. [Google Scholar] [CrossRef] [PubMed]
- González-Rosa, J.M.; Martín, V.; Peralta, M.; Torres, M.; Mercader, N. Extensive Scar Formation and Regression during Heart Regeneration after Cryoinjury in Zebrafish. Development 2011, 138, 1663–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-J.; Tu, C.-T.; Hsiao, C.-D.; Hsieh, F.-J.; Tsai, H.-J. Germ-Line Transmission of a Myocardium-Specific GFP Transgene Reveals Critical Regulatory Elements in the Cardiac Myosin Light Chain 2 Promoter of Zebrafish. Dev. Dyn. 2003, 228, 30–40. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellman, D.G.; Slaiman, I.M.; Mathiesen, S.B.; Andersen, K.S.; Hofmeister, W.; Ober, E.A.; Andersen, D.C. Apex Resection in Zebrafish (Danio rerio) as a Model of Heart Regeneration: A Video-Assisted Guide. Int. J. Mol. Sci. 2021, 22, 5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115865
Ellman DG, Slaiman IM, Mathiesen SB, Andersen KS, Hofmeister W, Ober EA, Andersen DC. Apex Resection in Zebrafish (Danio rerio) as a Model of Heart Regeneration: A Video-Assisted Guide. International Journal of Molecular Sciences. 2021; 22(11):5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115865
Chicago/Turabian StyleEllman, Ditte Gry, Ibrahim Mohamad Slaiman, Sabrina Bech Mathiesen, Kristian Skriver Andersen, Wolfgang Hofmeister, Elke Annette Ober, and Ditte Caroline Andersen. 2021. "Apex Resection in Zebrafish (Danio rerio) as a Model of Heart Regeneration: A Video-Assisted Guide" International Journal of Molecular Sciences 22, no. 11: 5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115865