
Distinctive Properties of Endothelial Cells from Tumor and Normal Tissue in Human Breast Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
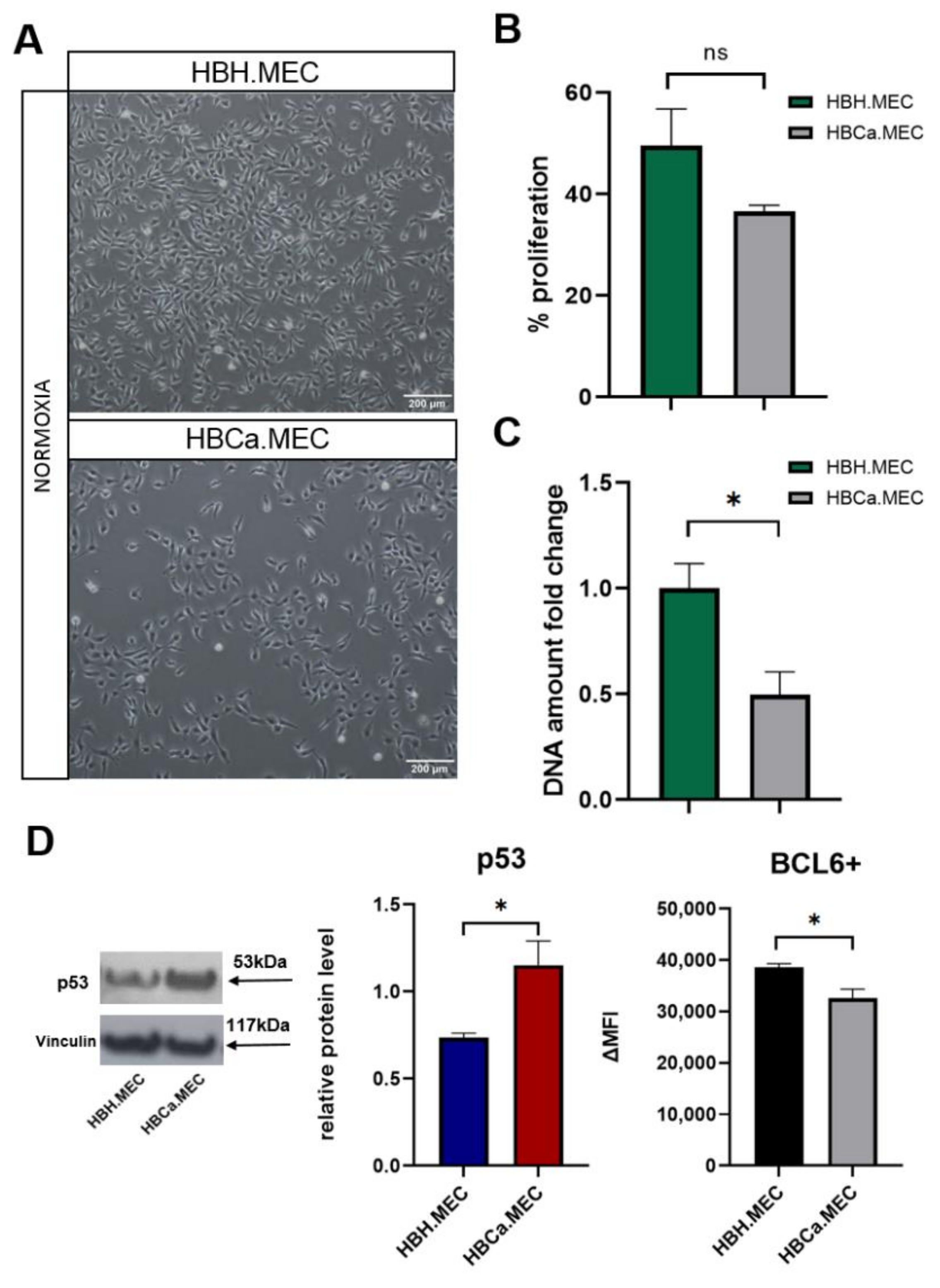
2.1. Tumor Microenvironment Influences EC Morphology, Proliferation and Expression of BCL6/p53
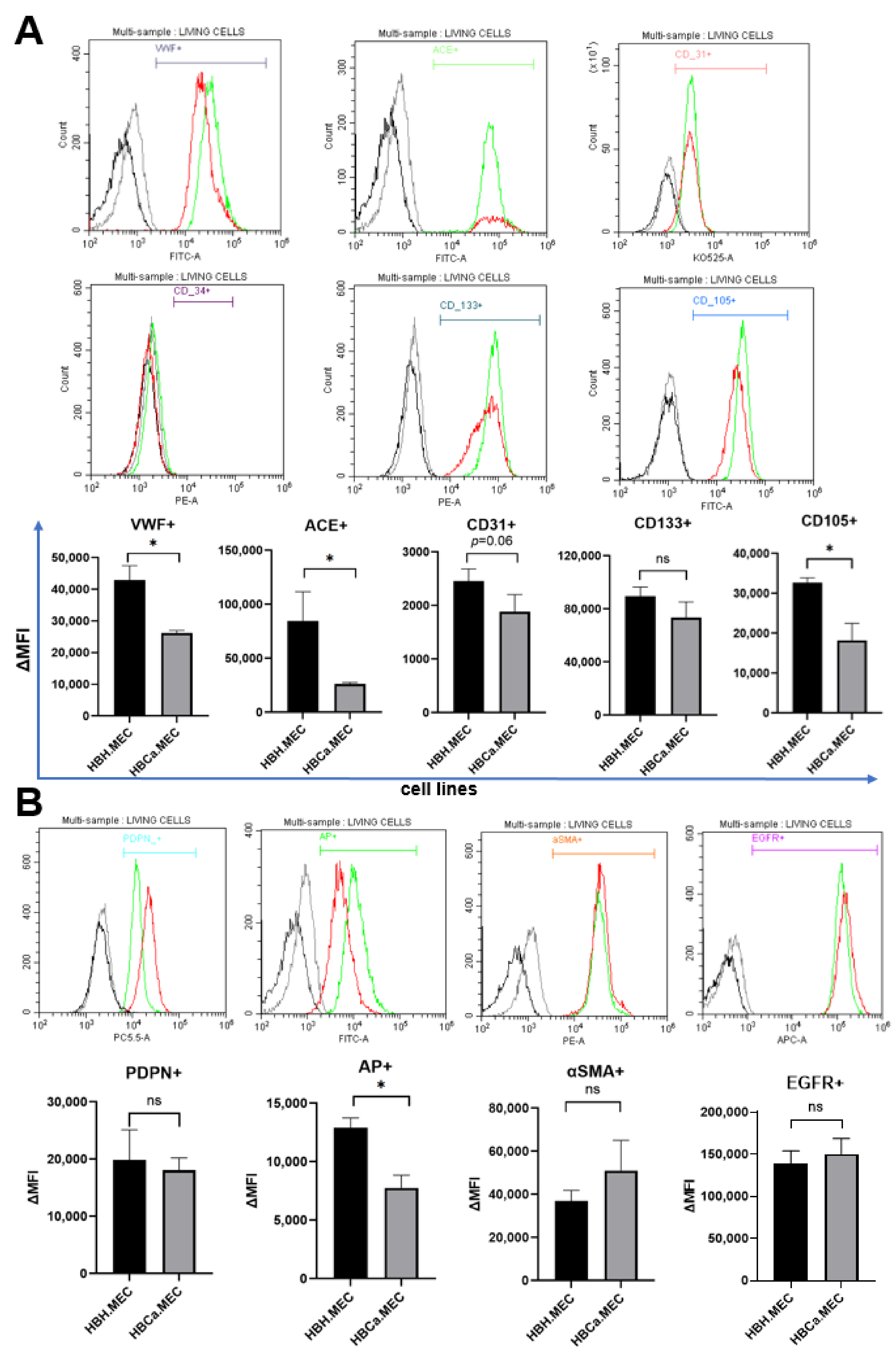
2.2. HBH.MEC and HBCa.MEC Lines Display ECs Phenotype
2.3. Hypoxia Induces the Production of VEGF-A by HBH.MEC and HBCa.MEC
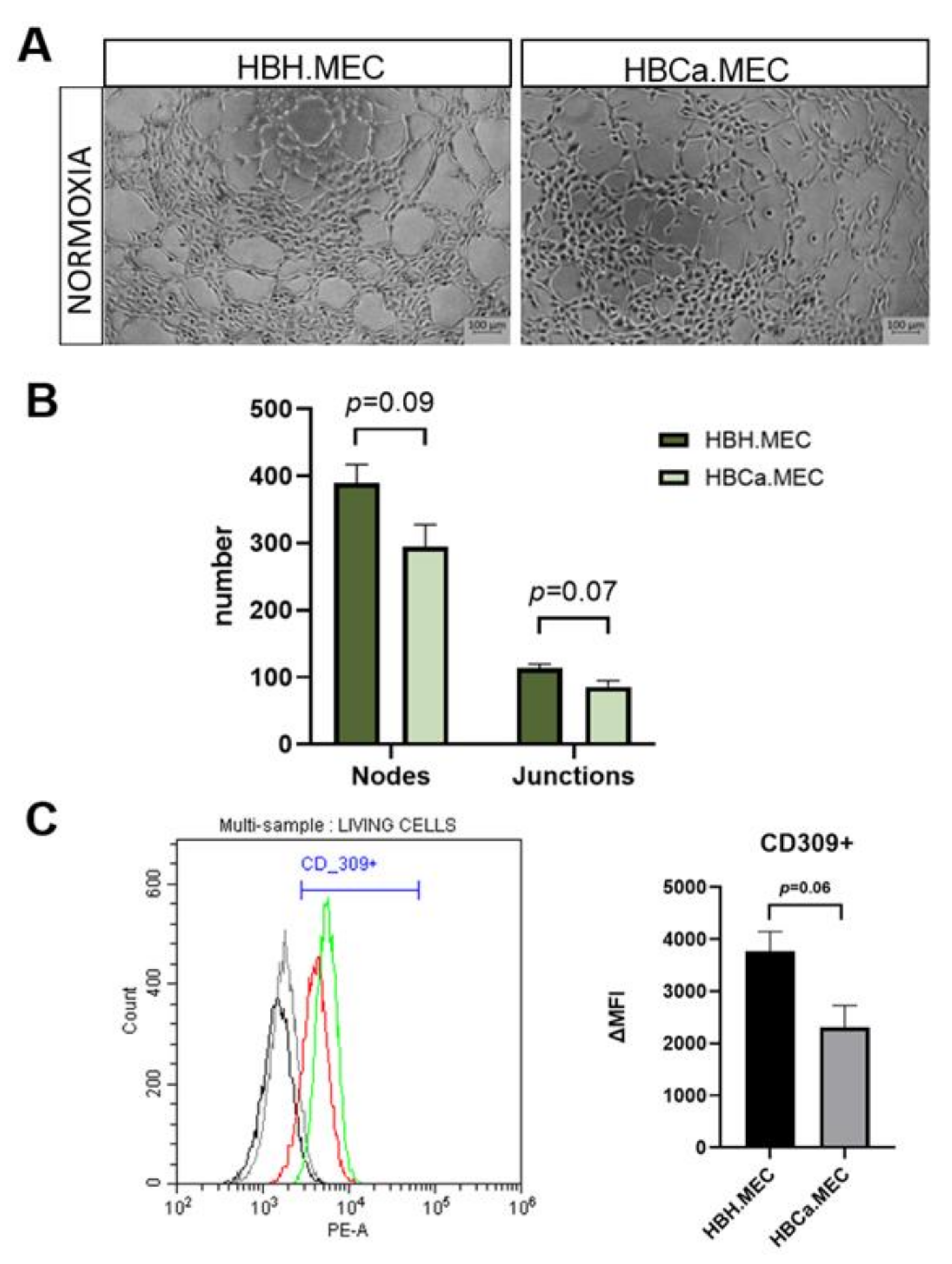
2.4. Proangiogenic Response In Vitro Depends on the Pathological State of ECs
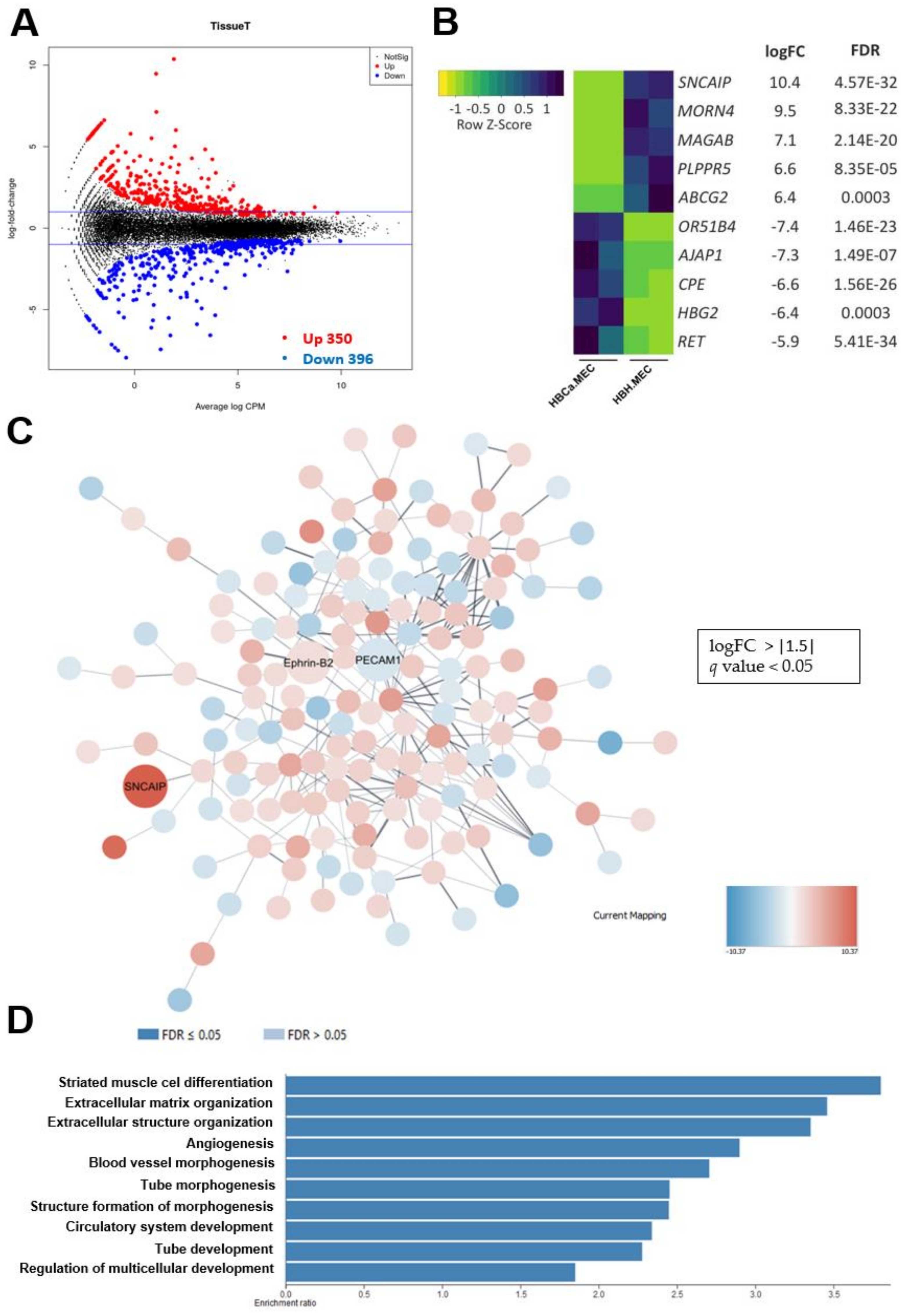
2.5. The Transcriptome Analysis of Breast Tumor-Derived ECs Compared with Healthy ECs Revealed a Distinct Expression Profile Implicated in Vascular Development and Angiogenesis
3. Discussion
4. Materials and Methods
4.1. Endothelial Cell Lines Culture
4.2. Cell Viability Assay
4.3. Flow Cytometry Analysis
4.4. Western Blotting
4.5. Next Generation Sequencing
4.6. Secretion of VEGF-A
4.7. In Vitro Angiogenesis Assay
4.8. Permeability Test
4.9. Cell Proliferation Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeSantis, C.E.; Bray, F.; Ferlay, J.; Lortet-Tieulent, J.; Anderson, B.O.; Jemal, A. International Variation in Female Breast Cancer Incidence and Mortality Rates. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1495–1506. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.A.; Camera-Benson, L.; Palmer-Crocker, R.; Pober, J.S. Variability among human umbilical vein endothelial cultures. Science 1995, 268, 447–448. [Google Scholar] [CrossRef] [Green Version]
