
Nuclear Magnetic Resonance Therapy Modulates the miRNA Profile in Human Primary OA Chondrocytes and Antagonizes Inflammation in Tc28/2a Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
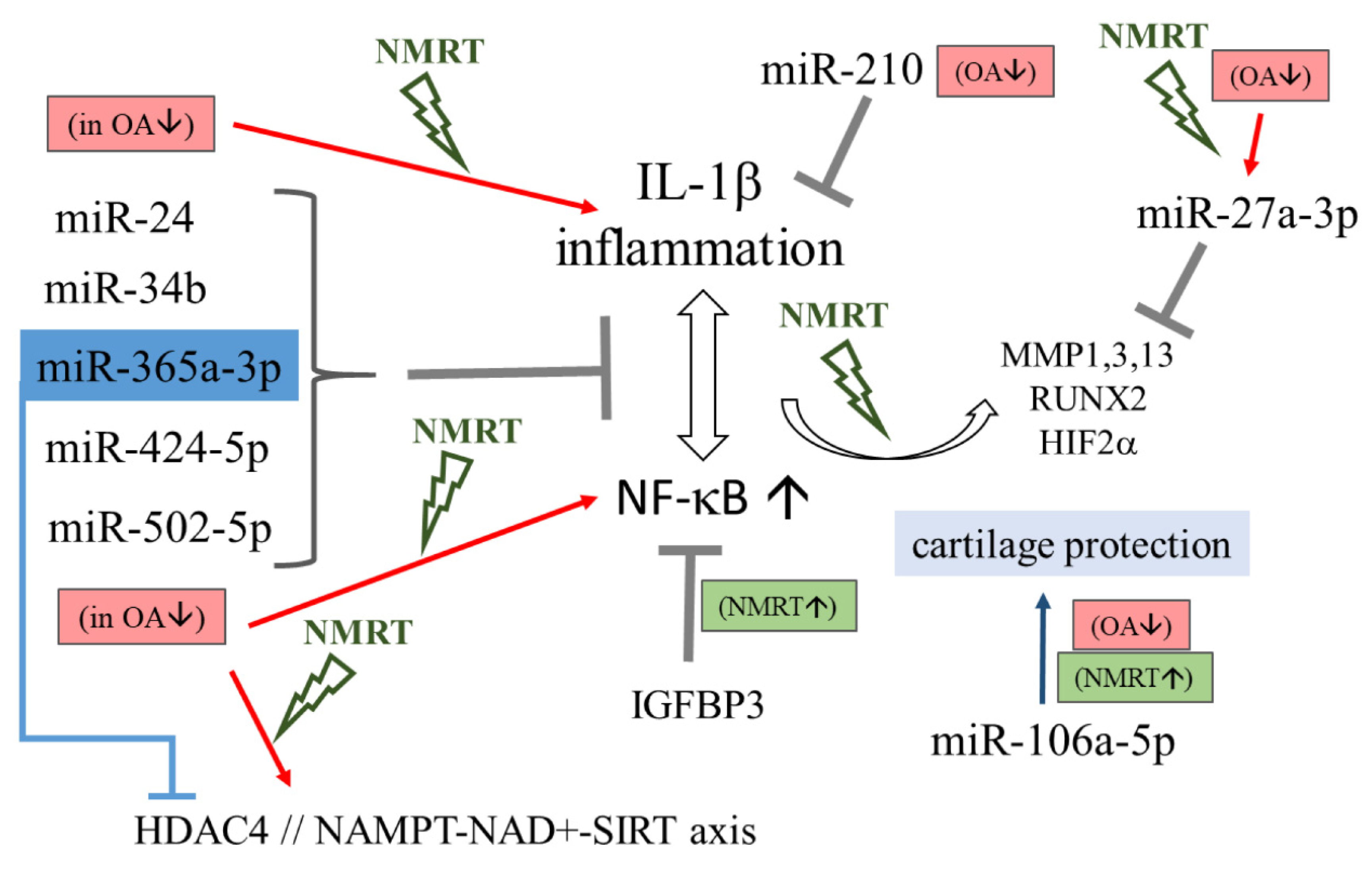
Abstract
:1. Introduction
2. Results
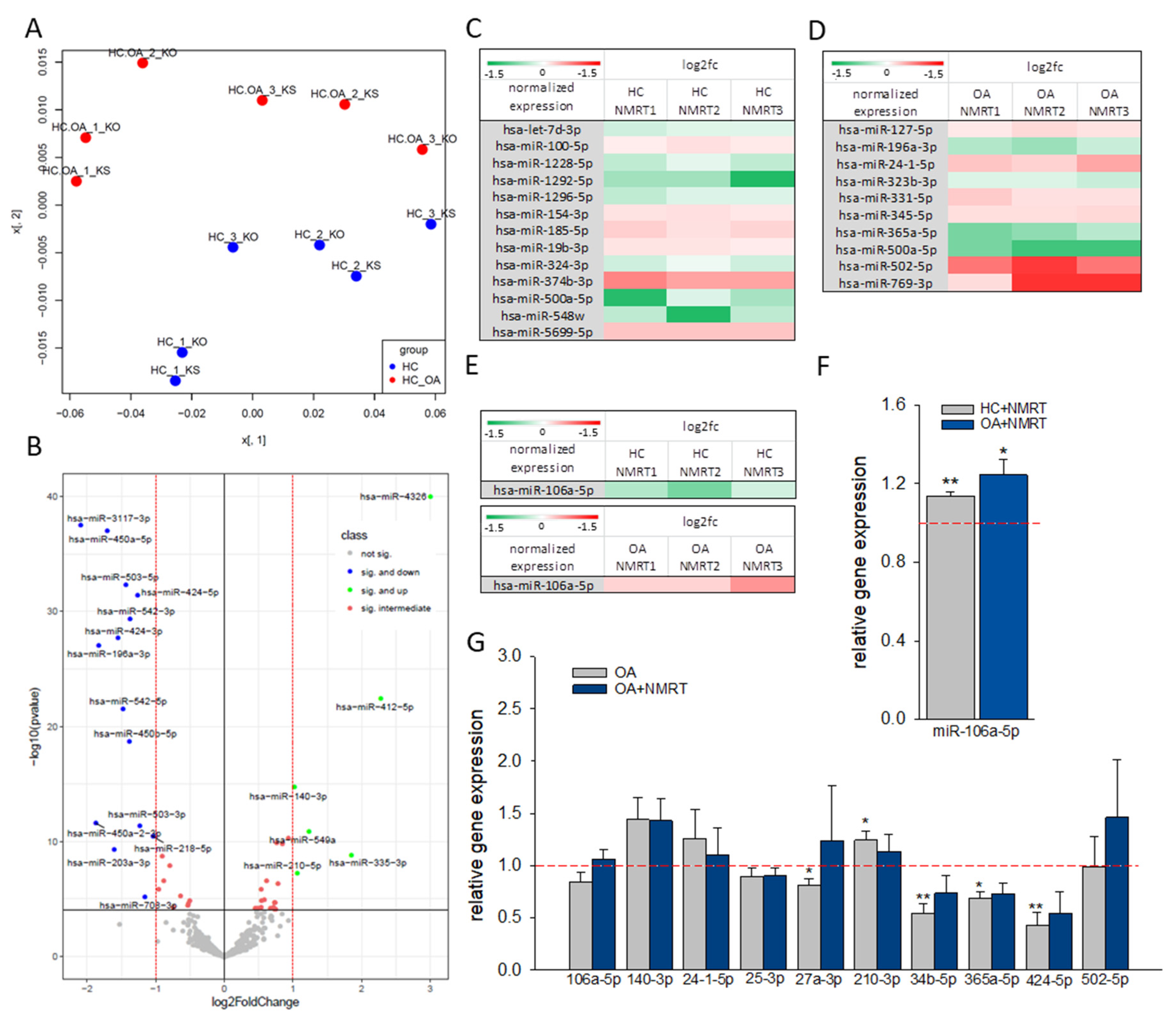
2.1. Differences in miR Expression in OA Cells and the Influence of NMRT
2.2. NMRT Affects Players Involved in miRNA Regulated Signaling
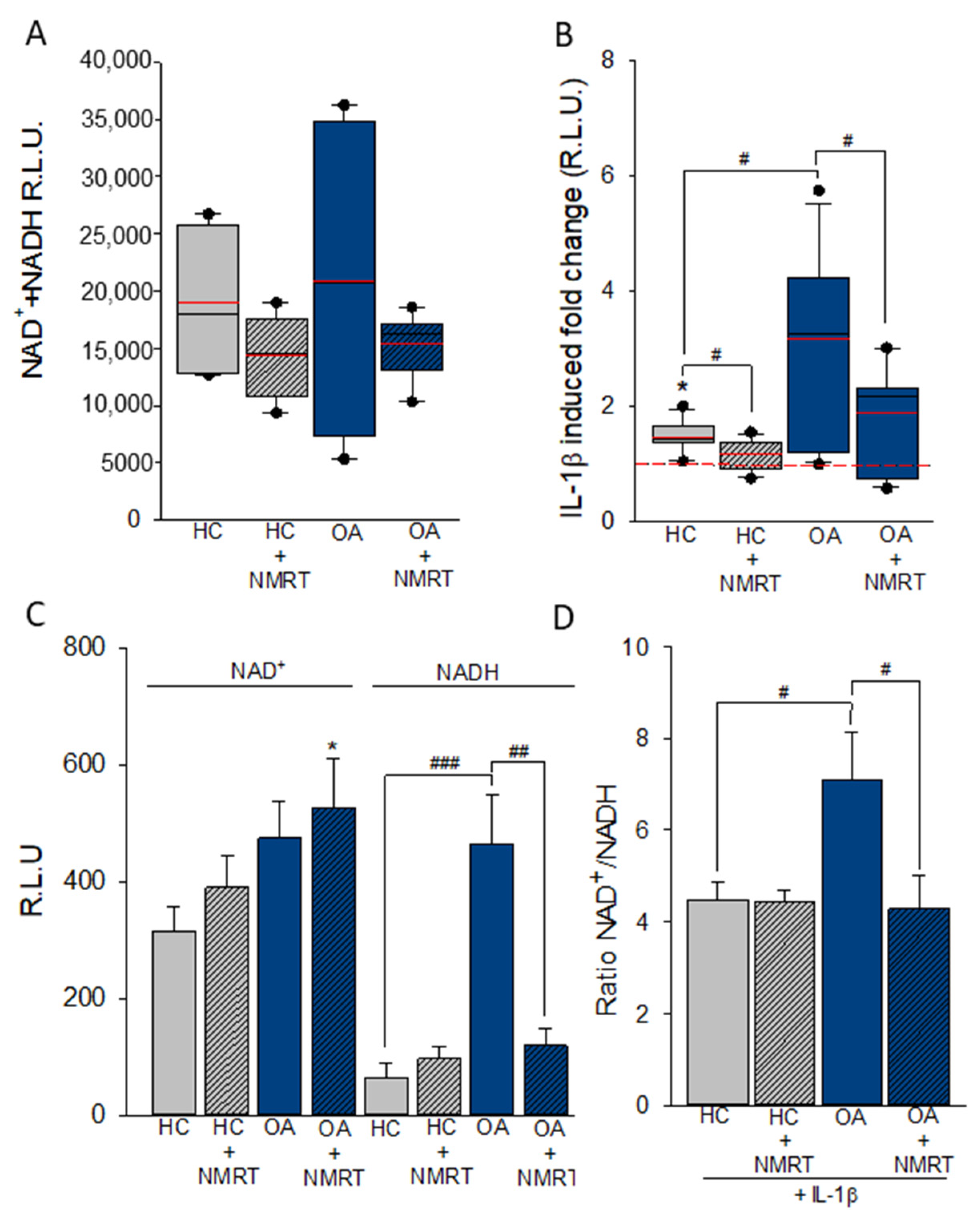
2.3. The Cofactor System NAD+/NADH Is Influenced by NMRT
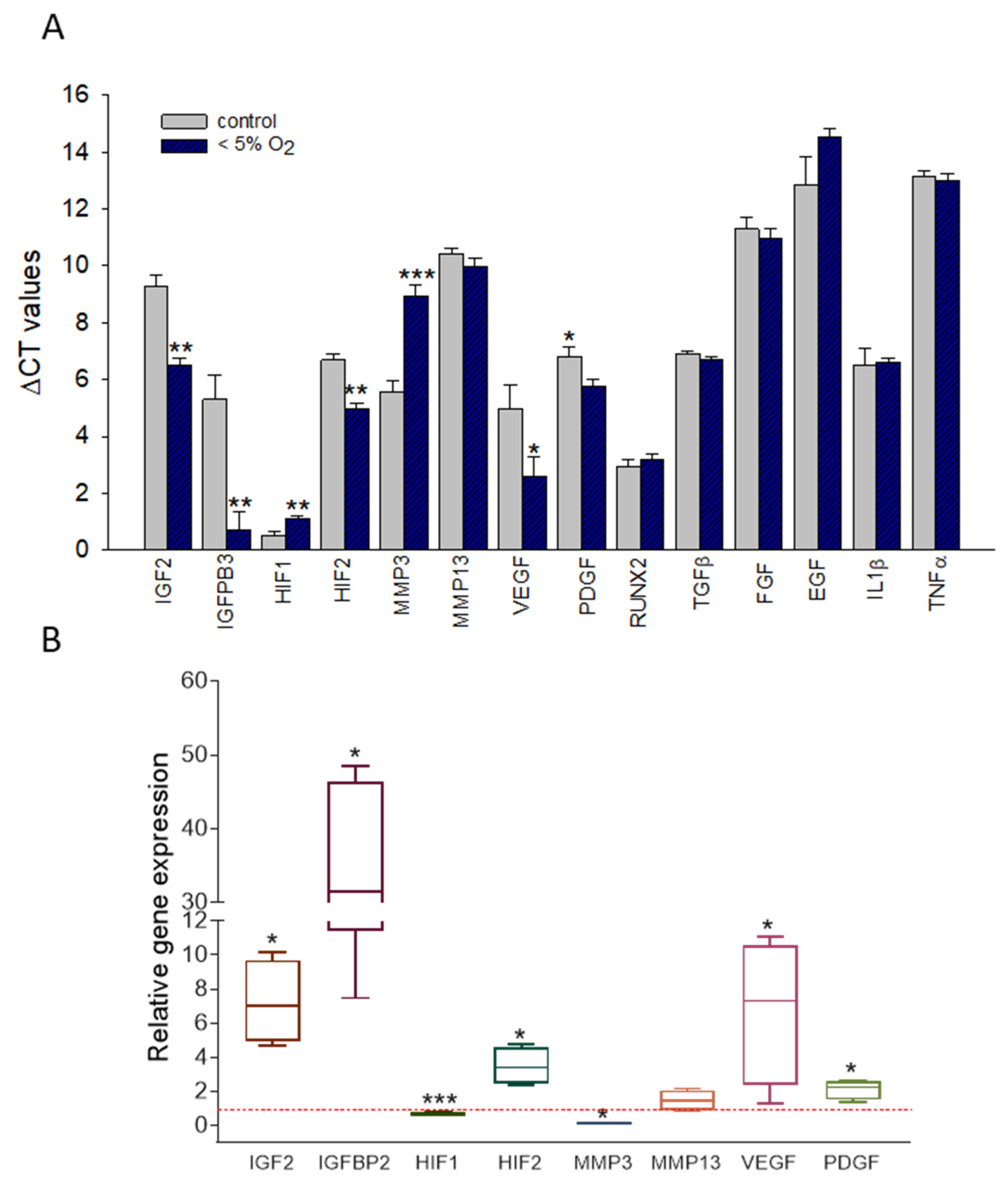
2.4. Hypoxia Induced Changes in the Expression of T/C-28a2 Cells
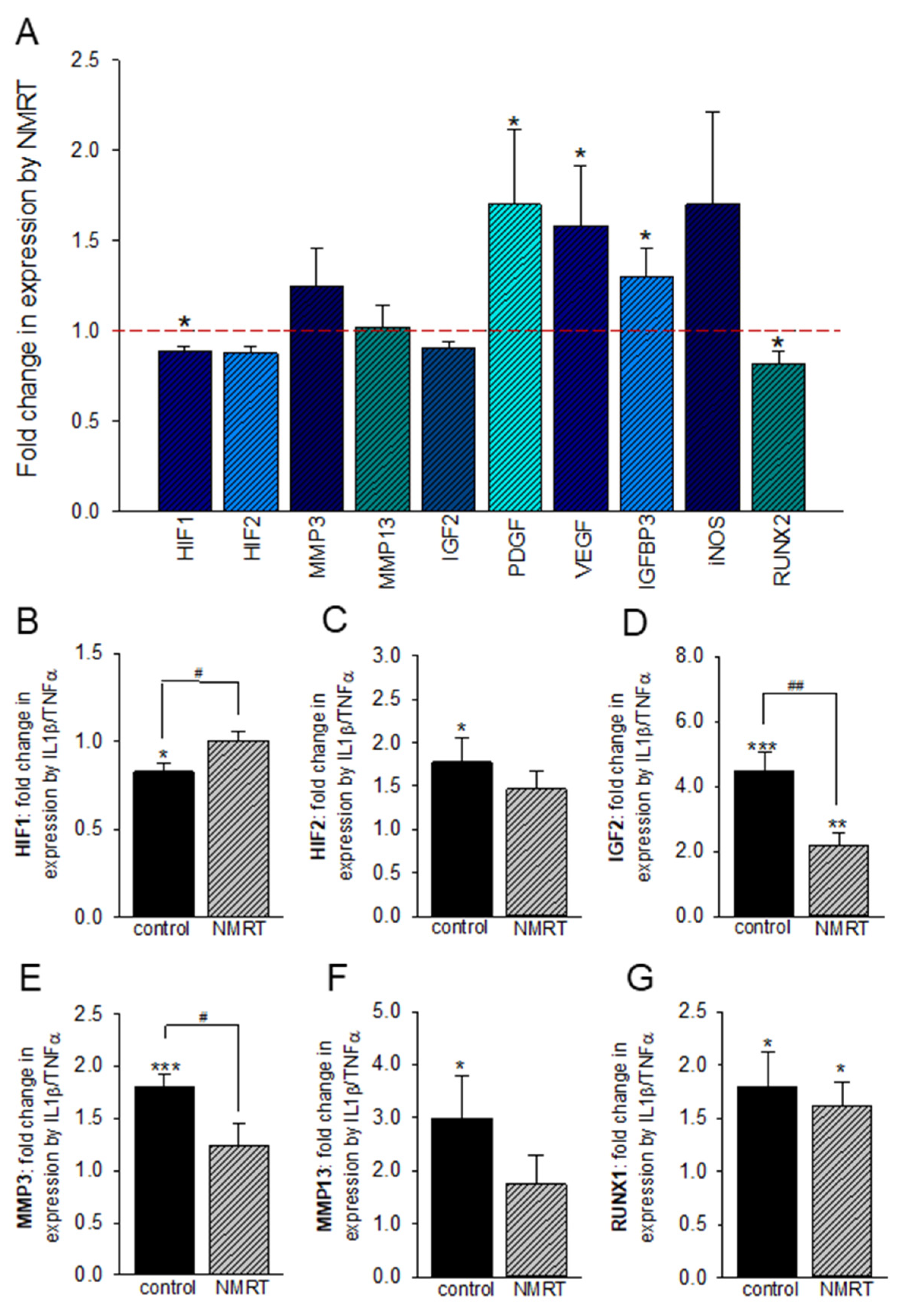
2.5. NMRT Influences IL-1β/TNFα Induced Changes in T/C-28a2 Chondrocytes under Hypoxic Conditions
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Nuclear Magnetic Resonance Therapy (NMRT)
4.3. Cell Culture under Hypoxic Conditions
4.4. miR Sequencing
4.5. miRs Expression
4.6. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.7. Western Blot Analysis
4.8. HDAC I/II Activity Measurement
4.9. NAD/NADH Detection
4.10. Annexin/PI Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levers, A.; Staat, M.; Van Laack, W. Analyse der Langzeitwirkung der MBST Kernspinresonanz-Therapie bei Gonarthrose. Orthopädische Praxis 2011, 47, 536–543. [Google Scholar]
