
Bioluminescent Optogenetics: A Novel Experimental Therapy to Promote Axon Regeneration after Peripheral Nerve Injury

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
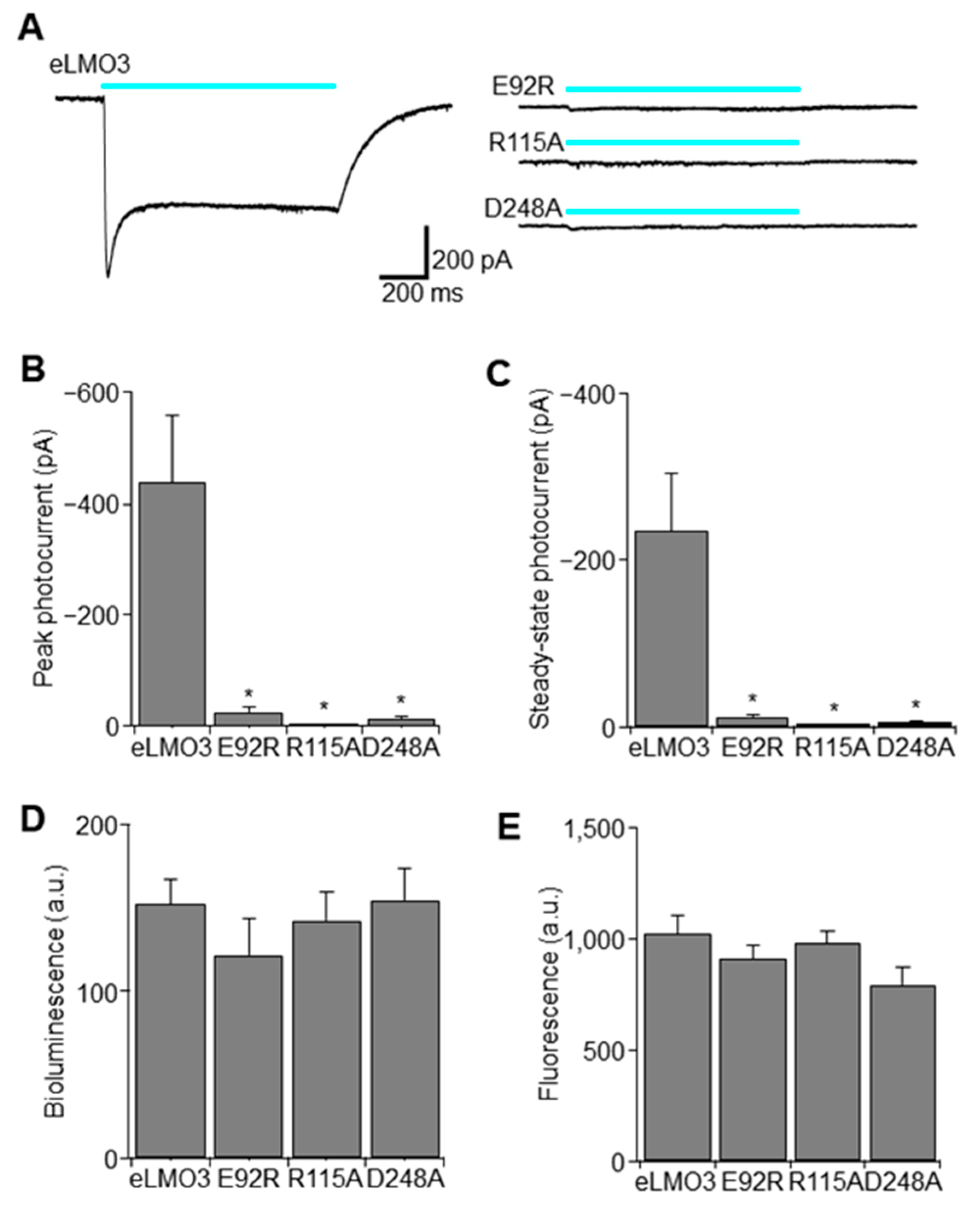
2.1. Point Mutation Renders Luminopsin Non-Functional without Compromising Expression Levels
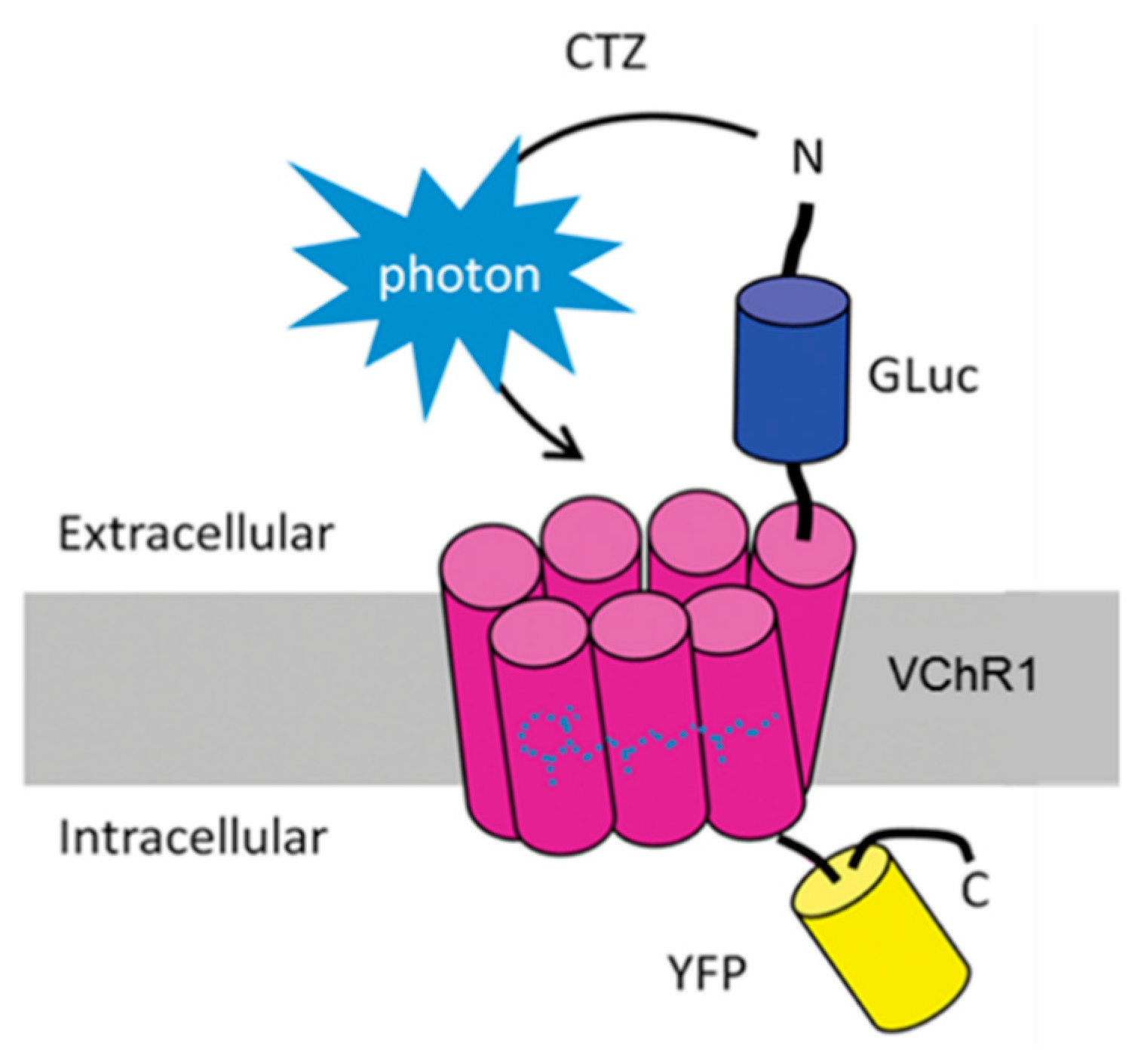
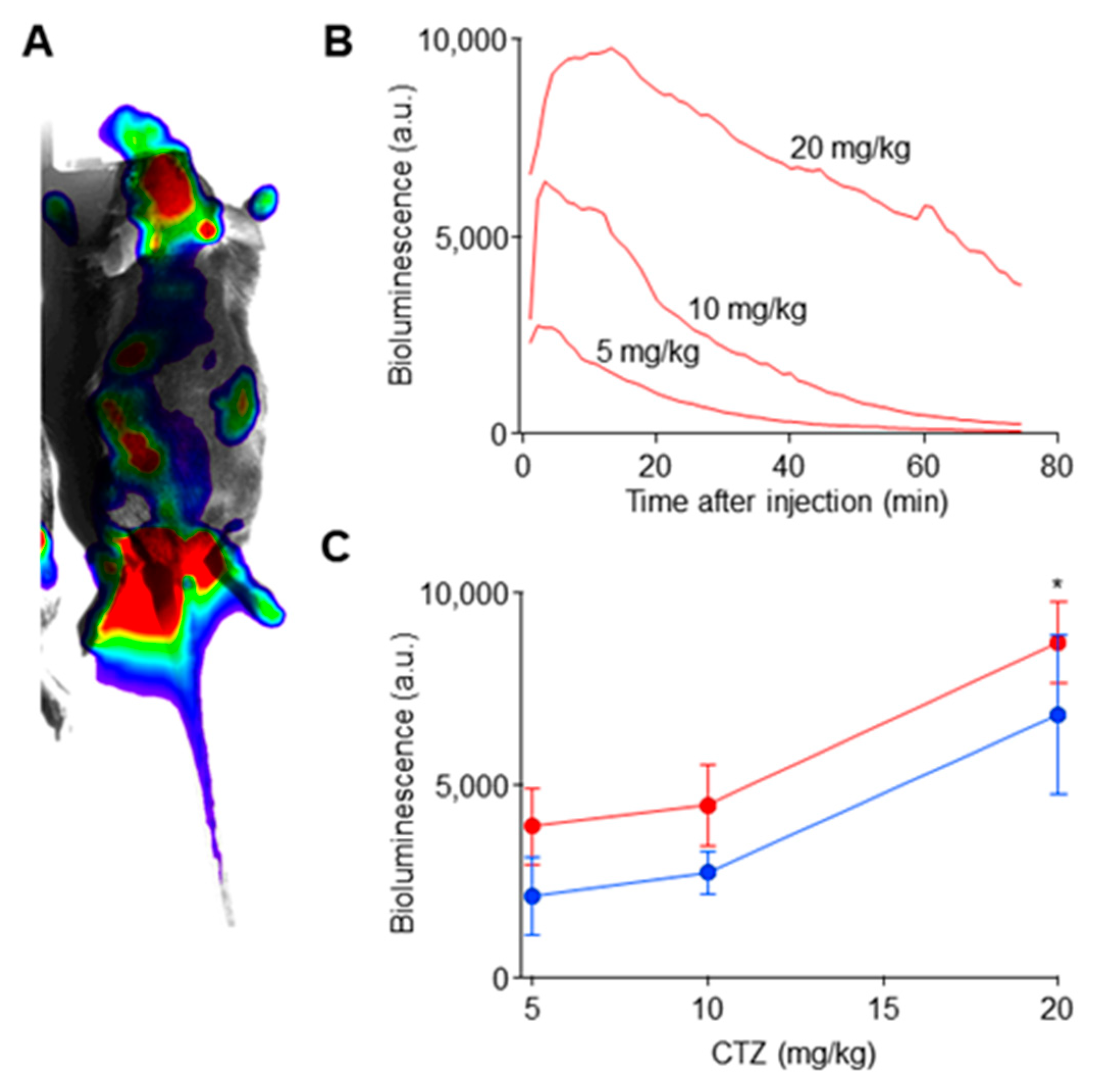
2.2. Coelenterazine Produces Bioluminescence in Luminopsin Expressing Nerves
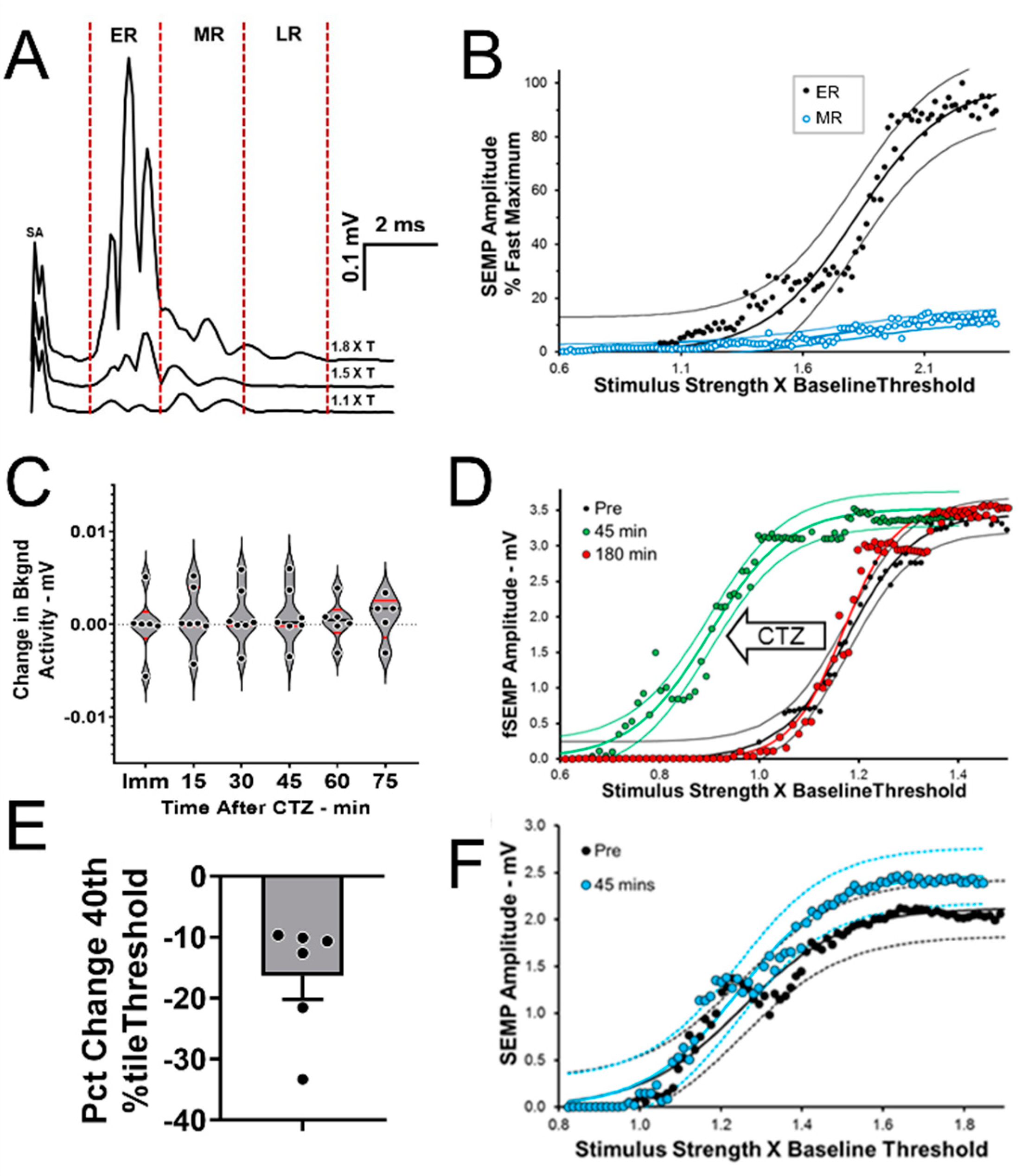
2.3. BL-OG Treatments Increase Motoneuron Excitability
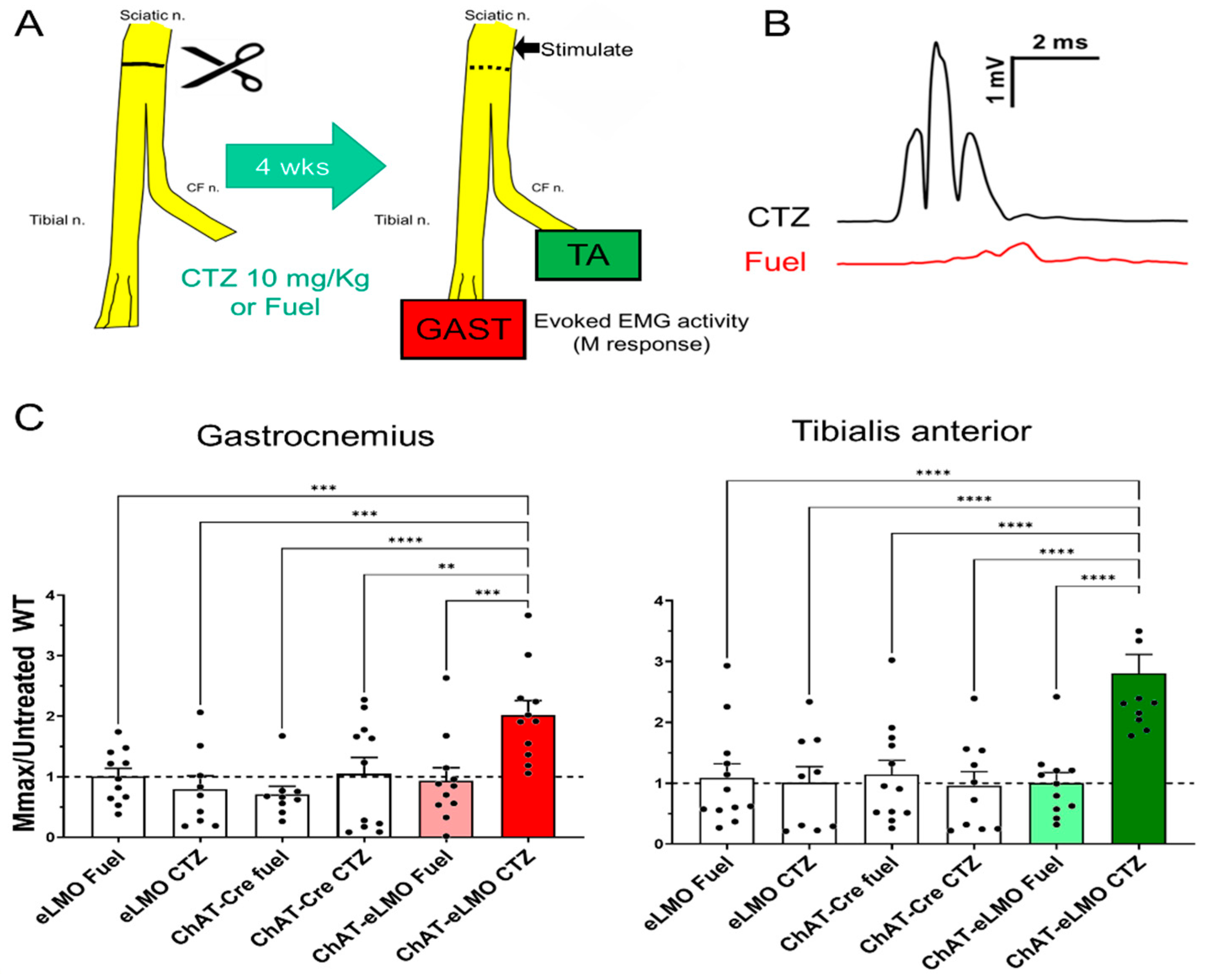
2.4. Effects of BL-OG Treatment on Axon Regeneration in Transgenic Mice
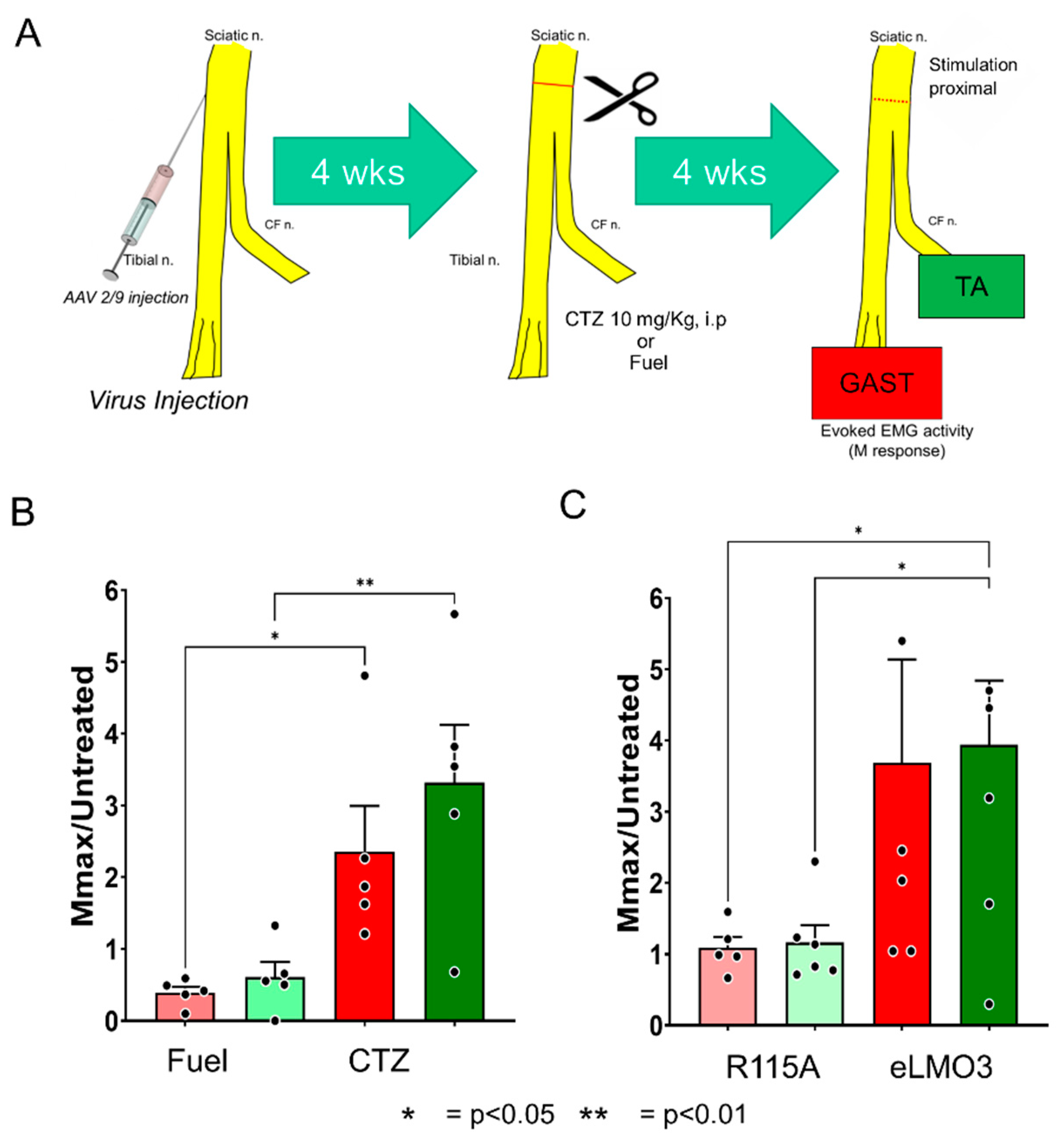
2.5. Effects of BL-OG Treatment on Axon Regeneration after Viral eLMO3 Transduction
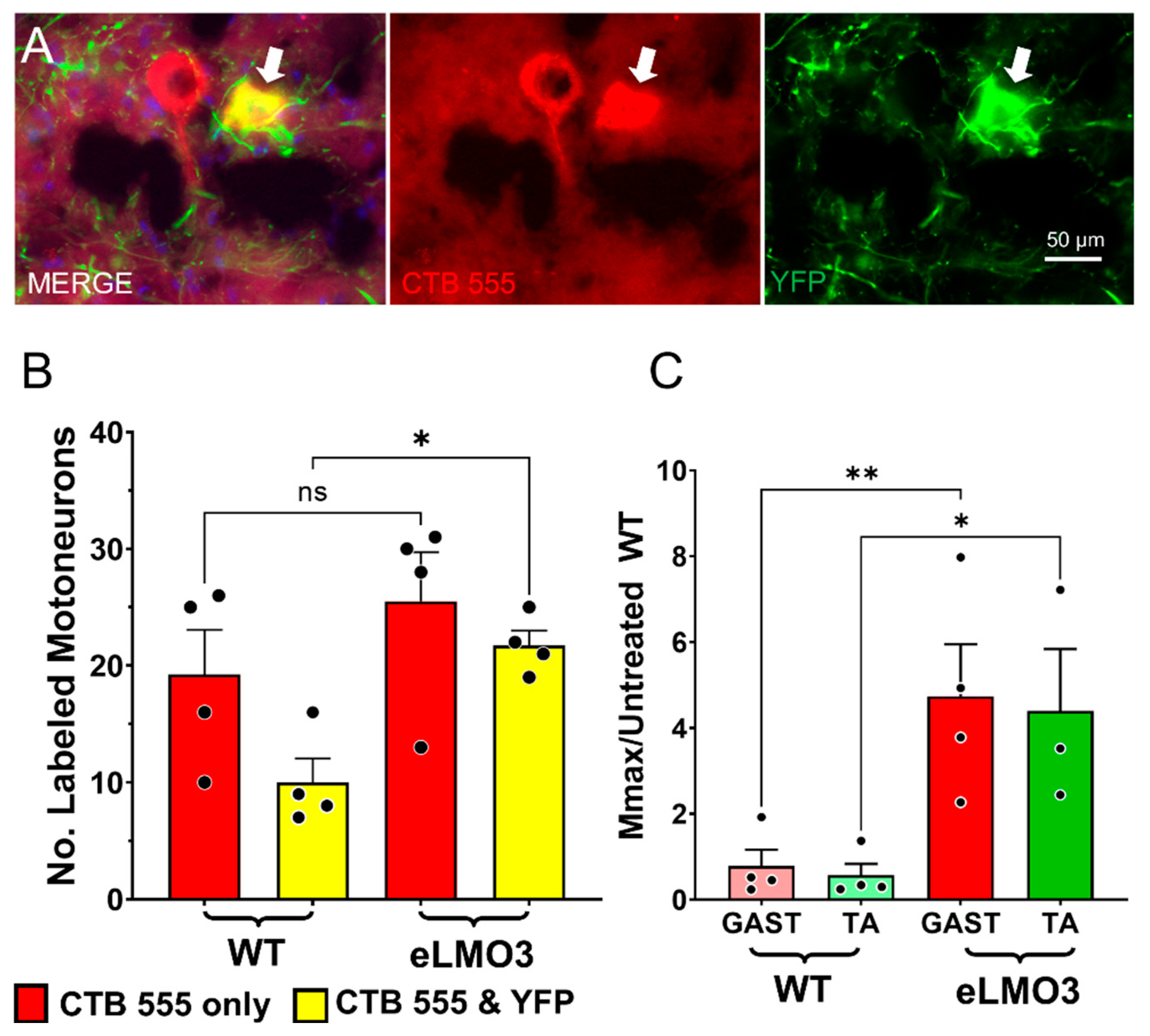
2.6. Retrograde Labeling
3. Discussion
4. Materials and Methods
4.1. Molecular Biology
4.2. Tissue Culture and Virus Production
4.3. Electrophysiology
4.4. Plate-reader Assays
4.5. Animals and Luminopsin Expression
4.6. Bioluminescence Experiments
4.7. Motoneuron Excitability Experiments
4.8. Nerve Transection Surgery
4.9. M Response Recovery
4.10. Retrograde Labeling