- Minshall, R.D.; Malik, A.B. Transport across the Endothelium: Regulation of Endothelial Permeability. In Handbook of Experimental Pharmacology; Springer: Berlin, Germany, 2006; pp. 107–144. [Google Scholar]
- Bassenge, E. Endothelial function in different organs. Prog. Cardiovasc. Dis. 1996, 39, 209–228. [Google Scholar] [CrossRef]
- Hida, K.; Maishi, N.; Annan, D.A.; Hida, Y. Contribution of Tumor Endothelial Cells in Cancer Progression. Int. J. Mol. Sci. 2018, 19, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, J.-T.; Chang, H.Y.; Haraldsen, G.; Jahnsen, F.L.; Troyanskaya, O.G.; Chang, D.S.; Wang, Z.; Rockson, S.G.; van de Rijn, M.; Botstein, D.; et al. Endothelial cell diversity revealed by global expression profiling. Proc. Natl. Acad. Sci. USA 2003, 100, 10623–10628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, E.A.; Hoyer, L.W.; Nachman, R.L. Synthesis of von Willebrand Factor by Cultured Human Endothelial Cells. Proc. Natl. Acad. Sci. USA 1974, 71, 1906–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albelda, S.M.; Muller, W.A.; Buck, C.A.; Newman, P.J. Molecular and cellular properties of PECAM-1 (endoCAM/CD31): A novel vascular cell-cell adhesion molecule. J. Cell Biol. 1991, 114, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Lebrin, F.; Goumans, M.J.; Jonker, L.; Carvalho, R.L.; Valdimarsdottir, G.; Thorikay, M.; Mummery, C.; Arthur, H.M.; ten Dijke, P. Endoglin promotes endothelial cell proliferation and TGF-beta/ALK1 signal transduction. EMBO J. 2004, 23, 4018–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhang, X.; Simons, M. Molecular Controls of Lymphatic VEGFR3 Signaling. Arter. Thromb. Vasc. Biol. 2015, 35, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Gale, N.W.; Baluk, P.; Pan, L.; Kwan, M.; Holash, J.; DeChiara, T.M.; McDonald, D.M.; Yancopoulos, G.D. Ephrin-B2 selectively marks arterial vessels and neovascularization sites in the adult. with expression in both endothelial and smooth-muscle cells. Dev. Biol. 2001, 230, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Makgoba, M.W.; Sanders, M.E.; Ginther Luce, G.E.; Dustin, M.L.; Springer, T.A.; Clark, E.A.; Mannoni, P.; Shaw, S. ICAM-1 a ligand for LFA-1-dependent adhesion of B, T and myeloid cells. Nature 1988, 331, 86–88. [Google Scholar] [CrossRef]
- Elices, M.J.; Osborn, L.; Takada, Y.; Crouse, C.; Luhowskyj, S.; Hemler, M.E.; Lobb, R.R. VCAM-1 on activated endothelium interacts with the leukocyte integrin VLA-4 at a site distinct from the VLA-4/fibronectin binding site. Cell 1990, 60, 577–584. [Google Scholar] [CrossRef]
