
Loss of ZNF677 Expression Is an Independent Predictor for Distant Metastasis in Middle Eastern Papillary Thyroid Carcinoma Patients


Abstract
:1. Introduction
2. Results
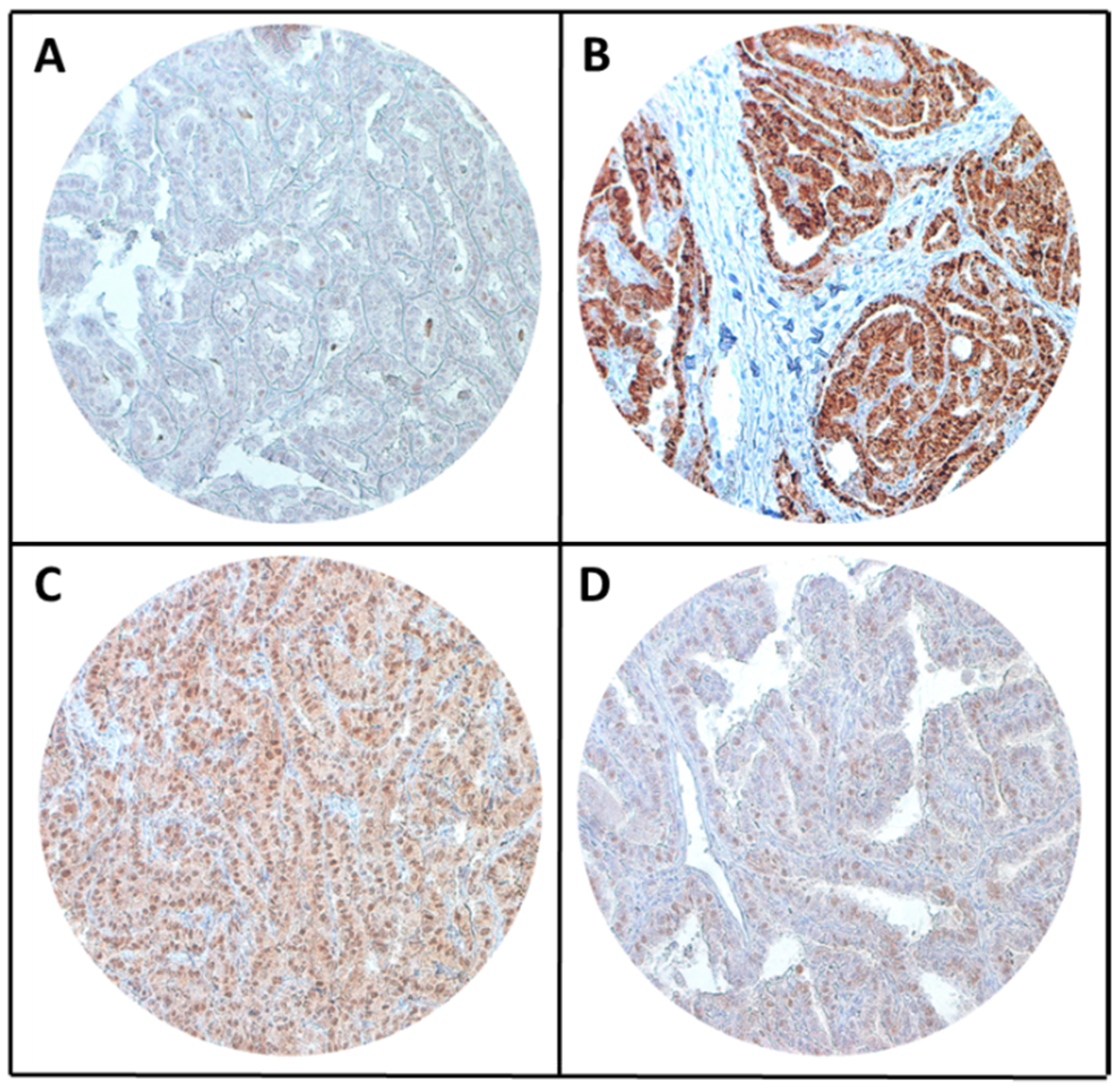
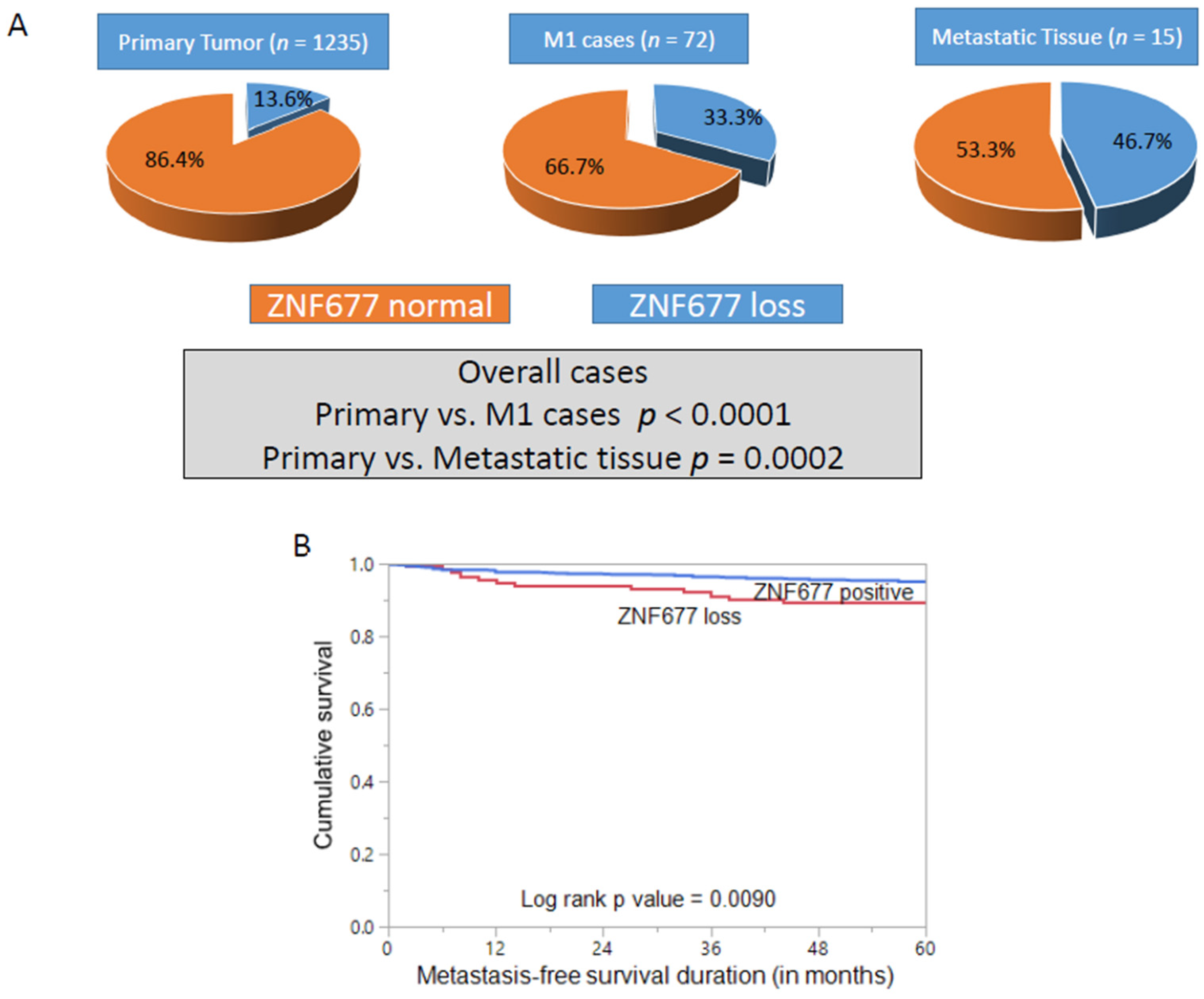
2.1. ZNF677 Expression and Its Clinico-Pathological Associations
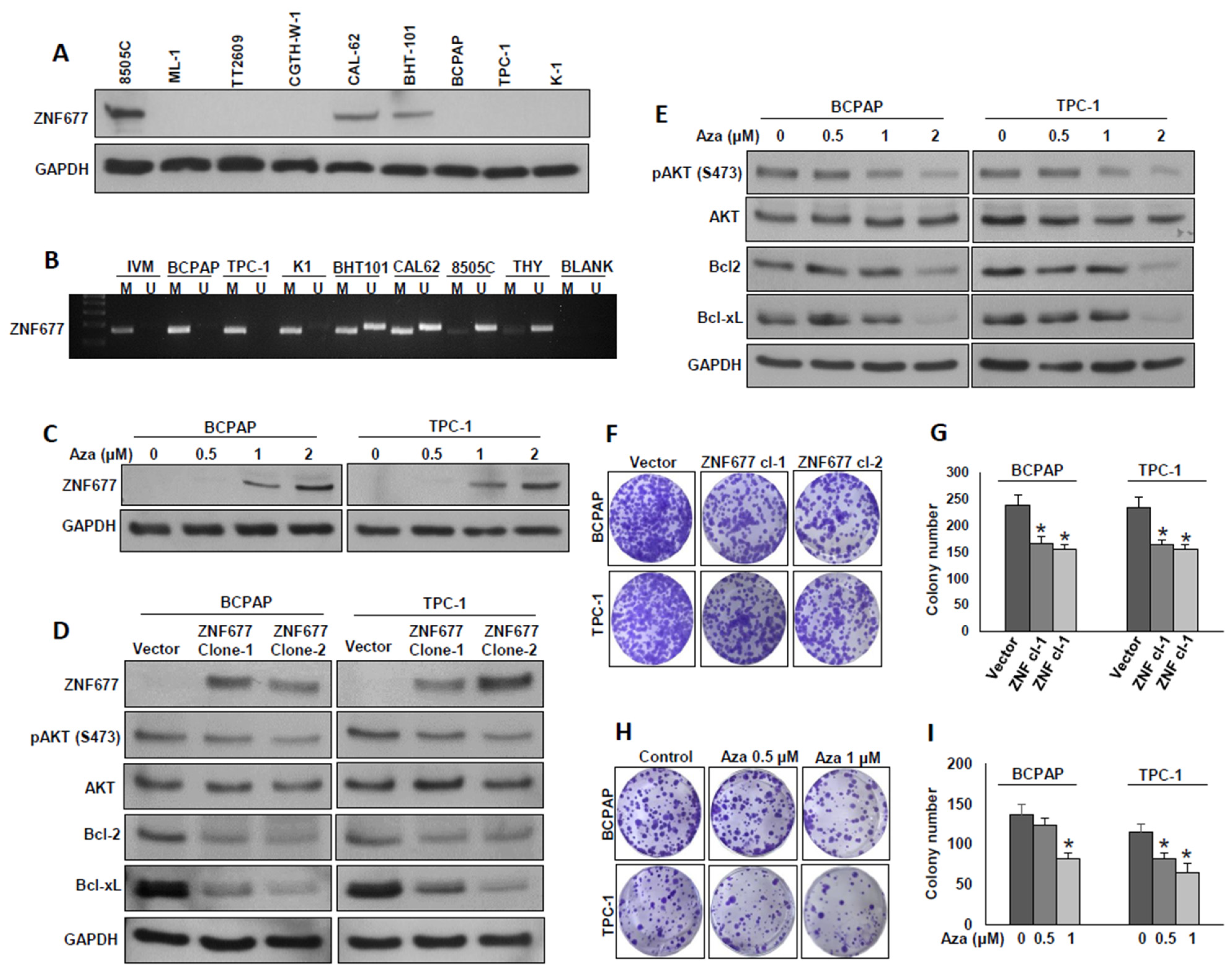
2.2. ZNF677 Impedes PTC Cell Growth In Vitro
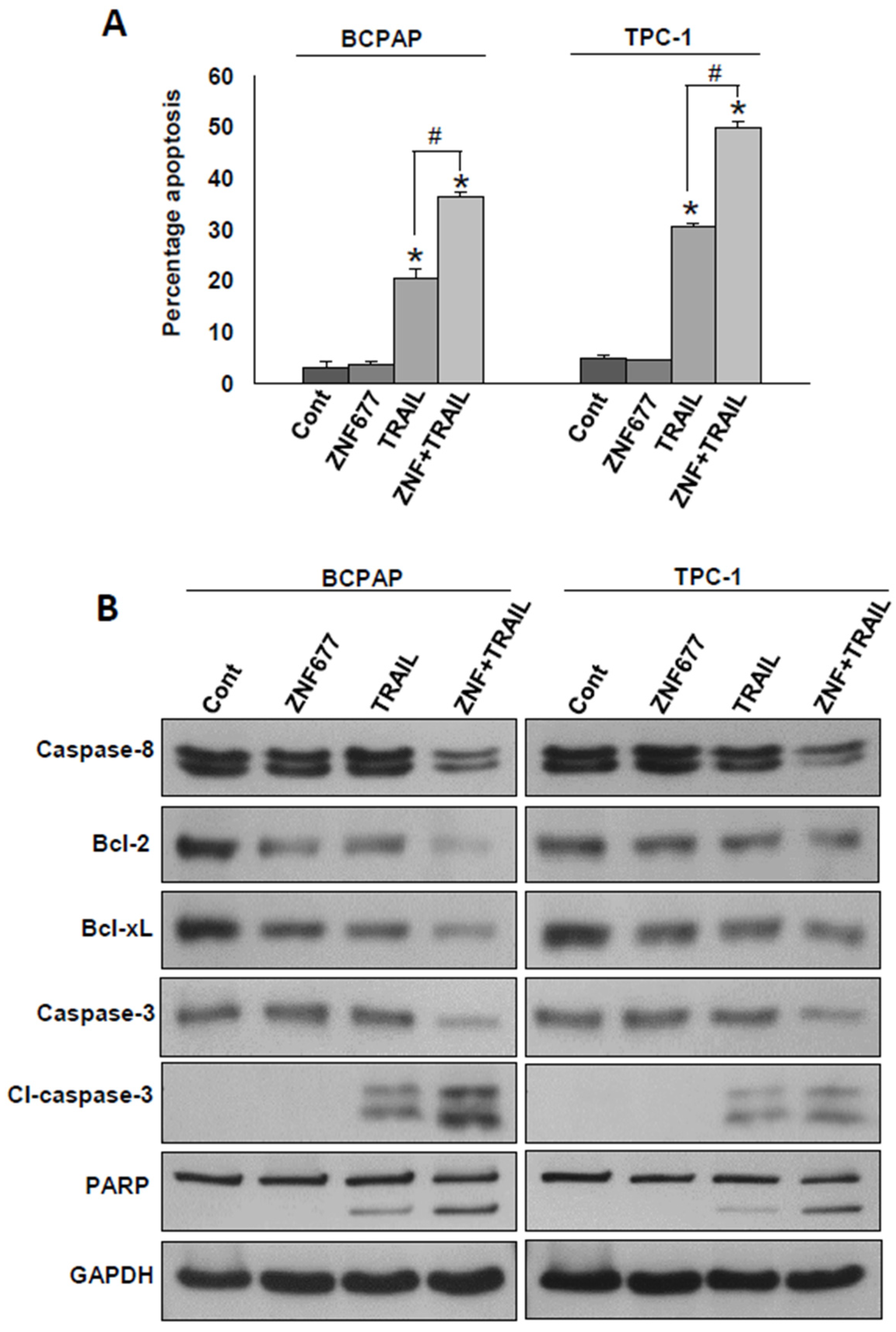
2.3. Ectopic Expression of ZNF677 Potentiates TRAIL-Induced Apoptosis in PTC Cells
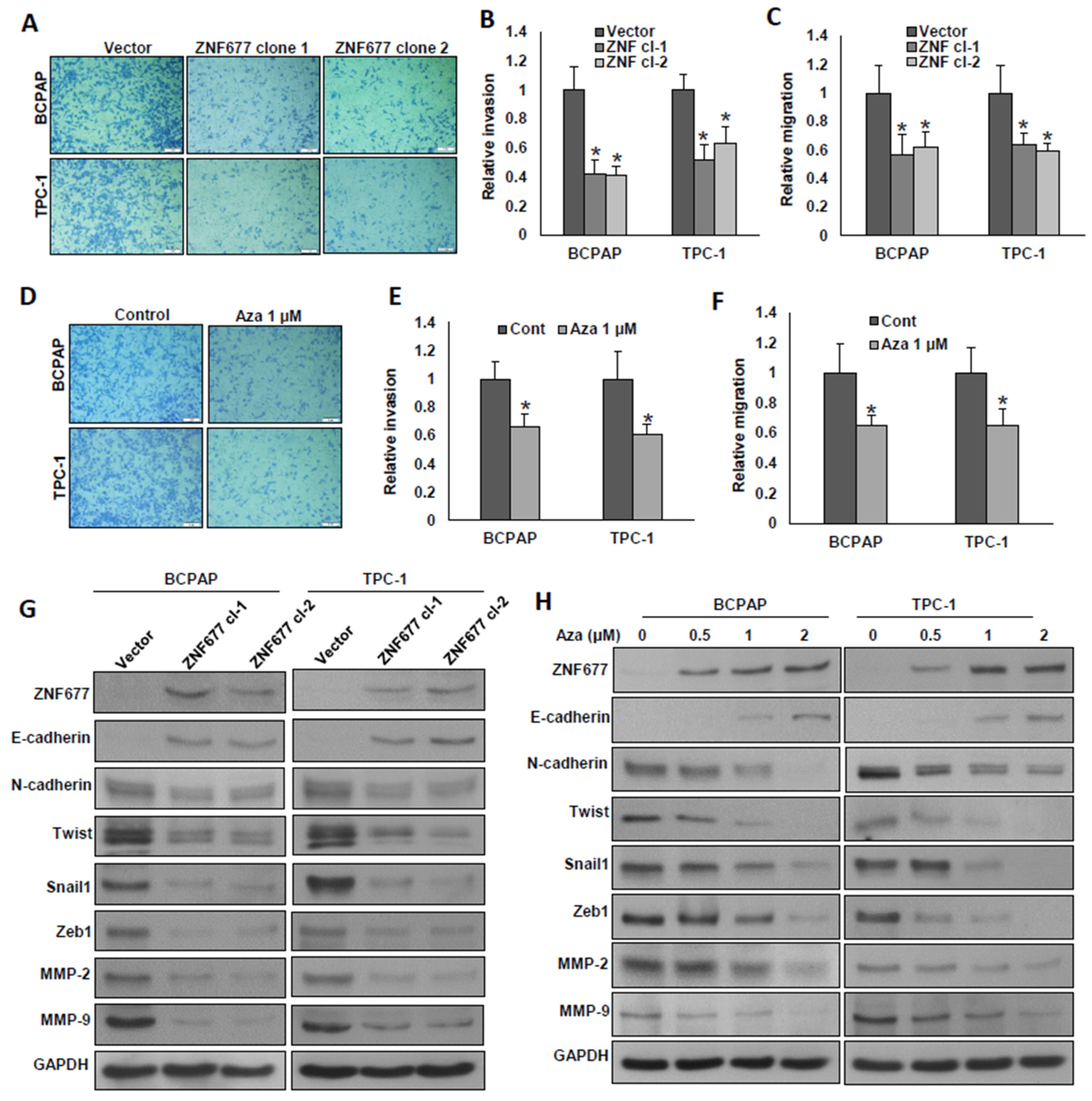
2.4. ZNF677 Decreases the Metastatic Potential of PTC Cells
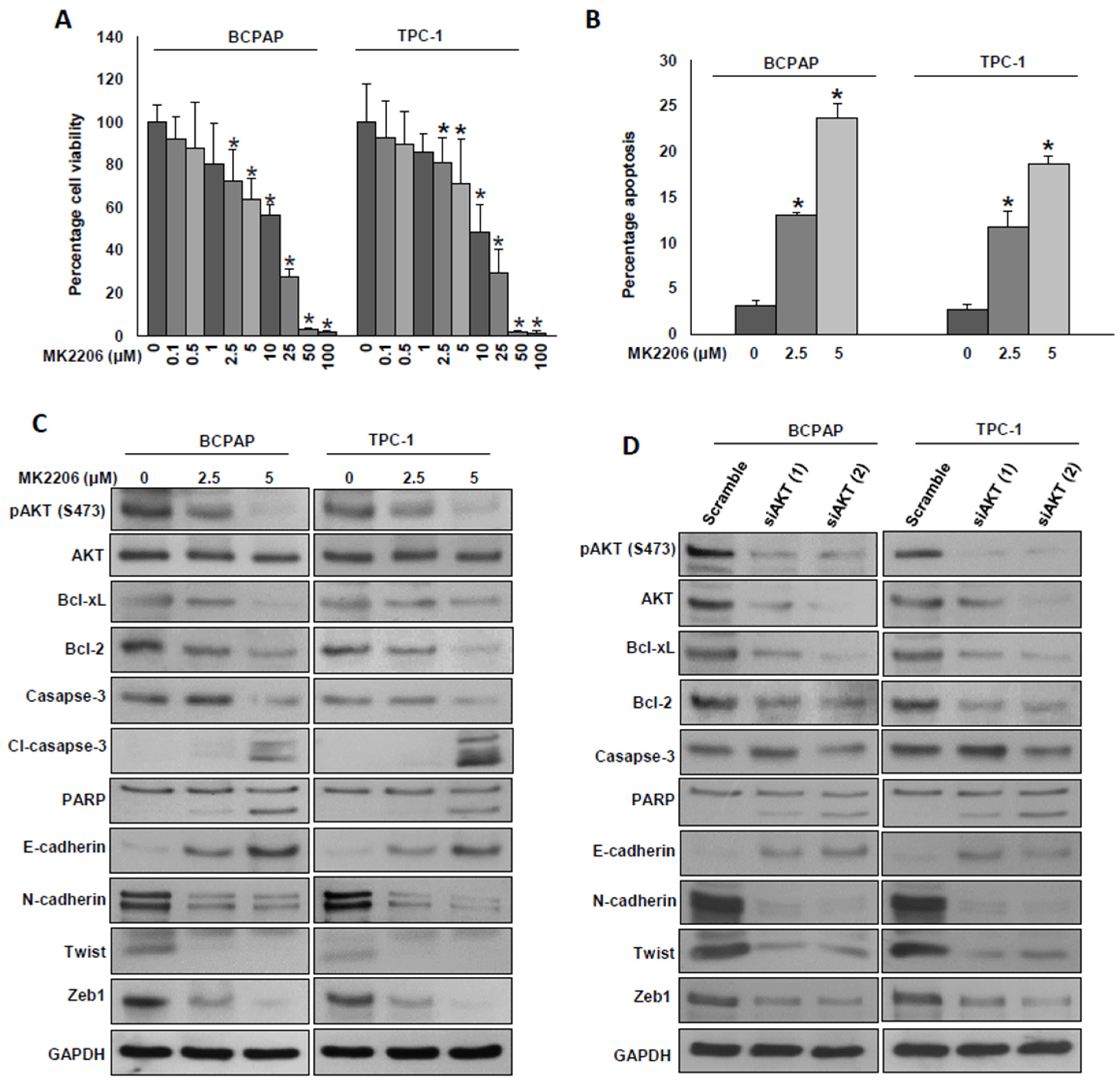
2.5. Inhibition of AKT Decreases Cell Growth and Attenuates EMT in PTC Cells
3. Discussion
4. Materials and Methods
4.1. Sample Selection
4.2. Tissue Microarray (TMA) Construction and Immunohistochemistry (IHC)
4.3. Bisulfite Modification and Methylation-Specific PCR
4.4. Cell Culture
4.5. Reagents and Antibodies
4.6. Clonogenic Assay
4.7. Annexin V Staining
4.8. Gene Silencing Using siRNA
4.9. Plasmid and Transfection
4.10. Cell Invasion and Migration Assays
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.; Dal Maso, L.; Vaccarella, S. Global trends in thyroid cancer incidence and the impact of overdiagnosis. Lancet Diabetes Endocrinol. 2020, 8, 468–470. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Devesa, S.S.; Sosa, J.A.; Check, D.; Kitahara, C.M. Trends in thyroid cancer incidence and mortality in the United States, 1974–2013. JAMA 2017, 317, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA A Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Kitahara, C.M.; Sosa, J.A. The changing incidence of thyroid cancer. Nat. Rev. Endocrinol. 2016, 12, 646–653. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.; Krapcho, M.; Garshell, J.; Miller, D.; Altekruse, S.; Kosary, C.; Yu, M.; Ruhl, J.; Tatalovich, Z. SEER Cancer Statistics Review, 1975–2012; National Cancer Institute: Bethesda, MD, USA, 2015. [Google Scholar]
