
Mechanical and Thermodynamic Properties of Non-Muscle Contractile Tissues: The Myofibroblast and the Molecular Motor Non-Muscle Myosin Type IIA


Abstract
:1. Introduction
2. The Myofibroblast
3. The Myofibroblast: A Contractile Cell Containing the Non-Muscle Myosin (NMIIA)
4. The Non-Muscle Myosin (NMII)
5. Mechanical and Thermodynamic Techniques for Studying Contractile Tissues
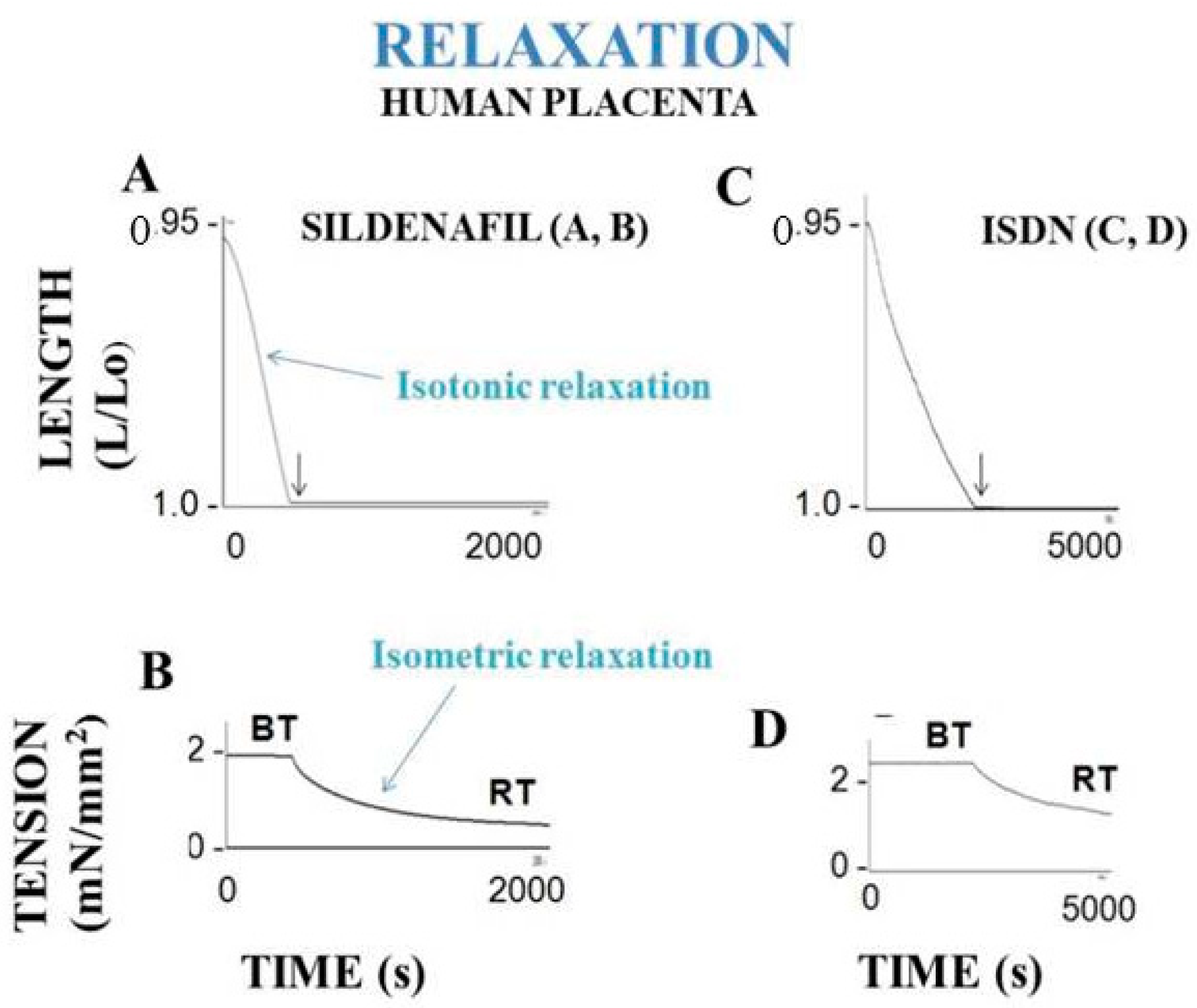
5.1. Numerous Techniques
5.2. Experimental Set-Up
5.3. A. Huxley Formalism
5.3.1. A. Huxley’s Equations
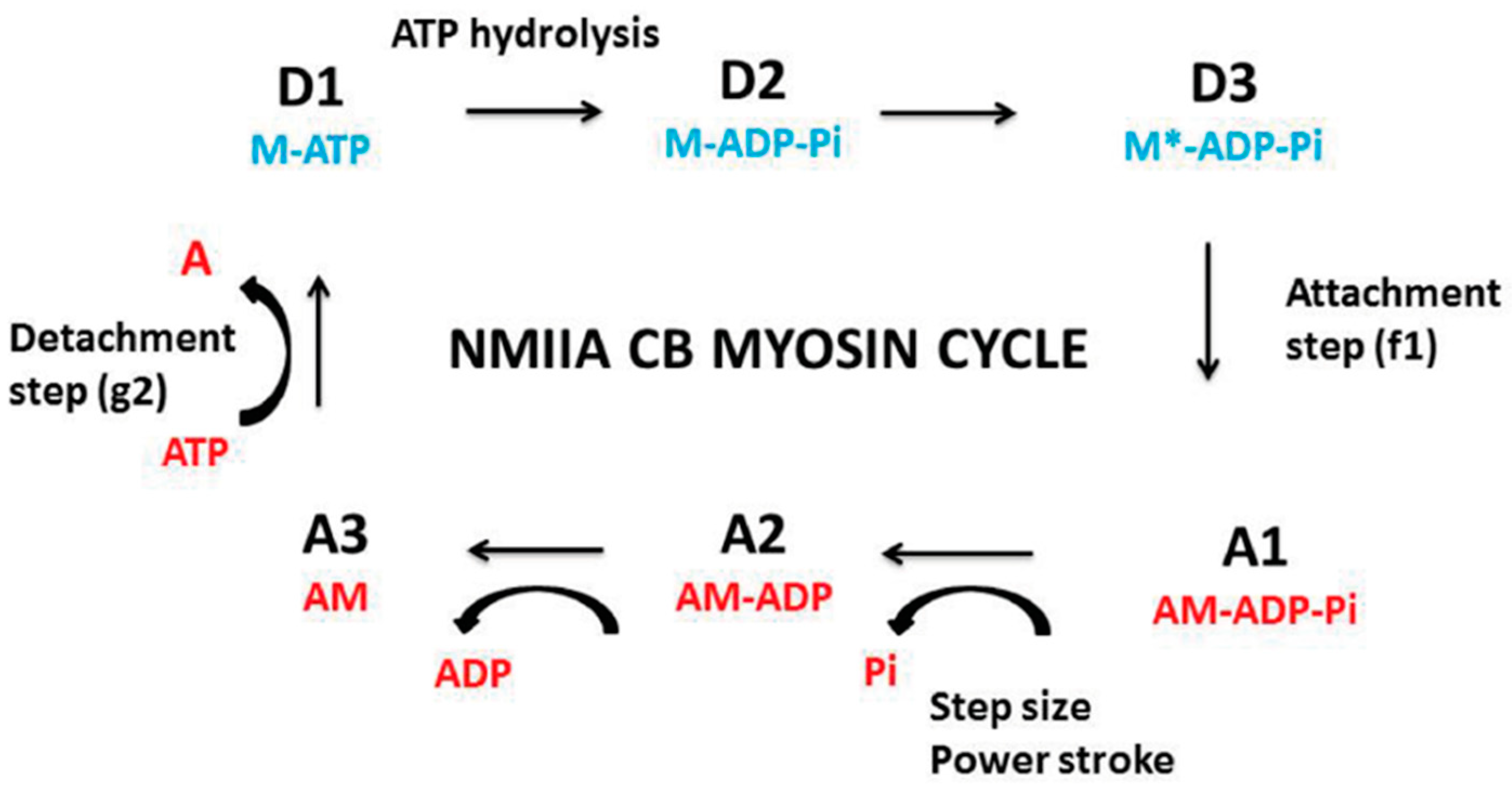
5.3.2. Computation of CB Probabilities of the 6 States of the CB Cycle
