
Regulatory T Cells Exhibit Interleukin-33-Dependent Migratory Behavior during Skin Barrier Disruption

 , and
, and
Abstract
:1. Introduction
2. Results
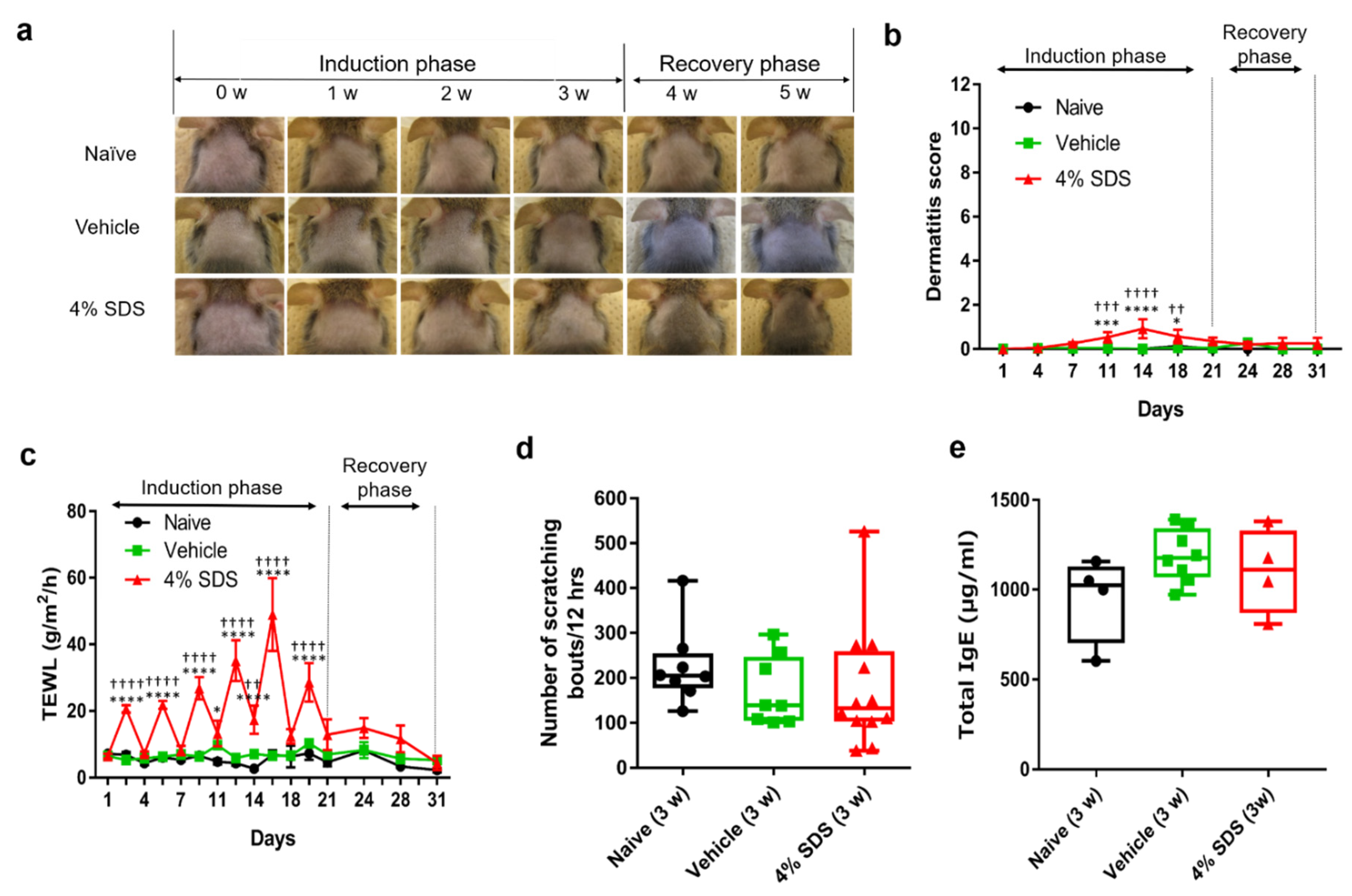
2.1. Establishment of Skin Barrier Disruption Model Mice by Repeated 4% SDS Application
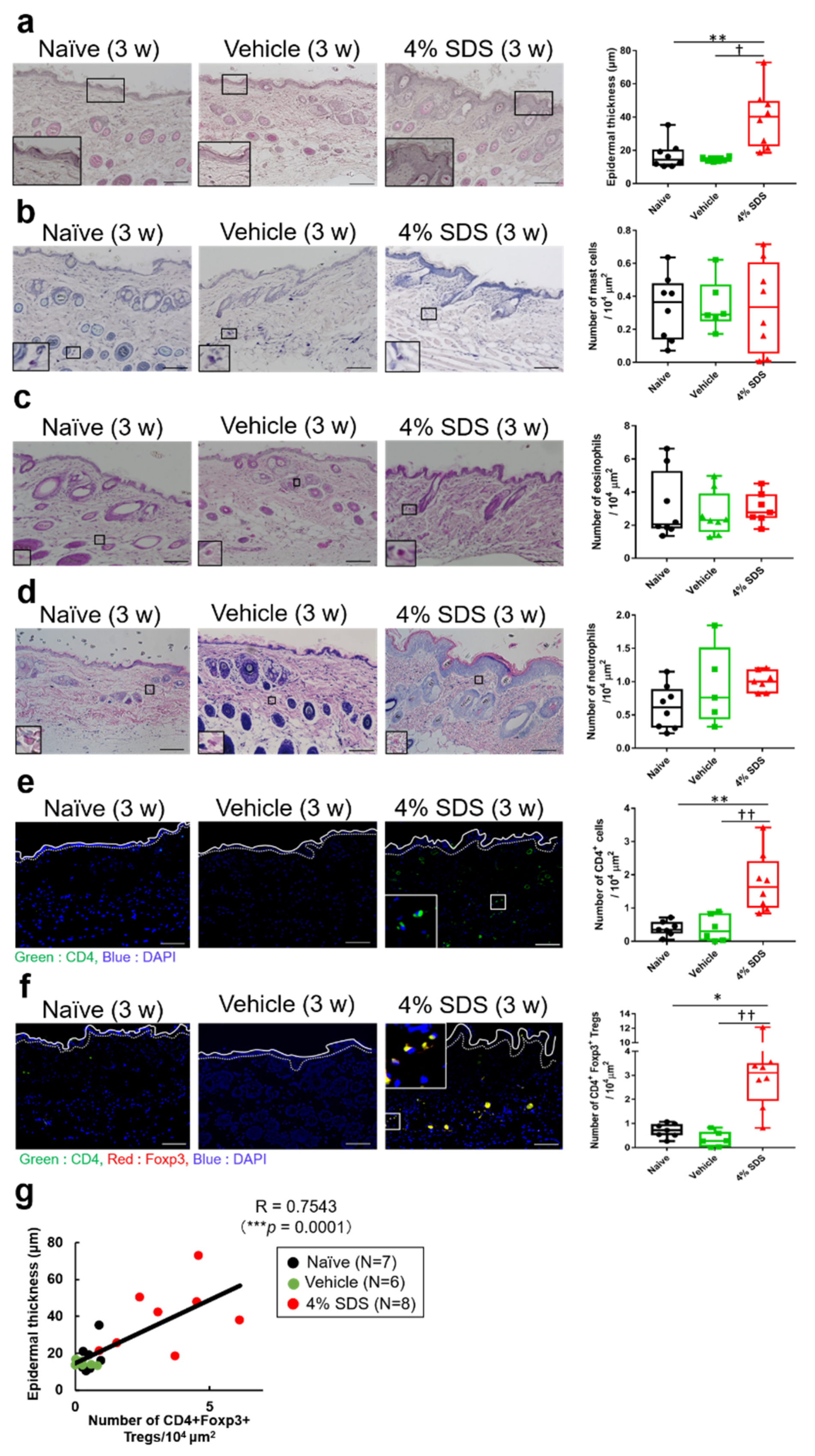
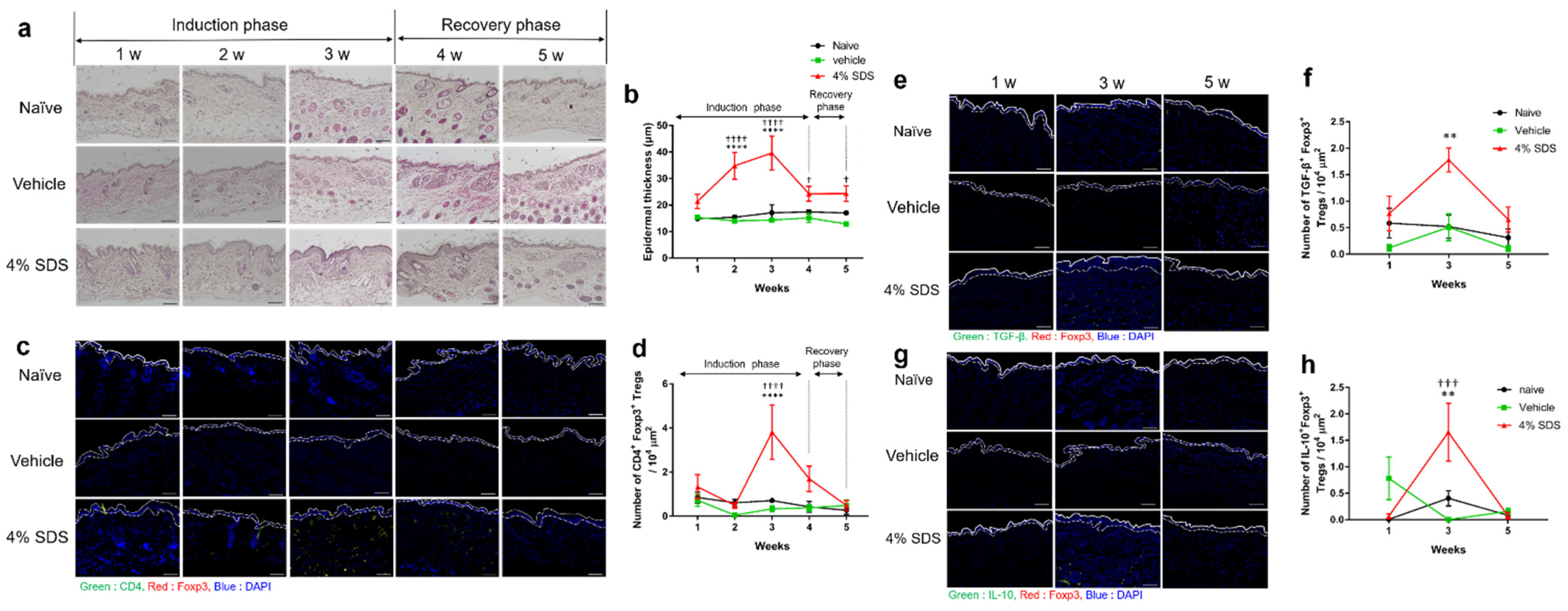
2.2. Effects of Skin Barrier Disruption on Acanthosis and Inflammatory Cells in Lesional Skin
2.3. Tregs Express Regulatory Cytokines in Barrier-Disrupted Skin
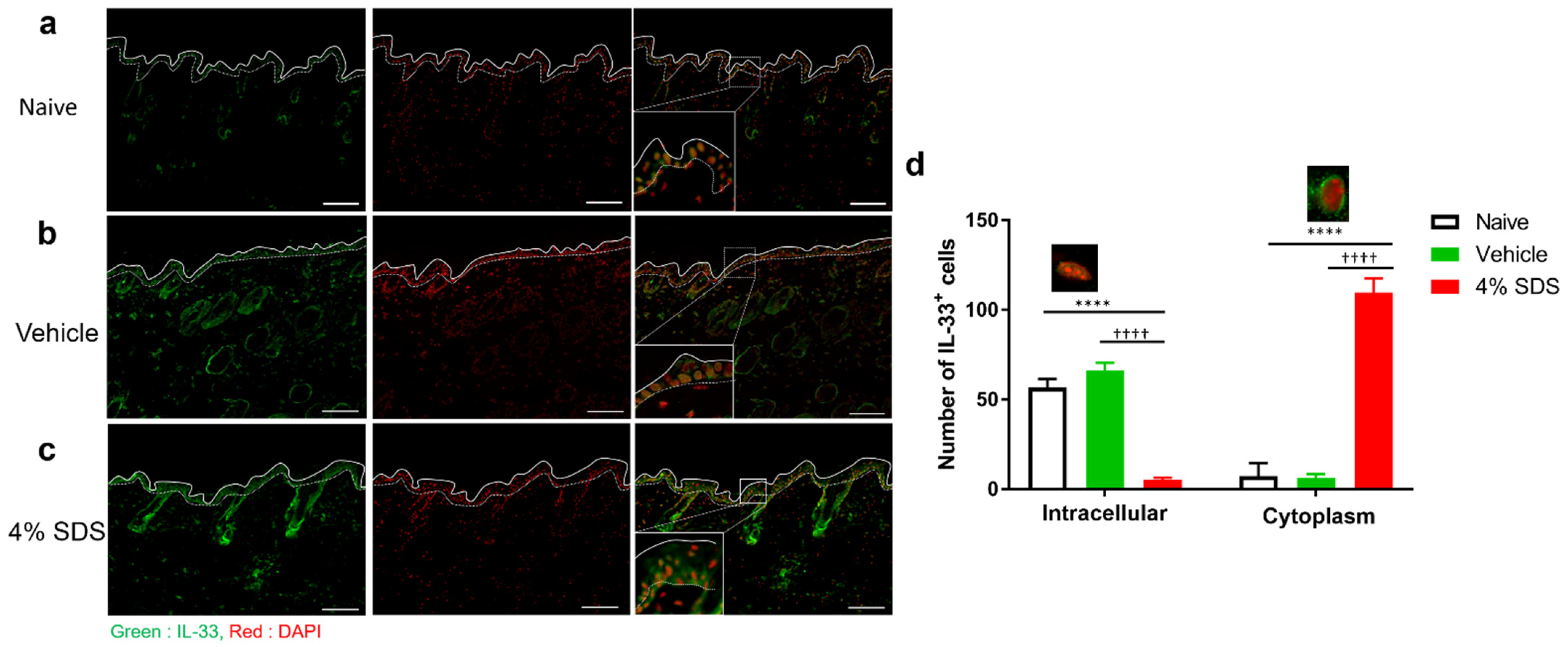
2.4. Localization of IL-33 in Barrier-Disrupted Skin
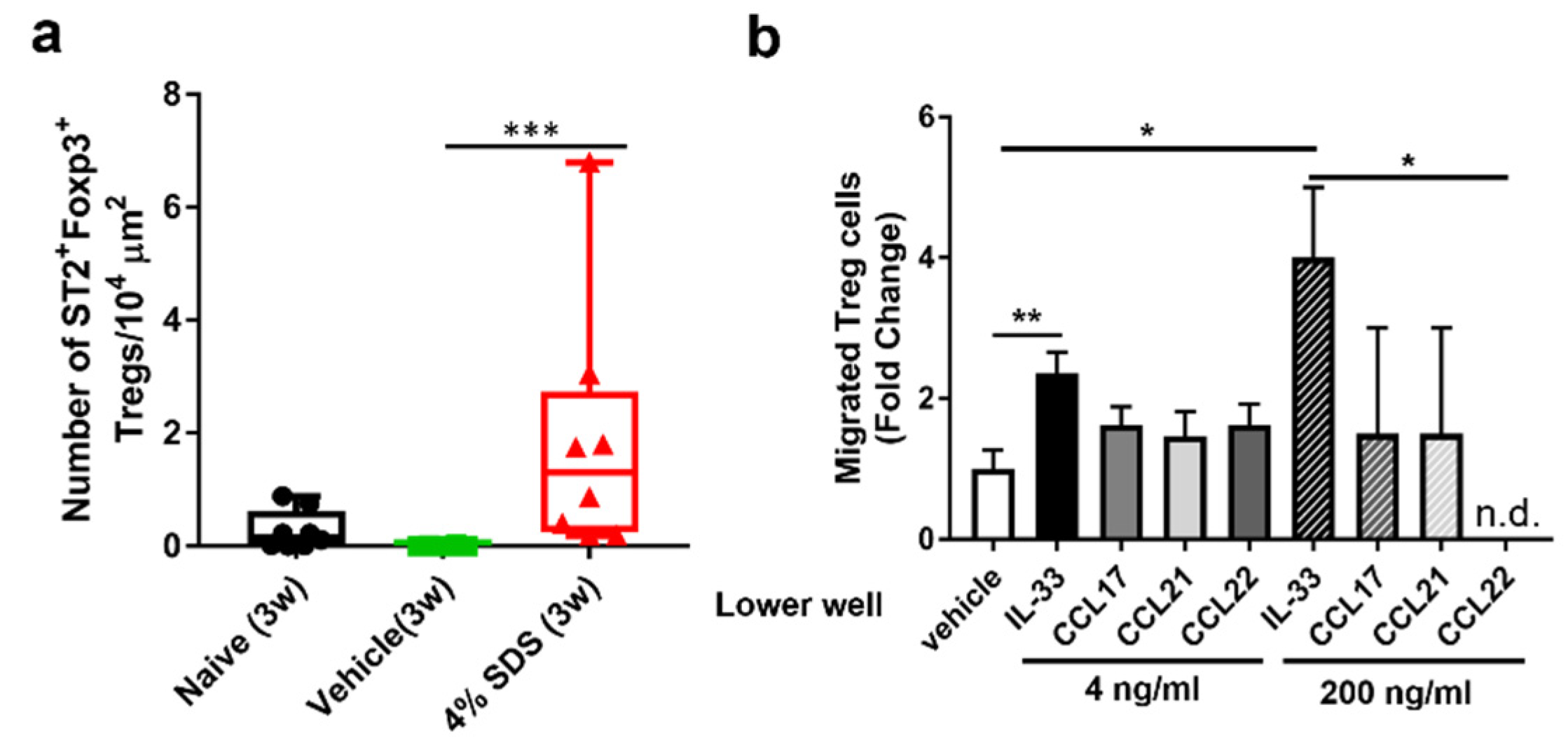
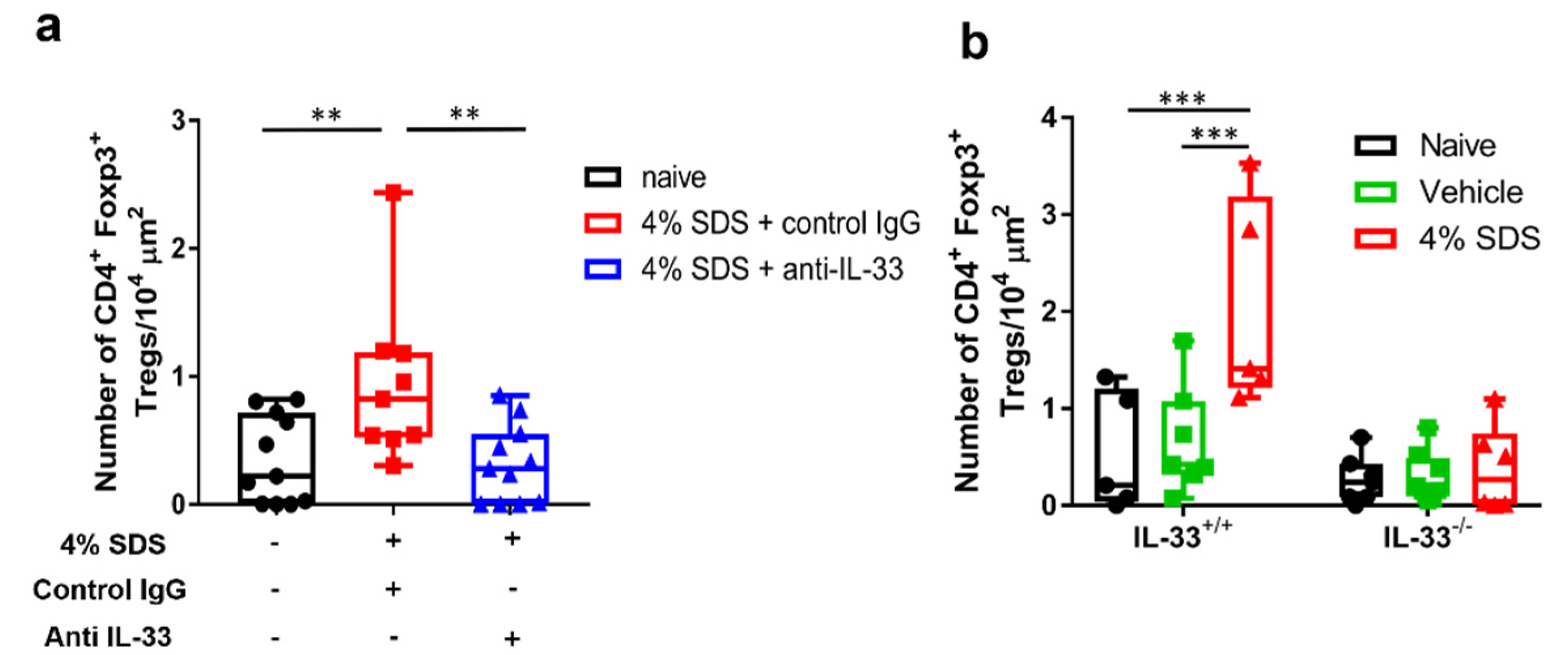
2.5. IL-33-Dependent Migratory Behavior of Tregs
3. Discussion
4. Materials and Methods
4.1. Animals
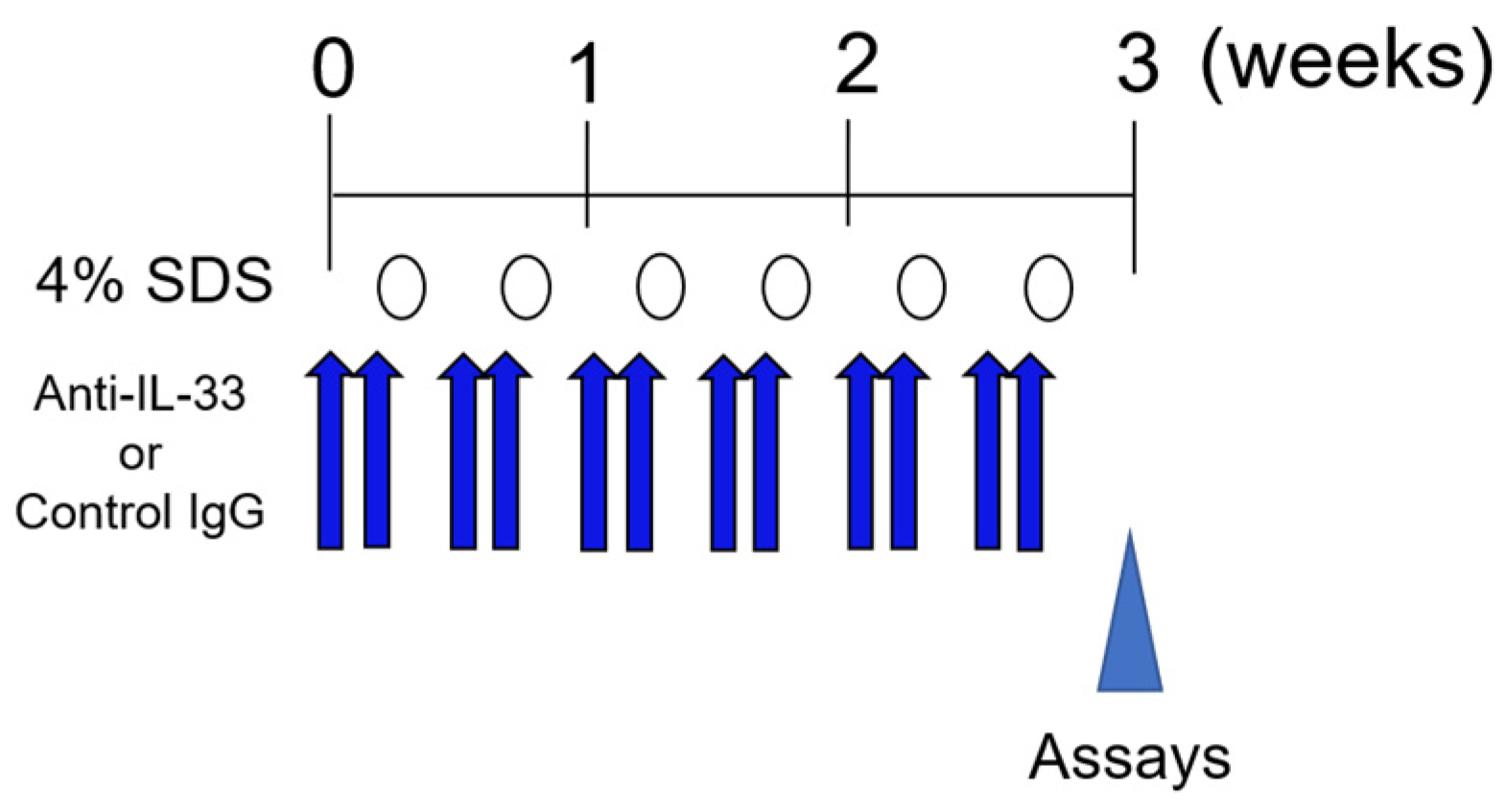
4.2. Skin Barrier-Disrupted Model Mice
4.3. Evaluation of Skin Condition and Dermatitis
4.4. Observation of Scratching Behavior
4.5. Measurement of Total IgE Levels in Sera
4.6. Histological and Immunohistochemical Analyses
4.7. In Situ Hybridization Using the RNAscope® Method
4.8. Quantitative Analyses from Histological Data
4.9. FACS Analysis
4.10. In Vitro Chemotaxis Assay
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rawlings, A.V.; Harding, C.R. Moisturization and Skin Barrier Function. Dermatol. Ther. 2004, 17 (Suppl. S1), 43–48. [Google Scholar] [CrossRef]
