
Effect of Inducible BMP-7 Expression on the Osteogenic Differentiation of Human Dental Pulp Stem Cells


Abstract
:1. Introduction
2. Results
2.1. Doxycycline-Regulated Gene Expression
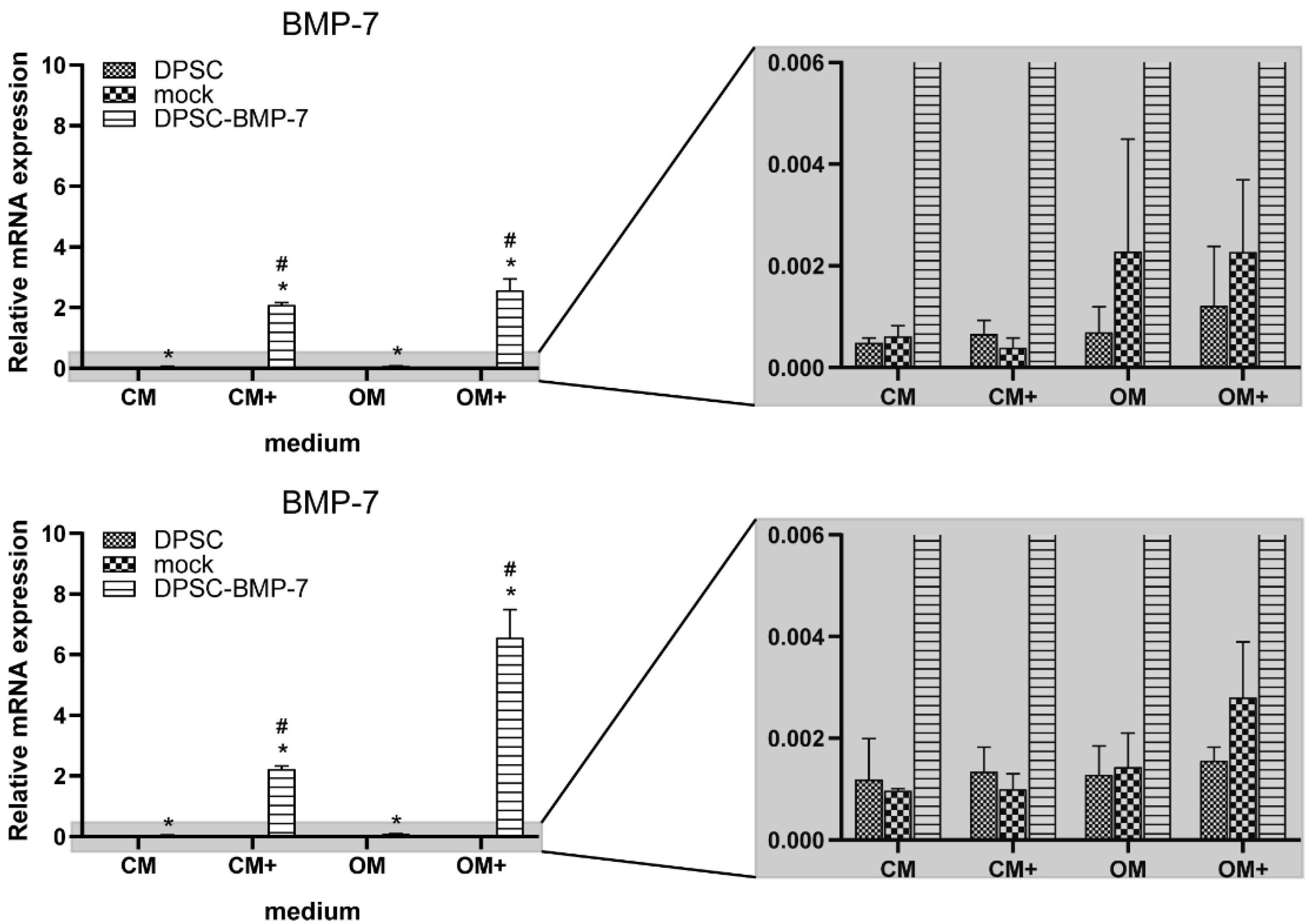
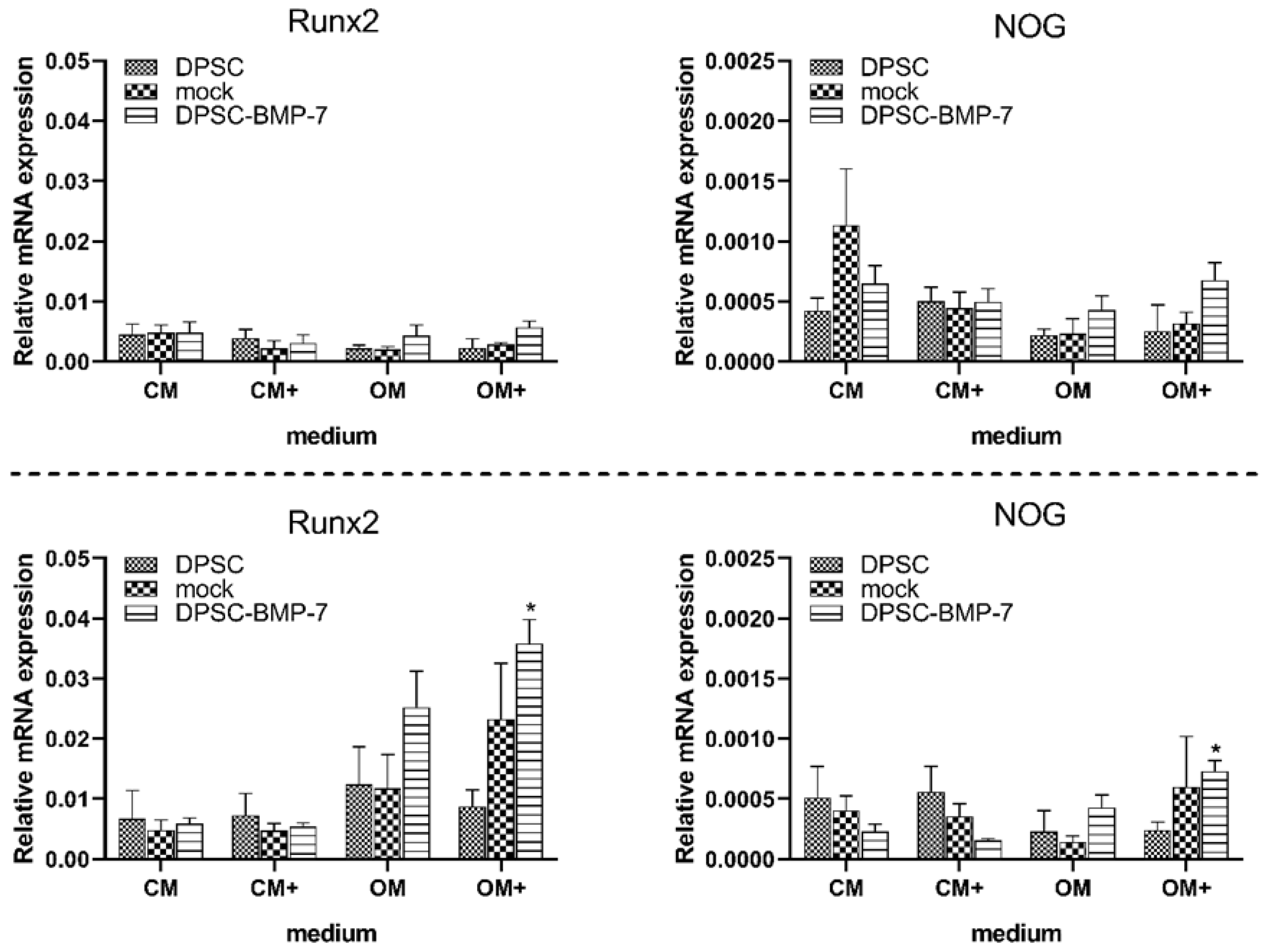
2.2. Quantitative Real-Time PCR Assays
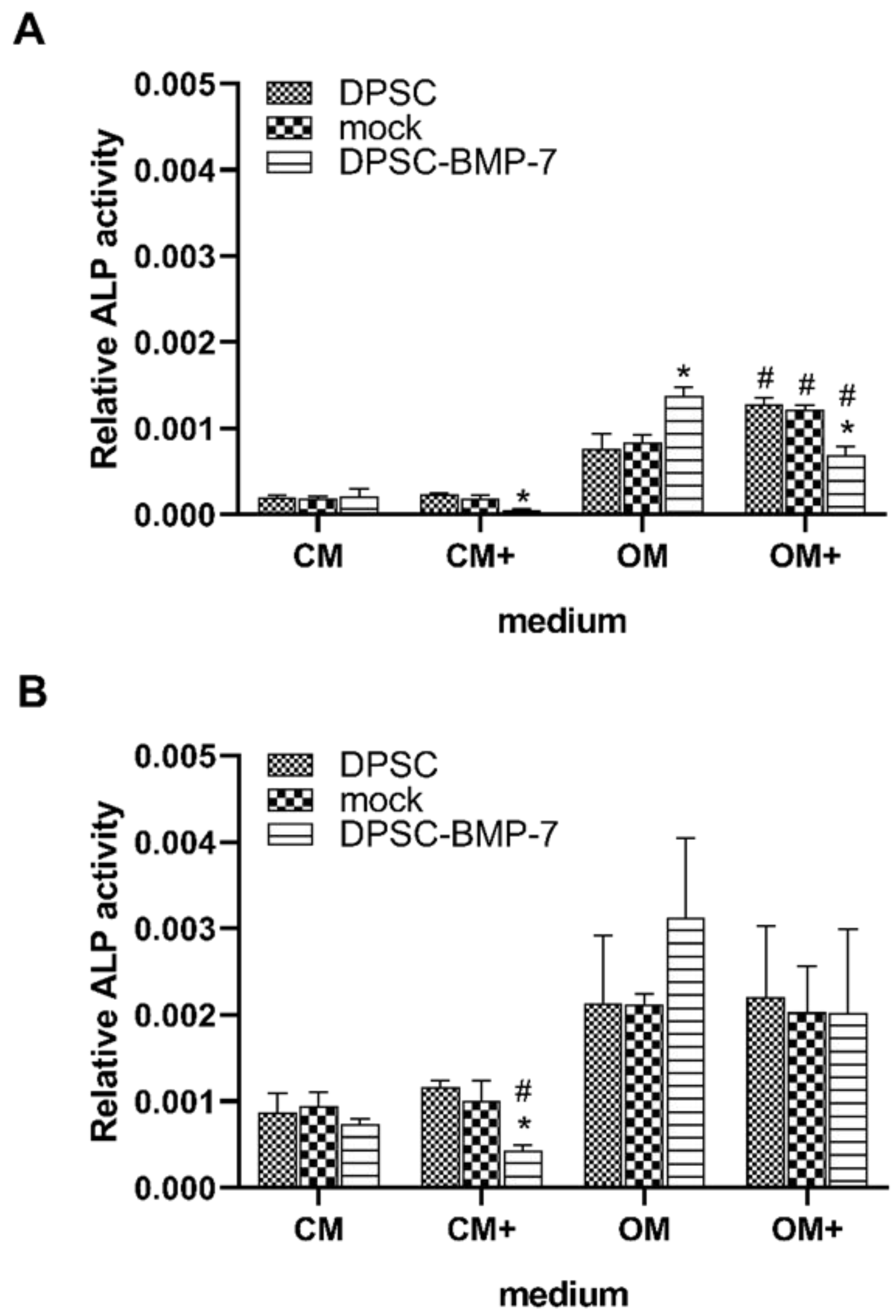
2.3. Alkaline Phosphatase Assay
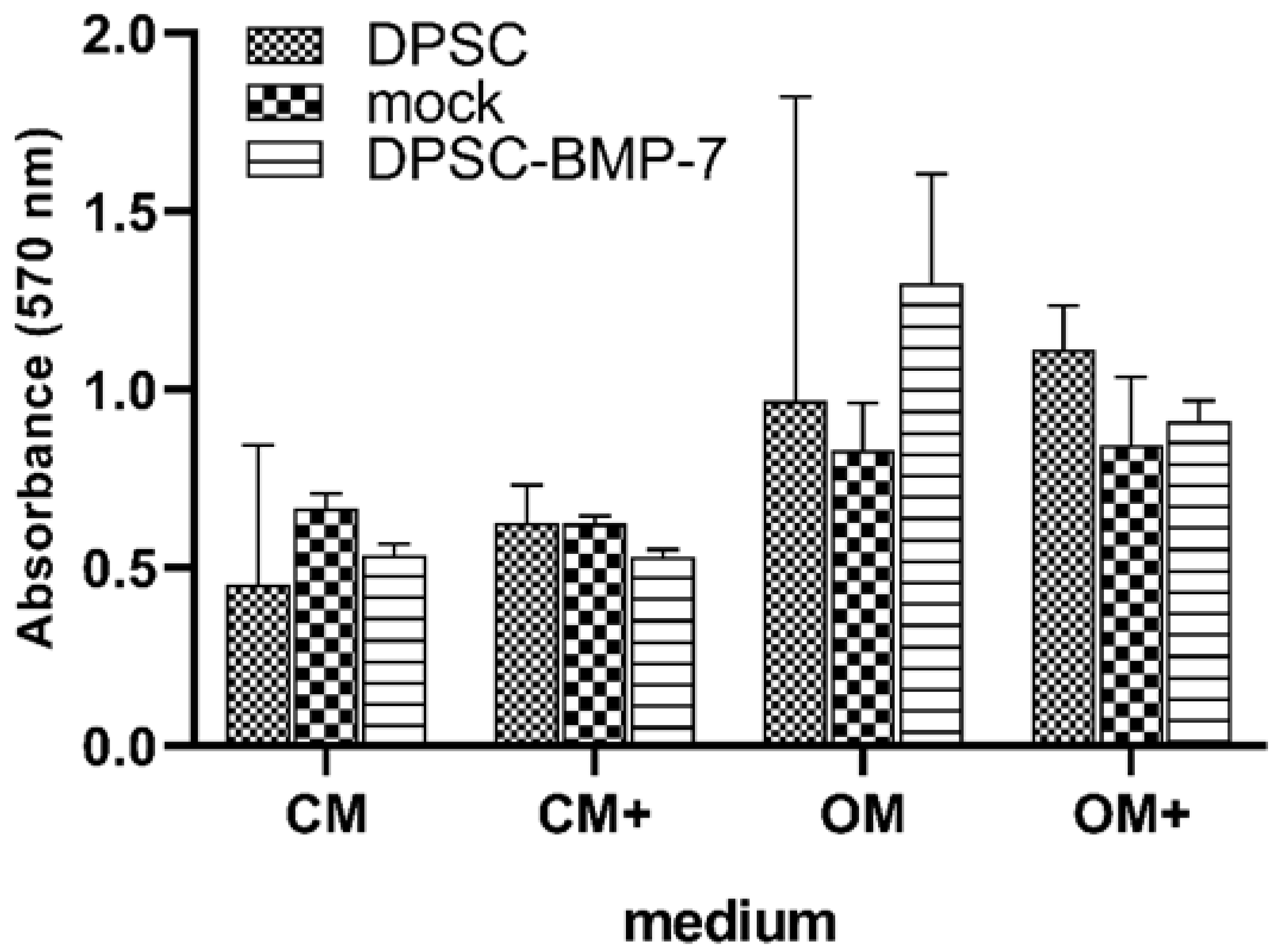
2.4. Mineralization Assay
3. Discussion
4. Materials and Methods
4.1. Construction of pTet-IRES-EGFP-BMP-7 Plasmid
4.2. Cell Culture
4.3. Lentivirus Preparation and Transduction
4.4. Western Blot Analysis
4.5. BMP-7 ELISA
4.6. Real-Time Quantitative PCR
4.7. Detection of Alkaline Phosphatase (ALP) Activity
4.8. Mineralization Assay
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mao, A.S.; Mooney, D.J. Regenerative medicine: Current therapies and future directions. Proc. Natl. Acad. Sci. USA 2015, 112, 14452–14459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amini, A.R.; Laurencin, C.T.; Nukavarapu, S.P. Bone tissue engineering: Recent advances and challenges. Crit. Rev. Biomed Eng. 2012, 40, 363–408. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, M.T.P.; Valera, M.C.; Nakashima, M.; Nör, J.E.; Bottino, M.C. Tissue-engineering-based strategies for regenerative endodontics. J. Dent. Res. 2014, 93, 1222–1231. [Google Scholar] [CrossRef]
- Fisher, J.N.; Peretti, G.M.; Scotti, C. Stem cells for bone regeneration: From cell-based therapies to decellularised engineered extracellular matrices. Stem. Cells Int. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.B.; Mauck, R.L. Tissue engineering and regenerative medicine: Recent innovations and the transition to translation. Tissue Eng. Part B Rev. 2013, 19, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, A.R.; Carrión, F.S.; Chaparro, A.P. Mesenchymal stem cells from the oral cavity and their potential value in tissue engineering. Periodontol 2000 2015, 67, 251–267. [Google Scholar] [CrossRef]