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.K.; Wolfe, S.W. Peripheral nerve injury and repair. J. Am. Acad. Orthop. Surg. 2000, 8, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sunitha, M.; Chung, K.C. How to measure outcomes of peripheral nerve surgery. Hand Clin. 2013, 29, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, T.; Chan, K.M.; Sulaiman, O.A.; Udina, E.; Amirjani, N.; Brushart, T.M. Accelerating axon growth to overcome limitations in functional recovery after peripheral nerve injury. Neurosurgery 2009, 65 (Suppl. 4), A132–A144. [Google Scholar] [CrossRef]
- Fu, S.Y.; Gordon, T. Contributing factors to poor functional recovery after delayed nerve repair: Prolonged denervation. J. Neurosci. 1995, 15, 3886–3895. [Google Scholar] [CrossRef]
- Fu, S.Y.; Gordon, T. Contributing factors to poor functional recovery after delayed nerve repair: Prolonged axotomy. J. Neurosci. 1995, 15, 3876–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, T.; English, A.W. Strategies to promote peripheral nerve regeneration: Electrical stimulation and/or exercise. Eur. J. Neurosci. 2016, 43, 336–350. [Google Scholar] [CrossRef] [Green Version]
- Udina, E.; Cobianchi, S.; Allodi, I.; Navarro, X. Effects of activity-dependent strategies on regeneration and plasticity after peripheral nerve injuries. Ann. Anat. 2011, 193, 347–353. [Google Scholar] [CrossRef]
