
The Role of Mitochondria-Derived Peptides in Cardiovascular Diseases and Their Potential as Therapeutic Targets


Abstract
:1. Introduction
2. MDP General Description
2.1. Mitochondrial Genome Overview
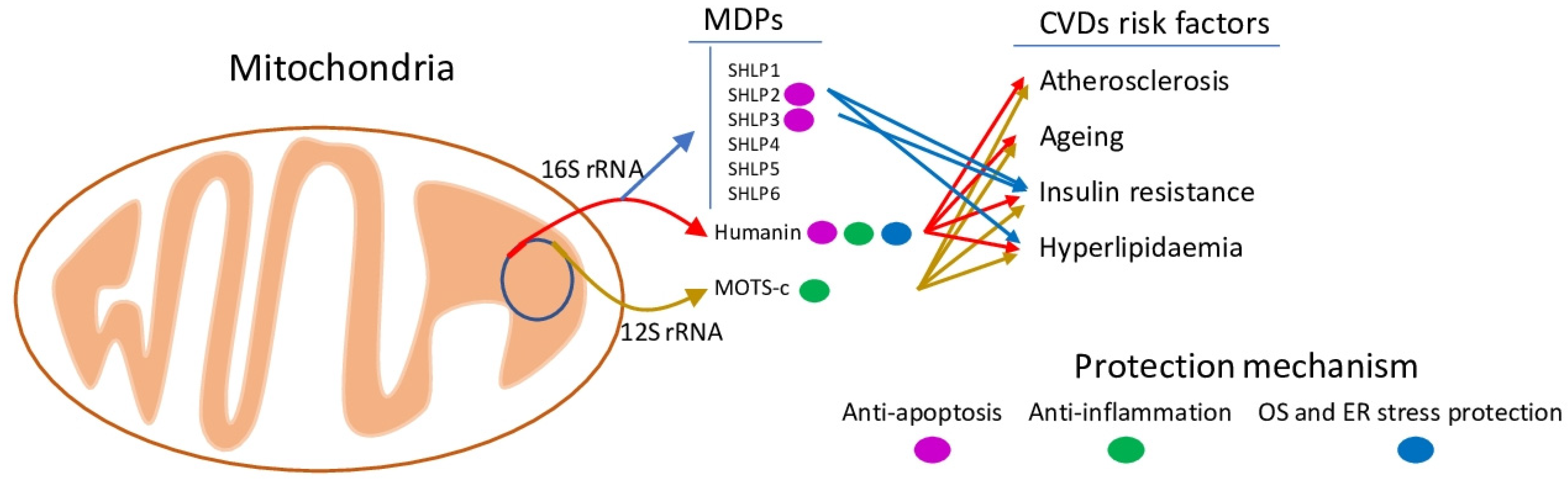
2.2. MDPs Generation and Functions
2.3. Nuclear-Encoded sORF Microproteins That Act on Mitochondria
2.4. Connection to Cardiovascular Risk Factors
2.5. Mechanisms of Action
3. Association between MDPs and CVDs
4. Therapeutic Application of Modified MDPs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABCA1 | ATP-binding cassette transporter A1 |
| ABCG1 | ATP-binding cassette sub-family G member 1 |
| ACC | acetyl CoA carboxylase |
| ADNF | activity-dependent neurotrophic factor |
| AhR | aryl hydrocarbon receptor |
| AICAR | 5-aminoimidazole-4-carboxamide ribonucleotide |
| AKT | protein kinase B |
| AMD | age-related macular degeneration |
| AMI | acute myocardial ischemia |
| AMPK | AMP-activated protein kinase |
| ARPE-19 | immortalized retinal pigment epithelial |
| AT-1 | angiotensin II type 1 |
| av-scO1 | avian strongly conserved sORFs 1 |
| BAX | BCL2 Associated X, apoptosis regulator |
| BCL2L11 | BCL2-like 11 apoptosis facilitator |
| Bid | BH3-interacting domain death agonist |
| BV-shHN | Baculovirus-encoded short-hairpin RNA |
| CAD | coronary artery disease |
| CD36 | fatty acid translocase |
| CLCC1 | chloride channel CLIC like 1 |
| CMD | coronary microvascular dysfunction |
| CNTF | ciliary neuronotrophic factor |
| COX2 | cyclooxygenase-2 |
| CVD | cardiovascular disease |
| Drp1 | dynamin-related protein 1 |
| ED | endothelial dysfunction |
| ELP | elastin-like polypeptides |
| eNOS | endothelial nitric oxide synthase |
| Erk1/2 | mitogen-activated protein kinase 1 and 3 |
| ET-1 | endothelin-1 |
| ET-B | endothelin B |
| FAS | fatty acid synthase |
| FGF2 | fibroblast growth factor 2 |
| FGF21 | fibroblast growth factor 21 |
| FPRL1 | formyl peptide receptor-like 1 |
| GLUT4 | glucose transporter type 4, insulin-responsive |
| GSK3B | glycogen synthase kinase 3β |
| HN | Humanin |
| HSP90 | heat shock protein 90 |
| ICAM-1 | intercellular adhesion molecule |
| IGFBP-3 | insulin-like growth factor-binding protein 3 |
| IL-6 | interleukin |
| IL27RA | interleukin 27 receptor subunit alpha |
| IL6ST | interleukin 6 cytokine family signal transducer |
| iNOS | nitric oxide synthase 2A inducible, hepatocytes |
| IR | insulin resistance |
| IRS-1 | insulin receptor substrate 1 |
| JNK | Janus kinase |
| KLF2 | Krüppel-like factor 2 |
| LOX-1 | lectin-type oxidized LDL receptor 1 |
| MAPK | mitogen-activated protein kinase |
| MAPK14 | mitogen-activated protein kinase 14 or p38 MAP kinase |
| MAPK3 | mitogen-activated protein kinase 3 |
| MCP-1 | monocyte chemotactic protein-1 |
| MDPs | mitochondria-derived peptides |
| MIEF1-MP | mitochondrial elongation factor 1 microprotein |
| MIM | mitochondrial inner membrane |
| MMP2 | matrix metallopeptidase 2 |
| MOM | mitochondrial outer membrane |
| MOTS-c | mitochondrial ORF of the 12S rDNA type-c |
| MOXI | micropeptide regulator of β-oxidation |
| MPM | micropeptide in mitochondria |
| MRI | myocardial reperfusion injury |
| mTOR | mechanistic target of rapamycin |
| mtTFA | mitochondrial transcription factor A |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NFE2L2 | nuclear factor erythroid 2-related factor 2 |
| NLRP3 | NOD-, LRR- and pyrin domain-containing protein 3 |
| NOX2 | superoxide-generating NADPH oxidase heavy chain subunit |
| ORF | open reading frames |
| OS | oxidative stress |
| Ox-LDL | oxidized low-density lipoprotein |
| OXPHOS | oxidative phosphorylation |
| PCA | percutaneous coronary angioplasty |
| PGC1α | peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PI3K | phosphatidylinositol 3-kinase |
| ROS | reactive oxygen species |
| RPE | retinal pigment epithelium |
| SASPs | senescence-associated secretory phenotypes |
| SCD1 | acyl-CoA desaturase |
| SCML1 | sex comb on midleg-like 1 |
| SHLP | small humanin-like peptide, 1 to 6 |
| SIRT1 | Sirtuin 1 |
| SNPs | single nucleotide polymorphisms |
| SOD1 | superoxide dismutase 1 |
| SREBP1 | sterol regulatory element-binding transcription factor 1 |
| STAT3 | signal transducer and activator of transcriptional 3 |
| TGFB1 | transforming growth factor-beta 1 |
| TNBC | triple-negative breast cancer |
| TNF-α | tumour necrosis factor-alpha |
| TxNIP | thioredoxin interacting protein |
| UPR | unfolded protein response |
| VCAM-1 | vascular cell adhesion molecule 1 |
| VEGF | vascular endothelial growth factor |
References
