
Bioactive Lipid O-cyclic phytosphingosine-1-phosphate Promotes Differentiation of Human Embryonic Stem Cells into Cardiomyocytes via ALK3/BMPR Signaling

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Embryonic Stem Cell (hESC) Maintenance
2.2. Cardiomyocyte Differentiation
2.3. Immunofluorescence Staining
2.4. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.5. Immunoblotting
2.6. Characterizing Spontaneous Contractions of Beating Cardiomyocytes
2.7. Statistical Analysis
3. Results
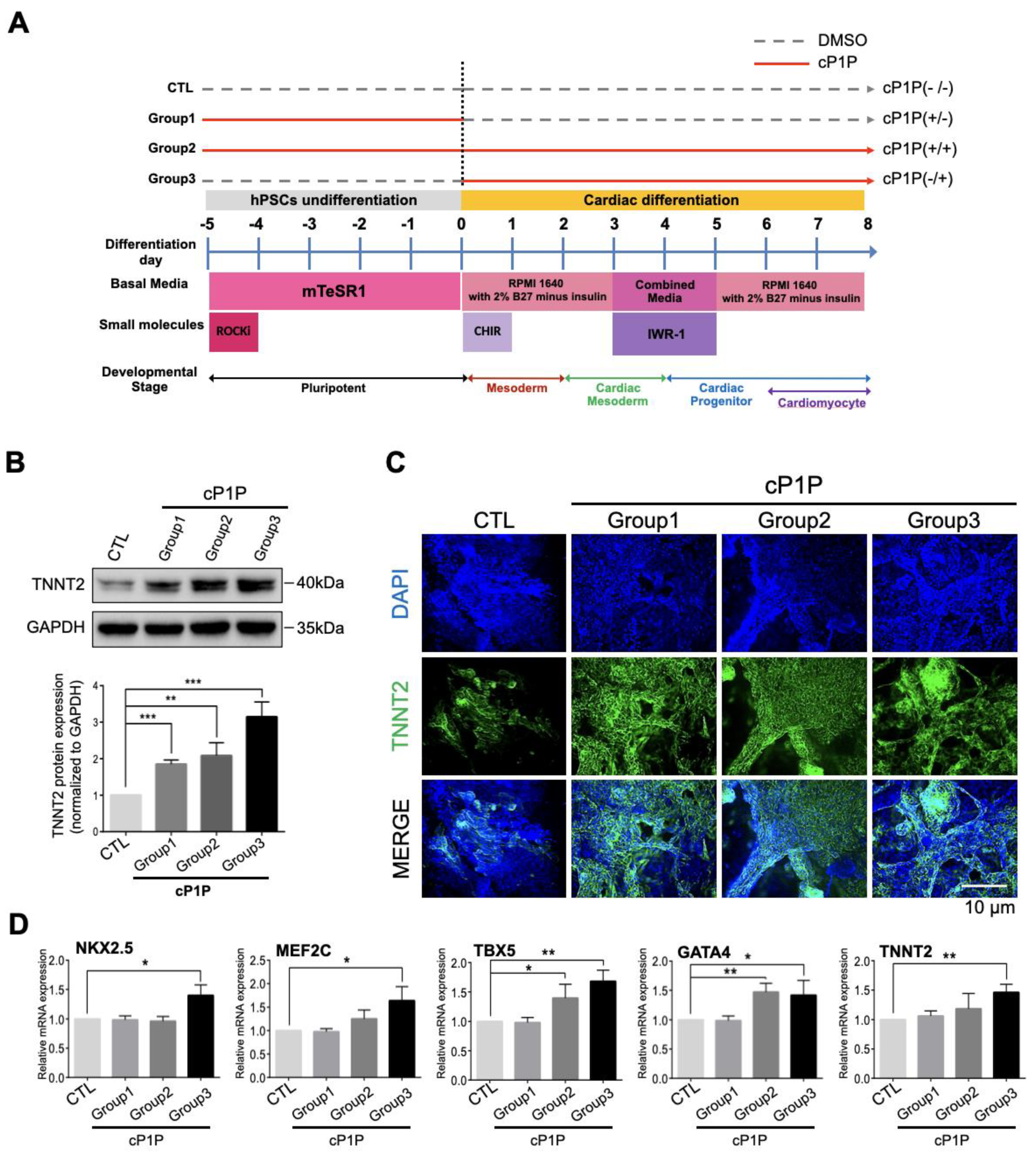
3.1. cP1P Enhances Cardiac Differentiation in Human Embryonic Stem Cells
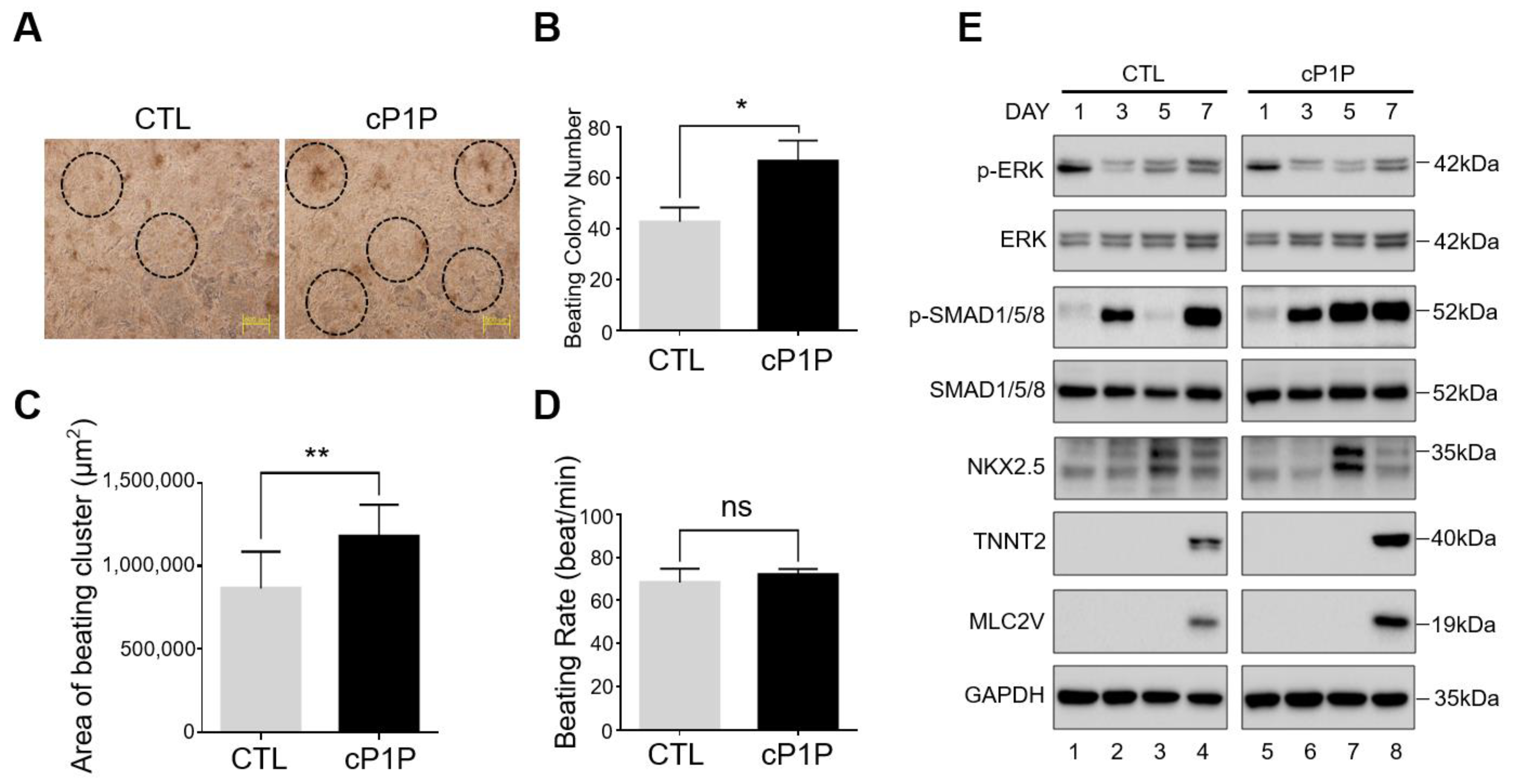
3.2. Bioactive Lipid cP1P Augments Beating Colony Number and Contracting Area in hESC-Derived Cardiomyocytes
3.3. Bioactive Lipid cP1P Activates SMAD Signaling during Cardiomyocyte Differentiation
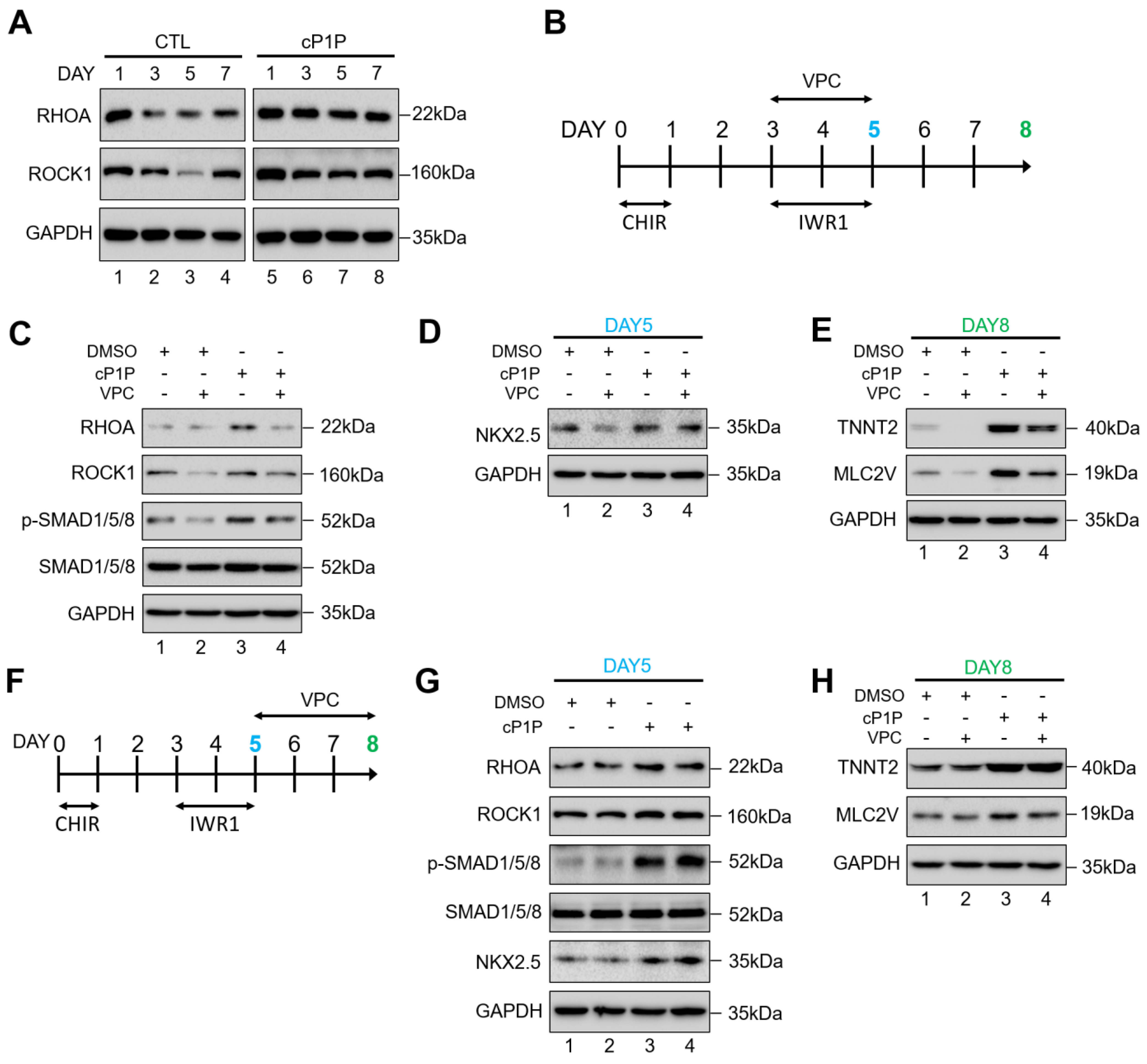
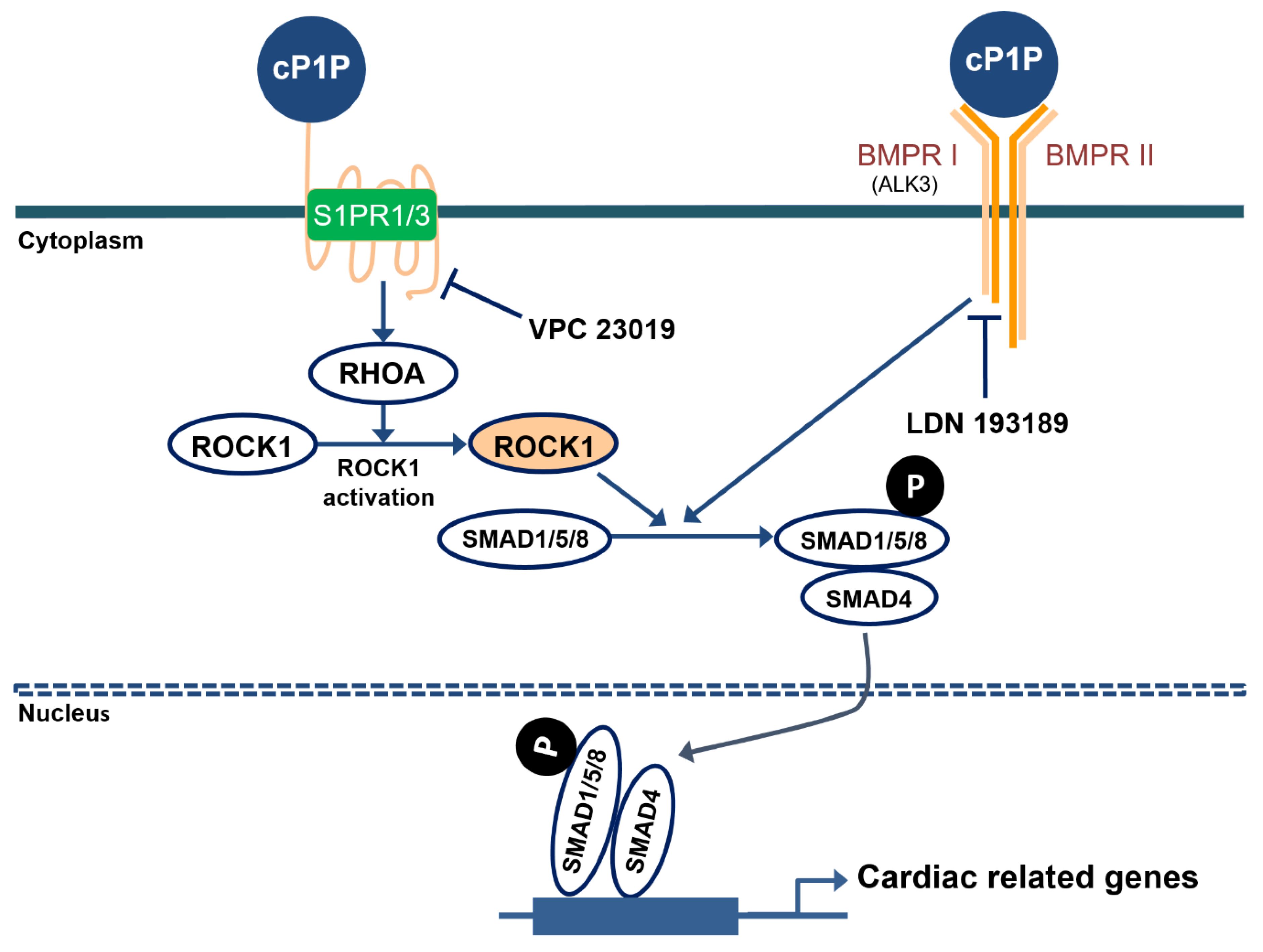
3.4. cP1P Promotes Cardiac Differentiation in hESCs by Partially Regulating S1PR-Mediated SMAD1/5/8 Signaling
3.5. cP1P Augments Cardiac Differentiation in hESCs Dependent on BMP Receptor-Mediated SMAD1/5/8 Signaling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xin, M.; Olson, E.N.; Bassel-Duby, R. Mending broken hearts: Cardiac development as a basis for adult heart regeneration and repair. Nat. Rev. Mol. Cell Biol. 2013, 14, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, J.T.; Moretti, A.; Laugwitz, K.L. Multipotent progenitor cells in regenerative cardiovascular medicine. Pediatr. Cardiol. 2009, 30, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Nsair, A.; MacLellan, W.R. Induced pluripotent stem cells for regenerative cardiovascular therapies and biomedical discovery. Adv. Drug Deliv. Rev. 2011, 63, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, J.; Khalil, M.; Shishechian, N.; Hannes, T.; Pfannkuche, K.; Liang, H.; Fatima, A.; Haustein, M.; Suhr, F.; Bloch, W.; et al. Comparison of contractile behavior of native murine ventricular tissue and cardiomyocytes derived from embryonic or induced pluripotent stem cells. FASEB J. 2010, 24, 2739–2751. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenkin, A.; Liang, H.; Xu, G.; Pfannkuche, K.; Eichhorn, H.; Fatima, A.; Luo, H.; Saric, T.; Wernig, M.; Jaenisch, R.; et al. Functional characterization of cardiomyocytes derived from murine induced pluripotent stem cells in vitro. FASEB J. 2009, 23, 4168–4180. [Google Scholar] [CrossRef]
- Ueno, S.; Weidinger, G.; Osugi, T.; Kohn, A.D.; Golob, J.L.; Pabon, L.; Reinecke, H.; Moon, R.T.; Murry, C.E. Biphasic role for Wnt/β-catenin signaling in cardiac specification in zebrafish and embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 9685–9690. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Gadue, P.; Huber, T.L.; Paddison, P.J.; Keller, G.M. Wnt and TGF-β signaling are required for the induction of an in vitro model of primitive streak formation using embryonic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 16806–16811. [Google Scholar] [CrossRef] [Green Version]
- Klaus, A.; Saga, Y.; Taketo, M.M.; Tzahor, E.; Birchmeier, W. Distinct roles of Wnt/β-catenin and Bmp signaling during early cardiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 18531–18536. [Google Scholar] [CrossRef] [Green Version]
- Schultheiss, T.M.; Burch, J.; Lassar, A.B. A role for bone morphogenetic proteins in the induction of cardiac myogenesis. Genes Dev. 1997, 11, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Avery, K.; Avery, S.; Shepherd, J.; Heath, P.R.; Moore, H. Sphingosine-1-Phosphate Mediates Transcriptional Regulation of Key Targets Associated with Survival, Proliferation, and Pluripotency in Human Embryonic Stem Cells. Stem Cells Dev. 2008, 17, 1195–1206. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Izpisúa Belmonte, J.C. Albumin-Associated Lipids Regulate Human Embryonic Stem Cell Self-Renewal. PLoS ONE 2008, 3, e1384. [Google Scholar] [CrossRef] [Green Version]
