
The CellBox-2 Mission to the International Space Station: Thyroid Cancer Cells in Space


 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
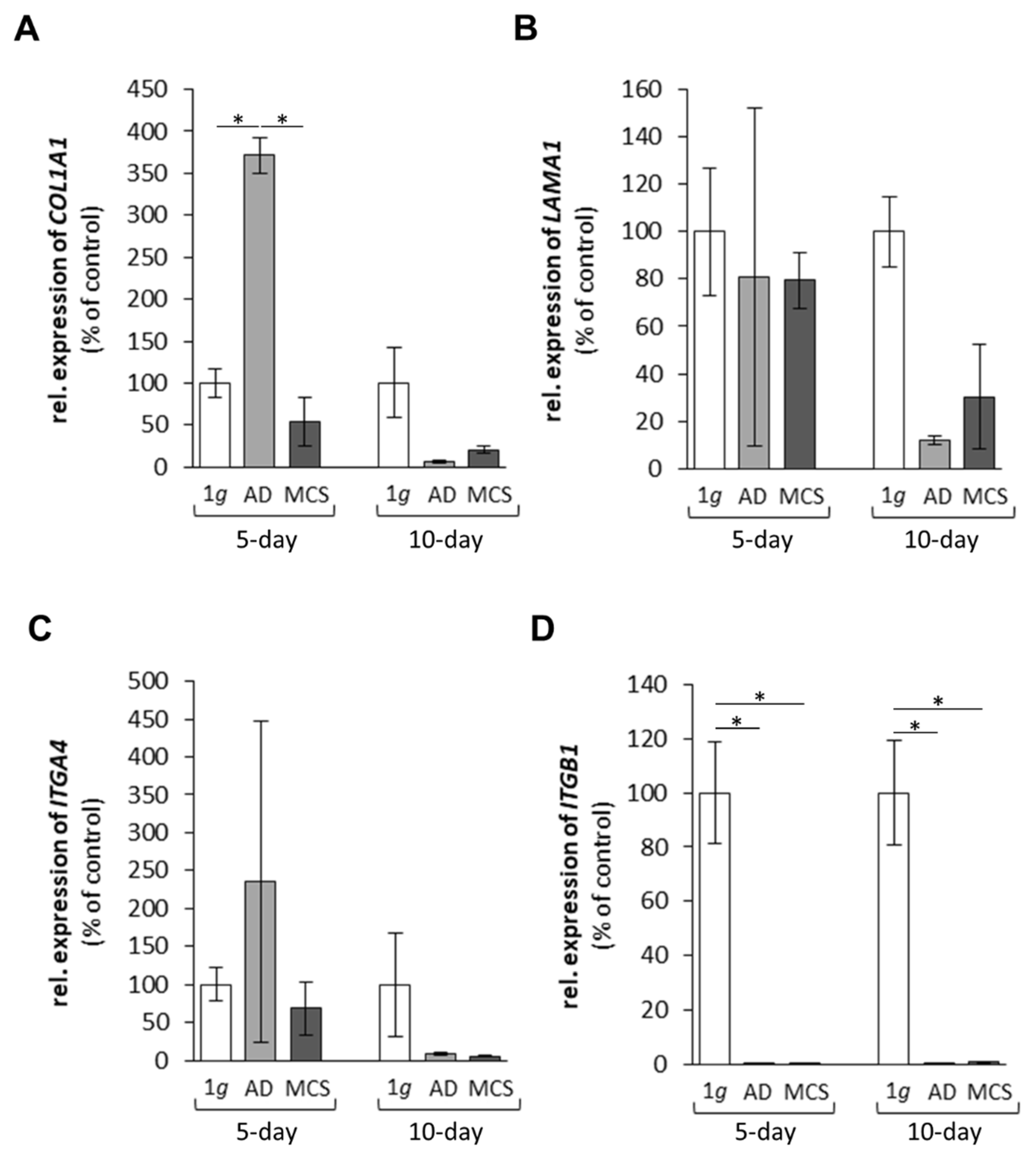
2.1. Impact of Microgravity on Selected Genes
2.1.1. Genes Encoding Extracellular Matrix Proteins
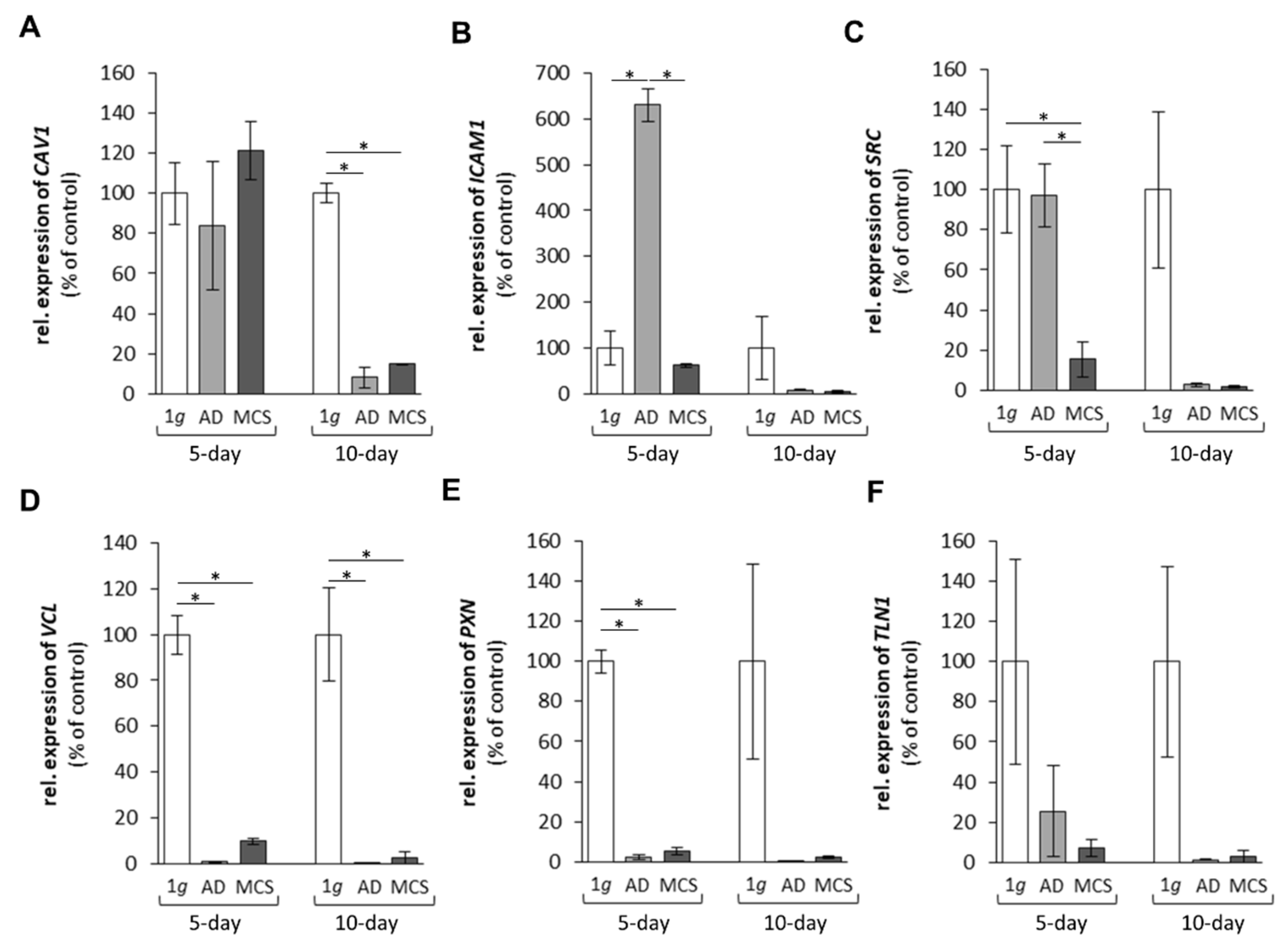
2.1.2. Genes Encoding for Membrane Proteins Involved in Cell Adhesion
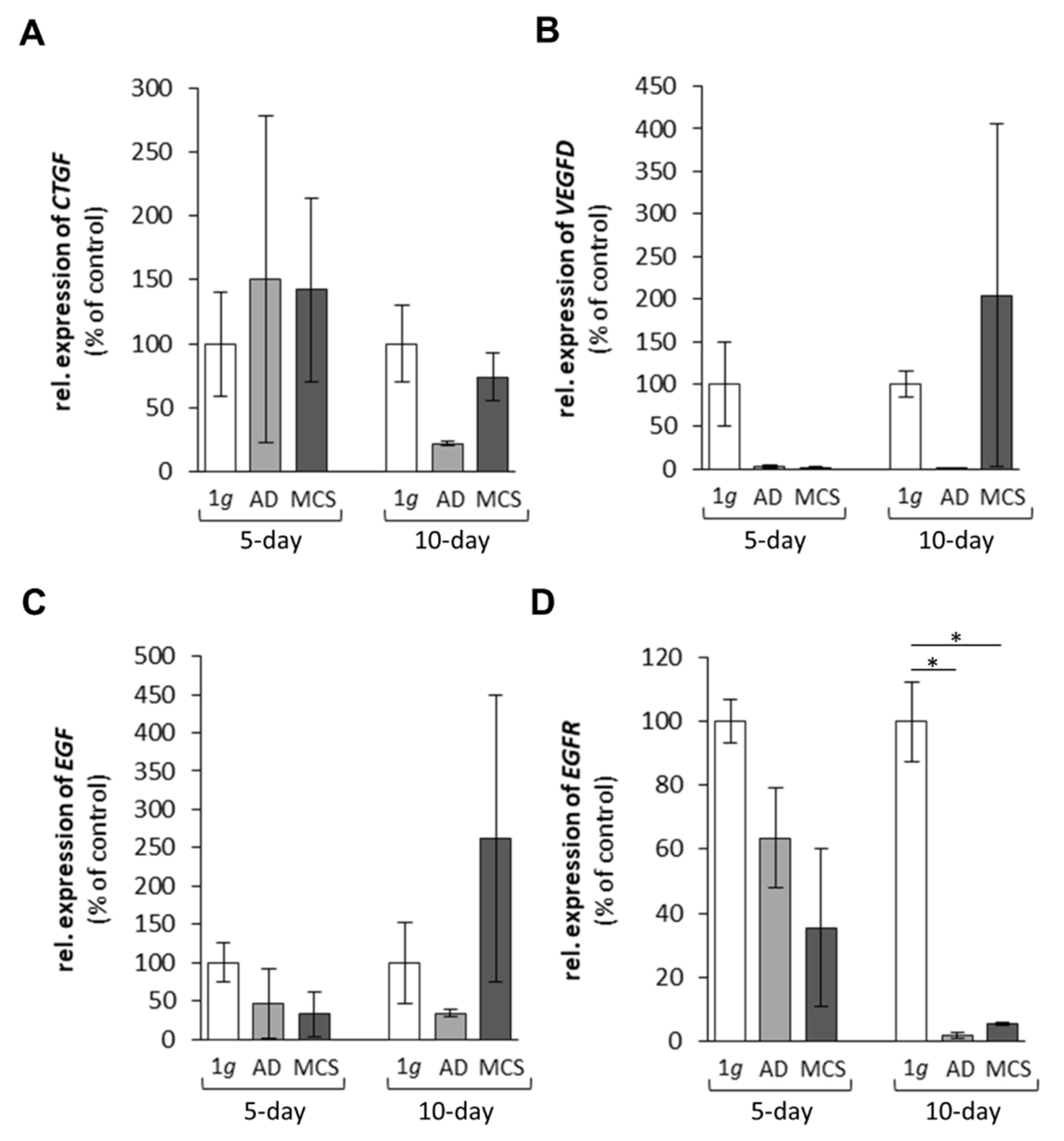
2.1.3. Genes Encoding for Factors Promoting Angiogenesis and Tumour Growth
2.1.4. Cytokines and the Extracellular Signal-Regulated Kinases
