
The Role of P-Selectin in COVID-19 Coagulopathy: An Updated Review

 ,
,
Abstract
:1. Introduction
2. Selectins
3. P-Selectin in Human Diseases
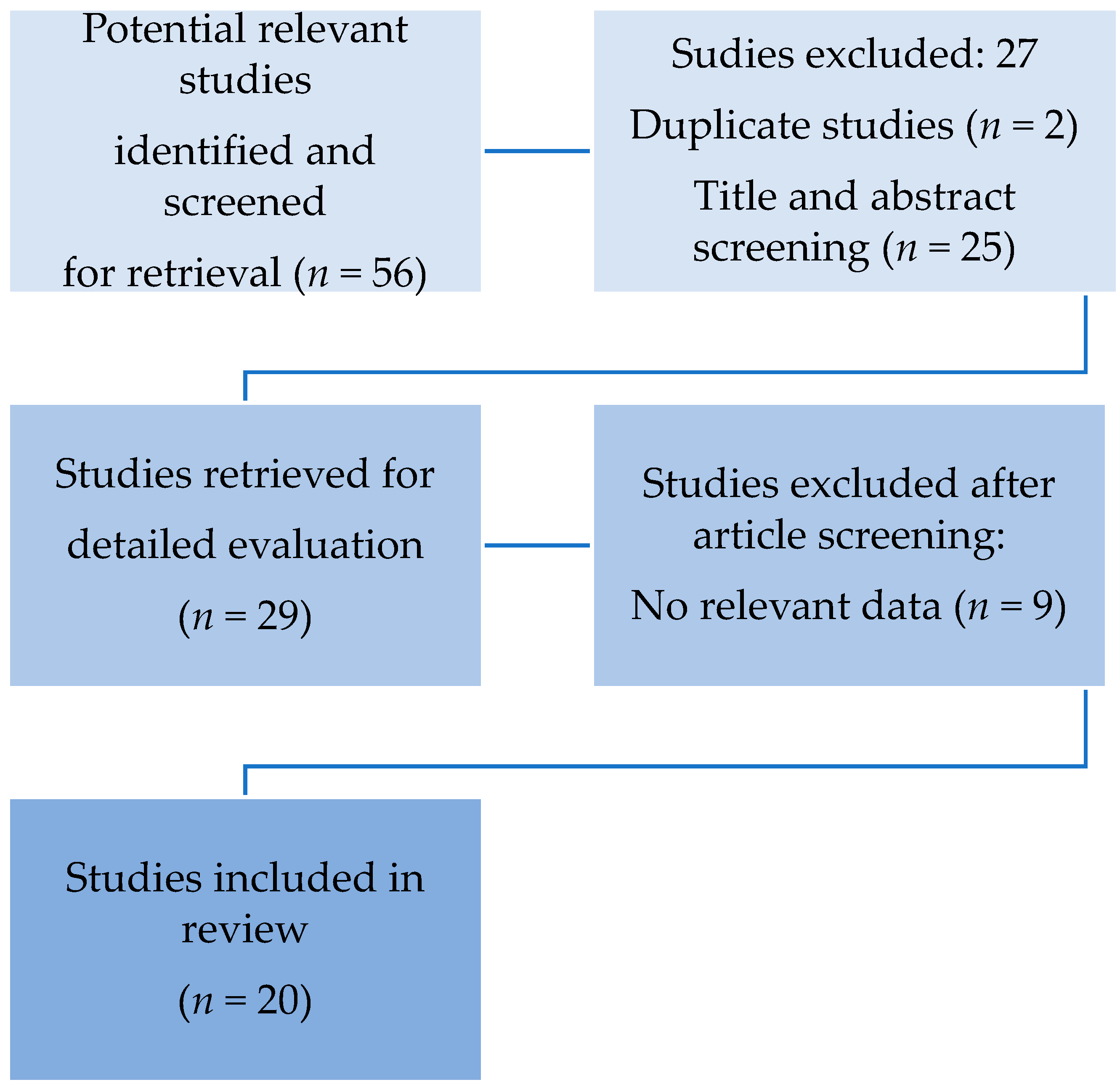
4. Methodology
5. Search Strategy
6. Results
7. Discussion
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nishiga, M.; Wang, D.W.; Han, Y.; Lewis, D.B.; Wu, J.C. COVID-19 and cardiovascular disease: From basic mechanisms to clinical perspectives. Nat. Rev. Cardiol. 2020, 17, 543–558. [Google Scholar] [CrossRef]
- Bonaventura, A.; Vecchié, A.; Dagna, L.; Martinod, K.; Dixon, D.L.; Van, T.B.W.; Dentali, F.; Montecucco, F.; Massberg, S.; Levi, M.; et al. Endothelial dysfunction and immunothrombosis as key pathogenic mechanisms in COVID. Nat. Rev. Immunol. 2021, 21, 319–329. [Google Scholar] [CrossRef]
- Stark, K.; Massberg, S. Interplay between inflammation and thrombosis in cardiovascular pathology. Nat. Rev. Cardiol. 2021, 6, 1–17. [Google Scholar] [CrossRef]
- Gu, S.X.; Tyagi, T.; Jain, K.; Gu, V.W.; Lee, S.H.; Hwa, J.M.; Kwan, J.M.; Krause, D.S.; Lee, A.I.; Halene, S.; et al. Thrombocytopathy and endotheliopathy: Crucial contributors to COVID-19 thromboinflammation. Nat. Rev. Cardiol. 2021, 18, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.A.H.; Bertozzi, C.R. The clinical impact of glycobiology: Targeting selectins, Siglecs and mammalian glycans. Nat. Rev. Drug. Discov. 2021, 20, 217–243. [Google Scholar] [CrossRef]
- Tvaroška, I.; Selvaraj, C.; Koča, J. Selectins-The Two Dr. Jekyll and Mr. Hyde Faces of Adhesion Molecules-A Review. Molecules 2020, 19, 2835. [Google Scholar] [CrossRef]
- Grobler, C.; Maphumulo, S.C.; Grobbelaar, L.M.; Bredenkamp, J.C.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Covid-19: The Rollercoaster of Fibrin(Ogen), D-Dimer, Von Willebrand Factor, P-Selectin and Their Interactions with Endothelial Cells, Platelets and Erythrocytes. Int. J. Mol. Sci. 2020, 21, 5168. [Google Scholar] [CrossRef]
- Polgar, J.; Matuskova, J.; Wagner, D.D. The P-selectin, tissue factor, coagulation triad. J. Thromb. Haemost. 2005, 3, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Furie, B. P-selectin and blood coagulation: It’s not only about inflammation any more. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 877–878. [Google Scholar] [CrossRef] [Green Version]
- Furie, B.; Furie, B.C. Role of platelet P-selectin and microparticle PSGL-1 in thrombus formation. Trends. Mol. Med. 2004, 10, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kato, G.J.; Piel, F.B.; Reid, C.D.; Gaston, M.H.; Ohene-Frempong, K.; Krishnamurti, L.; Smith, W.R.; Panepinto, J.A.; Weatherall, D.J.; Costa, F.F.; et al. Sickle cell disease. Nat. Rev. Dis. Primers. 2018, 15, 18010. [Google Scholar] [CrossRef] [Green Version]
