
Life and Death of Immature Neurons in the Juvenile and Adult Primate Amygdala


Abstract
:1. Introduction
1.1. A Brief History of the Study of Immature Neurons in the Primate Amygdala
1.2. Regulation of Immature Neurons in the Primate Amygdala
2. Results
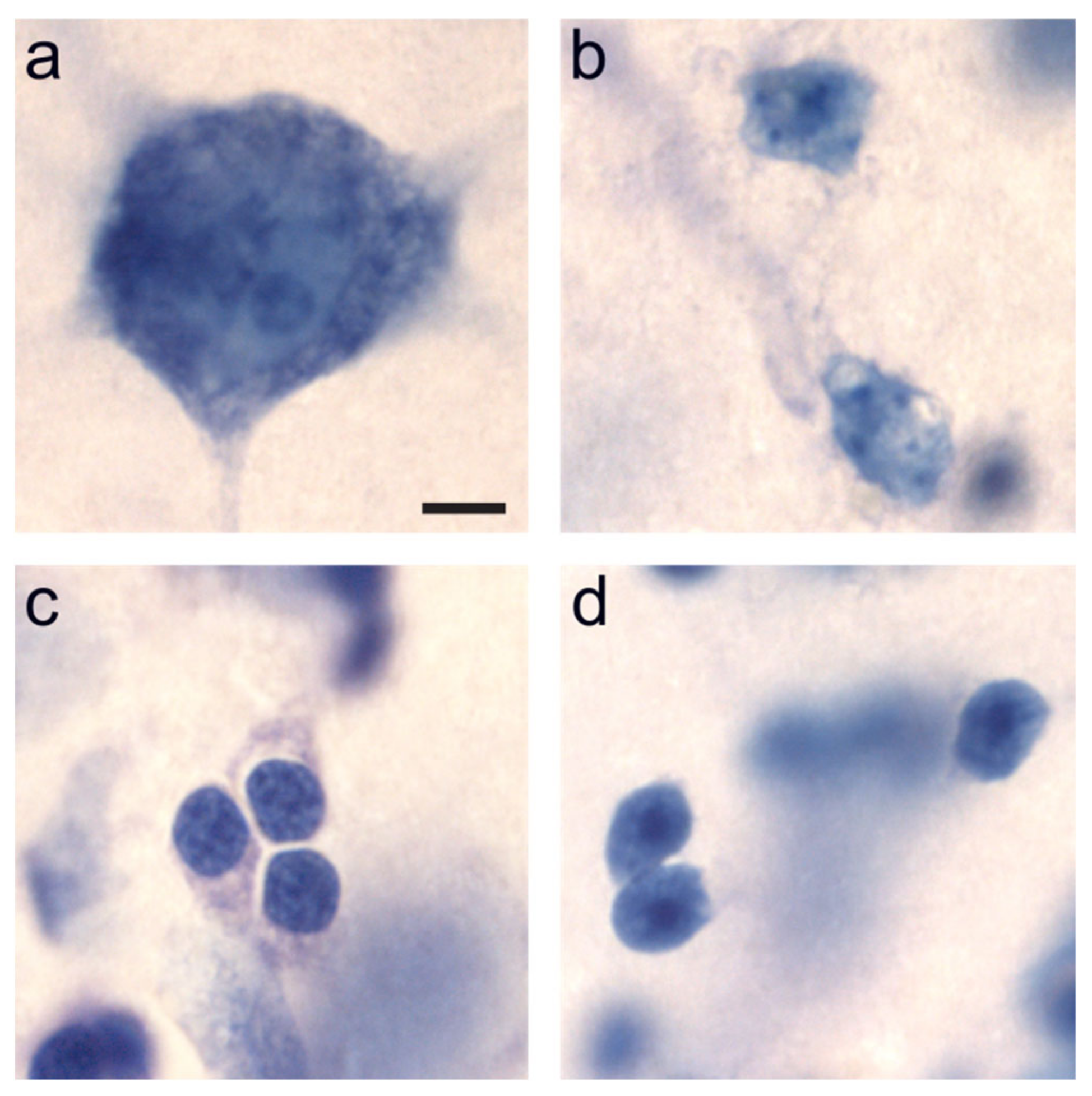
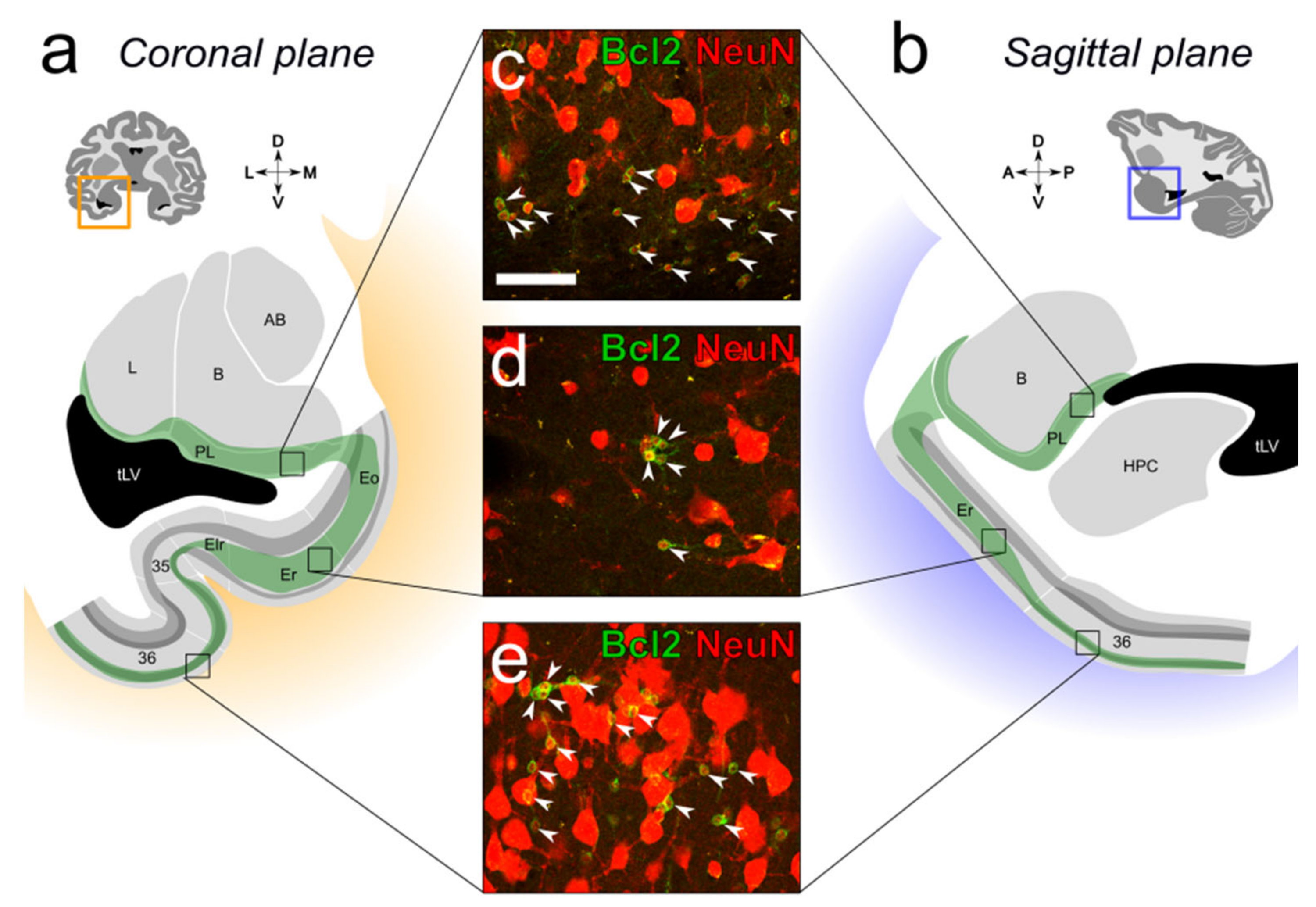
2.1. Distribution of Immature Neurons in the Primate Medial Temporal Lobe
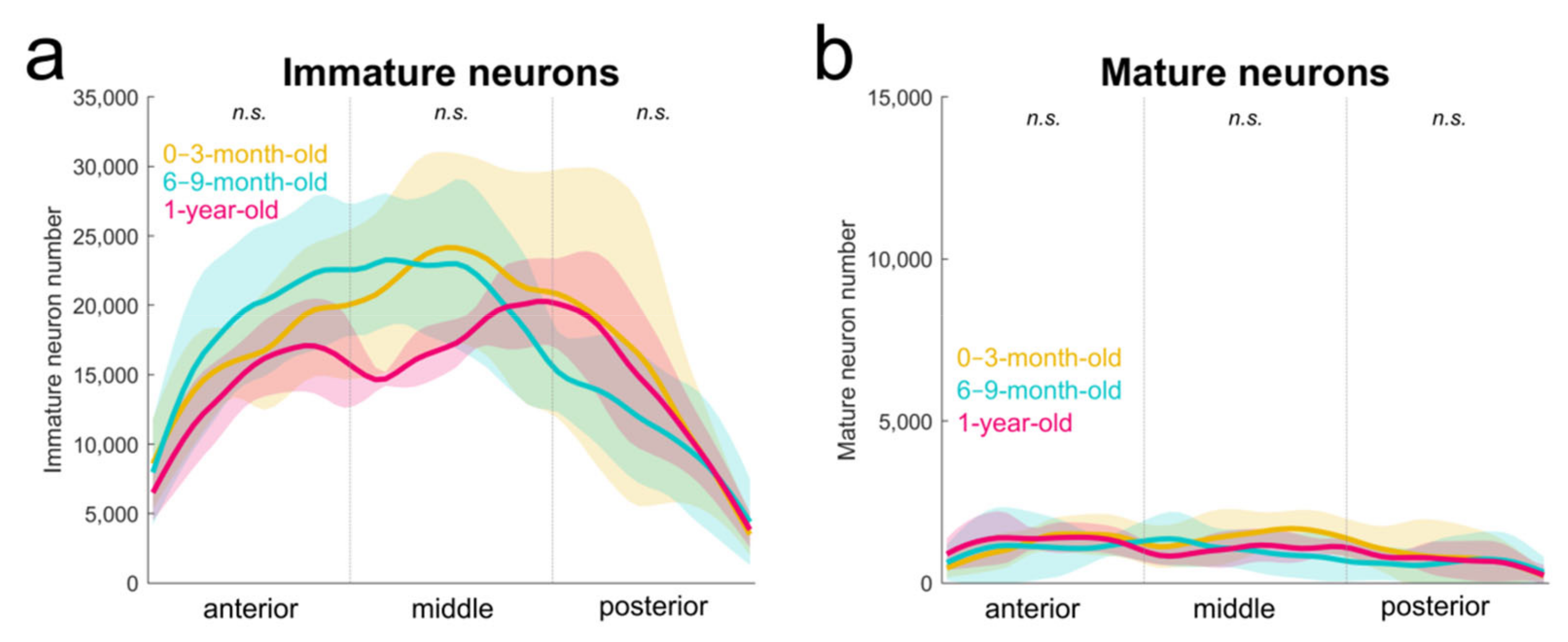
2.2. Immature Neurons from Birth to One Year of Age in Monkeys
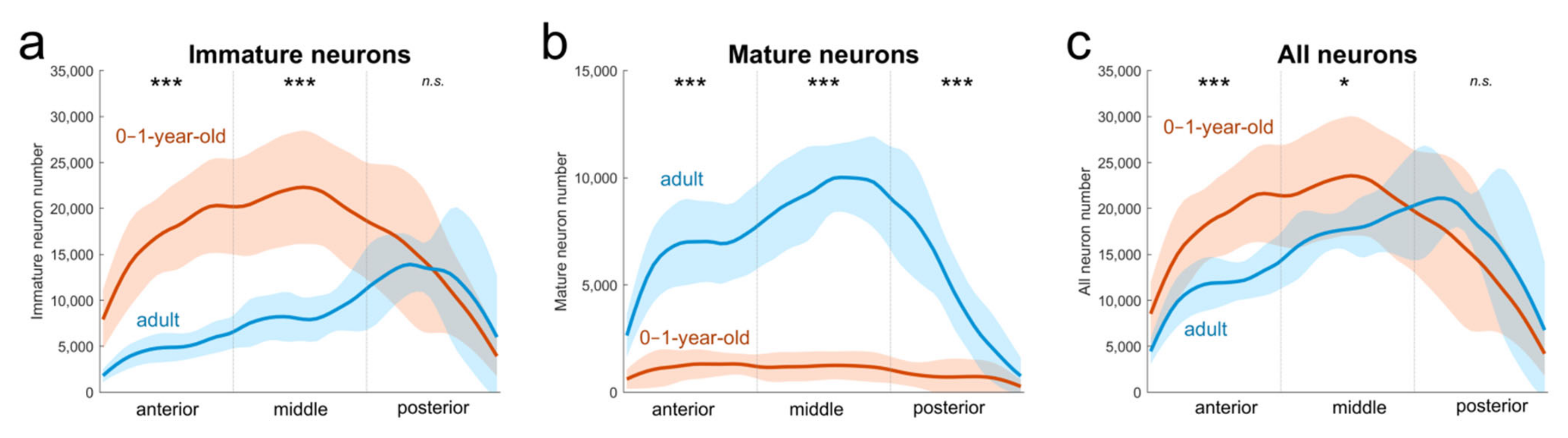
2.3. Immature Neurons from One Year to 5–9 Years of Age in Monkeys
2.4. Neuronal Migration Following Selective Lesion of the Hippocampus
3. Discussion
3.1. Distribution of Immature Neurons in the Primate Medial Temporal Lobe
3.2. Neuronal Migration during Development
3.3. Neuronal Migration Following Selective Lesion of the Hippocampus
3.3.1. Adult Hippocampus Lesion
3.3.2. Neonatal Hippocampus Lesion
3.4. Possible Functional Implications of Increased Neuronal Migration to the Amygdala
3.4.1. Increase in Neuron Density
3.4.2. Excitation-Inhibition Imbalance
3.4.3. Behavioral Alterations in Monkeys and Humans
4. Conclusions
5. Materials and Methods
5.1. Experimental Animals
5.1.1. Unoperated Control Monkeys
5.1.2. Adult Hippocampal-Lesioned Monkeys
5.1.3. Neonatal Hippocampal-Lesioned Monkeys
5.1.4. Experimental Lesion Surgeries
5.2. Histological Procedures
5.2.1. Brain Acquisition
5.2.2. Nissl Staining
5.2.3. Neuronal Phenotype Markers
5.3. Data Acquisition and Statistical Analyses
5.3.1. Stereological Analyses on Nissl-Stained Sections
5.3.2. Neuronal Phenotypes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernier, P.J.; Parent, A. The anti-apoptosis bcl-2 proto-oncogene is preferentially expressed in limbic structures of the primate brain. Neuroscience 1997, 82, 635–640. [Google Scholar] [CrossRef]
- Bernier, P.J.; Parent, A. Bcl-2 Protein as a Marker of Neuronal Immaturity in Postnatal Primate Brain. J. Neurosci. 1998, 18, 2486–2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yachnis, A.T.; Roper, S.N.; Love, A.; Fancey, J.T.; Muir, D. Bcl-2 immunoreactive cells with immature neuronal phenotype exist in the nonepileptic adult human brain. J. Neuropathol. Exp. Neurol. 2000, 59, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, P.J.; Bédard, A.; Vinet, J.; Lévesque, M.; Parent, A. Newly generated neurons in the amygdala and adjoining cortex of adult primates. Proc. Natl. Acad. Sci. USA 2002, 99, 11464–11469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fudge, J.L. Bcl-2 immunoreactive neurons are differentially distributed in subregions of the amygdala and hippocampus of the adult macaque. Neuroscience 2004, 127, 539–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlatt, M.W.; Philippens, I.; Manders, E.; Czéh, B.; Joels, M.; Krugers, H.; Lucassen, P.J. Distinct structural plasticity in the hippocampus and amygdala of the middle-aged common marmoset (Callithrix jacchus). Exp. Neurol. 2011, 230, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-M.; Cai, Y.; Chu, Y.; Chen, E.