- Pébay, A.; Bonder, C.S.; Pitson, S.M. Stem cell regulation by lysophospholipids. Prostaglandins Other Lipid Mediat. 2007, 84, 83–97. [Google Scholar] [CrossRef]
- Kleger, A.; Liebau, S.; Lin, Q.; von Wichert, G.; Seufferlein, T. The Impact of Bioactive Lipids on Cardiovascular Development. Stem Cells Int. 2011, 2011, 916180. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Kim, J.S.; Chae, C.W.; Lim, J.R.; Kim, S.Y.; Lee, J.E.; Park, M.C.; Yoon, J.H.; et al. O-cyclic phytosphingosine-1-phosphate stimulates HIF1α-dependent glycolytic reprogramming to enhance the therapeutic potential of mesenchymal stem cells. Cell Death Dis. 2019, 10, 590. [Google Scholar] [CrossRef]
- Lim, J.J.; Kim, H.J.; Rhie, B.-H.; Lee, M.R.; Choi, M.J.; Hong, S.-H.; Kim, K.-S. Maintenance of hPSCs under Xeno-Free and Chemically Defined Culture Conditions. Int. J. Stem Cells 2019, 12, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Shradhanjali, A.; Riehl, B.D.; Lee, J.S.; Ha, L.; Lim, J.Y. Enhanced cardiomyogenic induction of mouse pluripotent cells by cyclic mechanical stretch. Biochem. Biophys. Res. Commun. 2017, 488, 590–595. [Google Scholar] [CrossRef]
- Sharma, A.; Zhang, Y.; Buikema, J.W.; Serpooshan, V.; Chirikian, O.; Kosaric, N.; Churko, J.M.; Dzilic, E.; Shieh, A.; Burridge, P.W.; et al. Stage-specific Effects of Bioactive Lipids on Human iPSC Cardiac Differentiation and Cardiomyocyte Proliferation. Sci. Rep. 2018, 8, 6618. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Khanna, A. Role of smad-and wnt-dependent pathways in embryonic cardiac development. Stem Cells Dev. 2006, 15, 29–39. [Google Scholar] [CrossRef]