- Adam, M.A.; Pura, J.; Gu, L.; Dinan, M.A.; Tyler, D.S.; Reed, S.D.; Roman, S.A.; Sosa, J.A. Extent of surgery for papillary thyroid cancer is not associated with survival: An analysis of 61,775 patients. Ann. Surg. 2014, 260, 601. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.R.; Lai, P.; Hall, F.; Borglund, A.; Eski, S.; Freeman, J.L. Variables predicting distant metastases in thyroid cancer. Laryngoscope 2005, 115, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Benbassat, C.A.; Mechlis-Frish, S.; Hirsch, D. Clinicopathological characteristics and long-term outcome in patients with distant metastases from differentiated thyroid cancer. World J. Surg. 2006, 30, 1088–1095. [Google Scholar] [CrossRef]
- Casara, D.; Rubello, D.; Saladini, G.; Masarotto, G.; Favero, A.; Girelli, M.E.; Busnardo, B. Different features of pulmonary metastases in differentiated thyroid cancer: Natural history and multivariate statistical analysis of prognostic variables. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 1993, 34, 1626–1631. [Google Scholar]
- Wang, L.Y.; Palmer, F.L.; Nixon, I.J.; Thomas, D.; Patel, S.G.; Shaha, A.R.; Shah, J.P.; Tuttle, R.M.; Ganly, I. Multi-organ distant metastases confer worse disease-specific survival in differentiated thyroid cancer. Thyroid 2014, 24, 1594–1599. [Google Scholar] [CrossRef] [PubMed]
- Khan, U.; Al Afif, A.; Aldaihani, A.; MacKay, C.; Rigby, M.H.; Rajaraman, M.; Imran, S.A.; Bullock, M.J.; Taylor, S.M.; Trites, J.R. Patient and tumor factors contributing to distant metastasis in well-differentiated thyroid cancer: A retrospective cohort study. J. Otolaryngol. Head Neck Surg. 2020, 49, 78. [Google Scholar] [CrossRef]
- Lang, B.H.-H.; Wong, K.P.; Cheung, C.Y.; Wan, K.Y.; Lo, C.-Y. Evaluating the prognostic factors associated with cancer-specific survival of differentiated thyroid carcinoma presenting with distant metastasis. Ann. Surg. Oncol. 2013, 20, 1329–1335. [Google Scholar] [CrossRef] [Green Version]
