
Lysophosphatidic Acid Accelerates Bovine In Vitro-Produced Blastocyst Formation through the Hippo/YAP Pathway

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
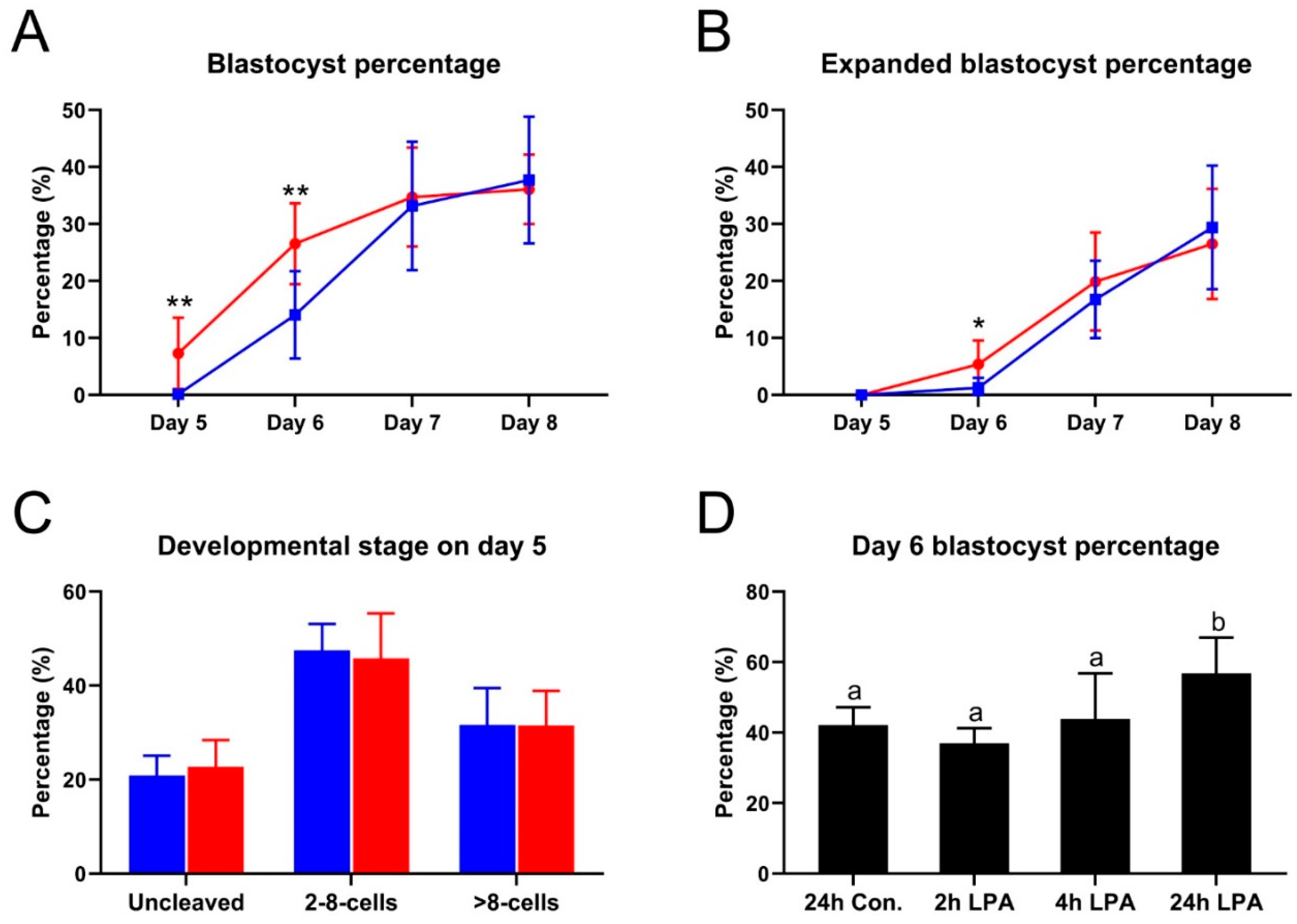
2.1. LPA Accelerates Blastocyst Formation
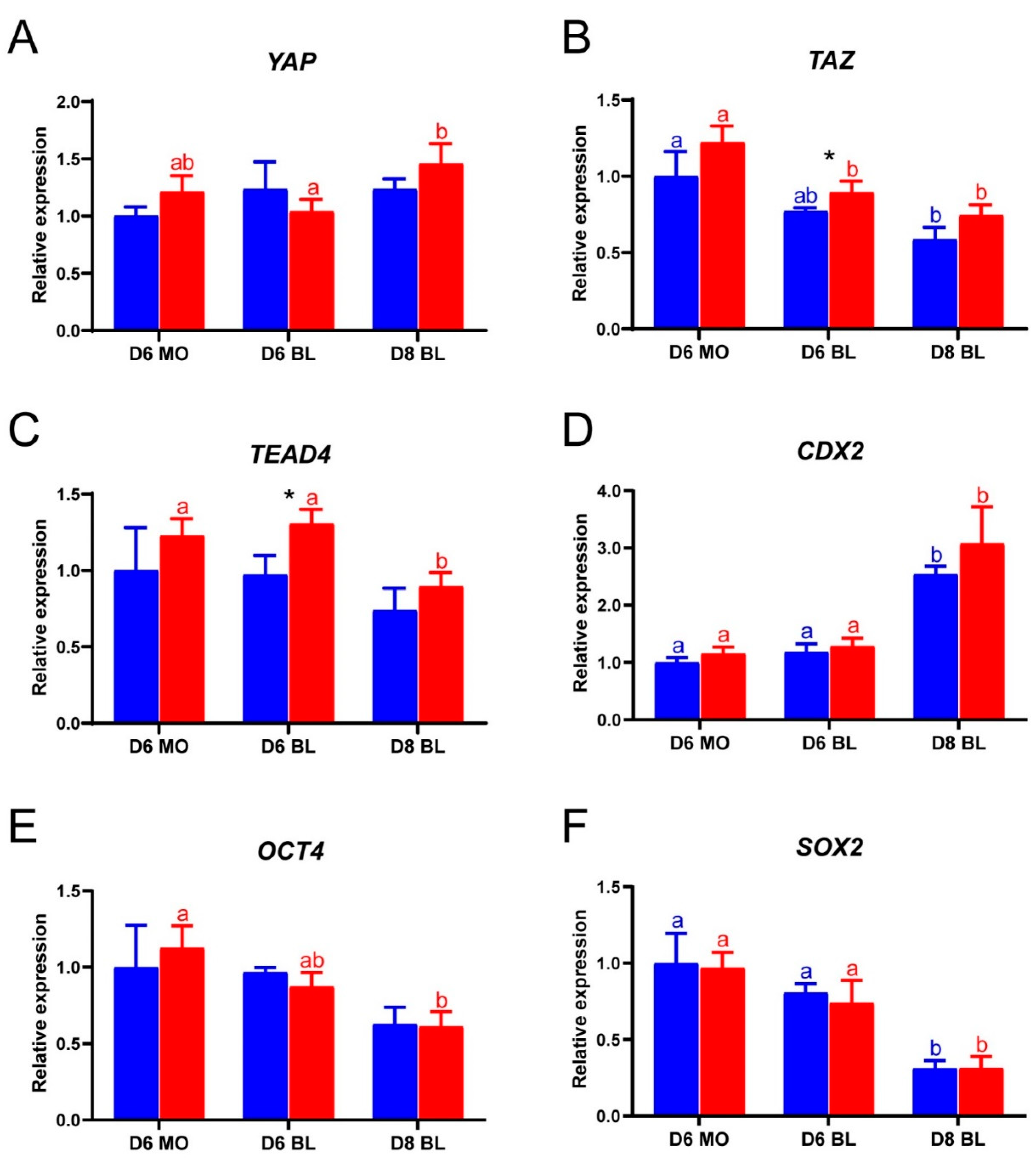
2.2. Gene Expression Levels of LPA-Cultured Embryos
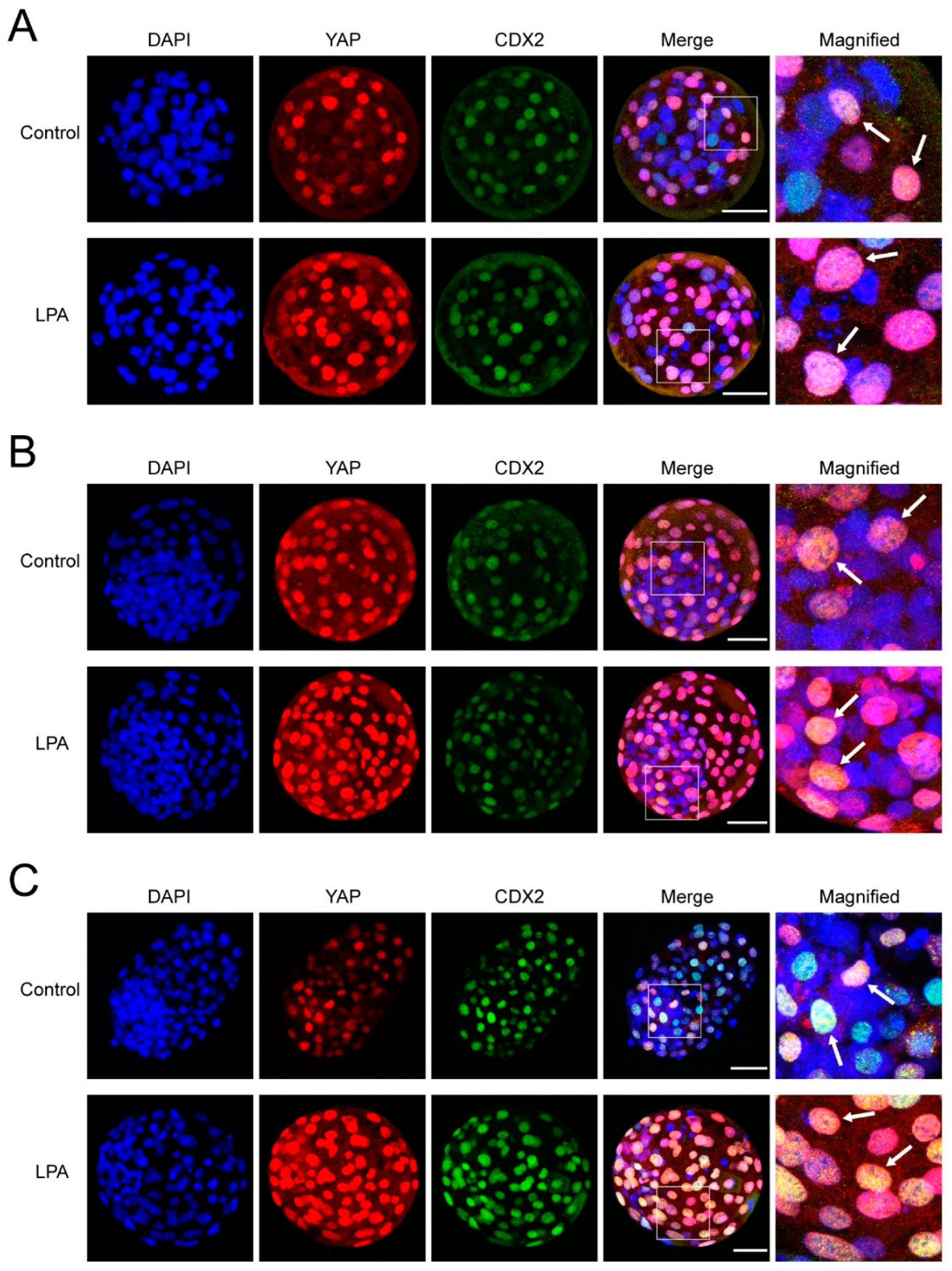
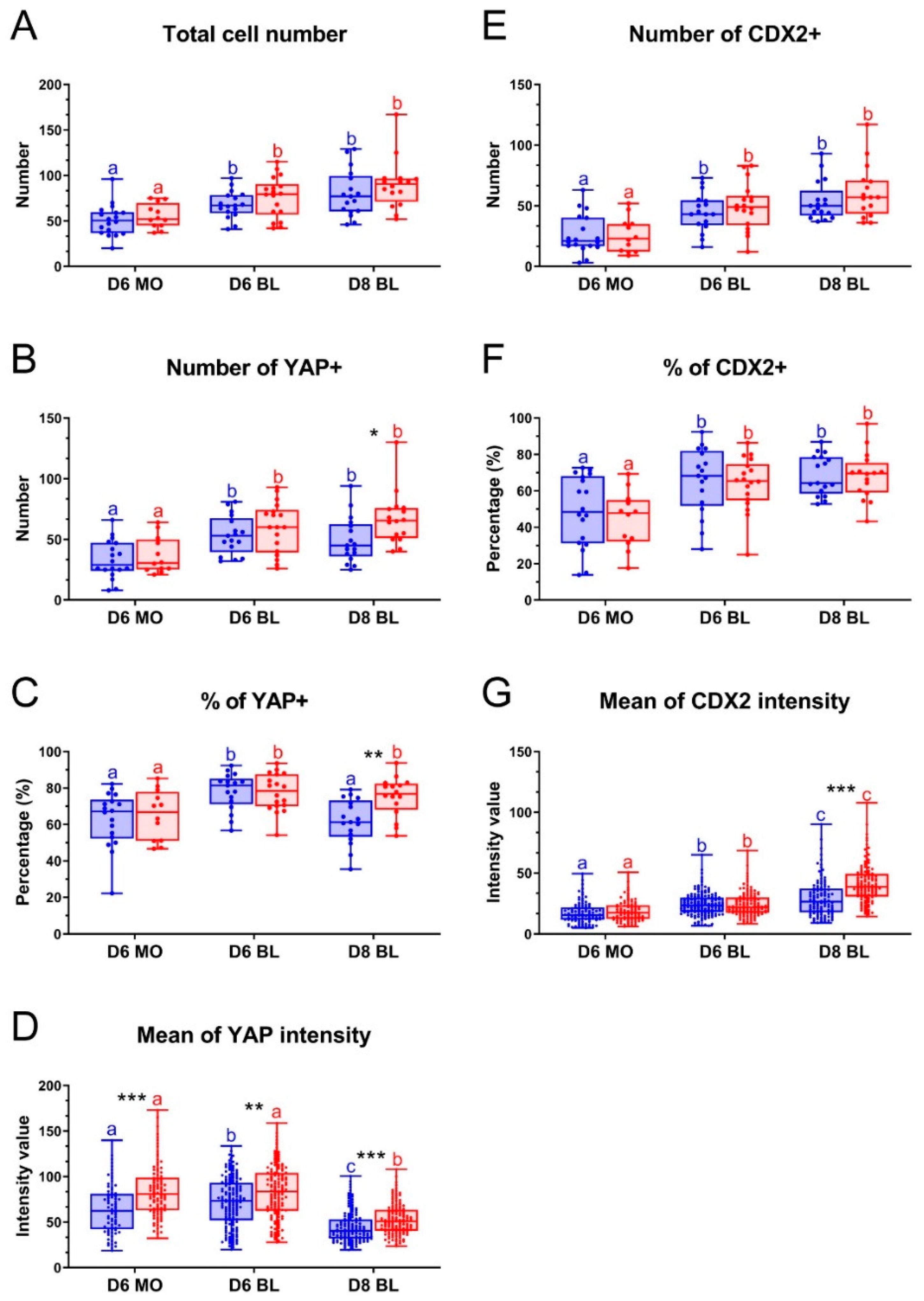
2.3. Effect of LPA on YAP and CDX2 Protein Expression
2.4. Colocalization of YAP and CDX2 in Bovine Embryos
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. In Vitro Embryo Production and LPA Stimulation
4.3. RNA Isolation, cDNA Generation and Quantitative Reverse Transcription-PCR
4.4. Immunofluorescence
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TE | Trophectoderm |
| ICM | Inner cell mass |
| LPA | Lysophosphatidic acid |
| RFU | Relative fluorescence units |
| COCs | Cumulus-oocyte complexes |
| IVF | in vitro fertilization |
| SOF | Synthetic oviductal fluid |
| SCNT | Somatic cell nuclear transfer |
References
- Huang, D.; Guo, G.; Yuan, P.; Ralston, A.; Sun, L.; Huss, M.; Mistri, T.; Pinello, L.; Ng, H.H.; Yuan, G.; et al. The role of Cdx2 as a lineage specific transcriptional repressor for pluripotent network during the first developmental cell lineage segregation. Sci. Rep. 2017, 7, 17156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Watt, K.I.; Harvey, K.F.; Gregorevic, P. Regulation of Tissue Growth by the Mammalian Hippo Signaling Pathway. Front. Physiol. 