- Higashi, K.; Matsuzaki, E.; Hashimoto, Y.; Takahashi-Yanaga, F.; Takano, A.; Anan, H.; Hirata, M.; Nishimura, F. Sphingosine-1-phosphate/S1PR2-mediated signaling triggers Smad1/5/8 phosphorylation and thereby induces Runx2 expression in osteoblasts. Bone 2016, 93, 1–11. [Google Scholar] [CrossRef]
- Gaussin, V.; Morley, G.E.; Cox, L.; Zwijsen, A.; Vance, K.M.; Emile, L.; Tian, Y.; Liu, J.; Hong, C.; Myers, D.; et al. Alk3/Bmpr1a Receptor Is Required for Development of the Atrioventricular Canal into Valves and Annulus Fibrosus. Circ. Res. 2005, 97, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Van Wijk, B.; Moorman, A.F.M.; van den Hoff, M.J.B. Role of bone morphogenetic proteins in cardiac differentiation. Cardiovasc. Res. 2007, 74, 244–255. [Google Scholar] [CrossRef]
- Mayeur, C.; Kolodziej, S.A.; Wang, A.; Xu, X.; Lee, A.; Yu, P.B.; Shen, J.; Bloch, K.D.; Bloch, D.B. Oral administration of a bone morphogenetic protein type I receptor inhibitor prevents the development of anemia of inflammation. Haematologica 2015, 100, e68–e71. [Google Scholar] [CrossRef] [Green Version]
- Kattman, S.J.; Witty, A.D.; Gagliardi, M.; Dubois, N.C.; Niapour, M.; Hotta, A.; Ellis, J.; Keller, G. Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 2011, 8, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Barron, M.; Gao, M.; Lough, J. Requirement for BMP and FGF signaling during cardiogenic induction in non-precardiac mesoderm is specific, transient, and cooperative. Dev. Dyn. 2000, 218, 383–393. [Google Scholar] [CrossRef]
- Dunn, K.K.; Palecek, S.P. Engineering Scalable Manufacturing of High-Quality Stem Cell-Derived Cardiomyocytes for Cardiac Tissue Repair. Front. Med. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, S.; Zhao, L.; Zhang, Y.; Li, Q.; Chai, X.; Zhang, Y. Resveratrol Enhances Cardiomyocyte Differentiation of Human Induced Pluripotent Stem Cells through Inhibiting Canonical WNT Signal Pathway and Enhancing Serum Response Factor-miR-1 Axis. Stem Cells Int. 2016, 2016, 2524092. [Google Scholar] [CrossRef] [Green Version]
