
Saccharomyces cerevisiae, a Powerful Model for Studying rRNA Modifications and Their Effects on Translation Fidelity


Abstract
:1. Introduction
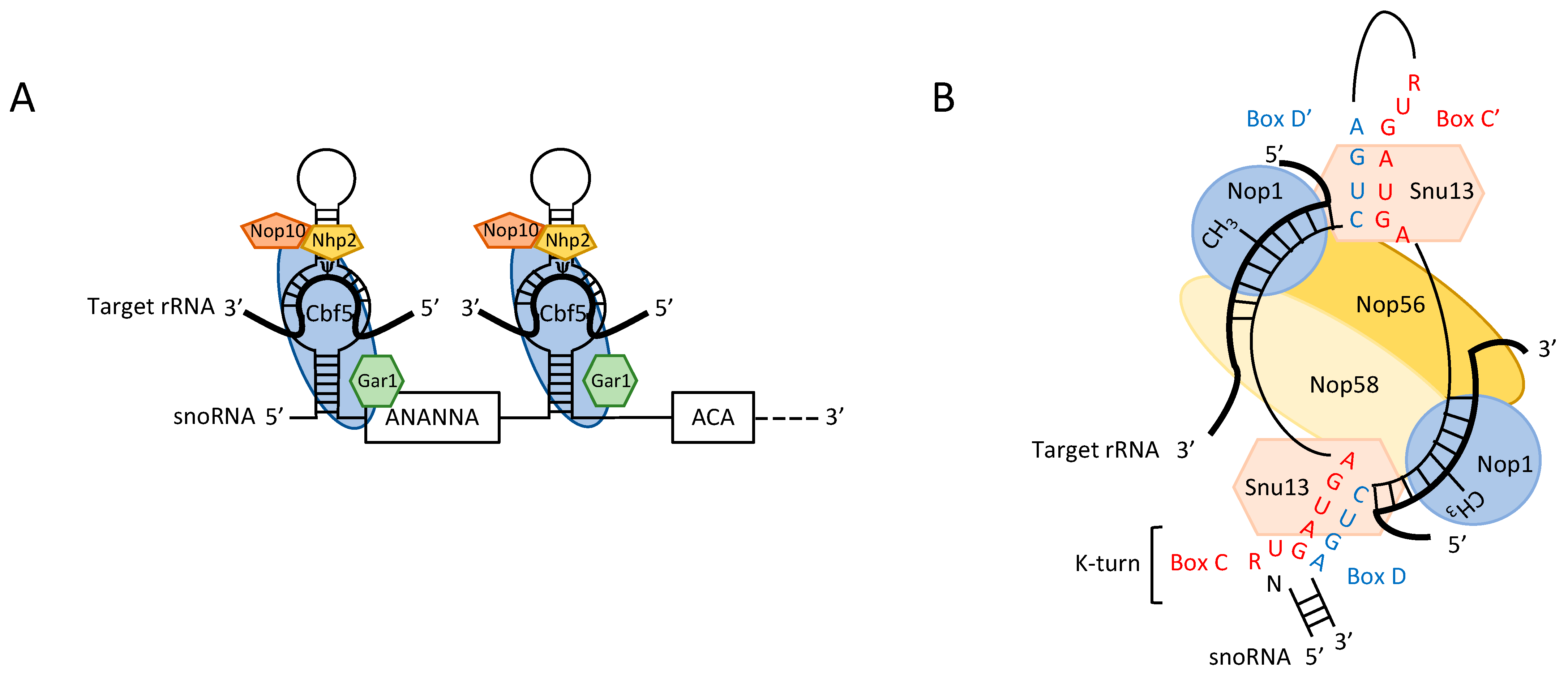
2. RNA-Guided Modifications
3. SnoRNA Engineering
4. Detection of rRNA Modifications
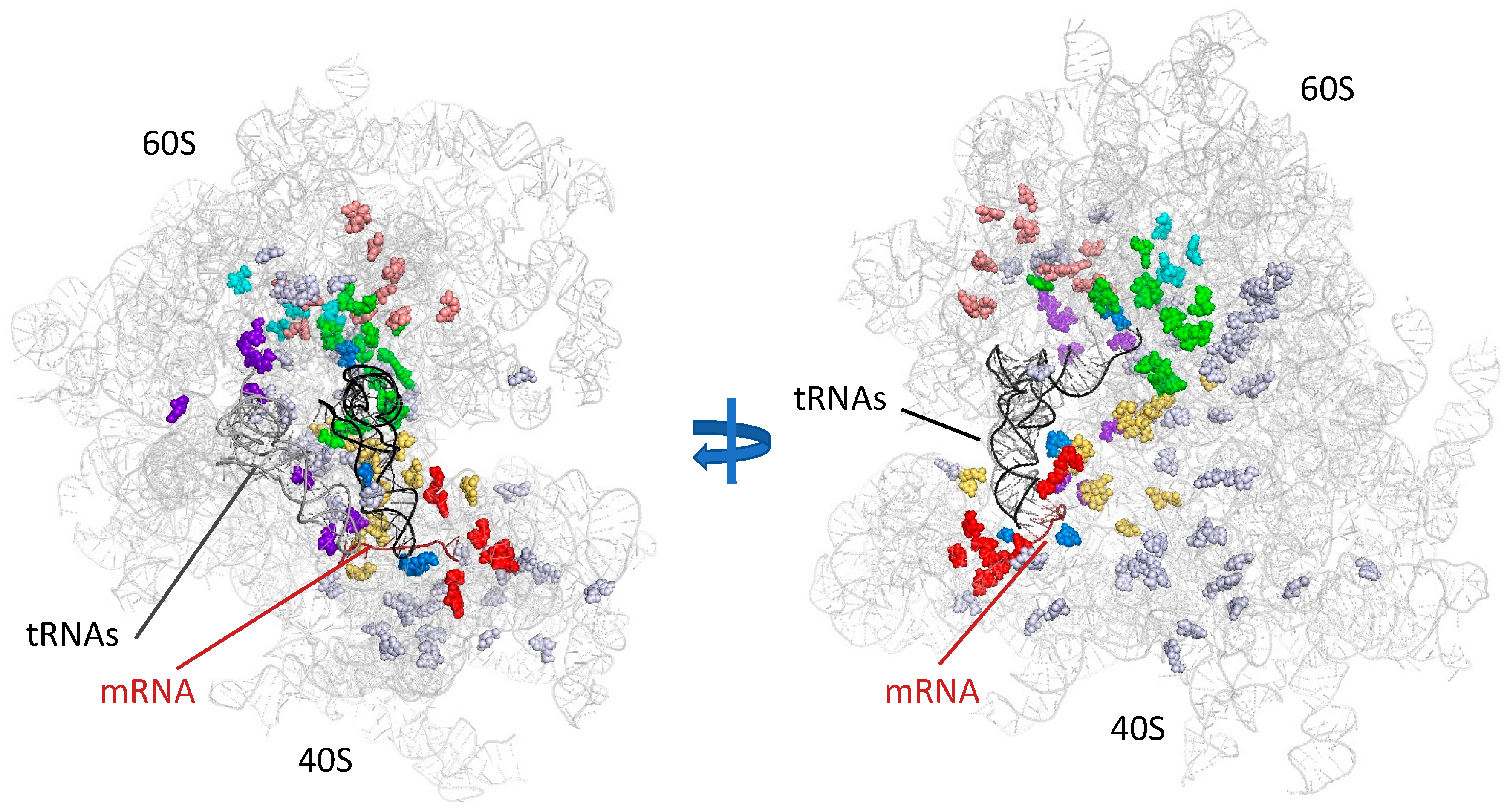
5. Function in Translation Fidelity
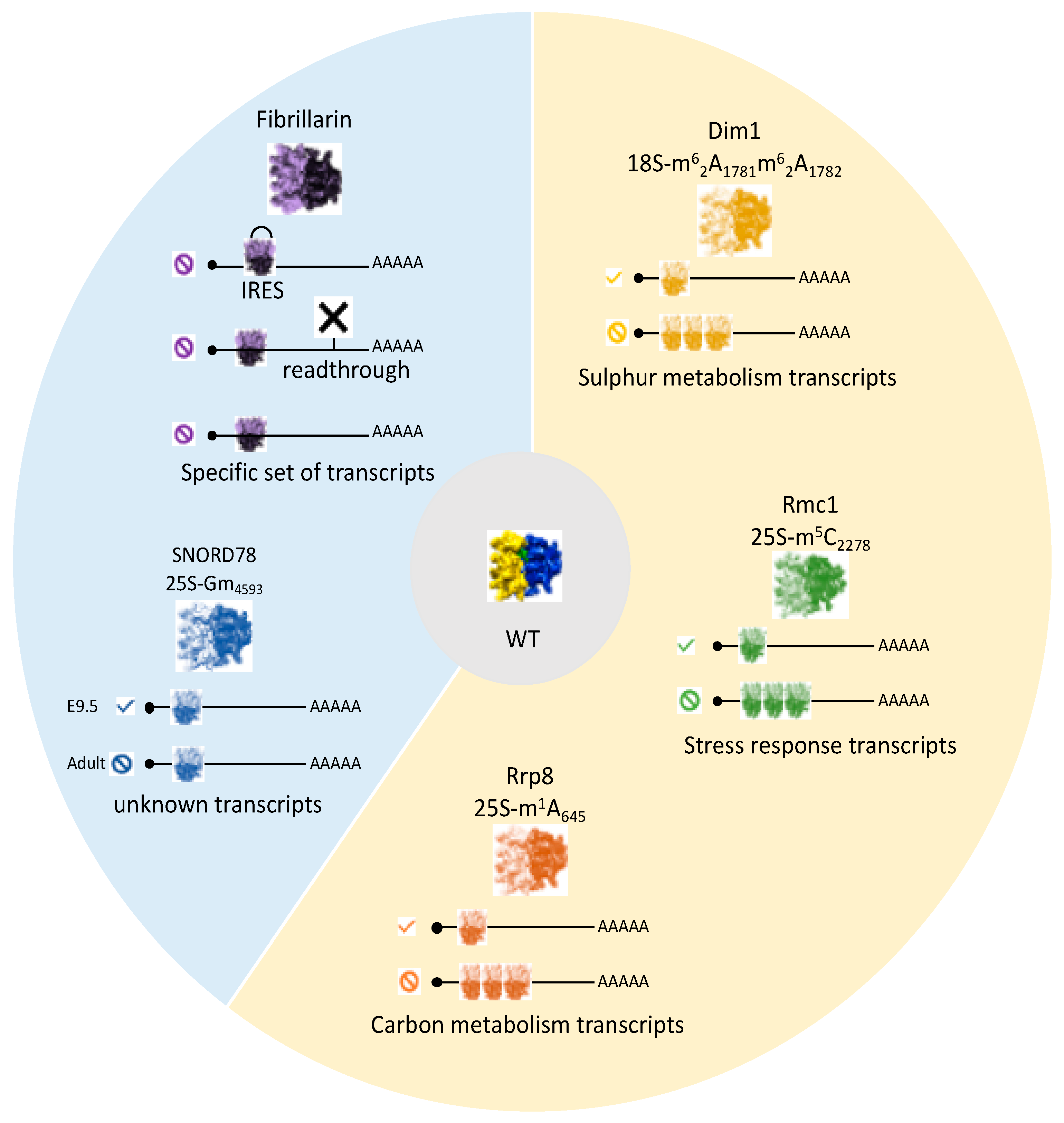
6. Specialized Ribosomes
7. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woolford, J.L.; Baserga, S.J. Ribosome Biogenesis in the Yeast Saccharomyces cerevisiae. Genetics 2013, 195, 643–681. [Google Scholar] [CrossRef] [Green Version]
- Bachellerie, J.P.; Michot, B.; Nicoloso, M.; Balakin, A.; Ni, J.; Fournier, M.J. Antisense snoRNAs: A family of nucleolar RNAs with long complementarities to rRNA. Trends Biochem. Sci. 1995, 20, 261–264. [Google Scholar] [CrossRef]
- Maxwell, E.S.; Fournier, M.J. The Small Nucleolar RNAs. Annu. Rev. Biochem. 1995, 64, 897–934. [Google Scholar] [CrossRef]
- Cavaille, J.; Nicoloso, M.; Bachellerie, J.P. Targeted ribose methylation of RNA in vivo directed by tailored an-tisense RNA guides. Nature 1996, 383, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Kiss-László, Z.; Henry, Y.; Bachellerie, J.-P.; Caizergues-Ferrer, M.; Kiss, T. Site-Specific Ribose Methylation of Preribosomal RNA: A Novel Function for Small Nucleolar RNAs. Cell 1996, 85, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Bortolin, M.; Ganot, P.; Kiss, T. Elements essential for accumulation and function of small nucleolar RNAs directing site-specific pseudouridylation of ribosomal RNAs. EMBO J. 1999, 18, 457–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganot, P.; Bortolin, M.-L.; Kiss, T. Site-Specific Pseudouridine Formation in Preribosomal RNA Is Guided by Small Nucleolar RNAs. Cell 1997, 89, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Balakin, A.G.; Smith, L.; Fournier, M.J. The RNA world of the nucleolus: Two major families of small RNAs defined by different box elements with related functions. Cell 1996, 86, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Chagot, M.-E.; Quinternet, M.; Rothé, B.; Charpentier, B.; Coutant, J.; Manival, X.; Lebars, I. The yeast C/D box snoRNA U14 adopts a “weak” K-turn like conformation recognized by the Snu13 core protein in solution. Biochimie 2019, 164, 70–82. [Google Scholar] [CrossRef]
- Sharma, S.; Yang, J.; Van Nues, R.; Watzinger, P.; Kötter, P.; Lafontaine, D.; Granneman, S.; Entian, K.-D. Specialized box C/D snoRNPs act as antisense guides to target RNA base acetylation. PLoS Genet. 2017, 13, e1006804. [Google Scholar] [CrossRef]
- Piekna-Przybylska, D.; Decatur, W.A.; Fournier, M.J. New bioinformatic tools for analysis of nucleotide modifications in eukaryotic rRNA. RNA 2007, 13, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Tiotiu, D.; Bragantini, B.; Marty, H.; Charpentier, B.; Massenet, S.; Labialle, S. Bcd1p controls RNA loading of the core protein Nop58 during C/D box snoRNP biogenesis. RNA 2019, 25, 496–506. [Google Scholar] [CrossRef]