- Osborn, L.; Hession, C.; Tizard, R.; Vassallo, C.; Luhowskyj, S.; Chi-Rosso, G.; Lobb, R. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell 1989, 59, 1203–1211. [Google Scholar] [CrossRef]
- Bernatchez, P.N.; Soker, S.; Sirois, M.G. Vascular Endothelial Growth Factor Effect on Endothelial Cell Proliferation, Migration, and Platelet-activating Factor Synthesis Is Flk-1-dependent. J. Biol. Chem. 1999, 274, 31047–31054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.F.; Berse, B.; Jackman, R.W.; Tognazzi, K.; Guidi, A.J.; Dvorak, H.F.; Senger, D.R.; Connolly, J.L.; Schnitt, S.J. Expression of vascular permeability factor (vascular endothelial growth factor) and its receptors in breast cancer. Hum. Pathol. 1995, 26, 86–91. [Google Scholar] [CrossRef]
- Johnson, A.R.; Erdös, E.G. Metabolism of vasoactive peptides by human endothelial cells in culture. Angiotensin I converting enzyme (kininase II) and angiotensinase. J. Clin. Investig. 1977, 59, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Grant, D.S.; Lelkes, P.I.; Fukuda, K.; Kleinman, H.K. Intracellular mechanisms involved in basement membrane induced blood vessel differentiation in vitro. In Vitro Cell. Dev. Biol. Anim. 1991, 27, 327–336. [Google Scholar] [CrossRef]
- Lawley, T.J.; Kubota, Y. Induction of Morphologic Differentiation of Endothelial Cells in Culture. J. Investig. Dermatol. 1989, 93, 59S–61S. [Google Scholar] [CrossRef]