- Sampson, E.; Brierley, J.D.; Le, L.W.; Rotstein, L.; Tsang, R.W. Clinical management and outcome of papillary and follicular (differentiated) thyroid cancer presenting with distant metastasis at diagnosis. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2007, 110, 1451–1456. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-D.; Hsueh, C.; Chao, T.-C. Long-term follow-up of the therapeutic outcomes for papillary thyroid carcinoma with distant metastasis. Medicine 2015, 94, e1063. [Google Scholar] [CrossRef] [PubMed]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M. 2015 American Thyroid Association management guidelines for adult patients with thyroid nodules and differentiated thyroid cancer: The American Thyroid Association guidelines task force on thyroid nodules and differentiated thyroid cancer. Thyroid 2016, 26, 1–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschellà, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef] [Green Version]
- Lupo, A.; Cesaro, E.; Montano, G.; Zurlo, D.; Izzo, P.; Costanzo, P. KRAB-zinc finger proteins: A repressor family displaying multiple biological functions. Curr. Genom. 2013, 14, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Ecco, G.; Imbeault, M.; Trono, D. KRAB zinc finger proteins. Development 2017, 144, 2719–2729. [Google Scholar] [CrossRef] [Green Version]
- Oleksiewicz, U.; Gładych, M.; Raman, A.T.; Heyn, H.; Mereu, E.; Chlebanowska, P.; Andrzejewska, A.; Sozańska, B.; Samant, N.; Fąk, K. TRIM28 and interacting KRAB-ZNFs control self-renewal of human pluripotent stem cells through epigenetic repression of pro-differentiation genes. Stem Cell Rep. 2017, 9, 2065–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, G.; Greenberg, D.; Macfarlan, T.S. Spotting the enemy within: Targeted silencing of foreign DNA in mammalian genomes by the Krüppel-associated box zinc finger protein family. Mob. DNA 2015, 6, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazanets, A.; Shorstova, T.; Hilmi, K.; Marques, M.; Witcher, M. Epigenetic silencing of tumor suppressor genes: Paradigms, puzzles, and potential. Biochim. Biophys. Acta (BBA) Rev. Cancer 2016, 1865, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA methyltransferases in cancer: Biology, paradox, aberrations, and targeted therapy. Cancers 2020, 12, 2123. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Peng, Y.; Gao, A.; Du, C.; Herman, J.G. Epigenetic heterogeneity in cancer. Biomark. Res. 2019, 7, 1–19. [Google Scholar] [CrossRef]
- Schmitges, F.W.; Radovani, E.; Najafabadi, H.S.; Barazandeh, M.; Campitelli, L.F.; Yin, Y.; Jolma, A.; Zhong, G.; Guo, H.; Kanagalingam, T. Multiparameter functional diversity of human C2H2 zinc finger proteins. Genome Res. 2016, 26, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.E.; Gadd, S.; Huff, V.; Gerhard, D.S.; Dome, J.S.; Perlman, E.J. A unique subset of low-risk Wilms tumors is characterized by loss of function of TRIM28 (KAP1), a gene critical in early renal development: A Children’s Oncology Group study. PLoS ONE 2018, 13, e0208936. [Google Scholar] [CrossRef] [Green Version]
- Heller, G.; Altenberger, C.; Schmid, B.; Marhold, M.; Tomasich, E.; Ziegler, B.; Müllauer, L.; Minichsdorfer, C.; Lang, G.; End-Pfützenreuter, A. DNA methylation transcriptionally regulates the putative tumor cell growth suppressor ZNF677 in non-small cell lung cancers. Oncotarget 2015, 6, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, Q.; Guan, H.; Shi, B.; Ji, M.; Hou, P. ZNF677 suppresses Akt phosphorylation and tumorigenesis in thyroid cancer. Cancer Res. 2018, 78, 5216–5228. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Dai, Q.; Su, X.; Fu, J.; Feng, X.; Peng, J. Role of PI3K/AKT pathway in cancer: The framework of malignant behavior. Mol. Biol. Rep. 2020, 47, 4587–4629. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Goffredo, P.; Sosa, J.A.; Roman, S.A. Differentiated thyroid cancer presenting with distant metastases: A population analysis over two decades. World J. Surg. 2013, 37, 1599–1605. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lim, Y.S.; Lee, J.-C.; Wang, S.-G.; Kim, I.-J.; Son, S.-M.; Lee, B.-J. Clinical implications of bilateral lateral cervical lymph node metastasis in papillary thyroid cancer: A risk factor for lung metastasis. Ann. Surg. Oncol. 2011, 18, 3486–3492. [Google Scholar] [CrossRef]
- Devlin, J.R.; Verschuren, E.W. More than a tumor suppressor: E-cadherin loss drives lung cancer metastasis. Am. Thorac. Soc. 2018, 59. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Orlandi, R.; Zhao, N.; Carcangiu, M.L.; Tagliabue, E.; Xu, J.; Bast, R.C.; Yu, Y. Tumor suppressor genes are frequently methylated in lymph node metastases of breast cancers. BMC Cancer 2010, 10, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, L.; Ding, Y. Screening of tumor suppressor genes in metastatic colorectal cancer. BioMed Res. Int. 2017, 2017, 2769140. [Google Scholar] [CrossRef]
- Justiniano, S.E.; McElroy, J.P.; Yu, L.; Yilmaz, A.S.; Coombes, K.R.; Senter, L.; Nagy, R.; Wakely, P., Jr.; Volinia, S.; Vinco, M. Genetic variants in thyroid cancer distant metastases. Endocr. Relat. Cancer 2016, 23, L33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Wang, Z.; Xu, L.; Sun, L.; Song, H.; Yin, H.; He, F. lncRNA SNHG3 acts as a novel Tumor Suppressor and regulates Tumor Proliferation and Metastasis via AKT/mTOR/ERK pathway in Papillary Thyroid Carcinoma. J. Cancer 2020, 11, 3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siraj, A.; Bavi, P.; Abubaker, J.; Jehan, Z.; Sultana, M.; Al-Dayel, F.; Al-Nuaim, A.; Alzahrani, A.; Ahmed, M.; Al-Sanea, O. Genome-wide expression analysis of Middle Eastern papillary thyroid cancer reveals c-MET as a novel target for cancer therapy. J. Pathol. 2007, 213, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Bavi, P.; Jehan, Z.; Atizado, V.; Al-Dossari, H.; Al-Dayel, F.; Tulbah, A.; Amr, S.S.; Sheikh, S.S.; Ezzat, A.; El-Solh, H. Prevalence of fragile histidine triad expression in tumors from saudi arabia: A tissue microarray analysis. Cancer Epidemiol. Prev. Biomark. 