- Berglund, K.; Tung, J.K.; Higashikubo, B.; Gross, R.E.; Moore, C.I.; Hochgeschwender, U. Combined Optogenetic and Chemogenetic Control of Neurons. Methods Mol. Biol. 2016, 1408, 207–225. [Google Scholar]
- Gomez-Ramirez, M.; More, A.I.; Friedman, N.G.; Hochgeschwender, U.; Moore, C.I. The BioLuminescent-OptoGenetic in vivo response to coelenterazine is proportional, sensitive, and specific in neocortex. J. Neurosci. Res. 2020, 98, 471–480. [Google Scholar] [CrossRef]
- Mistretta, O.C.; Lopez, A.; Wynans, C.; Berglund, K.; English, A.W. Use of bioluminescent optogenetics (BL-OG) to enhance motor axon regeneration after peripheral nerve transection and repair. Abstr. Soc. Neurosci. 2019. [Google Scholar]
- Tanimoto, S.; Sugiyama, Y.; Takahashi, T.; Ishizuka, T.; Yawo, H. Involvement of glutamate 97 in ion influx through photo-activated channelrhodopsin-2. Neurosci. Res. 2013, 75, 13–22. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Tung, J.K.; Wang, Z.; Yu, S.P.; Gross, R.E.; Wei, L.; Berglund, K. Improved trafficking and expression of luminopsins for more efficient optical and pharmacological control of neuronal activity. J. Neurosci. Res. 2020, 98, 481–490. [Google Scholar] [CrossRef]