- Kieda, C.; Paprocka, M.; Krawczenko, A.; Załęcki, P.; Dupuis, P.; Monsigny, M.; Radzikowski, C.; Duś, D. New Human Microvascular Endothelial Cell Lines with Specific Adhesion Molecules Phenotypes. Endothelium 2002, 9, 247–261. [Google Scholar] [CrossRef]
- Carreau, A.; Kieda, C.; Grillon, C. Nitric oxide modulates the expression of endothelial cell adhesion molecules involved in angiogenesis and leukocyte recruitment. Exp. Cell. Res. 2011, 317, 29–41. [Google Scholar] [CrossRef]
- Majewska, A.; Wilkus, K.; Brodaczewska, K.; Kieda, C. Endothelial Cells as Tools to Model Tissue Microenvironment in Hypoxia-Dependent Pathologies. Int. J. Mol. Sci. 2021, 22, 520. [Google Scholar] [CrossRef]
- Grange, C.; Bussolati, B.; Bruno, S.; Fonsato, V.; Sapino, A.; Camussi, G. Isolation and characterization of human breast tumor-derived endothelial cells. Oncol. Rep. 2006, 15, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.C.; Song, W.; Wang, D.; Zeng, Y.A. Identification of blood vascular endothelial stem cells by the expression of protein C receptor. Cell Res. 2016, 26, 1079–1098. [Google Scholar] [CrossRef] [Green Version]
- Ohga, N.; Ishikawa, S.; Maishi, N.; Akiyama, K.; Hida, Y.; Kawamoto, T.; Sadamoto, Y.; Osawa, T.; Yamamoto, K.; Kondoh, M.; et al. Heterogeneity of tumor endothelial cells: Comparison between tumor endothelial cells isolated from high- and low-metastatic tumors. Am. J. Pathol. 2012, 180, 1294–1307. [Google Scholar] [CrossRef]