- Minami, I.; Yamada, K.; Otsuji, T.G.; Yamamoto, T.; Shen, Y.; Otsuka, S.; Kadota, S.; Morone, N.; Barve, M.; Asai, Y.; et al. A Small Molecule that Promotes Cardiac Differentiation of Human Pluripotent Stem Cells under Defined, Cytokine- and Xeno-free Conditions. Cell Rep. 2012, 2, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Linask, K.L.; Mallon, B.; Johnson, K.; Klein, M.; Beers, J.; Xie, W.; Du, Y.; Liu, C.; Lai, Y.; et al. Heparin Promotes Cardiac Differentiation of Human Pluripotent Stem Cells in Chemically Defined Albumin-Free Medium, Enabling Consistent Manufacture of Cardiomyocytes. Stem Cells Transl. Med. 2017, 6, 527–538. [Google Scholar] [CrossRef]
- Qiu, X.X.; Liu, Y.; Zhang, Y.F.; Guan, Y.N.; Jia, Q.Q.; Wang, C.; Liang, H.; Li, Y.Q.; Yang, H.T.; Qin, Y.W.; et al. Rapamycin and CHIR99021 Coordinate Robust Cardiomyocyte Differentiation from Human Pluripotent Stem Cells via Reducing p53-Dependent Apoptosis. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, Y.; Ding, S. A chemical approach to stem-cell biology and regenerative medicine. Nature 2008, 453, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.; Mormeneo, D.; Long, J.S.; Delgado, A.; Pyne, N.J.; Pyne, S. Sphingosine 1-phosphate regulation of extracellular signal-regulated kinase-1/2 in embryonic stem cells. Stem Cells Dev. 2009, 18, 1319–1330. [Google Scholar] [CrossRef]
- Price, M.M.; Kapitonov, D.; Allegood, J.; Milstien, S.; Oskeritzian, C.A.; Spiegel, S. Sphingosine-1-phosphate induces development of functionally mature chymase-expressing human mast cells from hematopoietic progenitors. FASEB J. 2009, 23, 3506–3515. [Google Scholar] [CrossRef] [Green Version]
- Clay, H.; Wilsbacher, L.D.; Wilson, S.J.; Duong, D.N.; McDonald, M.; Lam, I.; Park, K.E.; Chun, J.; Coughlin, S.R. Sphingosine 1-phosphate receptor-1 in cardiomyocytes is required for normal cardiac development. Dev. Biol. 2016, 418, 157–165. [Google Scholar] [CrossRef]