2006, 15, 1708–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, S.; Siraj, A.K.; Al-Rasheed, M.; Ahmed, M.; Bu, R.; Myers, J.N.; Al-Nuaim, A.; Al-Sobhi, S.; Al-Dayel, F.; Bavi, P. Fatty acid synthase and AKT pathway signaling in a subset of papillary thyroid cancers. J. Clin. Endocrinol. Metab. 2008, 93, 4088–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, T.; Suzuki, J.; Wang, Y.V.; Menendez, S.; Morera, L.B.; Raya, A.; Wahl, G.M.; Izpisua Belmonte, J.C. Linking the p53 tumour suppressor pathway to somatic cell reprogramming. Nature 2009, 460, 1140–1144. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Sano, D.; Pickering, C.R.; Jasser, S.A.; Henderson, Y.C.; Clayman, G.L.; Sturgis, E.M.; Ow, T.J.; Lotan, R.; Carey, T.E. Assembly and initial characterization of a panel of 85 genomically validated cell lines from diverse head and neck tumor sites. Clin. Cancer Res. 2011, 17, 7248–7264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, R.; Siraj, A.K.; Divya, S.P.; Kong, Y.; Parvathareddy, S.K.; Al-Rasheed, M.; Al-Obaisi, K.A.; Victoria, I.G.; Al-Sobhi, S.S.; Al-Dawish, M. Telomerase reverse transcriptase mutations are independent predictor of disease-free survival in M iddle E astern papillary thyroid cancer. Int. J. Cancer 2018, 142, 2028–2039. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.R.; Siraj, A.K.; Ahmed, M.; Bu, R.; Pratheeshkumar, P.; Alrashed, A.M.; Qadri, Z.; Ajarim, D.; Al-Dayel, F.; Beg, S. XIAP over-expression is an independent poor prognostic marker in Middle Eastern breast cancer and can be targeted to induce efficient apoptosis. BMC Cancer 2017, 17, 640. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinico-Pathological Characteristics | n (%) |
|---|---|
| Age | |
| Median | 37.5 |
| Range (IQR) | 28.7–50.5 |
| <55 years | 1005 (81.4) |
| ≥55 years | 230 (18.6) |
| Gender | |
| Female | 938 (76.0) |
| Male | 297 (24.0) |
| Histopathology | |
| Classical Variant | 850 (68.8) |
| Follicular Variant | 194 (15.7) |