- Kullich, W.; Schwann, H.; Machreich, K.; Ausserwinkler, M. Additional outcome improvement in the rehabilitation of chronic low back pain after nuclear resonance therapy. Rheumatologia 2006, 20, 7–12. [Google Scholar]
- Kullich, W.; Ausserwinkler, M. Functional improvement in finger joint osteoarthritis with therapeutic use of nuclear magnetic resonance. Orthopädische Praxis 2008, 6, 287–290. [Google Scholar]
- Digel, I.; Kurulgan, E.; Linder, P.; Kayser, P.; Porst, D.; Braem, G.J.; Zerlin, K.; Artmann, G.M.; Temiz Artmann, A. Decrease in extracellular collagen crosslinking after NMR magnetic field application in skin fibroblasts. Med. Biol. Eng. Comput. 2007, 45, 91–97. [Google Scholar] [CrossRef]
- Steinecker-Frohnwieser, B.; Kullich, W.; Mann, A.; Kress, H.G.; Weigl, L. The therapeutic nuclear magnetic resonance changes the balance in intracellular calcium and reduces the interleukin-1β induced increase of NF-κB activity in chondrocytes. Clin. Exp. Rheumatol. 2018, 36, 294–301. [Google Scholar]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiologica 2017, 219, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Sondag, G.R.; Haqqi, T.M. The role of microRNAs and their targets in osteoarthritis. Curr. Rheumatol. Rep. 2016, 18, 56. [Google Scholar] [CrossRef]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB signaling pathways in osteoarthritic cartilage destruction. Cells 2019, 8, E734. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.M.; Haqqi, T.M. Epigenetics in osteoarthritis: Potential of HDAC inhibitors as therapeutics. Pharmacol. Res. 2018, 128, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Peffers, M.J.; Balaskas, P.; Smagul, A. Osteoarthritis year in review 2017: Genetics and epigenetics. Osteoarthr. Cartil. 2018, 26, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.; Kwak, J.-S.; Yang, S.; Gong, M.-K.; Kim, J.-H.; Rhee, J.; Kim, S.K.; Kim, H.-E.; Ryu, J.-H.; Chun, J.-S. Reciprocal regulation by hypoxia-inducible factor-2α and the NAMPT-NAD(+)-SIRT axis in articular chondrocytes is involved in osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2288–2296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.J.; Luo, W.; Lei, G.H. Role of HIF-1α and HIF-2α in osteoarthritis. Jt. Bone Spine 2015, 82, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Gao, J.; Si, Y.; Zhao, D. Combination of circulating miR-19b-3p, miR-122-5p and miR-486-5p expressions correlates with risk and disease severity of knee osteoarthritis. Am. J. Transl. Res. 2017, 9, 2852–2864. [Google Scholar]
- An, Y.; Wan, G.; Tao, J.; Cui, M.; Zhou, Q.; Hou, W. Down-regulation of microRNA-203a suppresses IL-1β-induced inflammation and cartilage degradation in human chondrocytes through Smad3 signaling. Biosci. Rep. 2020, 40, BSR20192723. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-H.; Liu, W.; Zhang, L.; Liu, X.-Y.; Wang, Y.; Xue, B.; Liu, B.; Duan, R.; Zhang, B.; Yang, J. Effects of microRNA-24 targeting C-myc on apoptosis, proliferation, and cytokine expressions in chondrocytes of rats with osteoarthritis via MAPK signaling pathway. J. Cell. Biochem. 2018, 119, 7944–7958. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.-J.; Yang, X.; Wei, L.; Chen, Q. MiR-365: A mechanosensitive microRNA stimulates chondrocyte differentiation through targeting histone deacetylase 4. FASEB J. 2011, 25, 4457–4466. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.S.; Park, S.J.; Lee, M.H.; Kim, H.A. MicroRNA-365 regulates IL-1β-induced catabolic factor expression by targeting HIF-2α in primary chondrocytes. Sci. Rep. 2017, 7, 17889. [Google Scholar] [CrossRef] [Green Version]
- Ntoumou, E.; Tzetis, M.; Braoudaki, M.; Lambrou, G.; Poulou, M.; Malizos, K.; Stefanou, N.; Anastasopoulou, L.; Tsezou, A. Serum microRNA array analysis identifies miR-140-3p, miR-33b-3p and miR-671-3p as potential osteoarthritis biomarkers involved in metabolic processes. Clin. Epigenetics 2017, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Sun, Y.; Wang, Y.; Liu, R.; Bao, Y.; Li, Q. MiR-502-5p inhibits IL-1β-induced chondrocyte injury by targeting TRAF2. Cell. Immunol. 2016, 302, 50–57. [Google Scholar] [CrossRef]