2.2. Impact of Microgravity on the Secretion of Proteins
Secreted Factors Involved in Angiogenesis
3. Discussion
3.1. Changes in Gene Expression Patterns of Thyroid Cancer Cells in Microgravity
3.1.1. Extracellular Matrix Proteins Altered by r-µg Exposure
3.1.2. Cell Adhesion Factors Changed in Space
3.1.3. Growth Factors and Signalling Molecules Altered in Microgravity
3.2. Interaction of Genes Involved in Angiogenesis and Spheroid Formation
4. Materials and Methods
4.1. Cell Cultures
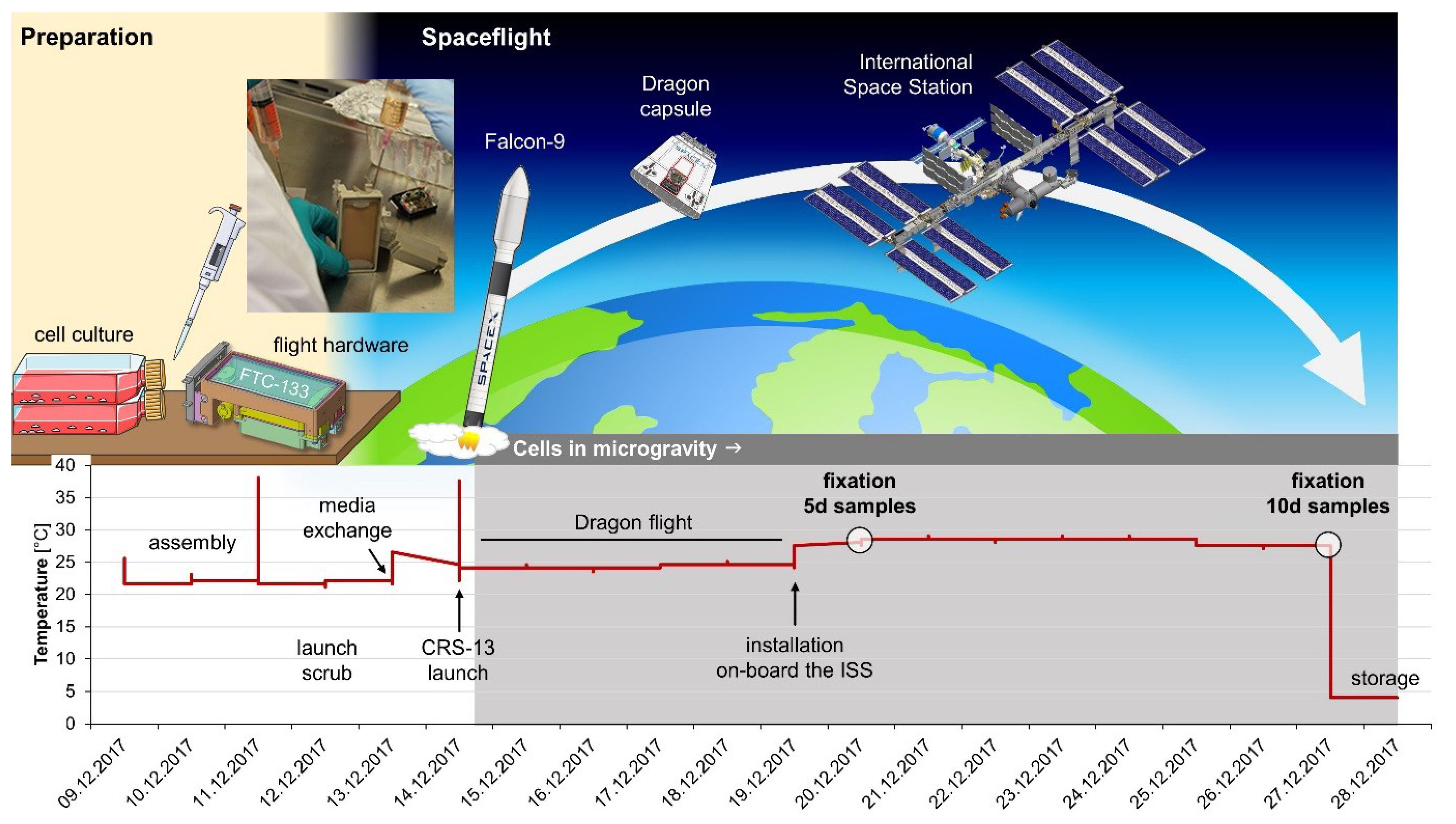
4.2. CellBox-2 Spaceflight Experiment
4.3. CellBox-2 Ground Control Experiment
4.4. RNA Extraction and Quantitative Polymerase Chain Reaction (qPCR)
4.5. Protein Measurements by Multi-Analyte Profiling Technology
4.6. Enzyme-Linked Immunosorbent Assay (ELISA) Measurements from Cell Culture Supernatants
4.7. Proteomic Profiling of Plasma Analytes
One-Way Analysis of Variance
4.8. Pathway Analysis
4.9. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- The American Cancer Society. Key Statistics for Thyroid Cancer. How Common is Thyroid Cancer? Available online: https://www.cancer.org/cancer/thyroid-cancer/about/key-statistics.html (accessed on 5 July 2021).