- Karki, N.R.; Kutlar, A. P-Selectin Blockade in the Treatment of Painful Vaso-Occlusive Crises in Sickle Cell Disease: A Spotlight on Crizanlizumab. J. Pain. Res. 2021, 30, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wu, G.; Chang, S.; Bie, L. Plasma P-selectin level is associated with severity of coronary heart disease in Chinese Han population. J. Int. Med. Res. 2020, 48, 300060519896437. [Google Scholar] [CrossRef]
- Hoel, H.; Pettersen, E.M.; Høiseth, L.Ø.; Mathiesen, I.; Seternes, A.; Seljeflot, I.; Hisdal, J. Effects of intermittent negative pressure treatment on circulating vascular biomarkers in patients with intermittent claudication. Vasc. Med. 2021, 13, 1358863X211007933. [Google Scholar] [CrossRef]
- Shen, L.; Yang, T.; Xia, K.; Yan, Z.; Tan, J.; Li, L.; Qin, Y.; Shi, W. P-selectin (CD62P) and soluble TREM-like transcript-1 (sTLT-1) are associated with coronary artery disease: A case control study. Cardiovasc. Disord. 2020, 24, 387. [Google Scholar] [CrossRef]
- Barale, C.; Russo, I. Influence of Cardiometabolic Risk Factors on Platelet Function. Int. J. Mol. Sci. 2020, 17, 623. [Google Scholar] [CrossRef] [Green Version]
- Hally, K.E.; Parker, O.M.; Brunton-O’Sullivan, M.M.; Harding, S.A.; Larsen, P.D. Linking Neutrophil Extracellular Traps and Platelet Activation: A Composite Biomarker Score for Predicting Outcomes after Acute Myocardial Infarction. Thromb. Haemost. 2021, 13, 1728763. [Google Scholar] [CrossRef]
- Nguyen, S.D.; Korhonen, E.A.; Lorey, M.B.; Hakanpää, L.; Mäyränpää, M.I.; Kovanen, P.T.; Saharinen, P.; Alitalo, K.; Öörni, K. Lysophosphatidylcholine in phospholipase A2-modified LDL triggers secretion of angiopoietin 2. Atherosclerosis 2021, 327, 87–99. [Google Scholar] [CrossRef]
- Collado, A.; Domingo, E.; Marquesm, P.; Perello, E.; Martínez-Hervás, S.; Piqueras, L.; Ascaso, J.F.; Real, J.T.; Sanz, M.J. Oral Unsaturated Fat Load Impairs Postprandial Systemic Inflammation in Primary Hypercholesterolemia Patients. Front. Pharmacol. 2021, 20, 656244. [Google Scholar] [CrossRef]
- Chan, L.W.; Luo, X.P.; Ni, H.C.; Shi, H.M.; Liu, L.; Wen, Z.C.; Gu, X.Y.; Qiao, J.; Li, J. High levels of LDL-C combined with low levels of HDL-C further increase platelet activation in hypercholesterolemic patients. Braz. J. Med. Biol. Res. 2015, 48, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Appleton, J.P.; Richardson, C.; Dovlatova, N.; May, J.; Sprigg, N.; Heptinstall, S.; Bath, P.M. Remote platelet function testing using P-selectin expression in patients with recent cerebral ischaemia on clopidogrel. Stroke Vasc. Neurol. 2021, 6, 103–108. [Google Scholar] [CrossRef]
- Atkinson, C.; Zhu, H.; Qiao, F.; Varela, J.C.; Yu, J.; Song, H.; Kindy, M.S.; Tomlinson, S. Complement-dependent P-selectin expression and injury following ischemic stroke. J. Immunol. 2006, 15, 7266–7274. [Google Scholar] [CrossRef] [PubMed]