- Grzechnik, P.; Szczepaniak, S.A.; Dhir, S.; Pastucha, A.; Parslow, H.; Matuszek, Z.; Mischo, H.E.; Kufel, J.; Proudfoot, N.J. Nuclear fate of yeast snoRNA is determined by co-transcriptional Rnt1 cleavage. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Massenet, S.; Bertrand, E.; Verheggen, C. Assembly and trafficking of box C/D and H/ACA snoRNPs. RNA Biol. 2017, 14, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Bonnerot, C.; Pintard, L.; Lutfalla, G. Functional Redundancy of Spb1p and a snR52-Dependent Mechanism for the 2′-O-Ribose Methylation of a Conserved rRNA Position in Yeast. Mol. Cell 2003, 12, 1309–1315. [Google Scholar] [CrossRef]
- Schmitt, H.G. Granulation tumor in the horse following barbed wire injury: X-ray therapy. Tierarztl. Prax. 1977, 5, 219–221. [Google Scholar]
- Ni, J.; Samarsky, D.; Liu, B.; Ferbeyre, G.; Cedergren, R.; Fournier, M.J. SnoRNAs as tools for RNA cleavage and modification. Nucleic Acids Symp. Ser. 1997, 36, 61–63. [Google Scholar]
- Liu, B.; Ni, J.; Fournier, M.J. Probing RNA in vivo with methylation guide small nucleolar RNAs. Methods 2001, 23, 276–286. [Google Scholar] [CrossRef]
- Liu, B.; Fournier, M.J. Interference probing of rRNA with snoRNPs: A novel approach for functional mapping of RNA in vivo. RNA 2004, 10, 1130–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Liang, X.-H.; Piekna-Przybylska, R.; Liu, Q.; Fournier, M.J. Mis-targeted methylation in rRNA can severely impair ribosome synthesis and activity. RNA Biol. 2008, 5, 249–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karijolich, J.; Yu, Y.-T. Converting nonsense codons into sense codons by targeted pseudouridylation. Nature 2011, 474, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Keith, G. Mobilities of modified ribonucleotides on two-dimensional cellulose thin-layer chromatography. Biochimie 1995, 77, 142–144. [Google Scholar] [CrossRef]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.-D.; Lafontaine, D.; Bohnsack, M.T. Tuning the ribosome: The influence of rRNA modification on eukaryotic ribosome biogenesis and function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Su, D.; Chan, C.T.Y.; Gu, C.; Lim, K.S.; Chionh, Y.H.; McBee, M.E.; Russell, B.S.; Babu, I.R.; Begley, T.J.; Dedon, P.C. Quantitative analysis of ribonucleoside modifications in tRNA by HPLC-coupled mass spectrometry. Nat. Protoc. 2014, 9, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jia, G. Detection methods of epitranscriptomic mark N6-methyladenosine. Essays Biochem. 2020, 64, 967–979. [Google Scholar] [PubMed]
- Marchand, V.; Pichot, F.; Neybecker, P.; Ayadi, L.; Bourguignon-Igel, V.; Wacheul, L.; Lafontaine, D.L.J.; Pinzano, A.; Helm, M.; Motorin, Y. HydraPsiSeq: A method for systematic and quantitative mapping of pseudouridines in RNA. Nucleic Acids Res. 2020, 48, e110. [Google Scholar] [CrossRef]