- Yang, J.W.; Shin, Y.Y.; Seo, Y.; Kim, H.-S. Therapeutic functions of stem cells from oral cavity: An update. Int. J. Mol. Sci. 2020, 21, 4389. [Google Scholar] [CrossRef] [PubMed]
- Leyendecker Junior, A.; Gomes Pinheiro, C.C.; Lazzaretti Fernandes, T.; Franco Bueno, D. The use of human dental pulp stem cells for in vivo bone tissue engineering: A systematic review. J. Tissue Eng. 2018, 9, 2041731417752766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Monjaraz, B.; Santiago-Osorio, E.; Monroy-García, A.; Ledesma-Martínez, E.; Mendoza-Núñez, V.M. Mesenchymal stem cells of dental origin for inducing tissue regeneration in periodontitis: A mini-review. Int. J. Mol. Sci. 2018, 19, 944. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Chen, J.; Liu, S.; Jin, Y. Stem cell-based bone and dental regeneration: A view of microenvironmental modulation. Int. J. Oral Sci. 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Di Vito, A.; Chiarella, E.; Baudi, F.; Scardamaglia, P.; Antonelli, A.; Giudice, D.; Barni, T.; Fortunato, L.; Giudice, A. Dose-dependent effects of zoledronic acid on human periodontal ligament stem cells: An in vitro pilot study. Cell Transpl. 2020, 29, 0963689720948497. [Google Scholar] [CrossRef] [PubMed]
- Hiraki, T.; Kunimatsu, R.; Nakajima, K.; Abe, T.; Yamada, S.; Rikitake, K.; Tanimoto, K. Stem cell-derived conditioned media from human exfoliated deciduous teeth promote bone regeneration. Oral Dis. 2020, 26, 381–390. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerényi, F.; Tarapcsák, S.; Hrubi, E.; Baráthne, S.Á.; Hegedüs, V.; Balogh, S.; Bágyi, K.; Varga, G.; Hegedüs, C. Comparison of sorting of fluorescently and magnetically labelled dental pulp stem cells. Fogorv. Szle. 2016, 109, 29–33. [Google Scholar]
- Takeda, T.; Tezuka, Y.; Horiuchi, M.; Hosono, K.; Iida, K.; Hatakeyama, D.; Miyaki, S.; Kunisada, T.; Shibata, T.; Tezuka, K. Characterization of dental pulp stem cells of human tooth germs. J. Dent. Res. 2008, 87, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Couble, M.L.; Farges, J.C.; Bleicher, F.; Perrat-Mabillon, B.; Boudeulle, M.; Magloire, H. Odontoblast differentiation