- Hashimoto, Y.; Niikura, T.; Ito, Y.; Sudo, H.; Hata, M.; Arakawa, E.; Abe, Y.; Kita, Y.; Nishimoto, I. Detailed Characterization of Neuroprotection by a Rescue Factor Humanin against Various Alzheimer’s Disease-Relevant Insults. J. Neurosci. 2001, 21, 9235–9245. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Niikura, T.; Tajima, H.; Yasukawa, T.; Sudo, H.; Ito, Y.; Kita, Y.; Kawasumi, M.; Kouyama, K.; Doyu, M.; et al. A Rescue Factor Abolishing Neuronal Cell Death by a Wide Spectrum of Familial Alzheimer’s Disease Genes and A. Proc. Natl. Acad. Sci. USA 2001, 98, 6336–6341. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.-J.; Mehta, H.; Hevener, A.L.; de Cabo, R.; et al. The Mitochondrial-Derived Peptide MOTS-c Promotes Metabolic Homeostasis and Reduces Obesity and Insulin Resistance. Cell Metab. 2015, 21, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobb, L.J.; Lee, C.; Xiao, J.; Yen, K.; Wong, R.G.; Nakamura, H.K.; Mehta, H.H.; Gao, Q.; Ashur, C.; Huffman, D.M.; et al. Naturally Occurring Mitochondrial-Derived Peptides Are Age-Dependent Regulators of Apoptosis, Insulin Sensitivity, and Inflammatory Markers. Aging 2016, 8, 796–809. [Google Scholar] [CrossRef] [Green Version]
- Merry, T.L.; Chan, A.; Woodhead, J.S.T.; Reynolds, J.C.; Kumagai, H.; Kim, S.-J.; Lee, C. Mitochondrial-Derived Peptides in Energy Metabolism. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E659–E666. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Kannan, R. Mechanisms of Protection of Retinal Pigment Epithelial Cells from Oxidant Injury by Humanin and Other Mitochondrial-Derived Peptides: Implications for Age-Related Macular Degeneration. Redox Biol. 2020, 37, 101663. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.F.; Asad, A.S.; Nicola Candia, A.J.; Gottardo, M.F.; Moreno Ayala, M.A.; Theas, M.S.; Seilicovich, A.; Candolfi, M. Mitochondrial-Derived Peptide Humanin as Therapeutic Target in Cancer and Degenerative Diseases. Expert Opin. Ther. Targets 2019, 23, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, A.R.; Larrick, J.W. Mitochondrial-Derived Peptides Exacerbate Senescence. Rejuvenation Res. 2018, 21, 369–373. [Google Scholar] [CrossRef]
- Rao, M.; Wu, Z.; Wen, Y.; Wang, R.; Zhao, S.; Tang, L. Humanin Levels in Human Seminal Plasma and Spermatozoa Are Related to Sperm Quality. Andrology 2019, 7, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Paris, V.; Walters, K.A. Humanin: A Potential Treatment for PCOS? Endocrinology 2021, 162, bqab085. [Google Scholar] [CrossRef] [PubMed]
- Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The Similarities between Human Mitochondria and Bacteria in the Context of Structure, Genome, and Base Excision Repair System. Molecules 2020, 25, 2857. [Google Scholar] [CrossRef] [PubMed]
- Vizioli, M.G.; Liu, T.; Miller, K.N.; Robertson, N.A.; Gilroy, K.; Lagnado, A.B.; Perez-Garcia, A.; Kiourtis, C.; Dasgupta, N.; Lei, X.; et al. Mitochondria-to-Nucleus Retrograde Signaling Drives Formation of Cytoplasmic Chromatin and Inflammation in Senescence. Genes Dev. 2020, 34, 428–445. [Google Scholar] [CrossRef]
- Gong, Z.; Tasset, I.; Diaz, A.; Anguiano, J.; Tas, E.; Cui, L.; Kuliawat, R.; Liu, H.; Kühn, B.; Cuervo, A.M.; et al. Humanin Is an Endogenous Activator of Chaperone-Mediated Autophagy. J. Cell Biol. 2018, 217, 635–647. [Google Scholar] [CrossRef]
- Gong, Z.; Tasset, I. Humanin Enhances the Cellular Response to Stress by Activation of Chaperone-Mediated Autophagy. Oncotarget 2018, 9, 10832–10833. [Google Scholar] [CrossRef]
- Qin, Q.; Jin, J.; He, F.; Zheng, Y.; Li, T.; Zhang, Y.; He, J. Humanin Promotes Mitochondrial Biogenesis in Pancreatic MIN6 β-Cells. Biochem. Biophys. Res. Commun. 2018, 497, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Logan, I.S. Pseudogenization of the Humanin gene is common in the mitochondrial DNA of many vertebrates. Zool. Res. 2017, 38, 198–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Choi, J.-W.; Namkung, J. Expression Profile of Mouse Gm20594, Nuclear-Encoded Humanin-Like Gene. J. Lifestyle Med. 2021, 11, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mortz, M.; Dégletagne, C.; Romestaing, C.; Duchamp, C. Comparative Genomic Analysis Identifies Small Open Reading Frames (SORFs) with Peptide-Encoding Features in Avian 16S RDNA. Genomics 2020, 112, 1120–1127. [Google Scholar] [CrossRef]
- Capt, C.; Passamonti, M.; Breton, S. The Human Mitochondrial Genome May Code for More than 13 Proteins. Mitochondrial DNA Part A 2016, 27, 3098–3101. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ren, K.; Jiang, T.; Zhao, G.-J. MOTS-c Attenuates Endothelial Dysfunction via Suppressing the MAPK/NF-ΚB Pathway. Int. J. Cardiol. 2018, 268, 40. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Son, J.M.; Benayoun, B.A.; Lee, C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress. Cell Metab. 2018, 28, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, J.C.; Lai, R.W.; Woodhead, J.S.T.; Joly, J.H.; Mitchell, C.J.; Cameron-Smith, D.; Lu, R.; Cohen, P.; Graham, N.A.; Benayoun, B.A.; et al. MOTS-c Is an Exercise-Induced Mitochondrial-Encoded Regulator of Age-Dependent Physical Decline and Muscle Homeostasis. Nat. Commun. 2021, 12, 470. [Google Scholar] [CrossRef]