- Li, X.; Yu, B.; Sun, Q.; Zhang, Y.; Ren, M.; Zhang, X.; Li, A.; Yuan, J.; Madisen, L.; Luo, Q.; et al. Generation of a whole-brain atlas for the cholinergic system and mesoscopic projectome analysis of basal forebrain cholinergic neurons. Proc. Natl. Acad. Sci. USA 2018, 115, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Gerasimenko, Y.P.; Lavrov, I.A.; Courtine, G.; Ichiyama, R.M.; Dy, C.J.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Spinal cord reflexes induced by epidural spinal cord stimulation in normal awake rats. J. Neurosci. Methods 2006, 157, 253–263. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Gorodnichev, R.; Moshonkina, T.; Sayenko, D.; Gad, P.; Reggie Edgerton, V. Transcutaneous electrical spinal-cord stimulation in humans. Ann. Phys. Rehabil. Med. 2015, 58, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayenko, D.G.; Angeli, C.; Harkema, S.J.; Edgerton, V.R.; Gerasimenko, Y.P. Neuromodulation of evoked muscle potentials induced by epidural spinal-cord stimulation in paralyzed individuals. J. Neurophysiol. 2014, 111, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Liu, C.Y.; Ward, P.J.; Jaiswal, P.B.; English, A.W. Effects of Repeated 20-Hz Electrical Stimulation on Functional Recovery Following Peripheral Nerve Injury. Neurorehabil. Neural