- Moin, A.S.M.; Al-Qaissi, A.; Sathyapalan, T.; Atkin, S.L.; Butler, A.E. Platelet Protein-Related Abnormalities in Response to Acute Hypoglycemia in Type 2 Diabetes. Front. Endocrinol. (Lausanne) 2021, 30, 651009. [Google Scholar] [CrossRef] [PubMed]
- Palella, E.; Cimino, R.; Pullano, S.A.; Fiorillo, A.S.; Gulletta, E.; Brunetti, A.; Foti, D.P.; Greco, M. Laboratory Parameters of Hemostasis, Adhesion Molecules, and Inflammation in Type 2 Diabetes Mellitus: Correlation with Glycemic Control. Int. J. Environ. Res. Public Health 2020, 17, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonopoulos, C.N.; Sfyroeras, G.S.; Kakisis, J.D.; Moulakakis, K.G.; Liapis, C.D. The role of soluble P selectin in the diagnosis of venous thromboembolism. Thromb. Res. 2014, 133, 17–24. [Google Scholar] [CrossRef]
- Ay, C.; Dunkler, D.; Marosi, C.; Chiriac, A.L.; Vormittag, R.; Simanek, R.; Quehenberger, P.; Zielinski, C.; Pabinger, I. Prediction of venous thromboembolism in cancer patients. Blood 2010, 116, 5377–5382. [Google Scholar] [CrossRef]
- Sánchez-López, V.; Gao, L.; Ferrer-Galván, M.; Arellano-Orden, E.; Elías-Hernández, T.; Jara-Palomares, L.; Asensio-Cruz, M.I.; Castro-Pérez, M.J.; Rodríguez-Martorell, F.J.; Lobo-Beristain, J.L.; et al. Differential biomarker profiles between unprovoked venous thromboembolism and cancer. Ann. Med. 2020, 52, 310–320. [Google Scholar] [CrossRef]
- Laursen, M.A.; Larsen, J.B.; Larsen, K.M.; Hvas, A.M. Platelet function in patients with septic shock. Thromb. Res. 2020, 185, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Widemann, A.; Pasero, C.; Arnaud, L.; Poullin, P.; Loundou, A.D.; Choukroun, G.; Sanderson, F.; Lacroix, R.; Sabatier, F.; Coppo, P.; et al. Circulating endothelial cells and progenitors as prognostic factors during autoimmune thrombotic thrombocytopenic purpura: Results of a prospective multicenter French study. J. Thromb. Haemost. 2014, 12, 1601–1609. [Google Scholar] [CrossRef]
- Campello, E.; Radu, C.M.; Duner, E.; Lombardi, A.M.; Spiezia, L.; Bendo, R.; Ferrari, S.; Simioni, P.; Fabris, F. Activated Platelet-Derived and Leukocyte-Derived Circulating Microparticles and the Risk of Thrombosis in Heparin-Induced Thrombocytopenia: A Role for PF4-Bearing Microparticles? Cytometry B Clin. Cytom. 2018, 94, 334–341. [Google Scholar] [CrossRef]
- Fabricius, H.Å.; Starzonek, S.; Lange, T. The Role of Platelet Cell Surface P-Selectin for the Direct Platelet-Tumor Cell Contact During Metastasis Formation in Human Tumors. Front. Oncol. 2021, 15, 642761. [Google Scholar] [CrossRef]
- Schlesinger, M. Role of platelets and platelet receptors in cancer metastasis. J. Hematol. Oncol. 2018, 11, 125. [Google Scholar] [CrossRef]
- Kappelmayer, J.; Nagy, B., Jr. The Interaction of Selectins and PSGL-1 as a Key Component in Thrombus Formation and Cancer Progression. Biomed. Res. Int. 2017, 2017, 6138145. [Google Scholar] [CrossRef]