- Krogh, N.; Nielsen, H. Sequencing-based methods for detection and quantitation of ribose methylations in RNA. Methods 2019, 156, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Angelova, M.T.; Dimitrova, D.G.; Da Silva, B.; Marchand, V.; Jacquier, C.; Achour, C.; Brazane, M.; Goyenvalle, C.; Bourguignon-Igel, V.; Shehzada, S.; et al. tRNA 2′-O-methylation by a duo of TRM7/FTSJ1 proteins modulates small RNA silencing in Drosophila. Nucleic Acids Res. 2020, 48, 2050–2072. [Google Scholar] [CrossRef] [Green Version]
- Marchand, V.; Pichot, F.; Thuring, K.; Ayadi, L.; Freund, I.; Dalpke, A.; Helm, M.; Motorin, Y. Next-Generation Sequencing-Based RiboMethSeq Protocol for Analysis of tRNA 2′-O-Methylation. Biomolecules 2017, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, V.; Blanloeil-Oillo, F.; Helm, M.; Motorin, Y. Illumina-based RiboMethSeq approach for mapping of 2′-O-Me residues in RNA. Nucleic Acids Res. 2016, 44, e135. [Google Scholar] [CrossRef] [Green Version]
- Krogh, N.; Jansson, M.; Häfner, S.; Tehler, D.E.; Birkedal, U.; Christensen-Dalsgaard, M.; Lund, A.H.; Nielsen, H. Profiling of 2′-O-Me in human rRNA reveals a subset of fractionally modified positions and provides evidence for ribosome heterogeneity. Nucleic Acids Res. 2016, 44, 7884–7895. [Google Scholar] [CrossRef] [Green Version]
- Marchand, V.; Ayadi, L.; Ernst, F.G.M.; Hertler, J.; Bourguignon-Igel, V.; Galvanin, A.; Kotter, A.; Helm, M.; Lafontaine, D.L.J.; Motorin, Y. AlkAniline-Seq: Profiling of m(7) G and m(3) C RNA Modifications at Single Nucleotide Resolution. Angew. Chem. 2018, 57, 16785–16790. [Google Scholar] [CrossRef]
- Zheng, H.-X.; Zhang, X.-S.; Sui, N. Advances in the profiling of N6-methyladenosine (m6A) modifications. Biotechnol. Adv. 2020, 45, 107656. [Google Scholar] [CrossRef]
- Liu, H.; Begik, O.; Lucas, M.C.; Ramírez, J.M.; Mason, C.E.; Wiener, D.; Schwartz, S.; Mattick, J.; Smith, M.A.; Novoa, E.M. Accurate detection of m6A RNA modifications in native RNA sequences. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Jain, M.; Mulroney, L.; Garalde, D.R.; Akeson, M. Reading canonical and modified nucleobases in 16S ribosomal RNA using nanopore native RNA sequencing. PLoS ONE 2019, 14, e0216709. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Olsen, H.; Akeson, M.; Abu-Shumays, R. Adaptation of Human Ribosomal RNA for Nanopore Sequencing of Canonical and Modified Nucleotides. In RNA Modifications; Humana: New York, NY, USA, 2021; pp. 53–74. [Google Scholar]
- Jenjaroenpun, P.; Wongsurawat, T.; Wadley, T.D.; Wassenaar, T.M.; Liu, J.; Dai, Q.; Wanchai, V.; Akel, N.S.; Jamshidi-Parsian, A.; Franco, A.T.; et al. Decoding the epitranscriptional landscape from native RNA sequences. Nucleic Acids Res. 2021, 49, e7. [Google Scholar] [CrossRef] [PubMed]