- Cork, M.J.; Danby, S.G.; Vasilopoulos, Y.; Hadgraft, J.; Lane, M.E.; Moustafa, M.; Guy, R.H.; Macgowan, A.L.; Tazi-Ahnini, R.; Ward, S.J. Epidermal Barrier Dysfunction in Atopic Dermatitis. J. Investig. Dermatol. 2009, 129, 1892–1908. [Google Scholar] [CrossRef] [PubMed]
- Danby, S.G. Biological Variation in Skin Barrier Function: From A (Atopic Dermatitis) to X (Xerosis). Curr. Probl. Dermatol. 2016, 49, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Morelli, P.; Gaspari, M.; Gabriele, C.; Dastoli, S.; Bennardo, L.; Pavel, A.B.; Patruno, C.; Del Duca, E.; Nistico, S.P. Proteomic Analysis from Skin Swabs Reveals a New Set of Proteins Identifying Skin Impairment in Atopic Dermatitis. Exp. Dermatol. 2021, 30, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S. Naturally Arising Foxp3-Expressing CD25+CD4+ Regulatory T Cells in Immunological Tolerance to Self and Non-self. Nat. Immunol. 2005, 6, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic Self-Tolerance Maintained by Activated T Cells Expressing IL-2 Receptor Alpha-Chains (CD25). Breakdown of a Single Mechanism of Self-Tolerance Causes Various Autoimmune Diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar]
- Khattri, R.; Cox, T.; Yasayko, S.A.; Ramsdell, F. An Essential Role for Scurfin in CD4+CD25+ T Regulatory Cells. Nat. Immunol. 2003, 4, 337–342. [Google Scholar] [CrossRef]