- Gaengel, K.; Niaudet, C.; Hagikura, K.; Laviña, B.; Muhl, L.; Hofmann, J.J.; Ebarasi, L.; Nyström, S.; Rymo, S.; Chen, L.L.; et al. The sphingosine-1-phosphate receptor S1PR1 restricts sprouting angiogenesis by regulating the interplay between VE-cadherin and VEGFR2. Dev. Cell 2012, 23, 587–599. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Dong, A.; Karasaki, K.; Sogabe, S.; Okamoto, D.; Saigo, M.; Ishida, M.; Yoshizumi, M.; Kokubo, H. Smad4 regulates the nuclear translocation of Nkx2-5 in cardiac differentiation. Sci. Rep. 2021, 11, 3588. [Google Scholar] [CrossRef]
- Xu, J.; Gruber, P.J.; Chien, K.R. SMAD4 Is Essential for Human Cardiac Mesodermal Precursor Cell Formation. Stem Cells 2019, 37, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.O., 3rd; Chi, X.; Garcia-Gras, E.; Shirai, M.; Feng, X.H.; Schwartz, R.J. The cardiac determination factor, Nkx2-5, is activated by mutual cofactors GATA-4 and Smad1/4 via a novel upstream enhancer. J. Biol. Chem. 2004, 279, 10659–10669. [Google Scholar] [CrossRef] [Green Version]
- Xin, C.; Ren, S.; Kleuser, B.; Shabahang, S.; Eberhardt, W.; Radeke, H.; Schäfer-Korting, M.; Pfeilschifter, J.; Huwiler, A. Sphingosine 1-phosphate cross-activates the Smad signaling cascade and mimics transforming growth factor-beta-induced cell responses. J. Biol. Chem. 2004, 279, 35255–35262. [Google Scholar] [CrossRef] [Green Version]
- Perino, M.G.; Yamanaka, S.; Riordon, D.R.; Tarasova, Y.; Boheler, K.R. Ascorbic acid promotes cardiomyogenesis through SMAD1 signaling in differentiating mouse embryonic stem cells. PLoS ONE 2017, 12, e0188569. [Google Scholar] [CrossRef] [Green Version]
- Yuasa, S.; Fukuda, K. Multiple roles for BMP signaling in cardiac development. Drug Discov. Today Ther. Strateg. 2008, 5, 209–214. [Google Scholar] [CrossRef]