2017, 8, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Han, H.; Seo, G.; Vargas, R.E.; Yang, B.; Chuc, K.; Zhao, H.; Wang, W. Systematic analysis of the Hippo pathway organization and oncogenic alteration in evolution. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Pan, D. The Hippo Signaling Pathway in Development and Disease. Dev. Cell 2019, 50, 264–282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Tumaneng, K.; Guan, K.-L. The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat. Cell Biol. 2011, 13, 877–883. [Google Scholar] [CrossRef]
- Kawashima, I.; Kawamura, K. Regulation of follicle growth through hormonal factors and mechanical cues mediated by Hippo signaling pathway. Syst. Biol. Reprod. Med. 2018, 64, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef]
- Masciangelo, R.; Hossay, C.; Chiti, M.C.; Manavella, D.D.; Amorim, C.A.; Donnez, J.; Dolmans, M.-M. Role of the PI3K and Hippo pathways in follicle activation after grafting of human ovarian tissue. J. Assist. Reprod. Genet. 2020, 37, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Frum, T.; Murphy, T.M.; Ralston, A. HIPPO signaling resolves embryonic cell fate conflicts during establishment of pluripotency in vivo. eLife 2018, 7. [Google Scholar] [CrossRef]
- Karasek, C.; Ashry, M.; Driscoll, C.S.; Knott, J.G. A tale of two cell-fates: Role of the Hippo signaling pathway and transcription factors in early lineage formation in mouse preimplantation embryos. Mol. Hum. Reprod. 2020, 26, 653–664. [Google Scholar] [CrossRef]