- Shimizu, J.; Yamazaki, S.; Sakaguchi, S. Induction of Tumor Immunity by Removing CD25(+)CD4(+) T Cells: A Common Basis between Tumor Immunity and Autoimmunity. J. Immunol. 1999, 163, 5211–5218. [Google Scholar]
- Rubtsov, Y.P.; Rasmussen, J.P.; Chi, E.Y.; Fontenot, J.; Castelli, L.; Ye, X.; Treuting, P.; Siewe, L.; Roers, A.; Henderson, W.R., Jr.; et al. Regulatory T Cell-Derived Interleukin-10 Limits Inflammation at Environmental Interfaces. Immunity 2008, 28, 546–558. [Google Scholar] [CrossRef]
- Worthington, J.J.; Kelly, A.; Smedley, C.; Bauche, D.; Campbell, S.; Marie, J.C.; Travis, M.A. Integrin Alphavbeta8-Mediated TGF-Beta Activation by Effector Regulatory T Cells Is Essential for Suppression of T-Cell-Mediated Inflammation. Immunity 2015, 42, 903–915. [Google Scholar] [CrossRef] [Green Version]
- Wildin, R.S.; Ramsdell, F.; Peake, J.; Faravelli, F.; Casanova, J.L.; Buist, N.; Levy-Lahad, E.; Mazzella, M.; Goulet, O.; Perroni, L.; et al. X-linked Neonatal Diabetes Mellitus, Enteropathy and Endocrinopathy Syndrome Is the Human Equivalent of Mouse Scurfy. Nat. Genet. 2001, 27, 18–20. [Google Scholar] [CrossRef]
- Brunkow, M.E.; Jeffery, E.W.; Hjerrild, K.A.; Paeper, B.; Clark, L.B.; Yasayko, S.A.; Wilkinson, J.E.; Galas, D.; Ziegler, S.F.; Ramsdell, F. Disruption of a New Forkhead/Winged-Helix Protein, Scurfin, Results in the Fatal Lymphoproliferative Disorder of the Scurfy Mouse. Nat. Genet. 