- Shahid, M.; Spagnolli, E.; Ernande, L.; Thoonen, R.; Kolodziej, S.A.; Leyton, P.A.; Cheng, J.; Tainsh, R.E.T.; Mayeur, C.; Rhee, D.K.; et al. BMP type I receptor ALK2 is required for angiotensin II-induced cardiac hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H984–H994. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Kohda, M.; Kanomata, K.; Nojima, J.; Nakamura, A.; Kamizono, J.; Noguchi, Y.; Iwakiri, K.; Kondo, T.; Kurose, J.; et al. Constitutively activated ALK2 and increased SMAD1/5 cooperatively induce bone morphogenetic protein signaling in fibrodysplasia ossificans progressiva. J. Biol. Chem. 2009, 284, 7149–7156. [Google Scholar] [CrossRef] [Green Version]
- Schille, C.; Heller, J.; Schambony, A. Differential requirement of bone morphogenetic protein receptors Ia (ALK3) and Ib (ALK6) in early embryonic patterning and neural crest development. BMC Dev. Biol. 2016, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.B.; Deng, D.Y.; Lai, C.S.; Hong, C.C.; Cuny, G.D.; Bouxsein, M.L.; Hong, D.W.; McManus, P.M.; Katagiri, T.; Sachidanandan, C.; et al. BMP type I receptor inhibition reduces heterotopic ossification. Nat. Med. 2008, 14, 1363–1369. [Google Scholar] [CrossRef]
- Wu, L.; Du, J.; Jing, X.; Yan, Y.; Deng, S.; Hao, Z.; She, Q. Bone morphogenetic protein 4 promotes the differentiation of Tbx18-positive epicardial progenitor cells to pacemaker-like cells. Exp. Ther. Med. 2019, 17, 2648–2656. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Catalogue No. | Dilution |
|---|---|---|
| OCT4 | Abcam (ab18976) | 1:1000 |
| BRACHYURY | Santa Cruz (sc-20109) | 1:1000 |
| MESP1 | Santa Cruz (sc-130461) | 1:1000 |
| NKX2.5 | Abcam (ab91196) | 1:1000 |
| TNNT2 | Invitrogen (MA5-12960) | 1:1000 |
| MLC2V | Proteintech (10906-1-AP) | 1:1000 |
| p-SMAD1/5/8 | Millipore (Ab3848-I) | 1:1000 |
| SMAD1/5/8 | Santa Cruz (sc-6031-R) | 1:1000 |