| Tall Cell Variant | 106 (8.6) |
| Others | 85 (6.9) |
| Extra Thyroidal Extension | |
| Absent | 640 (51.8) |
| Present | 595 (48.2) |
| pT | |
| T1 | 318 (25.7) |
| T2 | 250 (20.2) |
| T3 | 517 (41.9) |
| T4 | 105 (8.5) |
| Unknown | 45 (3.6) |
| pN | |
| N0 | 467 (37.8) |
| N1 | 633 (51.3) |
| Nx | 135 (10.9) |
| pM | |
| M0 | 1091 (88.4) |
| M1 | 72 (5.8) |
| Mx | 72 (5.8) |
| Stage | |
| I | 1001 (81.1) |
| II | 135 (10.9) |
| III | 20 (1.6) |
| IVA | 17 (1.4) |
| IVB | 27 (2.2) |
| Unknown | 35 (2.8) |
| Clinico-Pathological Characteristics | Total | ZNF677 Loss | ZNF677 Normal | p Value | |||
|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | ||
| No. of patients | 1235 | 168 | 13.6 | 1067 | 86.4 | ||
| Age (years) | |||||||
| <55 | 1005 | 81.4 | 136 | 13.5 | 869 | 86.5 | 0.8795 |
| ≥55 | 230 | 18.6 | 32 | 13.9 | 198 | 86.1 | |
| Sex | |||||||
| Female | 938 | 75.9 | 122 | 13.0 | 816 | 87.0 | 0.2829 |
| Male | 297 | 24.1 | 46 | 15.5 | 251 | 84.5 | |
| Extrathyroidal extension | |||||||
| Absent | 640 | 51.8 | 67 | 10.5 | 573 | 89.5 | 0.0008 * |
| Present | 595 | 48.2 | 101 | 17.0 | 494 | 83.0 | |
| pT | |||||||
| pT1 | 318 | 26.7 | 34 | 10.7 | 284 | 89.3 | 0.2812 |
| pT2 | 250 | 21.0 | 40 | 16.0 | 210 | 84.0 | |
| pT3 | 517 | 43.5 | 70 | 13.5 | 447 | 86.5 | |
| pT4 | 105 | 8.8 | 16 | 15.2 | 89 | 84.8 | |
| pN | |||||||
| pN0 | 467 | 42.5 | 67 | 14.4 | 400 | 85.6 | 0.5560 |
| pN1 | 633 | 57.5 | 83 | 13.1 | 550 | 86.9 | |
| pM | |||||||
| pM0 | 1091 | 93.8 | 142 | 13.0 | 949 | 87.0 | <0.0001 * |
| pM1 | 72 | 6.2 | 24 | 33.3 | 48 | 66.7 | |
| Stage | |||||||
| I | 1001 | 83.4 | 134 | 13.4 | 867 | 86.6 | 0.0431 * |
| II | 135 | 11.2 | 20 | 14.8 | 115 | 85.2 | |
| III | 20 | 1.7 | 3 | 15.0 | 17 | 85.0 | |
| IVA | 17 | 1.4 | 0 | 0.0 | 17 | 100.0 | |
| IVB | 27 | 2.3 | 8 | 29.6 | 19 | 70.4 | |
| Histology Type | |||||||
| Classical Variant | 850 | 68.8 | 113 | 13.3 | 737 | 86.7 | <0.0001 * |
| Follicular Variant | 194 | 15.7 | 43 | 22.2 | 151 | 77.8 | |