5.4. Statistical Mechanics Near Equilibrium
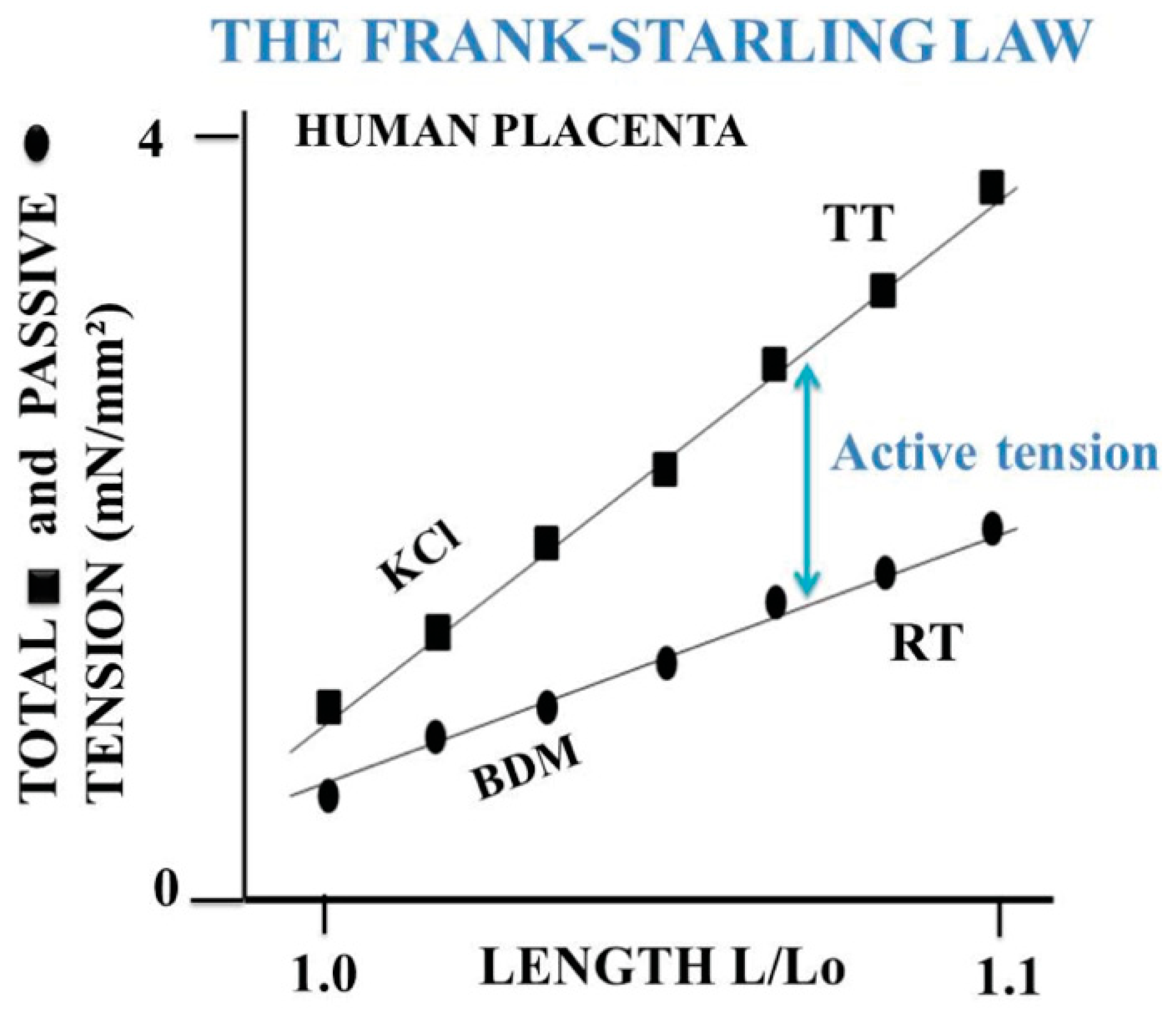
6. Mechanical Properties Shared by Muscles and Non-Muscle Tissues
7. Mechanics of Two Non-Muscle Contractile Tissues: A Physiological Tissue (Human Placenta) and an Engineered Tissue (MSC-Seeded in a Collagen Scaffold)
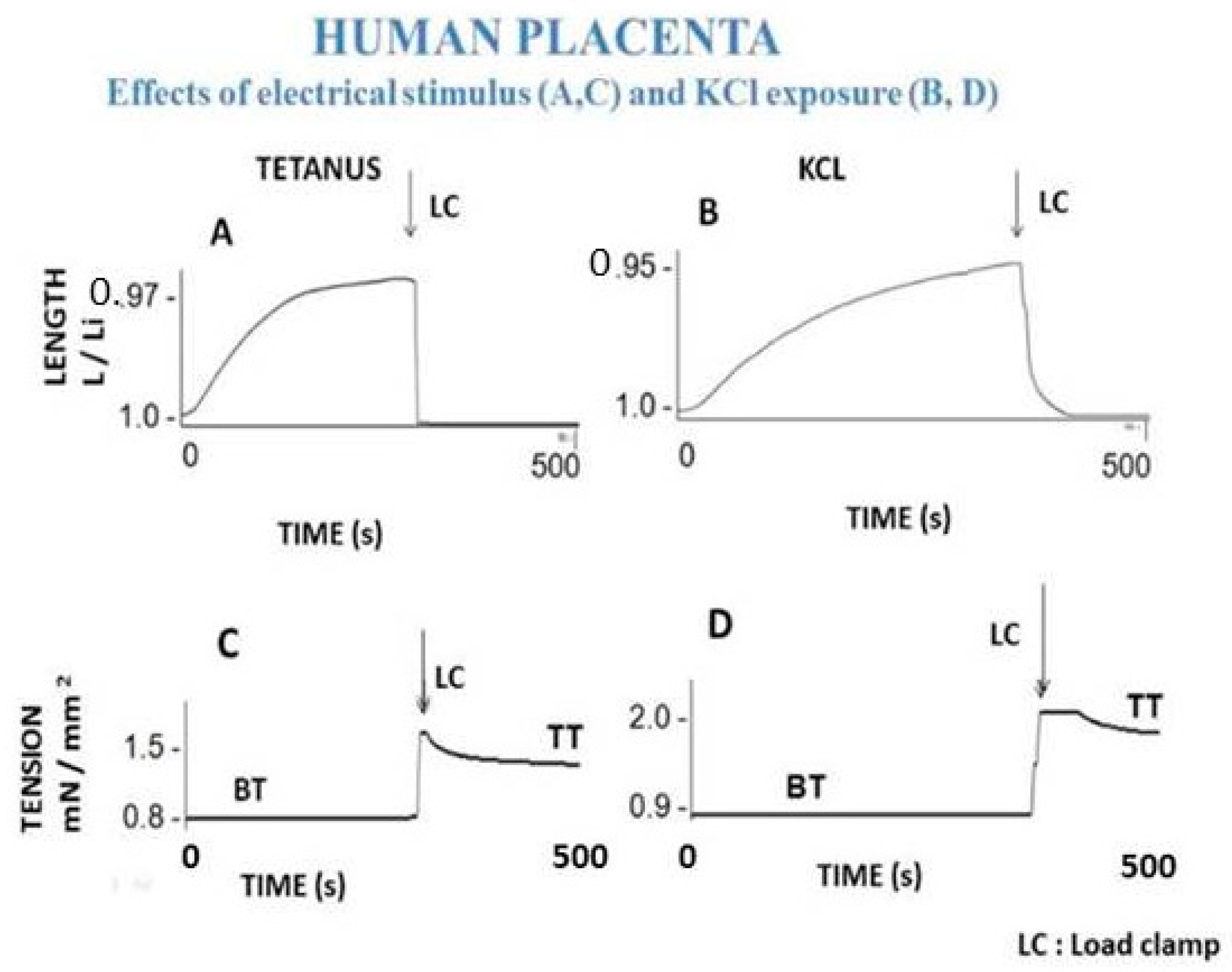