of human dental pulp cells in explant cultures. Calcif. Tissue Int. 2000, 66, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Kadar, K.; Kiraly, M.; Porcsalmy, B.; Molnar, B.; Racz, G.Z.; Blazsek, J.; Kallo, K.; Szabo, E.L.; Gera, I.; Gerber, G.; et al. Differentiation potential of stem cells from human dental origin-promise for tissue engineering. J. Physiol. Pharmacol. 2009, 60 (Suppl 7), 167–175. [Google Scholar] [PubMed]
- Sui, B.D.; Hu, C.H.; Liu, A.Q.; Zheng, C.X.; Xuan, K.; Jin, Y. Stem cell-based bone regeneration in diseased microenvironments: Challenges and solutions. Biomaterials 2019, 196, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Tamaoki, N.; Takahashi, K.; Tanaka, T.; Ichisaka, T.; Aoki, H.; Takeda-Kawaguchi, T.; Iida, K.; Kunisada, T.; Shibata, T.; Yamanaka, S.; et al. Dental pulp cells for induced pluripotent stem cell banking. J. Dent. Res. 2010, 89, 773–778. [Google Scholar] [CrossRef]
- Takeda-Kawaguchi, T.; Sugiyama, K.; Chikusa, S.; Iida, K.; Aoki, H.; Tamaoki, N.; Hatakeyama, D.; Kunisada, T.; Shibata, T.; Fusaki, N.; et al. Derivation of ipscs after culture of human dental pulp cells under defined conditions. PLoS ONE 2014, 9, e115392. [Google Scholar] [CrossRef] [PubMed]
- d’Aquino, R.; Papaccio, G.; Laino, G.; Graziano, A. Dental pulp stem cells: A promising tool for bone regeneration. Stem Cell Rev. 2008, 4, 21–26. [Google Scholar] [CrossRef]
- Hu, L.; Liu, Y.; Wang, S. Stem cell-based tooth and periodontal regeneration. Oral Dis. 2018, 24, 696–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, Y.; Kawase-Koga, Y.; Hojo, H.; Yano, F.; Sato, M.; Chung, U.; Ohba, S.; Chikazu, D. Bone regeneration by human dental pulp stem cells using a helioxanthin derivative and cell-sheet technology. Stem Cell Res. Ther. 2018, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathnam, C.; Chueng, S.T.D.; Yang, L.; Lee, K.B. Advanced gene manipulation methods for stem cell theranostics. Theranostics 2017, 7, 2775–2793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, H.; Yoshida, T.; Matsumoto, N.; Aoki, K.; Osada, Y.; Sugimura, T.; Terada, M. Growth regulation of human prostate cancer cells by bone morphogenetic Protein-2. Cancer Res. 1997, 57, 5022–5027. [Google Scholar] [PubMed]