2001, 27, 68–73. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 Programs the Development and Function of CD4+CD25+ Regulatory T Cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Gambineri, E.; Perroni, L.; Passerini, L.; Bianchi, L.; Doglioni, C.; Meschi, F.; Bonfanti, R.; Sznajer, Y.; Tommasini, A.; Lawitschka, A.; et al. Clinical and Molecular Profile of a New Series of Patients with Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome: Inconsistent Correlation between Forkhead Box Protein 3 Expression and Disease Severity. J Allergy Clin. Immunol. 2008, 122, 1105–1112.e1. [Google Scholar] [CrossRef]
- Wildin, R.S.; Smyk-Pearson, S.; Filipovich, A.H. Clinical and Molecular Features of the Immunodysregulation, Polyendocrinopathy, Enteropathy, X Linked (IPEX) Syndrome. J. Med. Genet 2002, 39, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Negi, O.; Tominaga, M.; Tengara, S.; Kamo, A.; Taneda, K.; Suga, Y.; Ogawa, H.; Takamori, K. Topically Applied Semaphorin 3A Ointment Inhibits Scratching Behavior and Improves Skin Inflammation in NC/Nga Mice with Atopic Dermatitis. J. Dermatol. Sci. 2012, 66, 37–43. [Google Scholar] [CrossRef]