7.1. Human Placenta
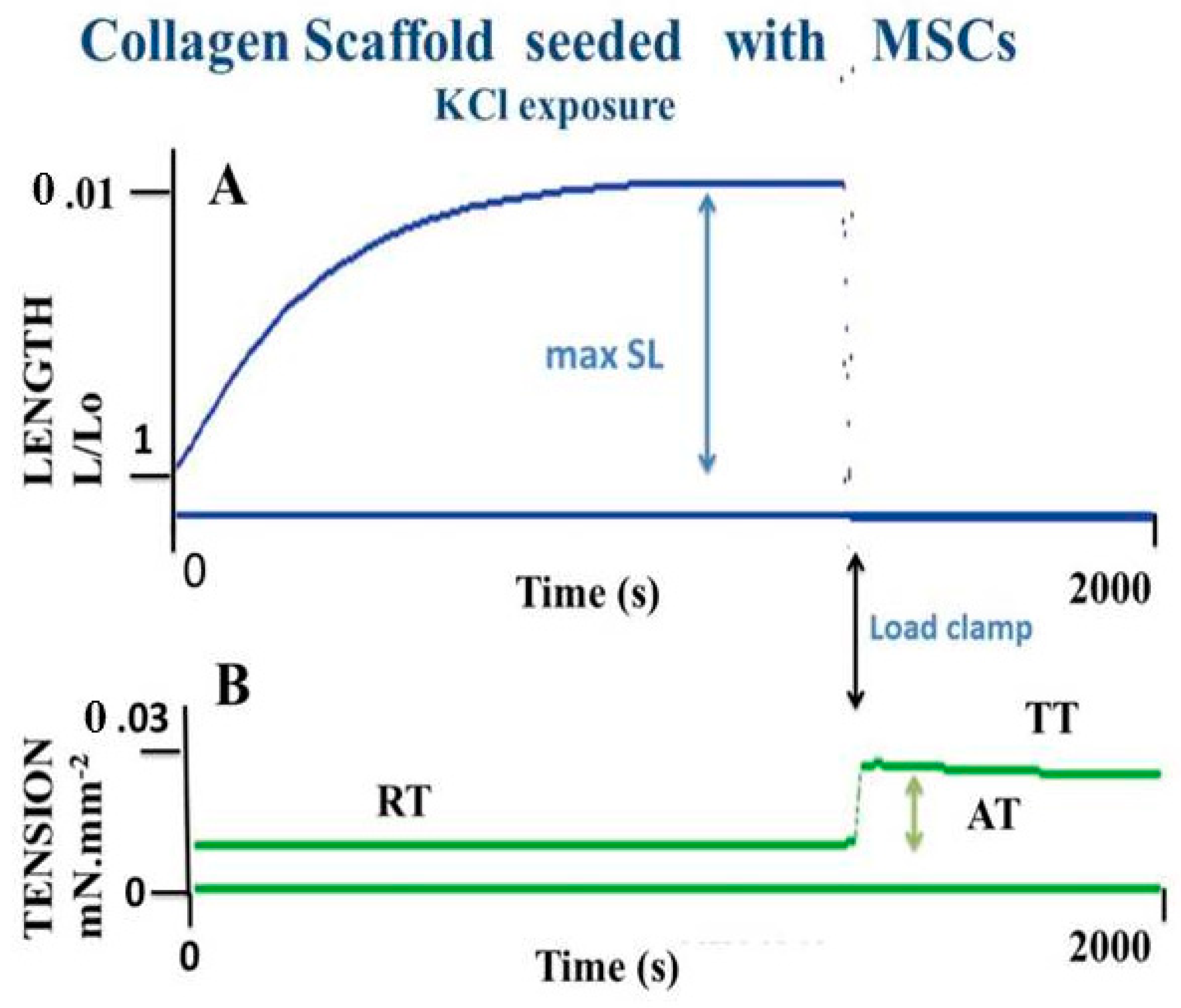
7.2. An Engineered Tissue: MSCs Seeded in Collagen Scaffolds
8. Synthesis
- Their maximum shortening velocity (Vmax) is very slow due to their rate constant of detachment (g2) which is very low compared to that of the heart (due to Vmax proportional to g2; see Equation (4)).