- Okada, A.K.; Teranishi, K.; Lobo, F.; Isas, J.M.; Xiao, J.; Yen, K.; Cohen, P.; Langen, R. The Mitochondrial-Derived Peptides, HumaninS14G and Small Humanin-like Peptide 2, Exhibit Chaperone-like Activity. Sci. Rep. 2017, 7, 7802. [Google Scholar] [CrossRef] [Green Version]
- Nashine, S.; Cohen, P.; Nesburn, A.B.; Kuppermann, B.D.; Kenney, M.C. Characterizing the Protective Effects of SHLP2, a Mitochondrial-Derived Peptide, in Macular Degeneration. Sci. Rep. 2018, 8, 15175. [Google Scholar] [CrossRef] [Green Version]
- Chu, Q.; Martinez, T.F.; Novak, S.W.; Donaldson, C.J.; Tan, D.; Vaughan, J.M.; Chang, T.; Diedrich, J.K.; Andrade, L.; Kim, A.; et al. Regulation of the ER Stress Response by a Mitochondrial Microprotein. Nat. Commun. 2019, 10, 4883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathore, A.; Chu, Q.; Tan, D.; Martinez, T.F.; Donaldson, C.J.; Diedrich, J.K.; Yates, J.R.; Saghatelian, A. MIEF1 Microprotein Regulates Mitochondrial Translation. Biochemistry 2018, 57, 5564–5575. [Google Scholar] [CrossRef]
- Stein, C.S.; Jadiya, P.; Zhang, X.; McLendon, J.M.; Abouassaly, G.M.; Witmer, N.H.; Anderson, E.J.; Elrod, J.W.; Boudreau, R.L. Mitoregulin: A LncRNA-Encoded Microprotein That Supports Mitochondrial Supercomplexes and Respiratory Efficiency. Cell Rep. 2018, 23, 3710–3720.e8. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Xiao, M.-H.; Chen, H.-X.; Meng, Y.; Zhao, N.; Yang, L.; Tang, H.; Wang, J.-L.; Liu, X.; Zhu, Y.; et al. A Novel Mitochondrial Micropeptide MPM Enhances Mitochondrial Respiratory Activity and Promotes Myogenic Differentiation. Cell Death Dis. 2019, 10, 528. [Google Scholar] [CrossRef] [PubMed]
- Makarewich, C.A.; Baskin, K.K.; Munir, A.Z.; Bezprozvannaya, S.; Sharma, G.; Khemtong, C.; Shah, A.M.; McAnally, J.R.; Malloy, C.R.; Szweda, L.I.; et al. MOXI Is a Mitochondrial Micropeptide That Enhances Fatty Acid β-Oxidation. Cell Rep. 2018, 23, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Karu, I.; Tahepold, P.; Ruusalepp, A.; Reimann, E.; Koks, S.; Starkopf, J. Exposure to Sixty Minutes of Hyperoxia Upregulates Myocardial Humanins in Patients with Coronary Artery Disease—A Pilot Study. J. Physiol Pharm. 2015, 66, 899–906. [Google Scholar]
- Eltermaa, M.; Jakobson, M.; Utt, M.; Kõks, S.; Mägi, R.; Starkopf, J. Genetic Variants in Humanin Nuclear Isoform Gene Regions Show No Association with Coronary Artery Disease. BMC Res. Notes 2019, 12, 759. [Google Scholar] [CrossRef]
- Zhu, Y.; Xian, X.; Wang, Z.; Bi, Y.; Chen, Q.; Han, X.; Tang, D.; Chen, R. Research Progress on the Relationship between Atherosclerosis and Inflammation. Biomolecules 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frambach, S.J.C.M.; de Haas, R.; Smeitink, J.A.M.; Rongen, G.A.; Russel, F.G.M.; Schirris, T.J.J. Brothers in Arms: ABCA1- and ABCG1-Mediated Cholesterol Efflux as Promising Targets in Cardiovascular Disease Treatment. Pharm. Rev. 2020, 72, 152–190. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, S.; Liu, Z.; Cao, Y.; Wang, F.; Wang, J.; Liu, C.; Xie, Y.; Xie, Y.; Zhang, Y. Gly[14]-Humanin Inhibits Ox-LDL Uptake and Stimulates Cholesterol Efflux in Macrophage-Derived Foam Cells. Biochem. Biophys. Res. Commun. 2017, 482, 93–99. [Google Scholar] [CrossRef]
- Ding, Y.; Feng, Y.; Zhu, W.; Zou, Y.; Xie, Y.; Wang, F.; Liu, C.; Zhang, Y.; Liu, H. [Gly14]-Humanin Prevents Lipid Deposition and Endothelial Cell Apoptosis in a Lectin-like Oxidized Low-density Lipoprotein Receptor-1-Dependent Manner. Lipids 2019, 54, 697–705. [Google Scholar] [CrossRef]
- Ding, Y.; Feng, Y.; Zou, Y.; Wang, F.; Liu, H.; Liu, C.; Zhang, Y. [Gly14]-Humanin Restores Cathepsin D Function via FPRL1 and Promotes Autophagic Degradation of Ox-LDL in HUVECs. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 2406–2416. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Yuan, Q.; Yu, H.; Xu, W.; Xie, X. New Insights into Oxidative Stress and Inflammation during Diabetes Mellitus-Accelerated Atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Das, H. KLF2 in Regulation of NF-ΚB-Mediated Immune Cell Function and Inflammation. Int. J. Mol. Sci. 2017, 18, 2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wu, Z.; He, Y.; Zhang, H.; Tian, L.; Zheng, C.; Shang, T.; Zhu, Q.; Li, D.; He, Y. Humanin Prevents High Glucose-Induced Monocyte Adhesion to Endothelial Cells by Targeting KLF2. Mol. Immunol. 2018, 101, 245–250. [Google Scholar] [CrossRef]