Repair 2019, 33, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, M.C.; Bostock, H.; Park, S.B.; Kaji, R.; Krarup, C.; Krishnan, A.V.; Kuwabara, S.; Lin, C.S.; Misawa, S.; Moldovan, M.; et al. Measurement of axonal excitability: Consensus guidelines. Clin. Neurophysiol. 2020, 131, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, M.; Redmon, N.; Schwartz, G.; English, A. Treadmill training promotes axon regeneration in injured peripheral nerves. Exp. Neurol. 2008, 211, 489–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, K.; Wilhelm, J.C.; Sabatier, M.J.; English, A.W. Sex differences in the effects of treadmill training on axon regeneration in cut peripheral nerves. Dev. Neurobiol. 2012, 72, 688–698. [Google Scholar] [CrossRef]
- Jaiswal, P.B.; Tung, J.K.; Gross, R.E.; English, A.W. Motoneuron activity is required for enhancements in functional recovery after peripheral nerve injury in exercised female mice. J. Neurosci. Res. 2020, 98, 448–457. [Google Scholar] [CrossRef]
- Ward, P.J.; English, A.W. Optical Stimulation and Electrophysiological Analysis of Regenerating Peripheral Axons. Bio-Protocol 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.B.; Mistretta, O.C.; Ward, P.J.; English, A.W. Chemogenetic Enhancement of Axon