| Tall-Cell Variant | 106 | 8.6 | 5 | 4.7 | 101 | 95.3 | |
| Other variants | 85 | 6.9 | 7 | 8.2 | 78 | 81.8 | |
| phospho-AKT IHC | |||||||
| High | 499 | 40.4 | 116 | 23.3 | 383 | 76.7 | <0.0001 * |
| Low | 736 | 59.6 | 52 | 7.1 | 684 | 92.9 | |
| Metastasis Free Survival | |||||||
| 5 years | 89.2 | 95.1 | 0.0090 * | ||||
| Clinico-Pathological Variables | Odds Ratio | 95% CI | p-Value |
|---|---|---|---|
| Age ≥55 years (vs. <55 years) | 0.66 | 0.25–1.72 | 0.3969 |
| Sex Male (vs. Female) | 2.31 | 1.17–4.56 | 0.0158 * |
| Extrathyroidal extension Present (vs. Absent) | 2.07 | 0.90–4.75 | 0.0876 |
| Histologic subtype Tall cell variant (vs. Other variants) | 0.73 | 0.23–2.28 | 0.5868 |
| Stage III–IV (vs. I–II) | 33.62 | 12.37–91.39 | <0.0001 * |
| ZNF677 IHC Loss (vs. positive) | 2.60 | 1.20–5.62 | 0.0155 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siraj, A.K.; Poyil, P.K.; Parvathareddy, S.K.; Alobaisi, K.; Ahmed, S.O.; Al-Sobhi, S.S.; Al-Dayel, F.; Al-Kuraya, K.S. Loss of ZNF677 Expression Is an Independent Predictor for Distant Metastasis in Middle Eastern Papillary Thyroid Carcinoma Patients. Int. J. Mol. Sci. 2021, 22, 7833. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157833
Siraj AK, Poyil PK, Parvathareddy SK, Alobaisi K, Ahmed SO, Al-Sobhi SS, Al-Dayel F, Al-Kuraya KS. Loss of ZNF677 Expression Is an Independent Predictor for Distant Metastasis in Middle Eastern Papillary Thyroid Carcinoma Patients. International Journal of Molecular Sciences. 2021; 22(15):7833. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157833
Chicago/Turabian StyleSiraj, Abdul K., Pratheesh Kumar Poyil, Sandeep Kumar Parvathareddy, Khadija Alobaisi, Saeeda O. Ahmed, Saif S. Al-Sobhi, Fouad Al-Dayel, and Khawla S. Al-Kuraya. 2021. "Loss of ZNF677 Expression Is an Independent Predictor for Distant Metastasis in Middle Eastern Papillary Thyroid Carcinoma Patients" International Journal of Molecular Sciences 22, no. 15: 7833. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157833