- Wei, M.; Gan, L.; Liu, Z.; Liu, L.; Chang, J.-R.; Yin, D.-C.; Cao, H.-L.; Su, X.-L.; Smith, W.W. Mitochondrial-Derived Peptide MOTS-c Attenuates Vascular Calcification and Secondary Myocardial Remodeling via Adenosine Monophosphate-Activated Protein Kinase Signaling Pathway. Cardiorenal Med. 2020, 10, 42–50. [Google Scholar] [CrossRef]
- Singh, K.D.; Karnik, S.S. Angiotensin Type 1 Receptor Blockers in Heart Failure. Curr. Drug Targets 2020, 21, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Delrio, S.; Wan, J.; Jay Widmer, R.; Cohen, P.; Lerman, L.O.; Lerman, A. Downregulation of Circulating MOTS-c Levels in Patients with Coronary Endothelial Dysfunction. Int. J. Cardiol. 2018, 254, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Ikonomidis, I.; Katogiannis, K.; Kyriakou, E.; Taichert, M.; Katsimaglis, G.; Tsoumani, M.; Andreadou, I.; Maratou, E.; Lambadiari, V.; Kousathana, F.; et al. β-Amyloid and Mitochondrial-Derived Peptide-c Are Additive Predictors of Adverse Outcome to High-on-Treatment Platelet Reactivity in Type 2 Diabetics with Revascularized Coronary Artery Disease. J. Thromb. Thrombolysis 2020, 49, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Miller, B.; Kumagai, H.; Silverstein, A.R.; Flores, M.; Yen, K. Mitochondrial-Derived Peptides in Aging and Age-Related Diseases. GeroScience 2021, 43, 1113–1121. [Google Scholar] [CrossRef]
- Reynolds, J.C.; Bwiza, C.P.; Lee, C. Mitonuclear Genomics and Aging. Hum. Genet. 2020, 139, 381–399. [Google Scholar] [CrossRef]
- Kim, S.-J.; Mehta, H.H.; Wan, J.; Kuehnemann, C.; Chen, J.; Hu, J.-F.; Hoffman, A.R.; Cohen, P. Mitochondrial Peptides Modulate Mitochondrial Function during Cellular Senescence. Aging 2018, 10, 1239–1256. [Google Scholar] [CrossRef]
- Conte, M.; Ostan, R.; Fabbri, C.; Santoro, A.; Guidarelli, G.; Vitale, G.; Mari, D.; Sevini, F.; Capri, M.; Sandri, M.; et al. Human Aging and Longevity Are Characterized by High Levels of Mitokines. J. Gerontol. Ser. A 2019, 74, 600–607. [Google Scholar] [CrossRef]
- Conte, M.; Sabbatinelli, J.; Chiariello, A.; Martucci, M.; Santoro, A.; Monti, D.; Arcaro, M.; Galimberti, D.; Scarpini, E.; Bonfigli, A.R.; et al. Disease-Specific Plasma Levels of Mitokines FGF21, GDF15, and Humanin in Type II Diabetes and Alzheimer’s Disease in Comparison with Healthy Aging. GeroScience 2021, 43, 985–1001. [Google Scholar] [CrossRef]
- Huang, T.-Y.; Zheng, D.; Houmard, J.A.; Brault, J.J.; Hickner, R.C.; Cortright, R.N. Overexpression of PGC-1α Increases Peroxisomal Activity and Mitochondrial Fatty Acid Oxidation in Human Primary Myotubes. Am. J. Physiol. Endocrinol. Metab. 2017, 312, E253–E263. [Google Scholar] [CrossRef] [Green Version]
- Römer, A.; Linn, T.; Petry, S.F. Lipotoxic Impairment of Mitochondrial Function in β-Cells: A Review. Antioxidants 2021, 10, 293. [Google Scholar] [CrossRef]
- Gidlund, E.-K.; von Walden, F.; Venojärvi, M.; Risérus, U.; Heinonen, O.J.; Norrbom, J.; Sundberg, C.J. Humanin Skeletal Muscle Protein Levels Increase after Resistance Training in Men with Impaired Glucose Metabolism. Physiol. Rep. 2016, 4, e13063. [Google Scholar] [CrossRef] [PubMed]
- Ramanjaneya, M.; Bettahi, I.; Jerobin, J.; Chandra, P.; Abi Khalil, C.; Skarulis, M.; Atkin, S.L.; Abou-Samra, A.-B. Mitochondrial-Derived Peptides Are Down Regulated in Diabetes Subjects. Front. Endocrinol. 2019, 10, 331. [Google Scholar] [CrossRef] [Green Version]
- Han, K.; Jia, N.; Zhong, Y.; Shang, X. S14G-humanin Alleviates Insulin Resistance and Increases Autophagy in Neurons of APP/PS1 Transgenic Mouse. J. Cell. Biochem. 2018, 119, 3111–3117. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Miller, B.; Kumagai, H.; Yen, K.; Cohen, P. MOTS-c: An Equal Opportunity Insulin Sensitizer. J. Mol. Med. 2019, 97, 487–490. [Google Scholar] [CrossRef]
- Kim, S.; Miller, B.; Mehta, H.H.; Xiao, J.; Wan, J.; Arpawong, T.E.; Yen, K.; Cohen, P. The Mitochondrial-derived Peptide MOTS-c Is a Regulator of Plasma Metabolites and Enhances Insulin Sensitivity. Physiol. Rep. 2019, 7, e14171. [Google Scholar] [CrossRef] [Green Version]
- Kwon, C.; Sun, J.L.; Jeong, J.H.; Jung, T.W. Humanin Attenuates Palmitate-Induced Hepatic Lipid Accumulation and Insulin Resistance via AMPK-Mediated Suppression of the MTOR Pathway. Biochem. Biophys. Res. Commun. 2020, 526, 539–545. [Google Scholar] [CrossRef]