- Bergsmedh, A.; Donohoe, M.E.; Hughes, R.-A.; Hadjantonakis, A.-K. Understanding the Molecular Circuitry of Cell Lineage Specification in the Early Mouse Embryo. Genes 2011, 2, 420–448. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, N.; Inoue, K.-I.; Adachi, K.; Kiyonari, H.; Ota, M.; Ralston, A.; Yabuta, N.; Hirahara, S.; Stephenson, R.O.; Ogonuki, N.; et al. The Hippo Signaling Pathway Components Lats and Yap Pattern Tead4 Activity to Distinguish Mouse Trophectoderm from Inner Cell Mass. Dev. Cell 2009, 16, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Zhang, W.; Shuai, L. The first cell fate decision in pre-implantation mouse embryos. Cell Regen. 2019, 8, 51–57. [Google Scholar] [CrossRef]
- Nishioka, N.; Yamamoto, S.; Kiyonari, H.; Sato, H.; Sawada, A.; Ota, M.; Nakao, K.; Sasaki, H. Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos. Mech. Dev. 2008, 125, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Lorthongpanich, C.; Issaragrisil, S. Emerging Role of the Hippo Signaling Pathway in Position Sensing and Lineage Specification in Mammalian Preimplantation Embryos1. Biol. Reprod. 2015, 92, 143. [Google Scholar] [CrossRef]
- Chen, Y.-A.; Lu, C.-Y.; Cheng, T.-Y.; Pan, S.-H.; Chen, H.-F.; Chang, N.-S. WW Domain-Containing Proteins YAP and TAZ in the Hippo Pathway as Key Regulators in Stemness Maintenance, Tissue Homeostasis, and Tumorigenesis. Front. Oncol. 