- Charette, M.; Gray, M.W. Pseudouridine in RNA: What, where, how, and why. IUBMB Life 2000, 49, 341–351. [Google Scholar]
- Assi, H.A.; Rangadurai, A.K.; Shi, H.; Liu, B.; Clay, M.C.; Erharter, K.; Kreutz, C.; Holley, C.L.; Al-Hashimi, H.M. 2′-O-Methylation can increase the abundance and lifetime of alternative RNA conformational states. Nucleic Acids Res. 2020, 48, 12365–12379. [Google Scholar] [PubMed]
- Elliott, B.A.; Ho, H.-T.; Ranganathan, S.V.; Vangaveti, S.; Ilkayeva, O.; Assi, H.A.; Choi, A.K.; Agris, P.F.; Holley, C.L. Modification of messenger RNA by 2′-O-methylation regulates gene expression in vivo. Nat. Commun. 2019, 10, 1–9. [Google Scholar]
- Decatur, W.; Fournier, M.J. rRNA modifications and ribosome function. Trends Biochem. Sci. 2002, 27, 344–351. [Google Scholar] [CrossRef]
- Tollervey, D.; Lehtonen, H.; Jansen, R.; Kern, H.; Hurt, E.C. Temperature-sensitive mutations demonstrate roles for yeast fibrillarin in pre-rRNA processing, pre-rRNA methylation, and ribosome assembly. Cell 1993, 72, 443–457. [Google Scholar] [CrossRef]
- Ojha, S.; Malla, S.; Lyons, S.M. snoRNPs: Functions in Ribosome Biogenesis. Biomolecules 2020, 10, 783. [Google Scholar] [CrossRef]
- Griffiths, C.J.; Cooper, B.; Gibson, G.J. A video system for investigating breathing disorders during sleep. Thorax 1991, 46, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhao, X.; Yu, Y. Pseudouridylation (Ψ) of U2 snRNA in S.cerevisiae is catalyzed by an RNA-independent mechanism. EMBO J. 2003, 22, 1889–1897. [Google Scholar] [CrossRef] [Green Version]
- Behm-Ansmant, I.; Urban, A.; Ma, X.; Yu, Y.T.; Motorin, Y.; Branlant, C. The Saccharomyces cerevisiae U2 snRNA: Pseudouridine-synthase Pus7p is a novel multisite-multisubstrate RNA:Psi-synthase also acting on tRNAs. RNA 2003, 9, 1371–1382. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; Leon-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouri-dylation of ncRNA and mRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Carlile, T.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Eddy, S.R. A computational screen for methylation guide snoRNAs in yeast. Science 1999, 283, 1168–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.-H.; Henras, A.; Lu, Y.-J.; Zhou, H.; Zhou, W.-X.; Zhu, Y.-Q.; Zhao, J.; Henry, Y.; Caizergues-Ferrer, M.; Bachellerie, J.-P. Seven Novel Methylation Guide Small Nucleolar RNAs Are Processed from a Common Polycistronic Transcript by Rat1p and RNase III in Yeast. Mol. Cell. Biol. 1999, 19, 1144–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter-Roshek, J.L.; Petrov, A.N.; Dinman, J.D. Optimization of Ribosome Structure and Function by rRNA Base Modification. PLoS ONE 2007, 2, e174. [Google Scholar] [CrossRef] [PubMed]
- Zebarjadian, Y.; King, T.; Fournier, M.J.; Clarke, L.; Carbon, J. Point Mutations in Yeast CBF5 Can Abolish In Vivo Pseudouridylation of rRNA. Mol. Cell. Biol. 1999, 19, 7461–7472. [Google Scholar] [CrossRef] [Green Version]