- Magaway, C.; Kim, E.; Jacinto, E. Targeting MTOR and Metabolism in Cancer: Lessons and Innovations. Cells 2019, 8, 1584. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Su, K.; Cui, L.; Tas, E.; Zhang, T.; Dong, H.H.; Yakar, S.; Muzumdar, R.H. Central Effects of Humanin on Hepatic Triglyceride Secretion. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E283–E292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, H.H.; Xiao, J.; Ramirez, R.; Miller, B.; Kim, S.-J.; Cohen, P.; Yen, K. Metabolomic Profile of Diet-Induced Obesity Mice in Response to Humanin and Small Humanin-like Peptide 2 Treatment. Metabolomics 2019, 15, 88. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Yu, Q.; Chang, B.; Guo, Q.; Xu, S.; Yi, X.; Cao, S. MOTS-c Interacts Synergistically with Exercise Intervention to Regulate PGC-1α Expression, Attenuate Insulin Resistance and Enhance Glucose Metabolism in Mice via AMPK Signaling Pathway. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2021, 1867, 166126. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, H.; Hu, G.; Ye, Z.; Zhai, D.; Yan, Z.; Wang, L.; Xiang, A.; Lu, Z. Earlier Changes in Mice after D-Galactose Treatment Were Improved by Mitochondria Derived Small Peptide MOTS-c. Biochem. Biophys. Res. Commun. 2019, 513, 439–445. [Google Scholar] [CrossRef]
- Zhai, D.; Ye, Z.; Jiang, Y.; Xu, C.; Ruan, B.; Yang, Y.; Lei, X.; Xiang, A.; Lu, H.; Zhu, Z.; et al. MOTS-c Peptide Increases Survival and Decreases Bacterial Load in Mice Infected with MRSA. Mol. Immunol. 2017, 92, 151–160. [Google Scholar] [CrossRef]
- Lei, W.; Liu, D.; Sun, M.; Lu, C.; Yang, W.; Wang, C.; Cheng, Y.; Zhang, M.; Shen, M.; Yang, Z.; et al. Targeting STAT3: A Crucial Modulator of Sepsis. J. Cell Physiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W. Aryl Hydrocarbon Receptor (AHR)-Mediated Inflammation and Resolution: Non-Genomic and Genomic Signaling. Biochem. Pharmacol. 2020, 182, 114220. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, D.; Yuan, W.; Wang, C.; Huang, Q.; Luo, J. Humanin Ameliorates Free Fatty Acid-Induced Endothelial Inflammation by Suppressing the NLRP3 Inflammasome. ACS Omega 2020, 5, 22039–22045. [Google Scholar] [CrossRef]
- Stitham, J.; Rodriguez-Velez, A.; Zhang, X.; Jeong, S.-J.; Razani, B. Inflammasomes: A Preclinical Assessment of Targeting in Atherosclerosis. Expert Opin. Ther. Targets 2020, 24, 825–844. [Google Scholar] [CrossRef]
- Hemenway, G.; Frishman, W.H. Therapeutic Implications of NLRP3-Mediated Inflammation in Coronary Artery Disease. Cardiol. Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- McGuire, P.J. Mitochondrial Dysfunction and the Aging Immune System. Biology 2019, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Soysal, P.; Arik, F.; Smith, L.; Jackson, S.E.; Isik, A.T. Inflammation, Frailty and Cardiovascular Disease. In Frailty and Cardiovascular Diseases; Veronese, N., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2020; Volume 1216, pp. 55–64. ISBN 978-3-030-33329-4. [Google Scholar]
- Li, Z.; Sreekumar, P.G.; Peddi, S.; Hinton, D.R.; Kannan, R.; MacKay, J.A. The Humanin Peptide Mediates ELP Nanoassembly and Protects Human Retinal Pigment Epithelial Cells from Oxidative Stress. Nanomed. Nanotechnol. Biol. Med. 2020, 24, 102111. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cui, Y.; Tang, Y.; Tang, X.; Yu, X.; Zhou, J.; Yin, Q.; Shentu, X. Cytoprotective Role of Humanin in Lens Epithelial Cell Oxidative Stress-induced Injury. Mol. Med. Rep. 2020, 22, 1467–1479. [Google Scholar] [CrossRef]
- Zhao, J.; Zeng, Y.; Wang, Y.; Shi, J.; Zhao, W.; Wu, B.; Du, H. Humanin Protects Cortical Neurons from Calyculin A-Induced Neurotoxicities by Increasing PP2A Activity and SOD. Int. J. Neurosci. 2021, 131, 527–535. [Google Scholar] [CrossRef]
- Gurunathan, S.; Jeyaraj, M.; Kang, M.H.; Kim, J.H. Mitochondrial Peptide Humanin Protects Silver Nanoparticles-Induced Neurotoxicity in Human Neuroblastoma Cancer Cells (SH-SY5Y). Int. J. Mol. Sci. 2019, 20, 4439. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, D.; Sreekumar, P.G.; Ishikawa, K.; Terasaki, H.; Barron, E.; Cohen, P.; Kannan, R.; Hinton, D.R. Humanin Protects RPE Cells from Endoplasmic Reticulum Stress-Induced Apoptosis by Upregulation of Mitochondrial Glutathione. PLoS ONE 2016, 11, e0165150. [Google Scholar] [CrossRef] [Green Version]
- Dubois-Deruy, E.; Peugnet, V.; Turkieh, A.; Pinet, F. Oxidative Stress in Cardiovascular Diseases. Antioxidants 2020, 9, 864. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, P.G.; Ishikawa, K.; Spee, C.; Mehta, H.H.; Wan, J.; Yen, K.; Cohen, P.; Kannan, R.; Hinton, D.R. The Mitochondrial-Derived Peptide Humanin Protects RPE Cells From Oxidative Stress, Senescence, and Mitochondrial Dysfunction. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1238. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, H.; Wu, J.; Yin, L.; Yan, L.-J.; Zhang, C. Humanin Attenuates NMDA-Induced Excitotoxicity by Inhibiting ROS-Dependent JNK/P38 MAPK Pathway. Int. J. Mol. Sci. 2018, 19, 2982. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, S.N.; Li, J.-M. Nox2 Dependent Redox-Regulation of Akt and ERK1/2 to Promote Left Ventricular Hypertrophy in Dietary Obesity of Mice. Biochem. Biophys. Res. Commun. 2020, 528, 506–513. [Google Scholar] [CrossRef]