- Pouilit, F.; Blais, A.; Labrie, C. Overexpression of a dominant negative type ii bone morphogenetic protein receptor inhibits the growth of human breast cancer cells. Cancer Res. 2003, 63, 277–281. [Google Scholar]
- Poynton, A.R.; Lane, J.M. Safety profile for the clinical use of bone morphogenetic proteins in the spine. Spine 2002, 27 (Suppl 1), 40–48. [Google Scholar] [CrossRef]
- Lee, K.-B.; Taghavi, C.E.; Murray, S.S.; Song, K.-J.; Keorochana, G.; Wang, J.C. BMP induced inflammation: A comparison of rhBMP-7 and rhBMP-2. J. Orthop. Res. 2012, 30, 1985–1994. [Google Scholar] [CrossRef]
- Moutsatos, L.K.; Turgeman, G.; Zhou, S.; Kurkalli, B.G.; Pelled, G.; Tzur, L.; Kelley, P.; Stumm, N.; Mi, S.; Müller, R.; et al. Exogenously regulated stem cell-mediated gene therapy for bone regeneration. Mol. Ther. 2001, 3, 449–461. [Google Scholar] [CrossRef]
- Gossen, M.; Bujardt, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl. Acad. Sci. USA 1992, 89, 5547–5551. [Google Scholar] [CrossRef] [Green Version]
- Tóth, F.; Gáll, J.M.; Tőzsér, J.; Hegedűs, C. Effect of inducible bone morphogenetic protein 2 expression on the osteogenic differentiation of dental pulp stem cells in vitro. Bone 2020, 132. [Google Scholar] [CrossRef]
- Gafni, Y.; Pelled, G.; Zilberman, Y.; Turgeman, G.; Apparailly, F.; Yotvat, H.; Galun, E.; Gazit, Z.; Jorgensen, C.; Gazit, D. Gene therapy platform for bone regeneration using an exogenously regulated, AAV-2-based gene expression system. Mol. Ther. 2004, 9, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Gersbach, C.A.; Le Doux, J.M.; Guldberg, R.E.; García, A.J. Inducible regulation of Runx2-stimulated osteogenesis. Gene Ther. 2006, 13, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thesleff, I.; Sharpe, P. Signalling networks regulating dental development. Mech. Dev. 1997, 67, 111–123. [Google Scholar] [CrossRef]