- Grimm, D.; Bauer, J.; Wise, P.; Kruger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of SOX family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67, 122–153. [Google Scholar] [CrossRef]
- Ancker, O.V.; Kruger, M.; Wehland, M.; Infanger, M.; Grimm, D. Multikinase Inhibitor Treatment in Thyroid Cancer. Int. J. Mol. Sci. 2019, 21, 10. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.N.; Shaha, A.R. Poorly differentiated thyroid cancer. Curr. Opin. Otolaryngol. Head Neck Surg. 2014, 22, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, A.; Kasai, N.; Sugano, H. Poorly differentiated carcinoma of the thyroid. A clinicopathologic entity for a high-risk group of papillary and follicular carcinomas. Cancer 1983, 52, 1849–1855. [Google Scholar] [CrossRef]
- Schlumberger, M.; Leboulleux, S. Current practice in patients with differentiated thyroid cancer. Nat. Rev. Endocrinol. 2021, 17, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Kruger, M.; Hemmersbach, R.; Lazzarini, A.; Cataldi, S.; Codini, M.; Beccari, T.; Ambesi-Impiombato, F.S.; Curcio, F. Impact of Gravity on Thyroid Cells. Int. J. Mol. Sci. 2017, 18, 972. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, D.; Bauer, J.; Hofstadter, F.; Riegger, G.A.; Kromer, E.P. Characteristics of multicellular spheroids formed by primary cultures of human thyroid tumor cells. Thyroid 1997, 7, 859–865. [Google Scholar] [CrossRef]
- Goretzki, P.E.; Frilling, A.; Simon, D.; Roeher, H.D. Growth regulation of normal thyroids and thyroid tumors in man. Recent Results Cancer Res. 1990, 118, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Schonberger, J.; Bauer, J.; Spruss, T.; Weber, G.; Chahoud, I.; Eilles, C.; Grimm, D. Establishment and characterization of the follicular thyroid carcinoma cell line ML-1. J. Mol. Med. 2000, 78, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.L.; Souza, G.R. Using space-based investigations to inform cancer research on Earth. Nat. Rev. Cancer 2013, 13, 315–327. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schoberger, J.; Pickenhahn, H.; Schulze-Tanzil, G.; Vetter, R.; Eilles, C.; Paul, M.; et al. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. FASEB J. 2002, 16, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Warnke, E.; Pietsch, J.; Wehland, M.; Bauer, J.; Infanger, M.; Gorog, M.; Hemmersbach, R.; Braun, M.; Ma, X.; Sahana, J.; et al. Spheroid formation of human thyroid cancer cells under simulated microgravity: A possible role of CTGF and CAV1. Cell Commun. Signal 2014, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Svejgaard, B.; Wehland, M.; Ma, X.; Kopp, S.; Sahana, J.; Warnke, E.; Aleshcheva, G.; Hemmersbach, R.; Hauslage, J.; Grosse, J.; et al. Common Effects on Cancer Cells Exerted by a Random Positioning Machine and a 2D Clinostat. PLoS ONE 2015, 10, e0135157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.; Kopp, S.; Schlagberger, E.M.; Grosse, J.; Sahana, J.; Riwaldt, S.; Wehland, M.; Luetzenberg, R.; Infanger, M.; Grimm, D. Proteome Analysis of Human Follicular Thyroid Cancer Cells Exposed to the Random Positioning Machine. Int. J. Mol. Sci. 2017, 18, 546. [Google Scholar] [CrossRef] [Green Version]
- Kopp, S.; Krüger, M.; Feldmann, S.; Oltmann, H.; Schütte, A.; Schmitz, B.; Bauer, J.; Schulz, H.; Saar, K.; Huebner, N.; et al. Thyroid cancer cells in space during the TEXUS-53 sounding rocket mission—The THYROID Project. Sci. Rep. 2018, 8, 10355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corydon, T.J.; Kopp, S.; Wehland, M.; Braun, M.; Schutte, A.; Mayer, T.; Hulsing, T.; Oltmann, H.; Schmitz, B.; Hemmersbach, R.; et al. Alterations of the cytoskeleton in human cells in space proved by life-cell imaging. Sci. Rep. 2016, 6, 20043. [Google Scholar] [CrossRef] [Green Version]
- Melnik, D.; Kruger, M.; Kopp, S.; Wehland, M.; Bauer, J.; Infanger, M.; Grimm, D. Microgravity-based Modulation of VEGF Expression in Human Thyroid Carcinoma Cells. Front. Physiol. 2019. [Google Scholar] [CrossRef]
- Riwaldt, S.; Pietsch, J.; Sickmann, A.; Bauer, J.; Braun, M.; Segerer, J.; Schwarzwalder, A.; Aleshcheva, G.; Corydon, T.J.; Infanger, M.; et al. Identification of proteins involved in inhibition of spheroid formation under microgravity. Proteomics 2015, 15, 2945–2952. [Google Scholar] [CrossRef]
- Pietsch, J.; Ma, X.; Wehland, M.; Aleshcheva, G.; Schwarzwälder, A.; Segerer, J.; Birlem, M.; Horn, A.; Bauer, J.; Infanger, M.; et al. Spheroid formation of human thyroid cancer cells in an automated culturing system during the Shenzhou-8 Space mission. Biomaterials 2013, 34, 7694–7705. [Google Scholar] [CrossRef]