- Jack, K.; Bellodi, C.; Landry, D.M.; Niederer, R.; Meskauskas, A.; Musalgaonkar, S.; Kopmar, N.; Krasnykh, O.; Dean, A.M.; Thompson, S.; et al. rRNA Pseudouridylation Defects Affect Ribosomal Ligand Binding and Translational Fidelity from Yeast to Human Cells. Mol. Cell 2011, 44, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Esguerra, J.; Warringer, J.; Blomberg, A. Functional importance of individual rRNA 2′-O-ribose methylations revealed by high-resolution phenotyping. RNA 2008, 14, 649–656. [Google Scholar] [CrossRef] [Green Version]
- King, T.H.; Liu, B.; McCully, R.R.; Fournier, M.J. Ribosome Structure and Activity Are Altered in Cells Lacking snoRNPs that Form Pseudouridines in the Peptidyl Transferase Center. Mol. Cell 2003, 11, 425–435. [Google Scholar] [CrossRef]
- Hirabayashi, N.; Sato, N.S.; Suzuki, T. Conserved Loop Sequence of Helix 69 in Escherichia coli 23 S rRNA Is Involved in A-site tRNA Binding and Translational Fidelity. J. Biol. Chem. 2006, 281, 17203–17211. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.-H.; Liu, Q.; Fournier, M.J. rRNA Modifications in an Intersubunit Bridge of the Ribosome Strongly Affect Both Ribosome Biogenesis and Activity. Mol. Cell 2007, 28, 965–977. [Google Scholar] [CrossRef]
- Baudin-Baillieu, A.; Fabret, C.; Liang, X.-H.; Piekna-Przybylska, D.; Fournier, M.J.; Rousset, J.-P. Nucleotide modifications in three functionally important regions of the Saccharomyces cerevisiae ribosome affect translation accuracy. Nucleic Acids Res. 2009, 37, 7665–7677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piekna-Przybylska, D.; Przybylski, P.; Baudin-Baillieu, A.; Rousset, J.-P.; Fournier, M.J. Ribosome Performance Is Enhanced by a Rich Cluster of Pseudouridines in the A-site Finger Region of the Large Subunit. J. Biol. Chem. 2008, 283, 26026–26036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.H.; Liu, Q.; Fournier, M.J. Loss of rRNA modifications in the decoding center of the ribosome impairs translation and strongly delays pre-rRNA processing. RNA 2009, 15, 1716–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigova, A.; Duggimpudi, S.; Pollex, T.; Schaefer, M.; Kos, M. A cluster of methylations in the domain IV of 25S rRNA is required for ribosome stability. RNA 2014, 20, 1632–1644. [Google Scholar] [CrossRef] [Green Version]
- Schosserer, M.; Minois, N.; Angerer, T.B.; Amring, M.; Dellago, H.; Harreither, E.; Calle-Perez, A.; Pircher, A.; Gerstl, M.P.; Pfeifenberger, S.; et al. Methylation of ribosomal RNA by NSUN5 is a conserved mechanism modulating organismal lifespan. Nat. Commun. 2015, 6, 6158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janin, M.; Ortiz-Barahona, V.; de Moura, M.C.; Martinez-Cardus, A.; Llinas-Arias, P.; Soler, M.; Nachmani, D.; Pelletier, J.; Schumann, U.; Calleja-Cervantes, M.E.; et al. Epigenetic loss of RNA-methyltransferase NSUN5 in glioma targets ribosomes to drive a stress adaptive translational program. Acta Neuropathol. 