- Ye, L.; Bokobza, S.M.; Jiang, W.G. Bone morphogenetic proteins in development and progression of breast cancer and therapeutic potential (review). Int. J. Mol. Med. 2009, 24, 591–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Surendran, K.; Zawaideh, M.A.; Mathew, S.; Hruska, K.A. Bone morphogenetic protein 7: A novel treatment for chronic renal and bone disease. Curr. Opin. Nephrol. Hypertens. 2004, 13, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Niu, L.Y.; An, G.; Guo, Y.S.; Lv, S.C.; Ren, X.P. Expression of recombinant BMP-7 gene increased ossification activity in the rabbit bone mesenchymal stem cells. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3056–3062. [Google Scholar] [PubMed]
- Rabie, A.B.; Wong, R.; Zuo, P.; John Yau, K. BMP7 transfection induces in-vitro osteogenic differentiation of dental pulp mesenchymal stem cells. APOS Trends Orthod. 2013, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Kotliarova, M.S.; Zhuikov, V.A.; Chudinova, Y.V.; Khaidapova, D.D.; Moisenovich, A.M.; Kon’kov, A.S.; Safonova, L.A.; Bobrova, M.M.; Arkhipova, A.Y.; Goncharenko, A.V.; et al. Induction of osteogenic differentiation of osteoblast-like cells MG-63 during cultivation on fibroin microcarriers. Moscow. Univ. Biol. Sci. Bull. 2016, 71, 212–217. [Google Scholar] [CrossRef]
- Zhu, L.; Ma, J.; Mu, R.; Zhu, R.; Chen, F.; Wei, X.; Shi, X.; Zang, S.; Jin, L. Bone morphogenetic protein 7 promotes odontogenic differentiation of dental pulp stem cells in vitro. Life Sci. 2018, 202, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, J.; Shi, B.; Wang, Y.; Chen, X.; Huang, C.; Yang, X.; Xu, D.; Cheng, X.; Chen, X. Combination of scaffold and adenovirus vectors expressing bone morphogenetic protein-7 for alveolar bone regeneration at dental implant defects. Biomaterials 2007, 28, 4635–4642. [Google Scholar] [CrossRef]