- Paprocka, M.; Krawczenko, A.; Dus, D.; Kantor, A.; Carreau, A.; Grillon, C.; Kieda, C. CD133 positive progenitor endothelial cell lines from human cord blood. Cytom. A 2011, 79, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, C.C.; Lamerant-Fayel, N.; Paprocka, M.; Mitterrand, M.; Gosset, D.; Dus, D.; Kieda, C. Selective human endothelial cell activation by chemokines as a guide to cell homing. Immunology 2009, 126, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, H.; Deguchi, M.; Fujimoto, M.; Fukushima, H. Alkaline phosphatase expression in cultured endothelial cells of aorta and brain microvessels: Induction by interleukin-6-type cytokines and suppression by transforming growth factor betas. Life Sci. 1997, 61, 2065–2072. [Google Scholar] [CrossRef]
- Deracinois, B.; Duban-Deweer, S.; Pottiez, G.; Cecchelli, R.; Karamanos, Y.; Flahaut, C. TNAP and EHD1 Are Over-Expressed in Bovine Brain Capillary Endothelial Cells after the Re-Induction of Blood-Brain Barrier Properties. PLoS ONE 2012, 7, e48428. [Google Scholar] [CrossRef] [Green Version]
- Sans, M.; Panés, J.; Ardite, E.; Elizalde, J.; Arce, Y.; Elena, M.; Palacín, A.; Fernández–Checa, J.; Anderson, D.C.; Lobb, R.; et al. VCAM-1 and ICAM-1 mediate leukocyte-endothelial cell adhesion in rat experimental colitis. Gastroenterology 1999, 116, 874–883. [Google Scholar] [CrossRef]