- Qi, C.; Wei, B.; Zhou, W. P-selectin-mediated platelet adhesion promotes tumor growth. Oncotarget 2015, 6, 6584–6596. [Google Scholar] [CrossRef] [Green Version]
- Tavares, L.P.; Teixeira, M.M.; Garcia, C.C. The inflammatory response triggered by Influenza virus: A two edged sword. Inflamm. Res. 2017, 66, 283–302. [Google Scholar] [CrossRef]
- Finsterbusch, M.; Schrottmaier, W.C.; Kral-Pointner, J.B.; Salzmann, M.; Assinger, A. Measuring and interpreting platelet-leukocyte aggregates. Platelets 2018, 29, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Nkambule, B.B.; Mxinwa, V.; Mkandla, Z.; Mutize, T.; Mokgalaboni, K.; Nyambuya, T.M.; Dludla, P.V. Platelet activation in adult HIV-infected patients on antiretroviral therapy: A systematic review and meta-analysis. BMC. Med. 2020, 18, 357. [Google Scholar] [CrossRef] [PubMed]
- Agrati, C.; Mazzotta, V.; Pinnetti, C.; Biava, G.; Bibas, M. Venous thromboembolism in people living with HIV infection (PWH). Transl. Res. 2021, 227, 89–99. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, J.A.; Dunne, E.; Gurwith, M.; Lambert, J.S.; Sheehan, G.J.; Feeney, E.R.; Pozniak, A.; Reiss, P.; Kenny, D.; Mallon, P. The effect of initiation of antiretroviral therapy on monocyte, endothelial and platelet function in HIV-1 infection. HIV Med. 2015, 16, 608–619. [Google Scholar] [CrossRef]
- O’Brien, M.P.; Hunt, P.W.; Kitch, D.W.; Klingman, K.; Stein, J.H.; Funderburg, N.T.; Berger, J.S.; Tebas, P.; Clagett, B.; Moisi, D.; et al. A Randomized Placebo Controlled Trial of Aspirin Effects on Immune Activation in Chronically Human Immunodeficiency Virus-Infected Adults on Virologically Suppressive Antiretroviral Therapy. Open. Forum. Infect. Dis. 2017, 4, 278. [Google Scholar] [CrossRef] [Green Version]
- Bibas, M.; Biavam, G.; Antinori, A. HIV-Associated Venous Thromboembolism. Infect. Dis. 2011, 3, e2011030. [Google Scholar] [CrossRef]
- Johansson, M.W.; Grill, B.M.; Barretto, K.T.; Favour, M.C.; Schira, H.M.; Swanson, C.M.; Lee, K.E.; Sorkness, R.L.; Mosher, D.F.; Denlinger, L.C.; et al. Plasma P-Selectin Is Inversely Associated with Lung Function and Corticosteroid Responsiveness in Asthma. Int. Arch. Allergy Immunol. 2020, 181, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Horváth, P.; Lázár, Z.; Gálffy, G.; Puskás, R.; Kunos, L.; Losonczy, G.; Mészáros, M.; Tárnoki, Á.D.; Tárnoki, D.L.; Bikov, A. Circulating P-Selectin Glycoprotein Ligand 1 and P-Selectin Levels in Obstructive Sleep Apnea Patients. Lung 2020, 198, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Xu, Z.; Liu, T.; Li, Y. Soluble P-selectin levels in patients with obstructive sleep apnea: A systematic review and meta-analysis. Eur. Arch. Otorhinolaryngol. 2021. [Google Scholar] [CrossRef]