- Pecina, M.; Giltaij, L.R.; Vukicevic, S. Orthopaedic applications of osteogenic protein-1 (BMP-7). Int. Orthop. 2001, 25, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, A.P.; Vaccaro, A.R.; Hall, J.A.; Whang, P.G.; Friel, B.C.; McKee, M.D. Clinical applications of BMP-7/OP-1 in fractures, nonunions and spinal fusion. Int. Orthop. 2007, 31, 735–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.-J.; Kim, C.-S.; Han, D.-K.; Cho, I.-H.; Jung, U.-W.; Choi, S.-H.; Kim, C.-K.; Cho, K.-S. The effect of a fibrin-fibronectin/β-tricalcium phosphate/recombinant human bone morphogenetic protein-2 system on bone formation in rat calvarial defects. Biomaterials 2006, 27, 3810–3816. [Google Scholar] [CrossRef]
- Haidar, Z.S.; Hamdy, R.C.; Tabrizian, M. Delivery of recombinant bone morphogenetic proteins for bone regeneration and repair. Part B: Delivery systems for BMPs in orthopaedic and craniofacial tissue engineering. Biotechnol. Lett. 2009, 31, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.J.; Xia, H.; Chen, L.; Ying, Q.S.; Yu, X.; Li, L.H.; Wang, J.H.; Zhang, Y. Efficient delivery of recombinant human bone morphogenetic protein (rhBMP-2) with dextran sulfate-chitosan microspheres. Exp. Ther. Med. 2018, 15, 3265–3272. [Google Scholar] [CrossRef] [Green Version]
- Tóth, F.; Tőzsér, J.; Hegedűs, C. Establishment and examination of an inducible bone morphogenetic protein 7 (BMP-7) expressing dental pulp stem cell line. S Hungarica 2018, 111, 38–43. [Google Scholar]
- Shuen, W.H.; Kan, R.; Yu, Z.; Lung, H.L.; Lung, M.L. Novel lentiviral-inducible transgene expression systems and versatile single-plasmid reporters for in vitro and in vivo cancer biology studies. Cancer Gene Ther. 2015, 22, 207–214. [Google Scholar] [CrossRef]
- Shi, Q.; Huo, N.; Wang, X.; Yang, S.; Wang, J.; Zhang, T. Exosomes from oral tissue stem cells: Biological effects and applications. Cell Biosci. 2020, 10, 108. [Google Scholar] [CrossRef]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H.; et al. BMP signaling in mesenchymal stem cell differentiation and bone formation. J. Biomed Sci. Eng. 2013, 6, 32–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, H.H.; Song, W.-X.; Luo, X.; Manning, D.; Luo, J.; Deng, Z.-L.; Sharff, K.A.; Montag, A.G.; Haydon, R.C.; He, T.-C. Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J. Orthop. Res. 2007, 25, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Horbelt, D.; Denkis, A.; Knaus, P. A portrait of transforming growth factor β superfamily signalling: Background matters. Int. J. Biochem. Cell Biol. 2012, 44, 469–474. [Google Scholar] [CrossRef]