- Ma, X.; Pietsch, J.; Wehland, M.; Schulz, H.; Saar, K.; Hübner, N.; Bauer, J.; Braun, M.; Schwarzwalder, A.; Segerer, J.; et al. Differential gene expression profile and altered cytokine secretion of thyroid cancer cells in space. FASEB J. 2014, 28, 813–835. [Google Scholar] [CrossRef]
- Riwaldt, S.; Bauer, J.; Pietsch, J.; Braun, M.; Segerer, J.; Schwarzwalder, A.; Corydon, T.J.; Infanger, M.; Grimm, D. The Importance of Caveolin-1 as Key-Regulator of Three-Dimensional Growth in Thyroid Cancer Cells Cultured under Real and Simulated Microgravity Conditions. Int. J. Mol. Sci. 2015, 16, 28296–28310. [Google Scholar] [CrossRef]
- Pietsch, J.; Sickmann, A.; Weber, G.; Bauer, J.; Egli, M.; Wildgruber, R.; Infanger, M.; Grimm, D. A proteomic approach to analysing spheroid formation of two human thyroid cell lines cultured on a random positioning machine. Proteomics 2011, 11, 2095–2104. [Google Scholar] [CrossRef]
- Topal, U.; Zamur, C. Microgravity, Stem Cells, and Cancer: A New Hope for Cancer Treatment. Stem Cells Int. 2021, 2021, 5566872. [Google Scholar] [CrossRef]
- Hammond, T.G.; Benes, E.; O’Reilly, K.C.; Wolf, D.A.; Linnehan, R.M.; Taher, A.; Kaysen, J.H.; Allen, P.L.; Goodwin, T.J. Mechanical culture conditions effect gene expression: Gravity-induced changes on the space shuttle. Physiol. Genom. 2000, 3, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.G.; Allen, P.L.; Birdsall, H.H. Effects of Space Flight on Mouse Liver versus Kidney: Gene Pathway Analyses. Int. J. Mol. Sci. 2018, 19, 4106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, E.M.; Yoshida, M.C.; Candelario, T.L.; Hughes-Fulford, M. Spaceflight and simulated microgravity cause a significant reduction of key gene expression in early T-cell activation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R480–R488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Wang, J.; Wang, C.; Xia, L.; Xu, J.; Xie, X.; Lu, W. Microenvironment remodeled by tumor and stromal cells elevates fibroblast-derived COL1A1 and facilitates ovarian cancer metastasis. Exp. Cell Res. 2020, 394, 112153. [Google Scholar] [CrossRef] [PubMed]
- Kopp, S.; Warnke, E.; Wehland, M.; Aleshcheva, G.; Magnusson, N.E.; Hemmersbach, R.; Corydon, T.J.; Bauer, J.; Infanger, M.; Grimm, D. Mechanisms of three-dimensional growth of thyroid cells during long-term simulated microgravity. Sci. Rep. 2015, 5, 16691. [Google Scholar] [CrossRef] [Green Version]
- Plosa, E.J.; Benjamin, J.T.; Sucre, J.M.; Gulleman, P.M.; Gleaves, L.A.; Han, W.; Kook, S.; Polosukhin, V.V.; Haake, S.M.; Guttentag, S.H.; et al. beta1 Integrin regulates adult lung alveolar epithelial cell inflammation. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Krüger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schütte, A.; et al. Real Microgravity Influences the Cytoskeleton and Focal Adhesions in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Li, Q.; Cao, Q.; Diao, Y.; Zhang, Y.; Yue, L.; Wei, L. EMT Transcription Factors Are Involved in the Altered Cell Adhesion under Simulated Microgravity Effect or Overloading by Regulation of E-cadherin. Int. J. Mol. Sci. 2020, 21, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, N.; Masiello, M.G.; Proietti, S.; Catizone, A.; Ricci, G.; Harrath, A.H.; Alwasel, S.H.; Cucina, A.; Bizzarri, M. Survival Pathways Are Differently Affected by Microgravity in Normal and Cancerous Breast Cells. Int. J. Mol. Sci. 2021, 22, 862. [Google Scholar] [CrossRef] [PubMed]
- Jokhadar, S.Z.; Majhenc, J.; Svetina, S.; Batista, U. Positioning of integrin beta1, caveolin-1 and focal adhesion kinase on the adhered membrane of spreading cells. Cell Biol. Int. 2013, 37, 1276–1284. [Google Scholar] [CrossRef]
- Tauber, S.; Lauber, B.A.; Paulsen, K.; Layer, L.E.; Lehmann, M.; Hauschild, S.; Shepherd, N.R.; Polzer, J.; Segerer, J.; Thiel, C.S.; et al. Cytoskeletal stability and metabolic alterations in primary human macrophages in long-term microgravity. PLoS ONE 2017, 12, e0175599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Golz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in cells of the monocyte/macrophage system in microgravity. Biomed. Res. Int. 2015, 2015, 538786. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, C.; Sun, S.; Zhang, C.; Lu, D.; Chen, Q.; Long, M. Microgravity-Induced Alterations of Inflammation-Related Mechanotransduction in Endothelial Cells on Board SJ-10 Satellite. Front. Physiol. 2018, 9, 1025. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Melnik, D.; Corydon, T.J.; Sahana, J.; Kruger, M.; Wehland, M.; Bauer, T.J.; Liemersdorf, C.; Hemmersbach, R.; et al. Short-Term Microgravity Influences Cell Adhesion in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 5730. [Google Scholar] [CrossRef] [Green Version]