- Goshua, G.; Pine, A.B.; Meizlish, M.L.; Chang, C.H.; Zhang, H.; Bahel, P.; Baluha, A.; Bar, N.; Bona, R.D.; Burns, A.J.; et al. Endotheliopathy in COVID-19-associated coagulopathy: Evidence from a single-centre, cross-sectional study. Lancet Haematol. 2020, 7, 575–582. [Google Scholar] [CrossRef]
- Hottz, E.D.; Azevedo-Quintanilha, I.G.; Palhinha, L.; Teixeira, L.; Barreto, E.A.; Pão, C.R.R.; Righy, C.; Franco, S.; Souza, T.M.L.; Kurtz, P.; et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood 2020, 136, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Campo, G.; Contoli, M.; Fogagnolo, A.; Vieceli, D.S.F.; Zucchetti, O.; Ronzoni, L.; Verri, M.; Fortini, F.; Pavasini, R.; Morandi, L.; et al. Over time relationship between platelet reactivity, myocardial injury and mortality in patients with SARS-CoV-2-associated respiratory failure. Platelets 2020, 3, 1–8. [Google Scholar] [CrossRef]
- Vassiliou, A.G.; Keskinidou, C.; Jahaj, E.; Gallos, P.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E. ICU Admission Levels of Endothelial Biomarkers as Predictors of Mortality in Critically Ill COVID-19 Patients. Cells 2021, 10, 186. [Google Scholar] [CrossRef]
- Barrett, T.J.; Lee, A.H.; Xia, Y.; Lin, L.H.; Black, M.; Cotzia, P.; Hochman, J.; Berger, J.S. Platelet and Vascular Biomarkers Associate With Thrombosis and Death in Coronavirus Disease. Circ. Res. 2020, 127, 945–947. [Google Scholar] [CrossRef]
- Agrati, C.; Bordoni, V.; Sacchi, A.; Petrosillo, N.; Nicastri, E.; Del, N.F.; D’Offizi, G.; Palmieri, F.; Marchioni, L.; Capobianchi, M.R.; et al. Elevated P-Selectin in Severe Covid-19: Considerations for Therapeutic Options. Mediterr. J. Hematol. Infect. Dis. 2021, 13, e2021016. [Google Scholar] [CrossRef] [PubMed]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet gene expression and function in patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, J.; Fang, Y.; Lu, S.; Wu, J.; Zheng, X.; Deng, F. SARS-CoV-2 interacts with platelets and megakaryocytes via ACE2-independent mechanism. J. Hematol. Oncol. 2021, 14, 72. [Google Scholar] [CrossRef]
- Petito, E.; Falcinelli, E.; Paliani, U.; Cesari, E.; Vaudo, G.; Sebastiano, M.; Cerotto, V.; Guglielmini, G.; Gori, F.; Malvestiti, M.; et al. Association of Neutrophil Activation, More Than Platelet Activation, With Thrombotic Complications in Coronavirus Disease. J. Infect. Dis. 2021, 223, 933–944. [Google Scholar] [CrossRef]
- Comer, S.P.; Cullivan, S.; Szklanna, P.B.; Weiss, L.; Cullen, S.; Kelliher, S.; Smolenski, A.; Murphy, C.; Altaie, H.; Curran, J.; et al. COVID-19 induces a hyperactive phenotype in circulating platelets. PLoS. Biol. 2021, 19, e3001109. [Google Scholar] [CrossRef]
- Fraser, D.D.; Patterson, E.K.; Slessarev, M.; Gill, S.E.; Martin, C.; Daley, M.; Miller, M.R.; Patel, M.A.; Dos Santos, C.C.; Bosma, K.J.; et al. Endothelial Injury and Glycocalyx Degradation in Critically Ill Coronavirus Disease 2019 Patients: Implications for Microvascular Platelet Aggregation. Crit. Care Explor. 2020, 2, e0194. [Google Scholar] [CrossRef]