- Bruhs, A.; Proksch, E.; Schwarz, T.; Schwarz, A. Disruption of the Epidermal Barrier Induces Regulatory T Cells via IL-33 in Mice. J. Investig. Dermatol. 2018, 138, 570–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Sakaguchi, S. Targeting Treg Cells in Cancer Immunotherapy. Eur. J. Immunol. 2019, 49, 1140–1146. [Google Scholar] [CrossRef] [Green Version]
- Panduro, M.; Benoist, C.; Mathis, D. Tissue Tregs. Annu. Rev. Immunol. 2016, 34, 609–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhagen, J.; Akdis, M.; Traidl-Hoffmann, C.; Schmid-Grendelmeier, P.; Hijnen, D.; Knol, E.F.; Behrendt, H.; Blaser, K.; Akdis, C.A. Absence of T-Regulatory Cell Expression and Function in Atopic Dermatitis Skin. J. Allergy Clin. Immunol. 2006, 117, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Gri, G.; Piconese, S.; Frossi, B.; Manfroi, V.; Merluzzi, S.; Tripodo, C.; Viola, A.; Odom, S.; Rivera, J.; Colombo, M.P.; et al. CD4+CD25+ regulatory T Cells Suppress Mast Cell Degranulation and Allergic Responses through OX40-OX40L Interaction. Immunity 2008, 29, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez Rodriguez, R.; Pauli, M.L.; Neuhaus, I.M.; Yu, S.S.; Arron, S.T.; Harris, H.W.; Yang, S.H.; Anthony, B.A.; Sverdrup, F.M.; Krow-Lucal, E.; et al. Memory Regulatory T Cells Reside in Human Skin. J. Clin. Investig. 