-Y.; Feng, J.-C.; Luo, X.-G.; Xiong, K.; Struble, R.G.; Clough, R.W.; Patrylo, P.R.; et al. Doublecortin-expressing cells persist in the associate cerebral cortex and amygdala in aged nonhuman primates. Front. Neuroanat. 2009, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decampo, D.M.; Fudge, J.L. Where and what is the paralaminar nucleus? A review on a unique and frequently overlooked area of the primate amygdala. Neurosci. Biobehav. Rev. 2012, 36, 520–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chareyron, L.J.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Postnatal development of the amygdala: A stereological study in macaque monkeys. J. Comp. Neurol. 2012, 520, 1965–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chareyron, L.J.; Lavenex, P.B.; Lavenex, P. Postnatal development of the amygdala: A stereological study in rats. J. Comp. Neurol. 2012, 520, 3745–3763. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, L.; Palazzo, O.; La Rosa, C.; Piumatti, M. Do large brains of long-living mammals prefer non-newly generated, immature neurons? Neural Regen. Res. 2018, 13, 633–634. [Google Scholar] [CrossRef]
- Jhaveri, D.J.; Tedoldi, A.; Hunt, S.; Sullivan, R.; Watts, N.R.; Power, J.M.; Bartlett, P.F.; Sah, P. Evidence for newly generated interneurons in the basolateral amygdala of adult mice. Mol. Psychiatry 2017, 23, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Chareyron, L.J.; Amaral, D.G.; Lavenex, P. Selective lesion of the hippocampus increases the differentiation of immature neurons in the monkey amygdala. Proc. Natl. Acad. Sci. USA 2016, 113, 14420–14425. [Google Scholar] [CrossRef] [Green Version]
- Chareyron, L.J.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Stereological analysis of the rat and monkey amygdala. J. Comp. Neurol. 2011, 519, 3218–3239. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, M.; Drevets, W.C.; Price, J.L. Glial reduction in amygdala in major depressive disorder is due to oligodendrocytes. Biol. Psychiatry 2004, 55, 563–569. [Google Scholar] [CrossRef]
- García-Cabezas, M.Á.; John, Y.J.; Barbas, H.; Zikopoulos, B. Distinction of Neurons, Glia and Endothelial Cells in the Cerebral Cortex: An Algorithm Based on Cytological Features. Front. Neuroanat. 2016, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Grady, M.S.; Charleston, J.S.; Maris, D.; Witgen, B.M.; Lifshitz, J. Neuronal and Glial Cell Number in the Hippocampus after Experimental Traumatic Brain Injury: Analysis by Stereological Estimation. J. Neurotrauma 2003, 20, 929–941. [Google Scholar] [CrossRef]