- Their total isometric tension is very low due to their low myosin content compared to that of the heart.
- The duration of their actin-myosin CB cycle (inverse of kcat) is very long due to their dramatically low CB attachment and detachment rate constants.
- The elementary force developed by a single CB and the thermodynamic force are approximately of the same order of magnitude in all three contractile systems described in this study.
- The thermodynamic flow and consequently the rate of entropy production (dis) are dramatically low compared to those of the heart; dis is the entropy change due to irreversible processes within the contractile system and is much lower in non-muscle contractile tissues than in the heart.
- The 3 contractile tissues described here operate near-equilibrium and in a stationary linear regime. However, non-muscle contractile tissues (human placenta and MSC-seeded collagen scaffolds) behave in a manner that is nearer to equilibrium than that observed for the heart.
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooke, R. Actomyosin interaction in striated muscle. Physiol. Rev. 1997, 77, 671–697. [Google Scholar] [CrossRef] [PubMed]
- Huxley, A.F. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 1957, 7, 255–318. [Google Scholar] [CrossRef]
- Atkins, P.W. Physical Chemistry, 5th ed.; Oxford University Press: Oxford, UK; Melbourne, VIC, Australia; Tokio, Japan, 1990; pp. 1–1031. [Google Scholar]
- Prigogine, I. Introduction to Thermodynamics of Irreversible Processes; John Wiley & Sons: New York, NY, USA, 1967. [Google Scholar]
- Kondepudi, D.; Prigogine, I. Modern Thermodynamics from Heat Engines to Dissipative Structures; Wiley & Sons: New York, NY, USA, 1999; pp. 1–486. [Google Scholar]
- Lecarpentier, Y.; Claes, V.; Lecarpentier, E.; Blanc, F.X.; Joseph, T.; Geraets, B.; Krokidis, X.; Hebert, J.L. Comparative statistical mechanics of myosin molecular motors in rat heart, diaphragm and tracheal smooth muscle. C. R. Biol. 2011, 334, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, Y.; Blanc, F.X.; Quillard, J.; Hebert, J.L.; Krokidis, X.; Coirault, C. Statistical mechanics of myosin molecular motors in skeletal muscles. J. Theor. Biol. 2005, 235, 381–392. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Claes, V.; Hebert, J.L.; Krokidis, X.; Blanc, F.X.; Michel, F.; Timbely, O. Statistical Mechanics of the Human Placenta: A Stationary State of a Near-Equilibrium System in a Linear Regime. PLoS ONE 2015, 10, e0142471. [Google Scholar] [CrossRef] [Green Version]
- Gabbiani, G.; Ryan, G.B.; Majne, G. Presence of modified fibroblasts in granulation tissue and their possible role in wound contraction. Experientia 1971, 27, 549–550. [Google Scholar] [CrossRef] [PubMed]
- Gabbiani, G.; Hirschel, B.J.; Ryan, G.B.; Statkov, P.R.; Majno, G. Granulation tissue as a contractile organ. A study of structure and function. J. Exp. Med. 1972, 135, 719–734. [Google Scholar] [CrossRef]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, Y.; Schussler, O.; Claes, V.; Vallée, A. The Myofibroblast: TGFβ-1, A Conductor which Plays a Key Role in Fibrosis by Regulating the Balance between PPARγ and the Canonical WNT Pathway. Nucl. Recept. Res. 2017, 4, 101299. [Google Scholar] [CrossRef] [Green Version]
- Vallee, A.; Lecarpentier, Y. TGF-beta in fibrosis by acting as a conductor for contractile properties of myofibroblasts. Cell Biosci. 2019, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Prunotto, M.; Desmouliere, A.; Varga, J.; De Wever, O.; Mareel, M.; Gabbiani, G. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am. J. Pathol. 2012, 180, 1340–1355. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Graff, A.; Pau, H.; Spahr, R.; Piper, H.M.; Skalli, O.; Gabbiani, G. Appearance of alpha-smooth muscle actin in human eye lens cells of anterior capsular cataract and in cultured bovine lens-forming cells. Differ. Res. Biol. Div. 1990, 43, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Krantz, E.K.; Parker, J.C. Contractile properties of the smooth muscle in the human placenta. Clin. Obstet. Gynecol. 1963, 6, 26–38. [Google Scholar] [CrossRef]