- Zhang, F.; Song, J.; Zhang, H.; Huang, E.; Song, D.; Tollemar, V.; Wang, J.; Wang, J.; Mohammed, M.; Wei, O.; et al. Wnt and BMP signaling crosstalk in regulating dental stem cells: Implications in dental tissue engineering. Genes Dis. 2016, 3, 263–276. [Google Scholar] [CrossRef] [Green Version]
- de Boer, J.; Siddappa, R.; Gaspar, C.; van Apeldoorn, A.; Fodde, R.; van Blitterswijk, C. Wnt signaling inhibits osteogenic differentiation of human mesenchymal stem cells. Bone 2004, 34, 818–826. [Google Scholar] [CrossRef]
- Moioli, E.K.; Hong, L.; Mao, J.J. Inhibition of osteogenic differentiation of human mesenchymal stem cells. Wound Repair Regen. 2007, 15, 413–421. [Google Scholar] [CrossRef]
- Lee, K.-M.; Park, K.H.; Hwang, J.S.; Lee, M.; Yoon, D.S.; Ryu, H.A.; Jung, H.S.; Park, K.W.; Kim, J.; Park, S.W.; et al. Inhibition of STAT5A promotes osteogenesis by DLX5 regulation. Cell Death Dis. 2018, 9, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Miyaki, S.; Ishitobi, H.; Matsuyama, S.; Nakasa, T.; Kamei, N.; Akimoto, T.; Higashi, Y.; Ochi, M. Mesenchymal-stem-cell-derived exosomes accelerate skeletal muscle regeneration. FEBS Lett. 2015, 589, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Phinney, D.G.; Pittenger, M.F. Concise review: Msc-derived exosomes for cell-free therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Giudice, A.; Antonelli, A.; Chiarella, E.; Baudi, F.; Barni, T.; Di Vito, A. The case of medication-related osteonecrosis of the jaw addressed from a pathogenic point of view. Innovative therapeutic strategies: Focus on the most recent discoveries on oral mesenchymal stem cell-derived exosomes. Pharmaceuticals 2020, 13, 423. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Ye, Y.; He, S.; Song, J. SHED-derived conditioned exosomes enhance the osteogenic differentiation of PDLSCs via Wnt and BMP signaling in vitro. Differentiation 2020, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Cardelli, P.; Merciaro, I.; Ettorre, V.; Traini, T.; Bedini, R.; Scionti, D.; Bramanti, A.; Nanci, A.; et al. Three-dimensional printed PLA scaffold and human gingival stem cell-derived extracellular vesicles: A new tool for bone defect repair. Stem Cell Res. Ther. 2018, 9, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Liu, J.; Zhuang, X.; Yu, S.; Zhu, S.; Liu, Y.; Chen, X. Identification and comparison of piRNA expression profiles of exosomes derived from human stem cells from the apical papilla and bone marrow mesenchymal stem cells. Stem Cells Dev. 2020, 29, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Song, Y.; Du, Z.; Yu, F.; Zhang, Y.; Jiang, N.; Ge, X. Exosomes derived from human exfoliated deciduous teeth ameliorate adult bone loss in mice through promoting osteogenesis. J. Mol. Histology 2020, 51, 455–466. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CM | CM+ | OM | OM+ | |
|---|---|---|---|---|
| Day 1 | 0.02 ng | 17.3 ng | 0.09 ng | 18.7 ng |
| Day 2 | 0.23 ng | 34.2 ng | 0.21 ng | 51.7 ng |
| Day 3 | 0.24 ng | 77.3 ng | 0.27 ng | 105.5 ng |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, F.; Tőzsér, J.; Hegedűs, C. Effect of Inducible BMP-7 Expression on the Osteogenic Differentiation of Human Dental Pulp Stem Cells. Int. J. Mol. Sci. 2021, 22, 6182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126182
Tóth F, Tőzsér J, Hegedűs C. Effect of Inducible BMP-7 Expression on the Osteogenic Differentiation of Human Dental Pulp Stem Cells. International Journal of Molecular Sciences. 2021; 22(12):6182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126182
Chicago/Turabian StyleTóth, Ferenc, József Tőzsér, and Csaba Hegedűs. 2021. "Effect of Inducible BMP-7 Expression on the Osteogenic Differentiation of Human Dental Pulp Stem Cells" International Journal of Molecular Sciences 22, no. 12: 6182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126182