- Fitting, S.; Booze, R.; Hasselrot, U.; Mactutus, C.F. Differential long-term neurotoxicity of HIV-1 proteins in the rat hippocampal formation: A design-based stereological study. Hippocampus 2008, 18, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.F.; Paredes, M.F.; Velmeshev, D.; Herranz-Pérez, V.; Sandoval, K.; Mayer, S.; Chang, E.F.; Insausti, R.; Kriegstein, A.R.; Rubenstein, J.L.; et al. Immature excitatory neurons develop during adolescence in the human amygdala. Nat. Commun. 2019, 10, 2748. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Boekhoorn, K.; Joels, M.; Lucassen, P.J. Increased proliferation reflects glial and vascular-associated changes, but not neurogenesis in the presenile Alzheimer hippocampus. Neurobiol. Dis. 2006, 24, 1–14. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Toni, N.; Kempermann, G.; Frisen, J.; Gage, F.H.; Swaab, D.F. Limits to human neurogenesis—really? Mol. Psychiatry 2020, 25, 2207–2209. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Fitzsimons, C.P.; Salta, E.; Maletic-Savatic, M. Adult neurogenesis, human after all (again): Classic, optimized, and future approaches. Behav. Brain Res. 2020, 381, 112458. [Google Scholar] [CrossRef]
- Fudge, J.; Decampo, D.; Becoats, K. Revisiting the hippocampal–amygdala pathway in primates: Association with immature-appearing neurons. Neuroscience 2012, 212, 104–119. [Google Scholar] [CrossRef] [Green Version]
- Avino, T.A.; Barger, N.; Vargas, M.V.; Carlson, E.L.; Amaral, D.G.; Bauman, M.D.; Schumann, C.M. Neuron numbers increase in the human amygdala from birth to adulthood, but not in autism. Proc. Natl. Acad. Sci. USA 2018, 115, 3710–3715. [Google Scholar] [CrossRef] [Green Version]
- Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Hippocampal Lesion Prevents Spatial Relational Learning in Adult Macaque Monkeys. J. Neurosci. 2006, 26, 4546–4558. [Google Scholar] [CrossRef]
- Bauman, M.D.; Lavenex, P.; Mason, W.A.; Capitanio, J.P.; Amaral, D.G. The Development of Mother-Infant Interactions after Neonatal Amygdala Lesions in Rhesus Monkeys. J. Neurosci. 2004, 24, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Chu-Wang, I.-W.; Oppenheim, R.W. Cell death of motoneurons in the chick embryo spinal cord. I. A light and electron microscopic study of naturally occurring and induced cell loss during development. J. Comp. Neurol. 1978, 177, 33–57. [Google Scholar] [CrossRef]
- Cowan, W.M.; Fawcett, J.W.; O’Leary, D.D.; Stanfield, B.B. Regressive events in neurogenesis. Science 1984, 225, 1258–1265. [Google Scholar] [CrossRef]
- Lossi, L.; Merighi, A. In vivo cellular and molecular mechanisms of neuronal apoptosis in the mammalian CNS. Prog. Neurobiol. 2003, 69, 287–312. [Google Scholar] [CrossRef]
- Dekkers, M.P.; Nikoletopoulou, V.; Barde, Y.-A. Death of developing neurons: New insights and implications for connectivity. J. Cell Biol. 2013, 203, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, R.W. Cell Death During Development of the Nervous System. Annu. Rev. Neurosci. 1991, 14, 453–501. [Google Scholar] [CrossRef] [PubMed]
- Petreanu, L.; Alvarez-Buylla, A. Maturation and Death of Adult-Born Olfactory Bulb Granule Neurons: Role of Olfaction. J. Neurosci. 2002, 22, 6106–6113. [Google Scholar] [CrossRef] [PubMed]
- Kirn, J.R.; Fishman, Y.; Sasportas, K.; Alvarez-Buylla, A.; Nottebohm, F. Fate of New Neurons in Adult Canary High Vocal Center during the First 30 Days after Their Formation. J. Comp. Neurol. 1999, 411, 487–494. [Google Scholar] [CrossRef]