- Farley, A.E.; Graham, C.H.; Smith, G.N. Contractile properties of human placental anchoring villi. Am. J. Physiol. Regulat. Integrat. Comp. Physiol. 2004, 287, R680–R685. [Google Scholar] [CrossRef]
- Feller, A.C.; Schneider, H.; Schmidt, D.; Parwaresch, M.R. Myofibroblast as a major cellular constituent of villous stroma in human placenta. Placenta 1985, 6, 405–415. [Google Scholar] [CrossRef]
- Lecarpentier, E.; Claes, V.; Timbely, O.; Hebert, J.L.; Arsalane, A.; Moumen, A.; Guerin, C.; Guizard, M.; Michel, F.; Lecarpentier, Y. Role of both actin-myosin cross bridges and NO-cGMP pathway modulators in the contraction and relaxation of human placental stem villi. Placenta 2013, 34, 1163–1169. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Schussler, O.; Sakic, A.; Rincon-Garriz, J.M.; Soulie, P.; Bochaton-Piallat, M.L.; Kindler, V. Human Bone Marrow Contains Mesenchymal Stromal Stem Cells That Differentiate In Vitro into Contractile Myofibroblasts Controlling T Lymphocyte Proliferation. Stem Cells Int. 2018, 2018, 6134787. [Google Scholar] [CrossRef] [Green Version]
- Leask, A.; Abraham, D.J. TGF-beta signaling and the fibrotic response. FASEB J. 2004, 18, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Fang, F.; Lam, A.P.; Sargent, J.L.; Hamburg, E.; Hinchcliff, M.E.; Gottardi, C.J.; Atit, R.; Whitfield, M.L.; Varga, J. Wnt/beta-catenin signaling is hyperactivated in systemic sclerosis and induces Smad-dependent fibrotic responses in mesenchymal cells. Arthritis Rheum. 2012, 64, 2734–2745. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Ghosh, A.K.; Sargent, J.L.; Komura, K.; Wu, M.; Huang, Q.Q.; Jain, M.; Whitfield, M.L.; Feghali-Bostwick, C.; Varga, J. PPARgamma downregulation by TGFss in fibroblast and impaired expression and function in systemic sclerosis: A novel mechanism for progressive fibrogenesis. PLoS ONE 2010, 5, e13778. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, S.P.; Reddy, A.T.; Banno, A.; Reddy, R.C. PPAR Agonists for the Prevention and Treatment of Lung Cancer. PPAR Res. 2017, 2017, 8252796. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Claes, V.; Duthoit, G.; Hebert, J.L. Circadian rhythms, Wnt/beta-catenin pathway and PPAR alpha/gamma profiles in diseases with primary or secondary cardiac dysfunction. Front. Physiol. 2014, 5, 429. [Google Scholar] [CrossRef] [Green Version]
- Serini, G.; Bochaton-Piallat, M.L.; Ropraz, P.; Geinoz, A.; Borsi, L.; Zardi, L.; Gabbiani, G. The fibronectin domain ED-A is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1. J. Cell Biol. 1998, 142, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Gabbiani, G.; Chaponnier, C.; Huttner, I. Cytoplasmic filaments and gap junctions in epithelial cells and myofibroblasts during wound healing. J. Cell Biol. 1978, 76, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.K.; Kugler, M.C.; Wolters, P.J.; Robillard, L.; Galvez, M.G.; Brumwell, A.N.; Sheppard, D.; Chapman, H.A. Alveolar epithelial cell mesenchymal transition develops in vivo during pulmonary fibrosis and is regulated by the extracellular matrix. Proc. Natl. Acad. Sci. USA 2006, 103, 13180–13185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, M.A.; Adelstein, R.S. Nonmuscle myosin II moves in new directions. J. Cell Sci. 2008, 121, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, S.; Sakurai, K.; Shinomiya, T.; Fujitani, N.; Key, K.; Ohashi, M. Biochemical and immunohistochemical characterization of the isoforms of myosin and actin in human placenta. Placenta 2011, 32, 347–355. [Google Scholar] [CrossRef]
- Chiavegato, A.; Bochaton-Piallat, M.L.; D’Amore, E.; Sartore, S.; Gabbiani, G. Expression of myosin heavy chain isoforms in mammary epithelial cells and in myofibroblasts from different fibrotic settings during neoplasia. Virchows Arch. 1995, 426, 77–86. [Google Scholar] [CrossRef]
- Parizi, M.; Howard, E.W.; Tomasek, J.J. Regulation of LPA-promoted myofibroblast contraction: Role of Rho, myosin light chain kinase, and myosin light chain phosphatase. Exp. Cell Res. 2000, 254, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef]