- Gökşen, N.; Çaliş, M.; Doğan, S.; Çaliş, H.T.; Özgöçmen, S. Magnetic resonance therapy for knee osteoarthritis: A randomized, double blind placebo controlled trial. Eur. J. Phys. Rehabil. Med. 2016, 52, 431–439. [Google Scholar]
- Zhang, D.; Cao, X.; Li, J.; Zhao, G. MiR-210 inhibits NF-κB signaling pathway by targeting DR6 in osteoarthritis. Sci. Rep. 2015, 5, 12775. [Google Scholar] [CrossRef]
- Zhang, W.; Hsu, P.; Zhong, B.; Guo, S.; Zhang, C.; Wang, Y.; Luo, C.; Zhan, Y.; Zhang, S. MiR-34a enhances chondrocyte apoptosis, senescence and facilitates development of osteoarthritis by targeting DLL1 and regulating PI3K/AKT pathway. Cell. Physiol. Biochem. 2018, 48, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Li, Z.; Abdalla, B.A.; Chen, Y.; Nie, Q. MiR-34b-5p mediates the proliferation and differentiation of myoblasts by targeting IGFBP2. Cells 2019, 8, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travascio, F.; Elmasry, S.; Asfour, S. Modeling the role of IGF-1 on extracellular matrix biosynthesis and cellularity in intervertebral disc. J. Biomech. 2014, 47, 2269–2276. [Google Scholar] [CrossRef]
- Zhang, X.L.; Li, H.H.; Cao, Y.P.; Peng, F.; Li, J.P. Insulin-like growth factor binding protein 3 inhibits inflammatory response and promotes apoptosis in fibroblast-like synoviocytes of osteoarthritis. Int. J. Clin. Exp. Pathol. 2017, 10, 3024–3032. [Google Scholar]