Regeneration Following Peripheral Nerve Injury in the SLICK-A Mouse. Brain Sci. 2018, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, F.J.; Rotterman, T.M.; Akhter, E.T.; Lane, A.R.; English, A.W.; Cope, T.C. Synaptic Plasticity on Motoneurons After Axotomy: A Necessary Change in Paradigm. Front. Mol. Neurosci. 2020, 13, 68. [Google Scholar] [CrossRef] [PubMed]
- Berglund, K.; Clissold, K.; Li, H.E.; Wen, L.; Park, S.Y.; Gleixner, J.; Klein, M.E.; Lu, D.; Barter, J.W.; Rossi, M.A.; et al. Luminopsins integrate opto- and chemogenetics by using physical and biological light sources for opsin activation. Proc. Natl. Acad. Sci. USA 2016, 113, E358–E367. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.P.; Tung, J.K.; Wei, Z.Z.; Chen, D.; Berglund, K.; Zhong, W.; Zhang, J.Y.; Gu, X.; Song, M.; Gross, R.E.; et al. Optochemogenetic Stimulation of Transplanted iPS-NPCs Enhances Neuronal Repair and Functional Recovery after Ischemic Stroke. J. Neurosci. 2019, 39, 6571–6594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothman, J.S.; Silver, R.A. NeuroMatic: An Integrated Open-Source Software Toolkit for Acquisition, Analysis and Simulation of Electrophysiological Data. Front. Neuroinform. 2018, 12, 14. [Google Scholar] [CrossRef]