| GAPDH | Santa Cruz (sc-32233) | 1:10,000 |
| ROCK1 | Santa Cruz (sc-6055) | 1:1000 |
| RHOA | Abcam (ab187027) | 1:2000 |
| ALK2 | Santa Cruz (sc-374523) | 1:1000 |
| ALK3 | R&D Systems (AF346) | 1:200 |
| p-Erk | Cell Signaling (#9101) | 1:1000 |
| Erk | Cell Signaling (#9102) | 1:1000 |
| Gene Name | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| NKX2.5 | AGTGTGCGTCTGCCTTTC | GTTGTCCGCCTCTGTCTTC |
| MEF2C | ATGGATGAACGTAACAGACAGGT | CGGCTCGTTGTACTCCGTG |
| TBX5 | TTGCATGTATGCCAGCTCTG | CTGGTAGGGTAGCCTGTCC |
| GATA4 | AGATGGGACGGGTCACTATC | CAGTTGGCACAGGAGAGG |
| TNNT2 | AGCGGAAAAGTGGGAAGAG | TCCAAGTTATAGATGCTCTGCC |
| GAPDH | GTCATCCCTGAGCTGAACGG | CCACCTGGTGCTCAGTGTAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, J.-H.; Kim, M.-S.; Antao, A.M.; Jo, W.-J.; Kim, H.-J.; Kim, S.-J.; Choi, M.-J.; Ramakrishna, S.; Kim, K.-S. Bioactive Lipid O-cyclic phytosphingosine-1-phosphate Promotes Differentiation of Human Embryonic Stem Cells into Cardiomyocytes via ALK3/BMPR Signaling. Int. J. Mol. Sci. 2021, 22, 7015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137015
Jang J-H, Kim M-S, Antao AM, Jo W-J, Kim H-J, Kim S-J, Choi M-J, Ramakrishna S, Kim K-S. Bioactive Lipid O-cyclic phytosphingosine-1-phosphate Promotes Differentiation of Human Embryonic Stem Cells into Cardiomyocytes via ALK3/BMPR Signaling. International Journal of Molecular Sciences. 2021; 22(13):7015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137015
Chicago/Turabian StyleJang, Ji-Hye, Min-Seong Kim, Ainsley Mike Antao, Won-Jun Jo, Hyung-Joon Kim, Su-Jin Kim, Myeong-Jun Choi, Suresh Ramakrishna, and Kye-Seong Kim. 2021. "Bioactive Lipid O-cyclic phytosphingosine-1-phosphate Promotes Differentiation of Human Embryonic Stem Cells into Cardiomyocytes via ALK3/BMPR Signaling" International Journal of Molecular Sciences 22, no. 13: 7015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137015