- Domingues, A.; Jolibois, J.; Marquet de Rougé, P.; Nivet-Antoine, V. The Emerging Role of TXNIP in Ischemic and Cardiovascular Diseases; A Novel Marker and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 1693. [Google Scholar] [CrossRef]
- Akyildiz, A.G.; Boran, T.; Jannuzzi, A.T.; Alpertunga, B. Mitochondrial Dynamics Imbalance and Mitochondrial Dysfunction Contribute to the Molecular Cardiotoxic Effects of Lenvatinib. Toxicol. Appl. Pharmacol. 2021, 423, 115577. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.-S.; Li, Y.; Zhai, H.; Bi, J.-W.; Zhang, F.-S.; Zhang, X.-Y.; Fan, S.-H. Humanin Analogue, S14G-Humanin, Has Neuroprotective Effects against Oxygen Glucose Deprivation/Reoxygenation by Reactivating Jak2/Stat3 Signaling through the PI3K/AKT Pathway. Exp. Ther. Med. 2017, 14, 3926–3934. [Google Scholar] [CrossRef] [Green Version]
- Song, H.-P.; Chu, Z.-G.; Zhang, D.-X.; Dang, Y.-M.; Zhang, Q. PI3K–AKT Pathway Protects Cardiomyocytes against Hypoxia-Induced Apoptosis by MitoKATP-Mediated Mitochondrial Translocation of PAKT. Cell Physiol. Biochem. 2018, 49, 717–727. [Google Scholar] [CrossRef]
- Cai, H.; Liu, Y.; Men, H.; Zheng, Y. Protective Mechanism of Humanin Against Oxidative Stress in Aging-Related Cardiovascular Diseases. Front. Endocrinol. 2021, 12, 683151. [Google Scholar] [CrossRef] [PubMed]
- Hazafa, A.; Batool, A.; Ahmad, S.; Amjad, M.; Chaudhry, S.N.; Asad, J.; Ghuman, H.F.; Khan, H.M.; Naeem, M.; Ghani, U. Humanin: A Mitochondrial-Derived Peptide in the Treatment of Apoptosis-Related Diseases. Life Sci. 2021, 264, 118679. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, D. Humanin Decreases Mitochondrial Membrane Permeability by Inhibiting the Membrane Association and Oligomerization of Bax and Bid Proteins. Acta Pharm. Sin. 2018, 39, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Kastner, D.W.; Johnson, S.; Strub, M.-P.; He, Y.; Bleck, C.K.E.; Lee, D.-Y.; Tjandra, N. Humanin Induces Conformational Changes in the Apoptosis Regulator BAX and Sequesters It into Fibers, Preventing Mitochondrial Outer-Membrane Permeabilization. J. Biol. Chem. 2019, 294, 19055–19065. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Johnson, S.; Bleck, C.K.E.; Lee, D.-Y.; Tjandra, N. Humanin Selectively Prevents the Activation of Pro-Apoptotic Protein BID by Sequestering It into Fibers. J. Biol. Chem. 2020, 295, 18226–18238. [Google Scholar] [CrossRef]
- Celvin, B.; Zaman, F.; Aulin, C.; Sävendahl, L. Humanin Prevents Undesired Apoptosis of Chondrocytes without Interfering with the Anti-Inflammatory Effect of Dexamethasone in Collagen-Induced Arthritis. Clin. Exp. Rheumatol. 2020, 38, 129–135. [Google Scholar]
- Jia, Y.; Lue, Y.; Swerdloff, R.S.; Lasky, J.L.; Panosyan, E.H.; Dai-Ju, J.; Wang, C. The Humanin Analogue (HNG) Prevents Temozolomide-Induced Male Germ Cell Apoptosis and Other Adverse Effects in Severe Combined Immuno-Deficiency (SCID) Mice Bearing Human Medulloblastoma. Exp. Mol. Pathol. 2019, 109, 42–50. [Google Scholar] [CrossRef]
- Ayala, M.A.M.; Gottardo, M.F.; Zuccato, C.F.; Pidre, M.L.; Nicola Candia, A.J.; Asad, A.S.; Imsen, M.; Romanowski, V.; Creton, A.; Isla Larrain, M.; et al. Humanin Promotes Tumor Progression in Experimental Triple Negative Breast Cancer. Sci. Rep. 2020, 10, 8542. [Google Scholar] [CrossRef] [PubMed]
- Vancheri, F.; Longo, G.; Vancheri, S.; Henein, M. Coronary Microvascular Dysfunction. J. Clin. Med. 2020, 9, 2880. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.J.; Flammer, A.J.; Herrmann, J.; Rodriguez-Porcel, M.; Wan, J.; Cohen, P.; Lerman, L.O.; Lerman, A. Circulating Humanin Levels Are Associated with Preserved Coronary Endothelial Function. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H393–H397. [Google Scholar] [CrossRef] [PubMed]
- López, B.; Ravassa, S.; Moreno, M.U.; José, G.S.; Beaumont, J.; González, A.; Díez, J. Diffuse Myocardial Fibrosis: Mechanisms, Diagnosis and Therapeutic Approaches. Nat. Rev. Cardiol. 2021, 18, 479–498. [Google Scholar] [CrossRef]
- Qin, Q.; Mehta, H.; Yen, K.; Navarrete, G.; Brandhorst, S.; Wan, J.; Delrio, S.; Zhang, X.; Lerman, L.O.; Cohen, P.; et al. Chronic Treatment with the Mitochondrial Peptide Humanin Prevents Age-Related Myocardial Fibrosis in Mice. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1127–H1136. [Google Scholar] [CrossRef] [Green Version]
- Liguori, T.T.A.; Liguori, G.R.; Moreira, L.F.P.; Harmsen, M.C. Fibroblast Growth Factor-2, but Not the Adipose Tissue-Derived Stromal Cells Secretome, Inhibits TGF-Β1-Induced Differentiation of Human Cardiac Fibroblasts into Myofibroblasts. Sci. Rep. 2018, 8, 16633. [Google Scholar] [CrossRef]