2014, 124, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aron, J.L.; Akbari, O. Regulatory T Cells and Type 2 Innate Lymphoid Cell-Dependent Asthma. Allergy 2017, 72, 1148–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, H.; Arae, K.; Unno, H.; Miyauchi, K.; Toyama, S.; Nambu, A.; Oboki, K.; Ohno, T.; Motomura, K.; Matsuda, A.; et al. An Interleukin-33-Mast Cell-Interleukin-2 Axis Suppresses Papain-Induced Allergic Inflammation by Promoting Regulatory T Cell Numbers. Immunity 2015, 43, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, E.; Massague, J. Transforming Growth Factor-beta Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef] [PubMed]
- Osabe, M.; Tajika, T.; Tohkin, M. Allopurinol Suppresses Expression of the Regulatory T-cell Migration Factors TARC/CCL17 and MDC/CCL22 in HaCaT keratinocytes via restriction of nuclear factor-kappaB activation. J. Appl. Toxicol. 2018, 38, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Tembhre, M.K.; Parihar, A.S.; Sharma, V.K.; Sharma, A.; Chattopadhyay, P.; Gupta, S. Alteration in Regulatory T Cells and Programmed Cell Death 1-Expressing Regulatory T Cells in Active Generalized Vitiligo and Their Clinical Correlation. Br. J. Dermatol. 2015, 172, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Eby, J.M.; Kang, H.K.; Tully, S.T.; Bindeman, W.E.; Peiffer, D.S.; Chatterjee, S.; Mehrotra, S.; Le Poole, I.C. CCL22 to Activate Treg Migration and Suppress Depigmentation in Vitiligo. J. Investig. Dermatol. 2015, 135, 1574–1580. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Ulzii, D.; Vu, Y.H.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T. Pathogenesis of Atopic Dermatitis: Current Paradigm. Iran. J. Immunol. 2019, 16, 97–107. [Google Scholar] [CrossRef]