2019, 138, 1053–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peifer, C.; Sharma, S.; Watzinger, P.; Lamberth, S.; Kotter, P.; Entian, K.D. Yeast Rrp8p, a novel méthyltransférase responsible for m1A 645 base modification of 25S rRNA. Nucleic Acids Res. 2013, 41, 1151–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Hartmann, J.D.; Watzinger, P.; Klepper, A.; Peifer, C.; Kötter, P.; Lafontaine, D.L.J.; Entian, K.-D. A single N1-methyladenosine on the large ribosomal subunit rRNA impacts locally its structure and the translation of key metabolic enzymes. Sci. Rep. 2018, 8, 11904. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Santos, D.A.; Hussmann, J.A.; Wang, Y.; Sutter, B.M.; Weissman, J.S.; Tu, B.P. Regulation of translation by methylation multiplicity of 18S rRNA. Cell Rep. 2021, 34, 108825. [Google Scholar] [CrossRef] [PubMed]
- Lafontaine, D.; Delcour, J.; Glasser, A.L.; Desgres, J.; Vandenhaute, J. The DIM1 gene responsible for the conserved m6(2)Am6(2)A dimethylation in the 3′-terminal loop of 18 S rRNA is essential in yeast. J. Mol. Biol. 1994, 241, 492–497. [Google Scholar] [CrossRef]
- Ingolia, N.T.; Hussmann, J.A.; Weissman, J.S. Ribosome Profiling: Global Views of Translation. Cold Spring Harb. Perspect. Biol. 2019, 11, a032698. [Google Scholar] [CrossRef]
- Baudin-Baillieu, A.; Hatin, I.; Legendre, R.; Namy, O. Translation Analysis at the Genome Scale by Ribosome Profiling. Methods Mol. Biol. 2016, 1361, 105–124. [Google Scholar]
- Bywater, M.J.; Poortinga, G.; Sanij, E.; Hein, N.; Peck, A.; Cullinane, C.; Wall, M.; Cluse, L.; Drygin, D.; Anderes, K.; et al. Inhibition of RNA Polymerase I as a Therapeutic Strategy to Promote Cancer-Specific Activation of p53. Cancer Cell 2012, 22, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.-M.; Thompson, I.; Yuan, Z.-M. Elevated snoRNA biogenesis is essential in breast cancer. Oncogene 2013, 33, 1348–1358. [Google Scholar] [CrossRef]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. p53 Acts as a Safeguard of Translational Control by Regulating Fibrillarin and rRNA Methylation in Cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erales, J.; Marchand, V.; Panthu, B.; Gillot, S.; Belin, S.; Ghayad, S.E.; Garcia, M.; Laforets, F.; Marcel, V.; Baudin-Baillieu, A.; et al. Evidence for rRNA 2′-O-methylation plasticity: Control of intrinsic translational capabilities of human ribosomes. Proc. Natl. Acad. Sci. USA 2017, 114, 12934–12939. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Modification | Position | Enzyme/snoRNA | Modification Rate | Conservation | Functional Domain |
|---|---|---|---|---|---|
| Ψ | 5S-50 | Pus7 | >85% | H | |
| Ψ | 5,8S-73 | snR43 | <85% | ||
| Am | 18S-28 | snR74 | >85% | H | |
| Am | 18S-100 | snR51 | <85% | H | |
| Ψ | 18S-106 | snR44 | >85% | H | |