- Karsli, E.; Sabirli, R.; Altintas, E.; Canacik, O.; Sabirli, G.T.; Kaymaz, B.; Kurt, Ö.; Koseler, A. Soluble P-selectin as a potential diagnostic and prognostic biomarker for COVID-19 disease: A case-control study. Life Sci. 2021, 277, 119634. [Google Scholar] [CrossRef]
- Taus, F.; Salvagno, G.; Canè, S.; Fava, C.; Mazzaferri, F.; Carrara, E.; Petrova, V.; Barouni, R.M.; Dima, F.; Dalbeni, A.; et al. Platelets Promote Thromboinflammation in SARS-CoV-2 Pneumonia. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2975–2989. [Google Scholar] [CrossRef]
- Chao, Y.; Rebetz, J.; Bläckberg, A.; Hovold, G.; Sunnerhagen, T.; Rasmussen, M.; Semple, J.W.; Shannon, O. Distinct phenotypes of platelet, monocyte, and neutrophil activation occur during the acute and convalescent phase of COVID. Platelets 2021, 17, 1–11. [Google Scholar] [CrossRef]
- Canzano, P.; Brambilla, M.; Porro, B.; Cosentino, N.; Tortorici, E.; Vicini, S.; Poggio, P.; Cascella, A.; Pengo, M.F.; Veglia, F.; et al. Platelet and Endothelial Activation as Potential Mechanisms Behind the Thrombotic Complications of COVID-19 Patients. JACC Basic. Transl. Sci. 2021, 6, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Bongiovanni, D.; Klug, M.; Lazareva, O.; Weidlich, S.; Biasi, M.; Ursu, S.; Warth, S.; Buske, C.; Lukas, M.; Spinner, C.D.; et al. SARS-CoV-2 infection is associated with a pro-thrombotic platelet phenotype. Cell Death Dis. 2021, 12, 50. [Google Scholar] [CrossRef]
- Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID. Int. J. Mol. Sci. 2020, 21, 8234. [Google Scholar] [CrossRef]
- Clark, C.C.; Jukema, B.N.; Barendrecht, A.D.; Spanjaard, J.S.; Jorritsma, N.K.N.; Smits, S.; de Maat, S.; Seinen, C.W.; Verhoef, S.; Parr, N.M.J.; et al. Thrombotic Events in COVID-19 Are Associated With a Lower Use of Prophylactic Anticoagulation Before Hospitalization and Followed by Decreases in Platelet Reactivity. Front. Med. (Lausanne) 2021, 8, 650129. [Google Scholar] [CrossRef]
- Bertolin, A.J.; Dalçóquio, T.F.; Salsoso, R.; Furtado, R.H.D.M.; Kalil-Filho, R.; Hajjar, L.A.; Siciliano, R.F.; Kallás, E.G.; Baracioli, L.M.; Lima, F.G.; et al. Platelet Reactivity and Coagulation Markers in Patients with COVID-19. Adv. Ther. 2021, 38, 3911–3923. [Google Scholar] [CrossRef]
- Spadaro, S.; Fogagnolo, A.; Campo, G.; Zucchetti, O.; Verri, M.; Ottaviani, I.; Tunstall, T.; Grasso, S.; Scaramuzzo, V.; Murgolo, F.; et al. Markers of endothelial and epithelial pulmonary injury in mechanically ventilated COVID-19 ICU patients. Crit. Care 2021, 25, 74. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Liu, S.; Sun, J.; Chen, Z.; Jiang, M.; Zhang, Q.; Wei, Y.; Wang, X.; Huang, Y.Y.; et al. Potential therapeutic effects of dipyridamole in the severely ill patients with COVID-19. Acta Pharm. Sin. B 2020, 10, 1205–1215. [Google Scholar] [CrossRef]