- Jang, H.; Park, S.; Kim, J.; Kim, J.H.; Kim, S.-Y.; Cho, S.; Park, S.G.; Park, B.C.; Kim, S.; Kim, J.-H. The tumor suppressor, p53, negatively regulates non-canonical NF-κB signaling through miRNA-induced silencing of NF-κB-inducing kinase. Mol. Cells 2020, 43, 23–33. [Google Scholar] [PubMed]
- Ji, Q.; Qi, D.; Xu, X.; Xu, Y.; Goodman, S.B.; Kang, L.; Song, Q.; Fan, Z.; Maloney, W.J.; Wang, Y. Cryptotanshinone protects cartilage against developing osteoarthritis through the miR-106a-5p/GLIS3 axis. Mol. Ther. Nucleic Acids 2018, 11, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Guan, Y.; Tian, S.; Wang, Y.; Sun, K.; Chen, Q. Mechanical and IL-1β responsive miR-365 contributes to osteoarthritis development by targeting histone deacetylase 4. Int. J. Mol. Sci. 2016, 17, 436. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, N.; Rasheed, Z.; Ramamurthy, S.; Anbazhagan, A.N.; Voss, F.R.; Haqqi, T.M. MicroRNA-27b regulates the expression of matrix metalloproteinase 13 in human osteoarthritis chondrocytes. Arthritis Rheumatol. 2010, 62, 1361–1371. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, L.; Wang, Q.; Huang, J.; Xu, S. LncRNA FOXD2-AS1 induces chondrocyte proliferation through sponging miR-27a-3p in osteoarthritis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1241–1247. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Qin, G.; Zhao, T.C. HDAC4: Mechanism of regulation and biological functions. Epigenomics 2014, 6, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Sun, Y.; Ge, Q.; Teng, H.; Jiang, Q. Histone deacetylase 4 alters cartilage homeostasis in human osteoarthritis. BMC Musculoskelet. Disord. 2014, 15, 438. [Google Scholar] [CrossRef] [Green Version]
- Markway, B.D.; Cho, H.; Johnstone, B. Hypoxia promotes redifferentiation and suppresses markers of hypertrophy and degeneration in both healthy and osteoarthritic chondrocytes. Arthritis Res. Ther. 2013, 15, R92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, M.B.; Birkhead, J.R.; Suen, L.F.; Yamin, R.; Mizuno, S. Interleukin-1 beta-modulated gene expression in immortalized human chondrocytes. J. Clin. Investig. 1994, 94, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
- Finger, F.; Schoerle, C.; Zien, A.; Gebhard, P.; Goldring, M.B. Molecular phenotyping of human chondrocyte cell lines T/C-28a2, T/C-28a4, and C-28/I2. Arthritis Rheumatol. 2003, 48, 3395–3403. [Google Scholar] [CrossRef]
- Goldring, M.B. Culture of immortalized chondrocytes and their use as models of chondrocyte function. Methods Mol. Med. 2004, 100, 37–52. [Google Scholar]