| Ψ | 18S-120 | snR49 | >85% | H | |
| Ψ | 18S-211 | snR49 | <85% | ||
| Ψ | 18S-302 | snR49 | >85% | ||
| Cm | 18S-414 | U14 | >85% | H | |
| Am | 18S-420 | snR52 | >85% | H | |
| Am | 18S-436 | snR87 | >85% d | H | |
| Ψ | 18S-466 | snR189 | <85% | ||
| Am | 18S-541 | snR41 | >85% | H | |
| Gm | 18S-562 | snR40 | >85% d | ||
| Um | 18S-578 | snR77 | >85% | H | DC-A site |
| Am | 18S-619 | snR47 | >85% | H | |
| Ψ | 18S-632 | snR161 | <85% | H | |
| Ψ | 18S-759 | snR80 | >85% | H | |
| Ψ | 18S-766 | snR161 | >85% | H | |
| Am | 18S-796 | snR53 | >85% | ||
| Am | 18S-974 | snR54 | >85% | H | |
| Ψ | 18S-999 | snR31 | <85% | H | IS bridge DC-E site |
| Cm | 18S-1007 | snR79 | >85% | IS bridge DC-E site | |
| Gm | 18S-1126 | snR41 | >85% | IS bridge | |
| Ψ | 18S-1181 | snR85 | >85% | H | IS bridge |
| Ψ | 18S-1187 | snR36 | >85% | H | DC-A site |
| m1acp3Ψ | 18S-1191 | snR35,Emg1, Tsr3 | >85% | H | DC-P site |
| Um | 18S-1269 | snR55 | >85% | H | DC-A site |
| Gm | 18S-1271 | snR40 | >85% | H | DC-A site |
| ac4C | 18S-1280 | snR4, Kre33 | >85% | H | DC-A site |
| Ψ | 18S-1290 | snR83 | >85% | H | |
| Ψ | 18S-1415 | snR83 | <85% | ||
| Gm | 18S-1428 | snR56 | >85% | H | DC-A site |
| Gm | 18S-1572 | snR57 | >85% | E-site | |
| m7G | 18S-1575 | Bud23 | >85% d | H | |
| Cm | 18S-1639 | snR70 | >85% d | Ec, H | DC-P site |
| ac4C | 18S-1773 | snR45, Kre33 | >85% | H | IS bridge |
| m26A | 18S-1781 | Dim1 | >85% | Ec, H | IS bridge |
| m26A | 18S-1782 | Dim1 | >85% | Ec, H | IS bridge |
| m1A | 25S-645 | Bmt1 (Rrp8) | >85% | H | PTC |
| Am | 25S-649 | U18 | >85% | H | Peptide exit tunnel |
| Cm | 25S-650 | U18 | >85% | Peptide exit tunnel | |
| Cm | 25S-663 | snR58 | <85% d | H | Peptide exit tunnel |
| Ψ | 25S-776 | snR80 | >85% | ||
| Gm | 25S-805 | snR39b | >85% | H | Peptide exit tunnel |
| Am | 25S-807 | snR39, snR59 | >85% | H | |
| Am | 25S-817 | snR60 | >85% | H | |
| Gm | 25S-867 | snR50 | >85% d | ||
| Am | 25S-876 | snR72 | >85% d | ||
| Um | 25S-898 | snR40 | >85% d | ||
| Gm | 25S-908 | snR60 | >85% | H | |
| Ψ | 25S-960 | snR8 | >85% | H | A site finger helix 37 |
| Ψ | 25S-966 | snR43 | >85% | H | A site finger helix 37 |
| Ψ | 25S-986 | snR8 | >85% | A site finger helix 38 | |
| Ψ | 25S-990 | snR49 | >85% | A site finger helix 38 | |
| Ψ | 25S-1004 | snR5 | <85% | H | A site finger helix 38 |
| Ψ | 25S-1042 | snR33 | >85% | H | A site finger helix 38 |
| Ψ | 25S-1052 | snR81 | >85% | H | A site finger helix 38 |
| Ψ | 25S-1056 | snR44 | >85% | A site finger helix 38 | |
| Ψ | 25S-1110 | snR82 | >85% | ||
| Ψ | 25S-1124 | snR5 | >85% | H | A site finger helix 39 |
| Am | 25S-1133 | snR61 | >85% | H | A site finger helix 39 |
| Cm | 25S-1437 | U24 | >85% | H | Peptide exit tunnel |
| Am | 25S-1449 | U24 | >85% d | H | |
| Gm | 25S-1450 | U24 | >85% | H | |
| Um | 25S-1888 | snR62 | >85% | H | |
| Ψ | 25S-2129 | snR5 | >85% | ||