- Medendorp, W.E.; Bjorefeldt, A.; Crespo, E.L.; Prakash, M.; Pal, A.; Waddell, M.L.; Moore, C.I.; Hochgeschwender, U. Selective postnatal excitation of neocortical pyramidal neurons results in distinctive behavioral and circuit deficits in adulthood. iScience 2021, 24, 102157. [Google Scholar] [CrossRef]
- Ward, P.J.; Clanton, S.L., 2nd; English, A.W. Optogenetically enhanced axon regeneration: Motor versus sensory neuron-specific stimulation. Eur J. Neurosci. 2018, 47, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Young, P.; Qiu, L.; Wang, D.; Zhao, S.; Gross, J.; Feng, G. Single-neuron labeling with inducible Cre-mediated knockout in transgenic mice. Nat. Neurosci. 2008, 11, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, J.C.; Cucoranu, D.; Xu, M.; Chmielewski, S.; Holmes, T.; Lau, K.S.; Bassell, G.J.; English, A.W. Cooperative roles of BDNF expression in neurons and Schwann cells are modulated by exercise to facilitate nerve regeneration. J. Neurosci. 2012, 32, 5002–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basmajian, J.V.; Stecko, G. The Role of Muscles in Arch Support of the Foot. J. Bone Jt. Surg. Am. 1963, 45, 1184–1190. [Google Scholar] [CrossRef]