- Chauhan, S.K.; Saban, D.R.; Dohlman, T.H.; Dana, R. CCL-21 Conditioned Regulatory T Cells Induce Allotolerance through Enhanced Homing to Lymphoid Tissue. J. Immunol. 2014, 192, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an Interleukin-1-Like Cytokine That Signals Via the IL-1 Receptor-Related Protein ST2 and Induces T helper Type 2-Associated Cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delacher, M.; Imbusch, C.D.; Weichenhan, D.; Breiling, A.; Hotz-Wagenblatt, A.; Trager, U.; Hofer, A.C.; Kagebein, D.; Wang, Q.; Frauhammer, F.; et al. Genome-Wide DNA-Methylation Landscape Defines Specialization of Regulatory T Cells in Tissues. Nat. Immunol. 2017, 18, 1160–1172. [Google Scholar] [CrossRef] [PubMed]
- Lemery, E.; Briancon, S.; Chevalier, Y.; Oddos, T.; Gohier, A.; Boyron, O.; Bolzinger, M.A. Surfactants Have Multi-Fold Effects on Skin Barrier Function. Eur. J. Dermatol. 2015, 25, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.T.; Ma, X.J.; Luo, Y. RNAscope: A Novel in Situ RNA Analysis Platform for Formalin-Fixed, Paraffin-Embedded Tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Probe | Source | Catalog Number |
|---|---|---|---|---|