- Dayer, A.G.; Ford, A.A.; Cleaver, K.M.; Yassaee, M.; Cameron, H.A. Short-term and long-term survival of new neurons in the rat dentate gyrus. J. Comp. Neurol. 2003, 460, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G.; Gast, D.; Kronenberg, G.; Yamaguchi, M.; Gage, F.H. Early determination and long-term persistence of adult-generated new neurons in the hippocampus of mice. Development 2003, 130, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Yuri, K.; Ozawa, H.; Lu, H.; Kawata, M. TheIn VivoTime Course for Elimination of Adrenalectomy-Induced Apoptotic Profiles from the Granule Cell Layer of the Rat Hippocampus. J. Neurosci. 1997, 17, 3981–3989. [Google Scholar] [CrossRef] [Green Version]
- Jabès, A.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Quantitative analysis of postnatal neurogenesis and neuron number in the macaque monkey dentate gyrus. Eur. J. Neurosci. 2010, 31, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Van der Beek, E.M.; Wiegant, V.M.; Schouten, W.G.; van Eerdenburg, F.J.C.M.; Loijens, L.W.; van der Plas, C.; Benning, M.A.; de Vries, H.; de Kloet, E.R.; Lucassen, P.J. Neuronal number, volume, and apoptosis of the left dentate gyrus of chronically stressed pigs correlate negatively with basal saliva cortisol levels. Hippocampus 2004, 14, 688–700. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Müller, M.B.; Holsboer, F.; Bauer, J.; Holtrop, A.; Wouda, J.; Hoogendijk, W.J.; De Kloet, E.R.; Swaab, D.F. Hippocampal Apoptosis in Major Depression Is a Minor Event and Absent from Subareas at Risk for Glucocorticoid Overexposure. Am. J. Pathol. 2001, 158, 453–468. [Google Scholar] [CrossRef] [Green Version]
- Lucassen, P.J.; Fuchs, E.; Czeh, B. Antidepressant treatment with tianeptine reduces apoptosis in the hippocampal dentate gyrus and temporal cortex. Biol. Psychiatry 2004, 55, 789–796. [Google Scholar] [CrossRef]
- Bonfanti, L. PSA-NCAM in mammalian structural plasticity and neurogenesis. Prog. Neurobiol. 2006, 80, 129–164. [Google Scholar] [CrossRef]
- Chang, E.H.; Adorjan, I.; Mundim, M.V.; Sun, B.; Dizon, M.L.V.; Szele, F.G. Traumatic Brain Injury Activation of the Adult Subventricular Zone Neurogenic Niche. Front. Neurosci. 2016, 10, 332. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.; Minami, M.; Lan, J.Q.; Mao, X.O.; Batteur, S.; Simon, R.P.; Greenberg, D.A. Neurogenesis in dentate subgranular zone and rostral subventricular zone after focal cerebral ischemia in the rat. Proc. Natl. Acad. Sci. USA 2001, 98, 4710–4715. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Isobe, Y.; Aihara, N.; Furuyama, F.; Misumi, S.; Kim, T.-S.; Nishino, H.; Hida, H. Increase in neurogenesis and neuroblast migration after a small intracerebral hemorrhage in rats. Neurosci. Lett. 2007, 425, 114–119. [Google Scholar] [CrossRef]
- Tonchev, A.B.; Yamashima, T.; Zhao, L.; Okano, H.J.; Okano, H. Proliferation of neural and neuronal progenitors after global brain ischemia in young adult macaque monkeys. Mol. Cell. Neurosci. 2003, 23, 292–301. [Google Scholar] [CrossRef]