- Li, B.; Wang, J.H. Fibroblasts and myofibroblasts in wound healing: Force generation and measurement. J. Tissue Viability 2011, 20, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecarpentier, Y.; Martin, J.L.; Claes, V.; Chambaret, J.P.; Migus, A.; Antonetti, A.; Hatt, P.Y. Real-time kinetics of sarcomere relaxation by laser diffraction. Circ. Res. 1985, 56, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brutsaert, D.L.; Claes, V.A.; Goethals, M.A. Effect of calcium on force-velocity-length relations of heart muscle of the cat. Circ. Res. 1973, 32, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.V. The heat of shortening and the dynamic constants of muscle. Proc. R. Soc. Lond. Biol. Sci. 1938, 126, 136–195. [Google Scholar]
- Lecarpentier, Y.; Chemla, D.; Blanc, F.X.; Pourny, J.C.; Joseph, T.; Riou, B.; Coirault, C. Mechanics, energetics, and crossbridge kinetics of rabbit diaphragm during congestive heart failure. FASEB J. 1998, 12, 981–989. [Google Scholar] [CrossRef]
- Woledge, R.C.; Curtin, A.N.; Homsher, E. Energetic Aspects of Muscle Contraction; Academic Press: London, UK, 1985; Volume 41, pp. 1–357. [Google Scholar]
- Veech, R.L.; Lawson, J.W.; Cornell, N.W.; Krebs, H.A. Cytosolic phosphorylation potential. J. Biol. Chem. 1979, 254, 6538–6547. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Claes, V.; Krokidis, X.; Vallée, A.A. Comparative Statistical Mechanics of Muscle and Non-Muscle Contractile Systems: Stationary States of Near-Equilibrium Systems in A Linear Regime. Entropy J. 2017, 19, 558. [Google Scholar] [CrossRef] [Green Version]
- Onsager, L. Reciprocal relations in irreversible processes II. Phys. Rev. 1931, 38, 405–426. [Google Scholar] [CrossRef]
- Prigogine, I. Life and physics. New perspectives. Cell Biophys. 1986, 9, 217–224. [Google Scholar] [CrossRef]
- De Donder, T. L’ affinité; Gauthiers-Villars: Paris, France, 1927. [Google Scholar]
- Frank, O. Zur Dynamik des Herzmuskels. Z. Biol. 1895, 32, 273–277. [Google Scholar]
- Starling, E.H. The Linacre Lecture on the Law of the Heart; Longmans, Green & Co: London, UK, 1918. [Google Scholar]
- Benirschke, K.; Kaufmann, P.; Baergen, R.N. Pathology of the Human Placenta; Springer: New York, NY, USA, 2006; p. 1069. [Google Scholar]
- Graf, R.; Langer, J.U.; Schonfelder, G.; Oney, T.; Hartel-Schenk, S.; Reutter, W.; Schmidt, H.H. The extravascular contractile system in the human placenta. Morphological and immunocytochemical investigations. Anat. Embryol. 1994, 190, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Demir, R.; Kosanke, G.; Kohnen, G.; Kertschanska, S.; Kaufmann, P. Classification of human placental stem villi: Review of structural and functional aspects. Microsc. Res. Tech. 1997, 38, 29–41. [Google Scholar] [CrossRef]
- Sparn, H.G.; Lieder-Ochs, B.A.; Franke, W.W. Immunohistochemical identification and characterization of a special type of desmin-producing stromal cells in human placenta and other fetal tissues. Differ. Res. Biol. Div. 1994, 56, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kohnen, G.; Castellucci, M.; Hsi, B.L.; Yeh, C.J.; Kaufmann, P. The monoclonal antibody GB 42—A useful marker for the differentiation of myofibroblasts. Cell Tissue Res. 1995, 281, 231–242. [Google Scholar] [CrossRef]
- Iizuka, S. Uber Verkammen von Muskelfasern in der menschlichen Placenta. Beitr. Geburtsh. Gynaek. 1916, 19, 101. [Google Scholar]
- Naujoks, H. Heben anatomische Veränderungen der kindlichen Eihäute einen Einfluss auf die Zeit des Blasensprunges. Z. Geburtsh. Gynaek. 1922, 84, 304. [Google Scholar]
- Dubreuil, G.; Rivière, M. Formations fibromusculaires du chorion et villosités du placenta humain. C. R. Soc. Biol. 1932, 111, 170–172. [Google Scholar]
- Spanner, R. Mütterlicher und kindlicher kreislauf der menschlichen Placenta und seine Strömbahnen. Z. Anat. Entwicklungsgesch. 1935, 105, 163–242. [Google Scholar] [CrossRef]
- Graf, R.; Schonfelder, G.; Muhlberger, M.; Gutsmann, M. The perivascular contractile sheath of human placental stem villi: Its isolation and characterization. Placenta 1995, 16, 57–66. [Google Scholar] [CrossRef]
- Graf, R.; Matejevic, D.; Schuppan, D.; Neudeck, H.; Shakibaei, M.; Vetter, K. Molecular anatomy of the perivascular sheath in human placental stem villi: The contractile apparatus and its association to the extracellular matrix. Cell Tissue Res. 1997, 290, 601–607. [Google Scholar] [CrossRef]