- Wang, Y.; Ge, C.; Chen, J.; Tang, K.; Liu, J. GSK-3β Inhibition Confers Cardioprotection Associated with the Restoration of Mitochondrial Function and Suppression of Endoplasmic Reticulum Stress in Sevoflurane Preconditioned Rats Following Ischemia/Reperfusion Injury. Perfusion 2018, 33, 679–686. [Google Scholar] [CrossRef] [PubMed]
- González-Montero, J.; Brito, R.; Gajardo, A.I.; Rodrigo, R. Myocardial Reperfusion Injury and Oxidative Stress: Therapeutic Opportunities. World J. Cardiol. 2018, 10, 74–86. [Google Scholar] [CrossRef]
- Arrigo, M.; Price, S.; Baran, D.A.; Pöss, J.; Aissaoui, N.; Bayes-Genis, A.; Bonello, L.; François, B.; Gayat, E.; Gilard, M.; et al. Optimising Clinical Trials in Acute Myocardial Infarction Complicated by Cardiogenic Shock: A Statement from the 2020 Critical Care Clinical Trialists Workshop. Lancet Respir. Med. 2021. [Google Scholar] [CrossRef]
- Thummasorn, S.; Shinlapawittayatorn, K.; Chattipakorn, S.C.; Chattipakorn, N. High-Dose Humanin Analogue Applied during Ischemia Exerts Cardioprotection against Ischemia/Reperfusion Injury by Reducing Mitochondrial Dysfunction. Cardiovasc. Ther. 2017, 35, e12289. [Google Scholar] [CrossRef]
- Thummasorn, S.; Apaijai, N.; Kerdphoo, S.; Shinlapawittayatorn, K.; Chattipakorn, S.C.; Chattipakorn, N. Humanin Exerts Cardioprotection against Cardiac Ischemia/Reperfusion Injury through Attenuation of Mitochondrial Dysfunction. Cardiovasc. Ther. 2016, 34, 404–414. [Google Scholar] [CrossRef]
- Thummasorn, S.; Shinlapawittayatorn, K.; Khamseekaew, J.; Jaiwongkam, T.; Chattipakorn, S.C.; Chattipakorn, N. Humanin Directly Protects Cardiac Mitochondria against Dysfunction Initiated by Oxidative Stress by Decreasing Complex I Activity. Mitochondrion 2018, 38, 31–40. [Google Scholar] [CrossRef]
- Kumfu, S.; Charununtakorn, S.T.; Jaiwongkam, T.; Chattipakorn, N.; Chattipakorn, S.C. Humanin Prevents Brain Mitochondrial Dysfunction in a Cardiac Ischaemia-Reperfusion Injury Model: Humanin and Brain after Cardiac Ischaemia-Reperfusion Injury. Exp. Physiol. 2016, 101, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumfu, S.; Charununtakorn, S.T.; Jaiwongkam, T.; Chattipakorn, N.; Chattipakorn, S.C. Humanin Exerts Neuroprotection During Cardiac Ischemia-Reperfusion Injury. J. Alzheimer’s Dis. 2018, 61, 1343–1353. [Google Scholar] [CrossRef]
- Sharp, T.E.; Gong, Z.; Scarborough, A.; Goetzman, E.S.; Ali, M.J.; Spaletra, P.; Lefer, D.J.; Muzumdar, R.H.; Goodchild, T.T. Efficacy of a Novel Mitochondrial-Derived Peptide in a Porcine Model of Myocardial Ischemia/Reperfusion Injury. JACC Basic Transl. Sci. 2020, 5, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Li, Q.; You, T.; Zhao, X.; Xu, X.; Tang, C.; Zhu, L. Humanin Analogue, HNG, Inhibits Platelet Activation and Thrombus Formation by Stabilizing Platelet Microtubules. J. Cell Mol. Med. 2020, 24, 4773–4783. [Google Scholar] [CrossRef] [PubMed]
- Gilon, C.; Gitlin-Domagalska, A.; Lahiani, A.; Yehoshua- Alshanski, S.; Shumacher-Klinger, A.; Gilon, D.; Taha, M.; Sekler, I.; Hoffman, A.; Lazarovici, P. Novel Humanin Analogs Confer Neuroprotection and Myoprotection to Neuronal and Myoblast Cell Cultures Exposed to Ischemia-like and Doxorubicin-Induced Cell Death Insults. Peptides 2020, 134, 170399. [Google Scholar] [CrossRef]
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline Chemotherapy and Cardiotoxicity. Cardiovasc. Drugs Ther. 2017, 31, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Lue, Y.; Gao, C.; Swerdloff, R.; Hoang, J.; Avetisyan, R.; Jia, Y.; Rao, M.; Ren, S.; Atienza, V.; Yu, J.; et al. Humanin Analog Enhances the Protective Effect of Dexrazoxane against Doxorubicin-Induced Cardiotoxicity. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H634–H643. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Shi, H.; He, Y.; Yuan, L.; Qu, X.; Zhang, J.; Wang, Z.; Cai, H.; Qi, J. Colivelin Ameliorates Impairments in Cognitive Behaviors and Synaptic Plasticity in APP/PS1 Transgenic Mice. J. Alzheimer’s Dis. 2017, 59, 1067–1078. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, Y.; Wei, C.; Li, Y.; Ma, H.; Wang, X.; Cui, Z.; Jin, W.-N.; Shi, F.-D. Colivelin Rescues Ischemic Neuron and Axons Involving JAK/STAT3 Signaling Pathway. Neuroscience 2019, 416, 198–206. [Google Scholar] [CrossRef]
- Solanki, A.; Smalling, R.; Parola, A.H.; Nathan, I.; Kasher, R.; Pathak, Y.; Sutariya, V. Humanin Nanoparticles for Reducing Pathological Factors Characteristic of Age-Related Macular Degeneration. Curr. Drug Deliv. 2019, 16, 226–232. [Google Scholar] [CrossRef]
- Pożarowska, D.; Pożarowski, P. The Era of Anti-Vascular Endothelial Growth Factor (VEGF) Drugs in Ophthalmology, VEGF and Anti-VEGF Therapy. CEJOI 2016, 3, 311–316. [Google Scholar] [CrossRef]