- Sun, Z.; Evans, J.; Bhagwate, A.; Middha, S.; Bockol, M.; Yan, H.; Kocher, J.P. CAP-miRSeq: A comprehensive analysis pipeline for microRNA sequencing data. BMC Genom. 2014, 15, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Li, W. Volcano plots in analyzing differential expressions with mRNA microarrays. J. Bioinform. Comput. Biol. 2012, 10, 1231003. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinecker-Frohnwieser, B.; Lohberger, B.; Eck, N.; Mann, A.; Kratschmann, C.; Leithner, A.; Kullich, W.; Weigl, L. Nuclear Magnetic Resonance Therapy Modulates the miRNA Profile in Human Primary OA Chondrocytes and Antagonizes Inflammation in Tc28/2a Cells. Int. J. Mol. Sci. 2021, 22, 5959. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115959
Steinecker-Frohnwieser B, Lohberger B, Eck N, Mann A, Kratschmann C, Leithner A, Kullich W, Weigl L. Nuclear Magnetic Resonance Therapy Modulates the miRNA Profile in Human Primary OA Chondrocytes and Antagonizes Inflammation in Tc28/2a Cells. International Journal of Molecular Sciences. 2021; 22(11):5959. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115959
Chicago/Turabian StyleSteinecker-Frohnwieser, Bibiane, Birgit Lohberger, Nicole Eck, Anda Mann, Cornelia Kratschmann, Andreas Leithner, Werner Kullich, and Lukas Weigl. 2021. "Nuclear Magnetic Resonance Therapy Modulates the miRNA Profile in Human Primary OA Chondrocytes and Antagonizes Inflammation in Tc28/2a Cells" International Journal of Molecular Sciences 22, no. 11: 5959. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115959