- King, T.M.; Groeschel-Stewart, U. Placental Contractile Protein. Am. J. Obstet. Gynecol. 1965, 93, 253–258. [Google Scholar] [CrossRef]
- Michael, C. Actomyosin content of the human placenta. J. Obstet. Gynaecol. 1974, 81, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Huszar, G.; Bailey, P. Isolation and characterization of myosin in the human term placenta. Am. J. Obstet. Gynecol. 1979, 135, 707–712. [Google Scholar] [CrossRef]
- Lecarpentier, E.E.; Claes, V.A.; Timbely, O.; Arsalane, A.; Wipff, J.A.; Hebert, J.L.; Michel, F.Y.; Lecarpentier, Y.C. Mechanics and energetics of myosin molecular motors from non-pregnant human myometrium. J. Appl. Physiol. 2011, 111, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, Y.; Claes, V.; Lecarpentier, E.; Guerin, C.; Hebert, J.L.; Arsalane, A.; Moumen, A.; Krokidis, X.; Michel, F.; Timbely, O. Ultraslow myosin molecular motors of placental contractile stem villi in humans. PLoS ONE 2014, 9, e108814. [Google Scholar]
- Kovacs, M.; Wang, F.; Hu, A.; Zhang, Y.; Sellers, J.R. Functional divergence of human cytoplasmic myosin II: Kinetic characterization of the non-muscle IIA isoform. J. Biol. Chem. 2003, 278, 38132–38140. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.M.; Huxley, A.F.; Julian, F.J. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. J. Physiol. 1966, 184, 170–192. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, Y.; Kindler, V.; Bochaton-Piallat, M.L.; Sakic, A.; Claes, V.; Hebert, J.L.; Vallee, A.; Schussler, O. Tripeptide Arg-Gly-Asp (RGD) modifies the molecular mechanical properties of the non-muscle myosin IIA in human bone marrow-derived myofibroblasts seeded in a collagen scaffold. PLoS ONE 2019, 14, e0222683. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, Y.; Kindler, V.; Krokidis, X.; Bochaton-Piallat, M.L.; Claes, V.; Hebert, J.L.; Vallee, A.; Schussler, O. Statistical Mechanics of Non-Muscle Myosin IIA in Human Bone Marrow-Derived Mesenchymal Stromal Cells Seeded in a Collagen Scaffold: A Thermodynamic Near-Equilibrium Linear System Modified by the Tripeptide Arg-Gly-Asp (RGD). Cells 2020, 9, 1510. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placenta | MSC-Seeded Scaffold | Heart | |
|---|---|---|---|
| Vmax (Lo/s) | 0.002 | 0.002 | 3.7 |
| Tension (mN) | 1.5 | 0.3 | 42 |
| kcat (s−1) | 0.003 | 0.004 | 20.6 |
| Unitary CB force (pN) | 2.0 | 2.1 | 1.6 |
| max Efficiency (%) | 36 | 38 | 28 |
| Myosin content (nmol/g) | 0.14 | 0.07 | 12.5 |
| G | 3.7 | 4.1 | 1.6 |
| PD3 (highest CB probability) | 0.80 | 0.82 | 0.58 |
| f1 (s−1) | 0.08 | 0.08 | 306 |
| g1 (s−1) | 0.03 | 0.03 | 194 |
| g2 (s−1) | 0.34 | 0.35 | 731 |
| Microcanonical partition function | 1.25 | 1.22 | 1.72 |
| Statistical entropy (J/T/mol) | 5.6 | 10 | 9.3 |
| Internal energy (J/mol) | 1097 | 2000 | 1401 |
| Affinity (J/mol) | 354 | 250 | 535 |
| Thermodynamic force | 1.2 | 0.9 | 1.8 |
| Thermodynamic flow (mol/L/s) | 4.8 × 10−13 | 1.6 × 10−13 | 2.8 |
| Entropy production rate (J/T/L/s) | 8.2 × 10−13 | 1.8 x 10−13 | 6.5 × 10 −7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lecarpentier, Y.; Claes, V.; Hébert, J.-L.; Schussler, O.; Vallée, A. Mechanical and Thermodynamic Properties of Non-Muscle Contractile Tissues: The Myofibroblast and the Molecular Motor Non-Muscle Myosin Type IIA. Int. J. Mol. Sci. 2021, 22, 7738. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147738
Lecarpentier Y, Claes V, Hébert J-L, Schussler O, Vallée A. Mechanical and Thermodynamic Properties of Non-Muscle Contractile Tissues: The Myofibroblast and the Molecular Motor Non-Muscle Myosin Type IIA. International Journal of Molecular Sciences. 2021; 22(14):7738. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147738
Chicago/Turabian StyleLecarpentier, Yves, Victor Claes, Jean-Louis Hébert, Olivier Schussler, and Alexandre Vallée. 2021. "Mechanical and Thermodynamic Properties of Non-Muscle Contractile Tissues: The Myofibroblast and the Molecular Motor Non-Muscle Myosin Type IIA" International Journal of Molecular Sciences 22, no. 14: 7738. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147738