- Gottardo, M.F.; Pidre, M.L.; Zuccato, C.; Asad, A.S.; Imsen, M.; Jaita, G.; Candolfi, M.; Romanowski, V.; Seilicovich, A. Baculovirus-Based Gene Silencing of Humanin for the Treatment of Pituitary Tumors. Apoptosis 2018, 23, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Nan, A.; Chen, L.; Zhang, N.; Jia, Y.; Li, X.; Zhou, H.; Ling, Y.; Wang, Z.; Yang, C.; Liu, S.; et al. Circular RNA CircNOL10 Inhibits Lung Cancer Development by Promoting SCLM1-Mediated Transcriptional Regulation of the Humanin Polypeptide Family. Adv. Sci. 2019, 6, 1800654. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| CVD Risk Factor | MDP | The Way of Action | References |
|---|---|---|---|
| Atherosclerosis | HNG | prevents CD36 and LOX-1 upregulation; upregulates the expression of ABCA1 and ABCG1 | [34] |
| suppress LOX-1 expression | [35] | ||
| promotes autophagic degradation of Ox-LDL throughout lysosomal enzyme cathepsin D and its receptor FPRL1 | [36] | ||
| NH | induces the expression of KLF2;inhibits the high glucose-induced secretion of TNF-α and IL-1β; reduces the expression of VCAM-1 and E-selectin | [39] | |
| MOTS-c | decreases the expression levels of the AT-1 and ET-B receptors; increased the level of phosphorylated AMPK | [40] | |
| Ageing | MOTS-c and HN | increase secretion of SASP components and rise mitochondrial respiration | [46] |
| stimulate the secretion of SASP components (cytokines IL-1β, IL-6, IL-8, IL-10 and TNF-α) | [8] | ||
| Insulin resistance | HN | increases insulin sensitivity in muscles and the liver; increase fatty acid metabolism signalling and insulin-mediated AKT-signalling | [51,52] |
| HNG | improves insulin sensitivity in the brain through the regulation of IRS-1/mTOR signalling pathway in the hippocampus | [53] | |
| SHLP2 and SHLP3 | enhance 3T3-L1 pre-adipocyte differentiation | [4] | |
| SHLP2 | SHLP2 increased glucose uptake and suppressed hepatic glucose production | [4] | |
| MOTS-c | increases fatty acids β-oxidation and mitochondria biogenesis; stimulates GLUT4 expression | [54,55] | |
| Hyperlipidaemia | HN | decreases lipid accumulation and down-regulates lipogenesis genes (SREBP1, FAS and SCD1); enhances AMPK phosphorylation-mediated effect on IR; suppresses phosphorylation of the mTOR signalling pathway | [56] |
| HNG | decreases liver triglyceride accumulation, visceral fat and body weight gain in high-fat diet-fed mice | [58] | |
| HNG and SHLP2 | acting via the glutathione and sphingolipid metabolism pathways alter the concentrations of amino acid and lipid metabolites in plasma | [59] | |
| MOTS-c | increases AICAR levels and activates AMPK | [3] | |
| activates AMPK, GLUT4 and AC; stimulating mitochondria biogenesis and increasing the level of fatty acid β-oxidation | [60] | ||
| improves mitochondria dynamics | [61] | ||
| regulates expression of ARE and NFE2L2/NRF2 | [21] |
| CVD | MDP | The Way of Action | References |
|---|---|---|---|
| CMD | HN | HN level is decreased in CMD patients; HN is a biomarker of ED | [92] |
| MOTS-c | MOTS-c level correlates with ED | [42] | |
| Myocardial fibrosis | HNG | reduces apoptosis, collagen deposition in aged hearts, cardiac fibroblast proliferation and expression of MMP2, FGF2 and TGFB1; up-regulates AKT/GSK3B pathway | [94] |
| Myocardial ischemia and reperfusion injury | HNG | attenuates the heart mitochondrial dysfunction; decreases left ventricular dysfunction, myocardial infarct size and cardiac arrhythmia | [99,100] |
| attenuates cardiac mitochondria swelling, depolarization and reduced ATP production; decreases mitochondrial complex I activity. | [101] | ||
| protects brain mitochondria during AMI and MRI, reducing tau hyperphosphorylation, Aβ accumulation and apoptosis | [102,103] | ||
| inhibits platelet aggregation, P-selectin expression, αIIbβ3 activation and adhesion under flow conditions; enhances tubulin acetylation and inhibits microtubule depolymerization | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabravolski, S.A.; Nikiforov, N.G.; Starodubova, A.V.; Popkova, T.V.; Orekhov, A.N. The Role of Mitochondria-Derived Peptides in Cardiovascular Diseases and Their Potential as Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 8770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168770
Dabravolski SA, Nikiforov NG, Starodubova AV, Popkova TV, Orekhov AN. The Role of Mitochondria-Derived Peptides in Cardiovascular Diseases and Their Potential as Therapeutic Targets. International Journal of Molecular Sciences. 2021; 22(16):8770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168770
Chicago/Turabian StyleDabravolski, Siarhei A., Nikita G. Nikiforov, Antonina V. Starodubova, Tatyana V. Popkova, and Alexander N. Orekhov. 2021. "The Role of Mitochondria-Derived Peptides in Cardiovascular Diseases and Their Potential as Therapeutic Targets" International Journal of Molecular Sciences 22, no. 16: 8770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168770