- MacGillivray, T.E. Fibrin Sealants and Glues. J. Card. Surg. 2003, 18, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Akhter, E.T.; Rotterman, T.M.; English, A.W.; Alvarez, F.J. Sciatic Nerve Cut and Repair Using Fibrin Glue in Adult Mice. Bio-Protocol 2019, 9. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
English, A.W.; Berglund, K.; Carrasco, D.; Goebel, K.; Gross, R.E.; Isaacson, R.; Mistretta, O.C.; Wynans, C. Bioluminescent Optogenetics: A Novel Experimental Therapy to Promote Axon Regeneration after Peripheral Nerve Injury. Int. J. Mol. Sci. 2021, 22, 7217. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137217
English AW, Berglund K, Carrasco D, Goebel K, Gross RE, Isaacson R, Mistretta OC, Wynans C. Bioluminescent Optogenetics: A Novel Experimental Therapy to Promote Axon Regeneration after Peripheral Nerve Injury. International Journal of Molecular Sciences. 2021; 22(13):7217. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137217
Chicago/Turabian StyleEnglish, Arthur W., Ken Berglund, Dario Carrasco, Katharina Goebel, Robert E. Gross, Robin Isaacson, Olivia C. Mistretta, and Carly Wynans. 2021. "Bioluminescent Optogenetics: A Novel Experimental Therapy to Promote Axon Regeneration after Peripheral Nerve Injury" International Journal of Molecular Sciences 22, no. 13: 7217. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137217