| CD4 | NM_013488.2 | Mm-Cd4 | Advanced Cell Diagnostics | 406841 |
| Foxp3 | NM_001199347.1 | Mm-Foxp3-C2 | Advanced Cell Diagnostics | 432611-C2 |
| IL-10 | NM_010548.2 | Mm-Il10 | Advanced Cell Diagnostics | 317261 |
| IL1RL1 | NM_001025602.3 | Mm-Il1rl1 | Advanced Cell Diagnostics | 440661 |
| TGF-β | NM_011577.1 | Mm-Tgfb1 | Advanced Cell Diagnostics | 406201 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyama, S.; Moniaga, C.S.; Nakae, S.; Kurosawa, M.; Ogawa, H.; Tominaga, M.; Takamori, K. Regulatory T Cells Exhibit Interleukin-33-Dependent Migratory Behavior during Skin Barrier Disruption. Int. J. Mol. Sci. 2021, 22, 7443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147443
Toyama S, Moniaga CS, Nakae S, Kurosawa M, Ogawa H, Tominaga M, Takamori K. Regulatory T Cells Exhibit Interleukin-33-Dependent Migratory Behavior during Skin Barrier Disruption. International Journal of Molecular Sciences. 2021; 22(14):7443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147443
Chicago/Turabian StyleToyama, Sumika, Catharina Sagita Moniaga, Susumu Nakae, Masaru Kurosawa, Hideoki Ogawa, Mitsutoshi Tominaga, and Kenji Takamori. 2021. "Regulatory T Cells Exhibit Interleukin-33-Dependent Migratory Behavior during Skin Barrier Disruption" International Journal of Molecular Sciences 22, no. 14: 7443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147443