2019, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, H.; Hejna, M.; Liu, Y.; Percharde, M.; Wossidlo, M.; Blouin, L.; Durruthy-Durruthy, J.; Wong, P.; Qi, Z.; Yu, J.; et al. YAP Induces Human Naive Pluripotency. Cell Rep. 2016, 14, 2301–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisca, F.; Sabbadini, R.A.; Goldshmit, Y.; Pébay, A. Biological Effects of Lysophosphatidic Acid in the Nervous System. Int. Rev. Cell Mol. Biol. 2012, 296, 273–322. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-E.; Herr, D.R.; Chun, J. Lysophosphatidic acid (LPA) receptors: Signaling properties and disease relevance. Prostaglandins Other Lipid Mediat. 2010, 91, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Yung, Y.C.; Chen, A.; Chun, J. Lysophosphatidic acid signalling in development. Development 2015, 142, 1390–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hama, K.; Aoki, J.; Inoue, A.; Endo, T.; Amano, T.; Motoki, R.; Kanai, M.; Ye, X.; Chun, J.; Matsuki, N.; et al. Embryo Spacing and Implantation Timing Are Differentially Regulated by LPA3-Mediated Lysophosphatidic Acid Signaling in Mice1. Biol. Reprod. 2007, 77, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Hama, K.; Contos, J.J.A.; Anliker, B.; Inoue, A.; Skinner, M.K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nat. Cell Biol. 2005, 435, 104–108. [Google Scholar] [CrossRef]