- Antithrombotic Trialists’ (ATT) Collaboration; Baigent, C.; Blackwell, L.; Collins, R.; Emberson, J.; Godwin, J.; Peto, R.; Buring, J.; Hennekens, C.; Kearney, P.; et al. Aspirin in the primary and secondary prevention of vascular disease: Collaborative meta-analysis of individual participant data from randomised trials. Lancet 2009, 373, 1849–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, J.H.; Khanna, A.K.; Kethireddy, S.; Yamane, D.; Levine, A.; Jackson, A.M.; McCurdy, M.T.; Tabatabai, A.; Kumar, G.; Park, P.; et al. Aspirin Use Is Associated With Decreased Mechanical Ventilation, Intensive Care Unit Admission, and In-Hospital Mortality in Hospitalized Patients With Coronavirus Disease. Anesth. Analg. 2021, 132, 930–941. [Google Scholar] [CrossRef] [PubMed]
- RECOVERY Collaborative Group; Peter, W.H.; Guilherme, P.A.; Natalie, S.; Jonathan, R.E.; Mark, C.; Enti, S.; Leon, P.; Nigel, J.B.; Simon, T.; et al. Aspirin in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label platform trial. Health Sci. 2021, 8, 21258132. [Google Scholar] [CrossRef]
- Patrick, R.L.; Ewan, C.G.; Jeffrey, S.B.; Matthew, D.N.; Bryan, J.M.; Jose, C.N.; Michelle, N.G.; Marc, C.; Robert, S.R.; Harmony, R.R. Therapeutic Anticoagulation in Non-Critically Ill Patients with Covid-19. Health Sci. 2021, 5, 21256846. [Google Scholar] [CrossRef]
- Sadeghipour, P.; Talasaz, A.H.; Rashidi, F.; Sharif-Kashani, B.; Beigmohammadi, M.T.; Farrokhpour, M.; Sezavar, S.H.; Payandemehr, P.; Dabbagh, A.; Moghadam, K.G.; et al. Effect of Intermediate-Dose vs Standard-Dose Prophylactic Anticoagulation on Thrombotic Events, Extracorporeal Membrane Oxygenation Treatment, or Mortality Among Patients With COVID-19 Admitted to the Intensive Care Unit: The INSPIRATION Randomized Clinical Trial. JAMA 2021, 325, 1620–1630. [Google Scholar] [PubMed]
- Blair, H.A. Crizanlizumab: First Approval. Drugs 2020, 80, 79–84. [Google Scholar] [CrossRef]


{kind=link}
| AUTHOR | COVID-19 pts | Age Median (Range) | Gender M/F | Moderate COVID-19+ (n) | Severe non- ICU COVID-19+ (n) | Severe ICU COVID-19+ (n) | Main Findings |
|---|---|---|---|---|---|---|---|
| Goshua [45] | 68 | 62 (20–93) | 27/41 | - | higher than HC (n = 20) | higher than non-ICU (n = 48) | Association with critical illness and death |
| Hottz [46] | 41 | 57 (47–64) | 19/22 | - | - | higher than controls (n = 35) | Association with critical illness and death |
| Campo [47] | 54 | 65 (57–73) | 40/14 | - | higher than HC (n = 17) | higher than non-ICU (n = 37) | Association with critical illness and death |
| Vassiliou [48] | 38 | 65(51–78) | 31/7 | - | higher than non-ICU (n = 38) | Association with critical illness and death | |
| Barrett [49] | 100 | 65 | 61/39 | higher than HC (n = 100) | Association with critical illness and death | ||