- Hayes, S.H.; Seigel, G.M. Immunoreactivity of ICAM-1 in human tumors, metastases and normal tissues. Int. J. Clin. Exp. Pathol. 2009, 2, 553–560. [Google Scholar] [PubMed]
- Irby, R.B.; Yeatman, T.J. Role of Src expression and activation in human cancer. Oncogene 2000, 19, 5636–5642. [Google Scholar] [CrossRef] [Green Version]
- Cho, N.L.; Lin, C.I.; Du, J.; Whang, E.E.; Ito, H.; Moore, F.D., Jr.; Ruan, D.T. Global tyrosine kinome profiling of human thyroid tumors identifies Src as a promising target for invasive cancers. Biochem. Biophys. Res. Commun. 2012, 421, 508–513. [Google Scholar] [CrossRef]
- Spanjaard, E.; de Rooij, J. Mechanotransduction: Vinculin provides stability when tension rises. Curr. Biol. 2013, 23, R159–R161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stervbo, U.; Roch, T.; Westhoff, T.H.; Gayova, L.; Kurchenko, A.; Seibert, F.S.; Babel, N. Repeated Changes to the Gravitational Field Negatively Affect the Serum Concentration of Select Growth Factors and Cytokines. Front. Physiol. 2019, 10, 402. [Google Scholar] [CrossRef] [PubMed]
- Rijken, P.J.; Boonstra, J.; Verkleij, A.J.; de Laat, S.W. Effects of gravity on the cellular response to epidermal growth factor. Adv. Space Biol. Med. 1994, 4, 159–188. [Google Scholar] [CrossRef]
- Jessup, J.M.; Frantz, M.; Sonmez-Alpan, E.; Locker, J.; Skena, K.; Waller, H.; Battle, P.; Nachman, A.; Thomas, D.A.; Weber, M.E.; et al. Microgravity culture reduces apoptosis and increases the differentiation of a human colorectal carcinoma cell line. In Vitro Cell Dev. Biol. Anim. 2000, 36, 367–373. [Google Scholar] [CrossRef]
- Nersita, R.; Matrone, A.; Klain, M.; Scavuzzo, F.; Vitolo, G.; Abbondanza, C.; Carlino, M.V.; Giacco, V.; Amato, G.; Carella, C. Decreased serum vascular endothelial growth factor-D levels in metastatic patients with differentiated thyroid carcinoma. Clin. Endocrinol. 2012, 76, 142–146. [Google Scholar] [CrossRef]
- Willett, C.G.; Duda, D.G.; di Tomaso, E.; Boucher, Y.; Ancukiewicz, M.; Sahani, D.V.; Lahdenranta, J.; Chung, D.C.; Fischman, A.J.; Lauwers, G.Y.; et al. Efficacy, safety, and biomarkers of neoadjuvant bevacizumab, radiation therapy, and fluorouracil in rectal cancer: A multidisciplinary phase II study. J. Clin. Oncol. 2009, 27, 3020–3026. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Manegold, P.C.; Hong, Y.K.; Zhang, W.; Pohl, A.; Lurje, G.; Winder, T.; Yang, D.; LaBonte, M.J.; Wilson, P.M.; et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int. J. Cancer 2011, 128, 2038–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.X.; Sahani, D.V.; Duda, D.G.; di Tomaso, E.; Ancukiewicz, M.; Catalano, O.A.; Sindhwani, V.; Blaszkowsky, L.S.; Yoon, S.S.; Lahdenranta, J.; et al. Efficacy, safety, and potential biomarkers of sunitinib monotherapy in advanced hepatocellular carcinoma: A phase II study. J. Clin. Oncol. 2009, 27, 3027–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Xie, K.; Wu, H.; Zhang, B.; Huang, C. Identification and functional study of cytokines and chemokines involved in tumorigenesis. Comb. Chem. High Throughput. Screen 2012, 15, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Pyo, J.S.; Kang, G.; Kim, D.H.; Chae, S.W.; Park, C.; Kim, K.; Do, S.I.; Lee, H.J.; Kim, J.H.; Sohn, J.H. Activation of nuclear factor-kappaB contributes to growth and aggressiveness of papillary thyroid carcinoma. Pathol. Res. Pract. 2013, 209, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Brown, R.E. Morphoproteomic confirmation of an activated nuclear factor-small ka, CyrillicBp65 pathway in follicular thyroid carcinoma. Int. J. Clin. Exp. Pathol. 2012, 5, 216–223. [Google Scholar]
- Busca, R.; Pouyssegur, J.; Lenormand, P. ERK1 and ERK2 Map Kinases: Specific Roles or Functional Redundancy? Front. Cell Dev. Biol. 2016, 4, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Li, R.; Tan, X.; Zhang, J.; Fan, C.; Zhao, Q.; Deng, Y.; Xu, A.; Lukong, K.E.; Genth, H.; et al. Simulated Microgravity Reduces Focal Adhesions and Alters Cytoskeleton and Nuclear Positioning Leading to Enhanced Apoptosis via Suppressing FAK/RhoA-Mediated mTORC1/NF-kappaB and ERK1/2 Pathways. Int. J. Mol. Sci. 2018, 19, 1994. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Tian, H.; Wang, P.; Li, L.; Zhang, Z.; Zhang, J.; Zhao, Y. Spaceflight and simulated microgravity suppresses macrophage development via altered RAS/ERK/NFkappaB and metabolic pathways. Cell Mol. Immunol. 2021, 18, 1489–1502. [Google Scholar] [CrossRef] [Green Version]