| Ψ | 25S-2133 | snR3 | >85% | H | |
| m1A | 25S-2142 | Bmt2 | >85% | Subunit surface | |
| Ψ | 25S-2191 | snR32 | >85% | H | |
| Cm | 25S-2197 | snR76 | >85% d | H | E-site |
| Am | 25S-2220 | snR47 | >85% | H | E-site |
| Am | 25S-2256 | snR63 | >85% | H | IS bridge helix 69 A site tRNA |
| Ψ | 25S-2258 | snR191 | >85% | Ec, H | IS bridge helix 69 A site tRNA |
| Ψ | 25S-2260 | snR191 | >85% | Ec, H | IS bridge helix 69 |
| Ψ | 25S-2264 | snR3 | >85% | H | IS bridge helix 69 |
| Ψ | 25S-2266 | snR84 | >85% | H | IS bridge helix 69 P-site tRNA |
| m5C | 25S-2278 | Bmt3 (Rcm1) | >85% | H | IS bridge |
| Am | 25S-2280 | snR13 | >85% | IS bridge | |
| Am | 25S-2281 | snR13 | >85% | H | IS bridge |
| Gm | 25S-2288 | snR75 | >85% | H | IS bridge |
| Ψ | 25S-2314 | snR86 | >85% | H | |
| Cm | 25S-2337 | snR64 | >85% | H | |
| Ψ | 25S-2340 | snR9 | >85% | H | |
| Um, Ψ, Ψm | 25S-2347 | snR65, snR9 | H | ||
| Ψ | 25S-2349 | snR82 | >85% | H | |
| Ψ | 25S-2351 | snR82 | >85% | ||
| Ψ | 25S-2416 | snR11 | >85% | H | |
| Um | 25S-2417 | snR66 | >85% | ||
| Um | 25S-2421 | snR78 | >85% | H | E-site |
| Gm | 25S-2619 | snR67 | >85% | Ec, H | P-site |
| m3U | 25S-2634 | Bmt5 | >85% | PTC | |
| Am | 25S-2640 | snR68 | >85% | ||
| Um | 25S-2724 | snR67 | >85% | ||
| Um | 25S-2729 | snR51 | >85% d | H | |
| Ψ | 25S-2735 | snR189 | >85% | H | |
| Gm | 25S-2791 | snR48 | >85% | E-site | |
| Gm | 25S-2793 | snR48 | >85% | H | E-site |
| Gm | 25S-2815 | snR38 | >85% | H | PTC |
| Ψ | 25S-2826 | snR34 | >85% | H | PTC |
| m3U | 25S-2843 | Bmt6 | >85% | H | |
| Ψ | 25S-2865 | snR46 | >85% | H | PTC |
| m5C | 25S-2870 | Bmt4 (Nop2) | >85% | Ec, H | PTC |
| Ψ | 25S-2880 | snR34 | >85% | H | PTC |
| Um | 25S-2921 | snR52, Spb1 | >85% | Ec, H | PTC |
| Gm | 25S-2922 | Spb1 | >85% | H | PTC |
| Ψ | 25S-2923 | snR10 | >85% | H | PTC |
| Ψ | 25S-2944 | snR37 | <85% | H | PTC |
| Am | 25S-2946 | snR71 | >85% | H | PTC |
| Cm | 25S-2948 | snR69 | >85% d | PTC | |
| Cm | 25S-2959 | snR73 | >85% | H | PTC |
| Ψ | 25S-2975 | snR42 | <85% | H | PTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baudin-Baillieu, A.; Namy, O. Saccharomyces cerevisiae, a Powerful Model for Studying rRNA Modifications and Their Effects on Translation Fidelity. Int. J. Mol. Sci. 2021, 22, 7419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147419
Baudin-Baillieu A, Namy O. Saccharomyces cerevisiae, a Powerful Model for Studying rRNA Modifications and Their Effects on Translation Fidelity. International Journal of Molecular Sciences. 2021; 22(14):7419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147419
Chicago/Turabian StyleBaudin-Baillieu, Agnès, and Olivier Namy. 2021. "Saccharomyces cerevisiae, a Powerful Model for Studying rRNA Modifications and Their Effects on Translation Fidelity" International Journal of Molecular Sciences 22, no. 14: 7419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147419