- Yu, F.-X.; Zhao, B.; Panupinthu, N.; Jewell, J.L.; Lian, I.; Wang, L.H.; Zhao, J.; Yuan, H.; Tumaneng, K.; Li, H.; et al. Regulation of the Hippo-YAP Pathway by G-Protein-Coupled Receptor Signaling. Cell 2012, 150, 780–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felley-Bosco, E.; Stahel, R. Hippo/YAP pathway for targeted therapy. Transl. Lung Cancer Res. 2014, 3, 75–83. [Google Scholar]
- Liu, H.; Du, S.; Lei, T.; Wang, H.; He, X.; Tong, R.; Wang, Y. Multifaceted regulation and functions of YAP/TAZ in tumors (Review). Oncol. Rep. 2018, 40, 16–28. [Google Scholar] [CrossRef]
- Moroishi, T.; Park, H.W.; Qin, B.; Chen, Q.; Meng, Z.; Plouffe, S.W.; Taniguchi, K.; Yu, F.-X.; Karin, M.; Pan, D.; et al. A YAP/TAZ-induced feedback mechanism regulates Hippo pathway homeostasis. Genes Dev. 2015, 29, 1271–1284. [Google Scholar] [CrossRef] [Green Version]
- Teo, S.T.; Yung, Y.C.; Herr, D.R.; Chun, J. Lysophosphatidic acid in vascular development and disease. IUBMB Life 2009, 61, 791–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Li, H.Y.; Lee, J.H.; Oh, Y.S.; Jun, H.-S. Lysophosphatidic acid increases mesangial cell proliferation in models of diabetic nephropathy via Rac1/MAPK/KLF5 signaling. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Yamano, S.; Murayama, S.; Ishikawa, H.; Tokumura, A.; Aono, T. Effect of lysophosphatidic acid on the preimplantation development of mouse embryos. FEBS Lett. 1994, 351, 38–40. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Li, L.; Gao, B.; Zhang, D.; Ren, Y.; Zheng, B.; Li, M.; Shi, D.; Huang, B. Early development of porcine parthenogenetic embryos and reduced expression of primed pluripotent marker genes under the effect of lysophosphatidic acid. Reprod. Domest. Anim. 2018, 53, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Gentile, L.; Fuchikami, T.; Sutter, J.; Psathaki, K.; Esteves, T.C.; Araúzo-Bravo, M.J.; Ortmeier, C.; Verberk, G.; Abe, K.; et al. Initiation of trophectoderm lineage specification in mouse embryos is independent of Cdx2. Development 2010, 137, 4159–4169. [Google Scholar] [CrossRef] [Green Version]
- Brinkhof, B.; Van Tol, H.T.A.; Koerkamp, M.J.A.G.; Riemers, F.M.; Ijzer, S.G.; Mashayekhi, K.; Haagsman, H.P.; Roelen, B.A.J. A mRNA landscape of bovine embryos after standard and MAPK-inhibited culture conditions: A comparative analysis. BMC Genom. 2015, 16, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Van Tol, H.T.A.; Stout, T.A.E.; Roelen, B.A.J. Cellular Fragments in the Perivitelline Space Are Not a Predictor of Expanded Blastocyst Quality. Front. Cell Dev. Biol. 2021, 8, 616801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, E.; Guan, H.; Wang, Q.; Zhang, S.; Liu, K.; Yan, F.; Tian, H.; Shan, D.; Xu, H.; et al. Vitamin C treatment of embryos, but not donor cells, improves the cloned embryonic development in sheep. Reprod. Domest. Anim. 2019, 55, 255–265. [Google Scholar] [CrossRef]