- Tonchev, A.B.; Yamashima, T.; Sawamoto, K.; Okano, H. Enhanced proliferation of progenitor cells in the subventricular zone and limited neuronal production in the striatum and neocortex of adult macaque monkeys after global cerebral ischemia. J. Neurosci. Res. 2005, 81, 776–788. [Google Scholar] [CrossRef]
- Koketsu, D.; Furuichi, Y.; Maeda, M.; Matsuoka, N.; Miyamoto, Y.; Hisatsune, T. Increased number of new neurons in the olfactory bulb and hippocampus of adult non-human primates after focal ischemia. Exp. Neurol. 2006, 199, 92–102. [Google Scholar] [CrossRef]
- Zheng, W.; Zhuge, Q.; Zhong, M.; Chen, G.; Shao, B.; Wang, H.; Mao, X.; Xie, L.; Jin, K. Neurogenesis in Adult Human Brain after Traumatic Brain Injury. J. Neurotrauma 2013, 30, 1872–1880. [Google Scholar] [CrossRef] [Green Version]
- Macas, J.; Nern, C.; Plate, K.H.; Momma, S. Increased Generation of Neuronal Progenitors after Ischemic Injury in the Aged Adult Human Forebrain. J. Neurosci. 2006, 26, 13114–13119. [Google Scholar] [CrossRef] [Green Version]
- Marti-Fabregas, J.; Romaguera-Ros, M.; Gomez-Pinedo, U.; Martinez-Ramirez, S.; Jimenez-Xarrie, E.; Marin, R.; Marti-Vilalta, J.-L.; García-Verdugo, J.M. Proliferation in the human ipsilateral subventricular zone after ischemic stroke. Neurology 2010, 74, 357–365. [Google Scholar] [CrossRef]
- Jin, K.; Sun, Y.; Xie, L.; Peel, A.; Mao, X.O.; Batteur, S.; Greenberg, D.A. Directed migration of neuronal precursors into the ischemic cerebral cortex and striatum. Mol. Cell. Neurosci. 2003, 24, 171–189. [Google Scholar] [CrossRef]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Parent, J.M.; Vexler, Z.S.; Gong, C.; Ma, N.D.; Ferriero, D.M. Rat forebrain neurogenesis and striatal neuron replacement after focal stroke. Ann. Neurol. 2002, 52, 802–813. [Google Scholar] [CrossRef]
- Wang, C.; Liu, F.; Liu, Y.-Y.; Zhao, C.-H.; You, Y.; Wang, L.; Zhang, J.; Wei, B.; Ma, T.; Zhang, Q.; et al. Identification and characterization of neuroblasts in the subventricular zone and rostral migratory stream of the adult human brain. Cell Res. 2011, 21, 1534–1550. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, S.; Takagi, Y.; Harada, J.; Teramoto, T.; Thomas, S.S.; Waeber, C.; Bakowska, J.C.; Breakefield, X.O.; Moskowitz, M.A. FGF-2 regulation of neurogenesis in adult hippocampus after brain injury. Proc. Natl. Acad. Sci. USA 2001, 98, 5874–5879. [Google Scholar] [CrossRef] [Green Version]
- Dempsey, R.J.; Sailor, K.A.; Bowen, K.K.; Türeyen, K.; Vemuganti, R. Stroke-induced progenitor cell proliferation in adult spontaneously hypertensive rat brain: Effect of exogenous IGF-1 and GDNF. J. Neurochem. 2003, 87, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Sanai, N.; Nguyen, T.; Ihrie, R.A.; Mirzadeh, Z.; Tsai, H.-H.; Wong, M.; Gupta, N.; Berger, M.S.; Huang, E.; García-Verdugo, J.M.; et al. Corridors of migrating neurons in the human brain and their decline during infancy. Nature 2011, 478, 382–386. [Google Scholar] [CrossRef]