| Agrati [50] | 46 | 67 (50–80) | 34/12 | - | higher than HC (n = 19) | no differences with non-ICU (n = 27) | Higher P-Sel regardless ICU admission |
| Manne [51] | 41 | 55 (33–77) | 19/22 | - | higher than HC (n = 24) | no differences with non-ICU (n = 17) | Higher P-Sel regardless ICU admission |
| Shen [52] | 62 | 66 (60–71) | 32/30 | - | higher than HC (n = 37) | higher than non-ICU (n = 25) | Higher values in COVID-19+ ICU patients. |
| Petito [53] | 36 | 70.6 (36–60) | 20/16 | - | higher than HC (n = 17) | higher than non-ICU (n = 19) | Higher values in COVID-19+ ICU patients |
| Comer [54] | 54 | 63 (47–84) | 34/20 | - | higher than HC (n = 20) | higher than non-ICU (n = 34) | Higher values in COVID-19+ ICU patients |
| Fraser [55] | 10 | 61 (54.8–67) | 3/7 | - | - | higher than HC (n = 10) | Higher values in COVID-19+ ICU patients |
| Karsli [56] | 80 | - | - | higher than HC (n = 50) | higher than non-ICU (n = 35) | Higher values in COVID-19+ ICU patients | |
| Taus [57] | 37 | 61.8 (47–94) | 18/19 | - | higher than HC (n = 37) | - | P-Sel higher than controls |
| Chao [58] | 15 | 71 (53–80) | 5/10 | higher than HC (n = 5) | - | - | P-Sel higher than controls |
| Canzano [59] | 46 | 72 (58–84) | 28/18 | - | higher than HC (n = 46) | - | P-Sel higher than controls |
| Bongiovanni [60] | 8 | 51.4 (39–64) | 5/3 | - | higher than HC (n = 8) | - | P-Sel higher than controls |
| Venter [61] | 30 | 53.1 (38–69) | - | lower than HC (n = 30) | - | - | Study reporting a lower P-Sel level in COVID-19+ |
| Clark [62] | 79 | 67 (54–75) | 43/34 | - | Similar to Controls (n = 79) | - | Study reporting a similar P-Sel level In COVID-19+ and Controls |
| Bertolin [63] | 60 | 52 (37–68) | 31/29 | Similar to Controls (n = 60) | Study reporting a similar P-Sel level In COVID-19+ and Controls | ||
| Spadaro [64] | 31 | 67 (55–75) | 26/5 | - | - | 31 lower value than COVID-19- ARDS (n = 31) | P-Sel lower than ARDS controls, but higher than normal value |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrati, C.; Sacchi, A.; Tartaglia, E.; Vergori, A.; Gagliardini, R.; Scarabello, A.; Bibas, M. The Role of P-Selectin in COVID-19 Coagulopathy: An Updated Review. Int. J. Mol. Sci. 2021, 22, 7942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157942
Agrati C, Sacchi A, Tartaglia E, Vergori A, Gagliardini R, Scarabello A, Bibas M. The Role of P-Selectin in COVID-19 Coagulopathy: An Updated Review. International Journal of Molecular Sciences. 2021; 22(15):7942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157942
Chicago/Turabian StyleAgrati, Chiara, Alessandra Sacchi, Eleonora Tartaglia, Alessandra Vergori, Roberta Gagliardini, Alessandra Scarabello, and Michele Bibas. 2021. "The Role of P-Selectin in COVID-19 Coagulopathy: An Updated Review" International Journal of Molecular Sciences 22, no. 15: 7942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157942