- Grosse, J.; Wehland, M.; Pietsch, J.; Schulz, H.; Saar, K.; Hübner, N.; Eilles, C.; Bauer, J.; Abou-El-Ardat, K.; Baatout, S.; et al. Gravity-sensitive signaling drives 3-dimensional formation of multicellular thyroid cancer spheroids. FASEB J. 2012, 26, 5124–5140. [Google Scholar] [CrossRef] [Green Version]
- Sahana, J.; Nassef, M.Z.; Wehland, M.; Kopp, S.; Krüger, M.; Corydon, T.J.; Infanger, M.; Bauer, J.; Grimm, D. Decreased E-Cadherin in MCF7 Human Breast Cancer Cells Forming Multicellular Spheroids Exposed to Simulated Microgravity. Proteomics 2018, 18, e1800015. [Google Scholar] [CrossRef]
- Li, T.; Guo, H.; Song, Y.; Zhao, X.; Shi, Y.; Lu, Y.; Hu, S.; Nie, Y.; Fan, D.; Wu, K. Loss of vinculin and membrane-bound beta-catenin promotes metastasis and predicts poor prognosis in colorectal cancer. Mol. Cancer 2014, 13, 263. [Google Scholar] [CrossRef] [Green Version]
- Vendramini-Costa, D.B.; Carvalho, J.E. Molecular link mechanisms between inflammation and cancer. Curr. Pharm. Des. 2012, 18, 3831–3852. [Google Scholar] [CrossRef]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumour. Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef]
- Huang, S.P.; Wu, M.S.; Shun, C.T.; Wang, H.P.; Lin, M.T.; Kuo, M.L.; Lin, J.T. Interleukin-6 increases vascular endothelial growth factor and angiogenesis in gastric carcinoma. J. Biomed. Sci. 2004, 11, 517–527. [Google Scholar] [CrossRef]
- Adachi, Y.; Aoki, C.; Yoshio-Hoshino, N.; Takayama, K.; Curiel, D.T.; Nishimoto, N. Interleukin-6 induces both cell growth and VEGF production in malignant mesotheliomas. Int. J. Cancer 2006, 119, 1303–1311. [Google Scholar] [CrossRef]
- Garbers, C.; Kuck, F.; Aparicio-Siegmund, S.; Konzak, K.; Kessenbrock, M.; Sommerfeld, A.; Haussinger, D.; Lang, P.A.; Brenner, D.; Mak, T.W.; et al. Cellular senescence or EGFR signaling induces Interleukin 6 (IL-6) receptor expression controlled by mammalian target of rapamycin (mTOR). Cell Cycle 2013, 12, 3421–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnke, E.; Pietsch, J.; Kopp, S.; Bauer, J.; Sahana, J.; Wehland, M.; Krüger, M.; Hemmersbach, R.; Infanger, M.; Lutzenberg, R.; et al. Cytokine Release and Focal Adhesion Proteins in Normal Thyroid Cells Cultured on the Random Positioning Machine. Cell. Physiol. Biochem. 2017, 43, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 12. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | LLOQ (pg/mL) | 5-Day µg (pg/mL) | 10-Day µg (pg/mL) | 5-Day 1g (pg/mL) | 10-Day 1g (pg/mL) | p | FDR q | Type |
|---|---|---|---|---|---|---|---|---|
| Angiopoietin-2 (Ang-2) | 33 | 1333.3 (3, 115.5) | 1150 (2, 212.1) | 1266.7 (3, 230.9) | 430 (2, 70.7) | 0.0054 | 0.097 | MAP |
| Carbonic anhydrase 9 (CA-9) | 4.2 | 64.3 (3, 32.5) | 40 (2, 21.2) | 18.3 (3, 1.53) | 17 (2, 4.24) | 0.11 | 0.64 | MAP |
| Collagen I alpha I (BR18) | --- | 1934.5 (3, 1281.4) | 1408.4 (2, 1748.8) | 1264.1 (3, 358.3) | 53.86 (2, 2.74) | 0.36 | 0.66 | ELISA |
| Epidermal Growth Factor (EGF) | 0.74 | 0.5 (1, 0.23) | --- | --- | 1 (2, 0) | --- | --- | MAP |
| Epidermal Growth Factor Receptor (EGFR) | 16 | 32.3 (3, 6.11) | 31 (2, 5.66) | 21.7 (3, 8.08) | 23.5 (2, 2.12) | 0.23 | 0.66 | MAP |
| Fatty Acid-Binding Protein, adipocyte (FABP, adipocyte) | 30 | 87 (3, 23) | 69.5 (2, 33.2) | --- | --- | --- | --- | MAP |
| Fibronectin (BR56) | --- | 606,423.3 (3, 417,312.8) | 412,019.5 (2, 521,272.7) | 200,447 (3, 56,096.2) | 105,046.5 (2, 22,128.9) | 0.37 | 0.66 | ELISA |
| Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) | 5.9 | 9.17 (3, 2.63) | 6.33 (1, 4.77) | --- | 5.38 (1, 3.43) | --- | --- | MAP |
| Heparin-Binding EGF-Like Growth Factor (HB-EGF) | 0.7 | 1.43 (3, 0.31) | 1.15 (2, 0.64) | 1.3 (3, 0.4) | 1.5 (2, 0.71) | 0.88 | 0.88 | MAP |
| Hepatocyte Growth Factor receptor (HGF receptor) | 52 | 2110 (3, 1358.4) | 1430 (2, 1654.6) | 1240 (3, 341.8) | 1150 (2, 212.1) | 0.72 | 0.88 | MAP |
| Insulin-like Growth Factor-Binding Protein 1 (IGFBP-1) | 120 | 3600 (3, 556.8) | 4700 (2, 1697.1) | 3193.3 (3, 2174.4) | 3110 (2, 3804.2) | 0.86 | 0.88 | MAP |