- La Rosa, C.; Ghibaudi, M.; Bonfanti, L. Newly Generated and Non-Newly Generated “Immature” Neurons in the Mammalian Brain: A Possible Reservoir of Young Cells to Prevent Brain Aging and Disease? J. Clin. Med. 2019, 8, 685. [Google Scholar] [CrossRef] [Green Version]
- Selemon, L.D.; Goldman-Rakic, P.S. The reduced neuropil hypothesis: A circuit based model of schizophrenia. Biol. Psychiatry 1999, 45, 17–25. [Google Scholar] [CrossRef]
- Bakhshi, K.; Chance, S. The neuropathology of schizophrenia: A selective review of past studies and emerging themes in brain structure and cytoarchitecture. Neuroscience 2015, 303, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Sohal, V.S.; Rubenstein, J.L.R. Excitation-inhibition balance as a framework for investigating mechanisms in neuropsychiatric disorders. Mol. Psychiatry 2019, 24, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Ikrar, T.; Guo, N.; He, K.; Besnard, A.; Levinson, S.; Hill, A.; Lee, H.-K.; Hen, R.; Xu, X.; Sahay, A. Adult neurogenesis modifies excitability of the dentate gyrus. Front. Neural Circuits 2013, 7, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauman, M.D.; Lavenex, P.; Mason, W.A.; Capitanio, J.P.; Amaral, D.G. The Development of Social Behavior Following Neonatal Amygdala Lesions in Rhesus Monkeys. J. Cogn. Neurosci. 2004, 16, 1388–1411. [Google Scholar] [CrossRef]
- Bauman, M.D.; Toscano, J.E.; Mason, W.A.; Lavenex, P.; Amaral, D.G. The expression of social dominance following neonatal lesions of the amygdala or hippocampus in rhesus monkeys (Macaca mulatta). Behav. Neurosci. 2006, 120, 749–760. [Google Scholar] [CrossRef]
- Bauman, M.D.; Toscano, J.E.; Babineau, B.A.; Mason, W.A.; Amaral, D.G. Emergence of stereotypies in juvenile monkeys (Macaca mulatta) with neonatal amygdala or hippocampus lesions. Behav. Neurosci. 2008, 122, 1005–1015. [Google Scholar] [CrossRef] [Green Version]
- Bliss-Moreau, E.; Toscano, J.E.; Bauman, M.D.; Mason, W.A.; Amaral, D.G. Neonatal amygdala or hippocampus lesions influence responsiveness to objects. Dev. Psychobiol. 2010, 52, 487–503. [Google Scholar] [CrossRef] [Green Version]
- Bliss-Moreau, E.; Moadab, G.; Bauman, M.D.; Amaral, D.G. The Impact of Early Amygdala Damage on Juvenile Rhesus Macaque Social Behavior. J. Cogn. Neurosci. 2013, 25, 2124–2140. [Google Scholar] [CrossRef] [Green Version]
- Bliss-Moreau, E.; Toscano, J.E.; Bauman, M.; Mason, W.A.; Amaral, D.G. Neonatal amygdala lesions alter responsiveness to objects in juvenile macaques. Neuroscience 2011, 178, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Bliss-Moreau, E.; Bauman, M.D.; Amaral, D.G. Neonatal amygdala lesions result in globally blunted affect in adult rhesus macaques. Behav. Neurosci. 2011, 125, 848–858. [Google Scholar] [CrossRef]
- Moadab, G.; Bliss-Moreau, E.; Amaral, D.G. Adult social behavior with familiar partners following neonatal amygdala or hippocampus damage. Behav. Neurosci. 2015, 129, 339–350. [Google Scholar] [CrossRef]
- Moadab, G.; Bliss-Moreau, E.; Bauman, M.D.; Amaral, D.G. Early amygdala or hippocampus damage influences adolescent female social behavior during group formation. Behav. Neurosci. 2017, 131, 68–82. [Google Scholar] [CrossRef]