- Samiec, M.; Skrzyszowska, M.; Opiela, J. Creation of cloned pig embryos using contact-inhibited or serum-starved fibroblast cells analysed intravitam for apoptosis occurrence/Uzyskiwanie klonalnych zarodków świni z wykorzystaniem komórek fibroblastycznych poddanych inhibicji kontaktowej lub deprywacji troficznej oraz analizowanych przyżyciowo w kierunku apoptozy. Ann. Anim. Sci. 2013, 13, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Olivera, R.; Moro, L.N.; Jordan, R.; Luzzani, C.; Miriuka, S.; Radrizzani, M.; Donadeu, F.X.; Vichera, G. In Vitro and In Vivo Development of Horse Cloned Embryos Generated with iPSCs, Mesenchymal Stromal Cells and Fetal or Adult Fibroblasts as Nuclear Donors. PLoS ONE 2016, 11, e0164049. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, B.; van Tol, H.T.A.; Oei, C.H.Y.; Stout, T.A.E.; Roelen, B.A.J. Lysophosphatidic Acid Accelerates Bovine In Vitro-Produced Blastocyst Formation through the Hippo/YAP Pathway. Int. J. Mol. Sci. 2021, 22, 5915. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115915
Yu B, van Tol HTA, Oei CHY, Stout TAE, Roelen BAJ. Lysophosphatidic Acid Accelerates Bovine In Vitro-Produced Blastocyst Formation through the Hippo/YAP Pathway. International Journal of Molecular Sciences. 2021; 22(11):5915. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115915
Chicago/Turabian StyleYu, Bo, Helena T. A. van Tol, Christine H. Y. Oei, Tom A. E. Stout, and Bernard A. J. Roelen. 2021. "Lysophosphatidic Acid Accelerates Bovine In Vitro-Produced Blastocyst Formation through the Hippo/YAP Pathway" International Journal of Molecular Sciences 22, no. 11: 5915. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115915