| Intercellular Adhesion Molecule 1 (ICAM-1) | 570 | 396.7 (1, 193.4) | 452.5 (1, 236.9) | 443.3 (1, 274.2) | 845 (2, 120.2) | --- | --- | MAP |
| Interleukin-6 (IL-6) | 0.81 | 128 (3, 146.6) | 61.5 (2, 2.12) | 65.3 (3, 25) | 71.5 (2, 23.3) | 0.78 | 0.88 | MAP |
| Interleukin-8 (IL-8) | 0.63 | 6890 (3, 7913.4) | 2505 (2, 1322.3) | 2380 (3, 1796.4) | 5180 (2, 2616.3) | 0.66 | 0.88 | MAP |
| Laminin | --- | 527.67 (3, 240.19) | 525 (2, 237.59) | 306.33 (3, 225.019) | 13,393 (2, 10,875.3) | 0.055 | 0.497 | ELISA |
| Lipocalin-2/NGAL (BR53) | --- | 251.85 (3, 17.2) | 243.3 (2, 12.36) | 243.32 (3, 8.74) | 243.3 (2, 12.36) | 0.83 | 0.88 | ELISA |
| Macrophage Inflammatory Protein-1 beta (MIP-1 beta) | 8.2 | 7.07 (1, 5.14) | --- | 6.07 (1, 3.4) | 13.5 (2, 4.9) | --- | --- | MAP |
| Macrophage Migration Inhibitory Factor (MIF) | 2.1 | 1560 (3, 561.1) | 1300 (2, 141.4) | 850 (3, 130.8) | 1000 (2, 282.8) | 0.18 | 0.66 | MAP |
| Matrix Metalloproteinase-3 (MMP-3) | 10 | 154 (3, 104.9) | 90.5 (2, 98.3) | 115 (3, 27.8) | 94 (2, 7.07) | 0.76 | 0.88 | MAP |
| Monocyte Chemotactic Protein 1 (MCP-1) | 27 | 30.5 (2, 14.8) | 22.3 (1, 12.4) | 33.3 (3, 2.52) | 39 (2, 8.49) | --- | --- | MAP |
| Osteopontin (BR54) | --- | 8524.3 (3, 197.3) | 8412.0 (2, 264.5) | 8399.7 (3, 188.2) | 8896 (2, 420.0) | 0.26 | 0.66 | ELISA |
| Placenta Growth Factor (PLGF) | 12 | 13 (1, 12.1) | --- | --- | --- | --- | --- | MAP |
| Tenascin-C (TN-C) | 1700 | 233,000 (3, 157,515.1) | 155,800 (2, 211,000.7) | 62,666.7 (3, 17,897.9) | 52,000 (2, 1414.2) | 0.37 | 0.66 | MAP |
| Urokinase-type plasminogen activator receptor (uPAR) | 250 | 883.3 (3, 281.8) | 662.5 (1, 760.1) | 793.3 (3, 83.3) | 885 (2, 7.07) | --- | --- | MAP |
| VEGF-D (BR73) | --- | 137.74 (3, 3.52) | 132.45 (2, 2.51) | 135.4 (3, 2.038) | 135.99 (2, 2.5) | 0.31 | 0.66 | ELISA |
| Gene | Primer Name | Sequence |
|---|---|---|
| 18S rRNA | 18S-F | GGAGCCTGCGGCTTAATTT |
| 18S-R | CAACTAAGAACGGCCATGCA | |
| CAV1 | CAV1-F | CCTCCTCACAGTTTCATCCA |
| CAV1-R | TGTAGATGTTGCCCTGTTCC | |
| COL1A1 | Col1A-F | ACGAAGACATCCCACCAATCAC |
| Col1A-R | CGTTGTCGCAGACGCAGAT | |
| CTGF | CTGF-F | ACAAGGGCCTCTTCTGTGACTT |
| CTGF-R | GGTACACCGTACCACCGAAGAT | |
| EGF | EGF-F | TGCCAGCTGCACAAATACAGA |
| EGF-R | TCTTACGGAATAGTGGTGGTCATC | |
| EGFR | EGFR-F | TTGCCGCAAAGTGTGTAACG |
| EGFR-R | GAGATCGCCACTGATGGAGG | |
| ERK1 | ERK1-F | ACCTGCGACCTTAAGATTTGTGA |
| ERK1-R | AGCCACATACTCCGTCAGGAA | |
| ERK2 | ERK2-F | TTCCAACCTGCTGCTCAACA |
| ERK2-R | TCTGTCAGGAACCCTGTGTGAT | |
| ICAM1 | ICAM1-F | CGGCTGACGTGTGCAGTAAT |
| ICAM1-R | CTTCTGAGACCTCTGGCTTCGT | |
| IL6 | IL6-F | CGGGAACGAAAGAGAAGCTCTA |
| IL6-R | GAGCAGCCCCAGGGAGAA | |
| CXCL8 | IL8-F | TGGCAGCCTTCCTGATTTCT |
| IL8-R | GGGTGGAAAGGTTTGGAGTATG | |
| ITGA4 | ITGA4-F | CCAGCTGGGTAGCCCTAATG |
| ITGA4-R | CCTGGCTGTCTGGAAAGTGT | |
| ITGB1 | ITGB1-F | GAAAACAGCGCATATCTGGAAATT |
| ITGB1-R | CAGCCAATCAGTGATCCACAA | |
| LAMA1 | LAMA1-F | TGGGAATGGCACAGTTGTCA |
| LAMA1-R | AGCCACTCTCCTCTGGGTGTT | |
| MKI67 | MKI67-F | TGGGGAAAGTAGGTGTGAAAGAAG |
| MKI67-R | CTCCTTAAACGTTCTGATGCTCTTG | |
| RELA | NFKBp65-F | CGCTTCTTCACACACTGGATTC |
| NFKBp65-R | ACTGCCGGGATGGCTTCT | |
| PXN | PXN-F | CATGGACGACCTCGACGC |
| PXN-R | CAAGAACACAGGCCGTTTGG | |
| TLN1 | TLN1-F | GATGGCTATTACTCAGTACAGACAACTGA |
| TLN1-R | CATAGTAGACTCCTCATCTCCTTCCA | |
| VCL | VCL-F | GTCTCGGCTGCTCGTATCTT |
| VCL-R | GTCCACCAGCCCTGTCATTT | |
| VEGFD | VEGFD-F | TGCAGGAGGAAAATCCACTTG |
| VEFGD-R | CTCGCAACGATCTTCGTCAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnik, D.; Krüger, M.; Schulz, H.; Kopp, S.; Wehland, M.; Bauer, J.; Baselet, B.; Vermeesen, R.; Baatout, S.; Corydon, T.J.; et al. The CellBox-2 Mission to the International Space Station: Thyroid Cancer Cells in Space. Int. J. Mol. Sci. 2021, 22, 8777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168777
Melnik D, Krüger M, Schulz H, Kopp S, Wehland M, Bauer J, Baselet B, Vermeesen R, Baatout S, Corydon TJ, et al. The CellBox-2 Mission to the International Space Station: Thyroid Cancer Cells in Space. International Journal of Molecular Sciences. 2021; 22(16):8777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168777
Chicago/Turabian StyleMelnik, Daniela, Marcus Krüger, Herbert Schulz, Sascha Kopp, Markus Wehland, Johann Bauer, Bjorn Baselet, Randy Vermeesen, Sarah Baatout, Thomas J. Corydon, and et al. 2021. "The CellBox-2 Mission to the International Space Station: Thyroid Cancer Cells in Space" International Journal of Molecular Sciences 22, no. 16: 8777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168777