- Bliss-Moreau, E.; Moadab, G.; Santistevan, A.; Amaral, D.G. The effects of neonatal amygdala or hippocampus lesions on adult social behavior. Behav. Brain Res. 2017, 322, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Chareyron, L.J.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Functional organization of the medial temporal lobe memory system following neonatal hippocampal lesion in rhesus monkeys. Brain Struct. Funct. 2017, 222, 3899–3914. [Google Scholar] [CrossRef]
- Lavenex, P.; Lavenex, P.B.; Amaral, D.G. Spatial relational learning persists following neonatal hippocampal lesions in macaque monkeys. Nat. Neurosci. 2006, 10, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Maguire, E.A.; Vargha-Khadem, F.; Mishkin, M. The effects of bilateral hippocampal damage on fMRI regional activations and interactions during memory retrieval. Brain 2001, 124, 1156–1170. [Google Scholar] [CrossRef]
- Amaral, D.G.; Bauman, M.D.; Capitanio, J.P.; Lavenex, P.; Mason, W.A.; Mauldin-Jourdain, M.L.; Mendoza, S.P. The amygdala: Is it an essential component of the neural network for social cognition? Neuropsychologia 2003, 41, 517–522. [Google Scholar] [CrossRef]
- Adolphs, R. What does the amygdala contribute to social cognition? Ann. N. Y. Acad. Sci. 2010, 1191, 42–61. [Google Scholar] [CrossRef] [Green Version]
- Järvinen-Pasley, A.; Adolphs, R.; Yam, A.; Hill, K.J.; Grichanik, M.; Reilly, J.; Mills, D.; Reiss, A.L.; Korenberg, J.R.; Bellugi, U. Affiliative behavior in Williams syndrome: Social perception and real-life social behavior. Neuropsychologia 2010, 48, 2110–2119. [Google Scholar] [CrossRef] [Green Version]
- Lew, C.H.; Groeniger, K.M.; Bellugi, U.; Stefanacci, L.; Schumann, C.M.; Semendeferi, K. A postmortem stereological study of the amygdala in Williams syndrome. Brain Struct. Funct. 2017, 223, 1897–1907. [Google Scholar] [CrossRef]
- Bostelmann, M.; Ruggeri, P.; Circelli, A.R.; Costanzo, F.; Menghini, D.; Vicari, S.; Lavenex, P.; Lavenex, P.B. Path Integration and Cognitive Mapping Capacities in Down and Williams Syndromes. Front. Psychol. 2020, 11, 571394. [Google Scholar] [CrossRef]
- Bostelmann, M.; Fragnière, E.; Costanzo, F.; Di Vara, S.; Menghini, D.; Vicari, S.; Lavenex, P.; Lavenex, P.B. Dissociation of spatial memory systems in Williams syndrome. Hippocampus 2017, 27, 1192–1203. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Mervis, C.B.; Sarpal, D.; Koch, P.; Steele, S.; Kohn, P.; Marenco, S.; Morris, C.A.; Das, S.; Kippenhan, S.; et al. Functional, structural, and metabolic abnormalities of the hippocampal formation in Williams syndrome. J. Clin. Investig. 2005, 115, 1888–1895. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Lindenberg, A.; Mervis, C.B.; Berman, K.F. Neural mechanisms in Williams syndrome: A unique window to genetic influences on cognition and behaviour. Nat. Rev. Neurosci. 2006, 7, 380–393. [Google Scholar] [CrossRef]
- Jabès, A.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Postnatal development of the hippocampal formation: A stereological study in macaque monkeys. J. Comp. Neurol. 2011, 519, 1051–1070. [Google Scholar] [CrossRef] [Green Version]
- Piguet, O.; Chareyron, L.J.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Stereological analysis of the rhesus monkey entorhinal cortex. J. Comp. Neurol. 2018, 526, 2115–2132. [Google Scholar] [CrossRef] [Green Version]
- Piguet, O.; Chareyron, L.J.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Postnatal development of the entorhinal cortex: A stereological study in macaque monkeys. J. Comp. Neurol. 2020, 528, 2308–2332. [Google Scholar] [CrossRef] [PubMed]
- Lavenex, P.; Lavenex, P.B.; Bennett, J.L.; Amaral, D.G. Postmortem changes in the neuroanatomical characteristics of the primate brain: Hippocampal formation. J. Comp. Neurol. 2009, 512, 27–51. [Google Scholar] [CrossRef] [Green Version]
- Mullen, R.; Buck, C.; Smith, A. NeuN, a neuronal specific nuclear protein in vertebrates. Development 1992, 116, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.G.; Jensen, E.B. The efficiency of systematic sampling in stereology and its prediction*. J. Microsc. 1987, 147, 229–263. [Google Scholar] [CrossRef] [PubMed]
- West, M.J.; Gundersen, H.J.G. Unbiased stereological estimation of the number of neurons in the human hippocampus. J. Comp. Neurol. 1990, 296, 1–22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immature neurons | ||||
| 0–3-month | 6–9-month | 1-year | Adult | |
| Anterior | 320,116 ± 52,162 | 365,778 ± 72,720 | 278,854 ± 34,880 | 92,609 ± 22,334 |
| Middle | 447,066 ± 117,968 | 429,655 ± 84,484 | 349,521 ± 21,913 | 168,363 ± 41,566 |
| Posterior | 275,609 ± 139,239 | 217,477 ± 68,508 | 264,992 ± 67,537 | 237,238 ± 69,921 |
| TOTAL | 1,042,791 ± 250,175 | 1,012,910 ± 193,789 | 893,367 ± 71,509 | 498,210 ± 82,884 |
| Mature neurons | ||||
| 0–3-month | 6–9-month | 1-year | Adult | |
| Anterior | 23,708 ± 5,900 | 21,473 ± 14,113 | 25,880 ± 4,387 | 127,076 ± 26,604 |
| Middle | 28,775 ± 9,750 | 20,744 ± 13,227 | 20,660 ± 6,018 | 186,252 ± 30,750 |
| Posterior | 15,990 ± 13,555 | 12,178 ± 12,696 | 14,100 ± 6,516 | 93,796 ± 21,515 |
| TOTAL | 68,473 ± 14,508 | 54,395 ± 31,898 | 60,640 ± 5,811 | 407,123 ± 37,270 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chareyron, L.J.; Banta Lavenex, P.; Amaral, D.G.; Lavenex, P. Life and Death of Immature Neurons in the Juvenile and Adult Primate Amygdala. Int. J. Mol. Sci. 2021, 22, 6691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136691
Chareyron LJ, Banta Lavenex P, Amaral DG, Lavenex P. Life and Death of Immature Neurons in the Juvenile and Adult Primate Amygdala. International Journal of Molecular Sciences. 2021; 22(13):6691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136691
Chicago/Turabian StyleChareyron, Loïc J., Pamela Banta Lavenex, David G. Amaral, and Pierre Lavenex. 2021. "Life and Death of Immature Neurons in the Juvenile and Adult Primate Amygdala" International Journal of Molecular Sciences 22, no. 13: 6691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136691