- Rzeniewicz, K.; Newe, A.; Rey Gallardo, A.; Davies, J.; Holt, M.R.; Patel, A.; Charras, G.T.; Stramer, B.; Molenaar, C.; Tedder, T.F.; et al. L-selectin shedding is activated specifically within transmigrating pseudopods of monocytes to regulate cell polarity in vitro. Proc. Natl. Acad. Sci. USA 2015, 112, E1461-70. [Google Scholar] [CrossRef] [Green Version]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-selectin: A Major Regulator of Leukocyte Adhesion. Migration and Signaling. Front. Immunol. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukoji, E.; Yamashita, A.; Nakamura, K.; Hirai, T.; Asada, Y. Podoplanin expression on endothelial cells promotes superficial erosive injury and thrombus formation in rat carotid artery: Implications for plaque erosion. Thromb. Res. 2019, 183, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, K.; Kaneko, M.K.; Kato, Y.; Ishikawa, T.; Nishihira, K.; Tsujimoto, Y.; Shibata, Y.; Ozaki, Y.; Asada, Y. Podoplanin expression in advanced atherosclerotic lesions of human aortas. Thromb. Res. 2012, 129, e70–e76. [Google Scholar] [CrossRef] [PubMed]
- Noren, N.K.; Lu, M.; Freeman, A.L.; Koolpe, M.; Pasquale, E.B. Interplay between EphB4 on tumor cells and vascular ephrin-B2 regulates tumor growth. Proc. Natl. Acad. Sci. USA 2004, 101, 5583–5588. [Google Scholar] [CrossRef] [Green Version]
- Doyle, L.; Ross, D.D. Multidrug resistance mediated by the breast cancer resistance protein BCRP (ABCG2). Oncogene 2003, 22, 7340–7358. [Google Scholar] [CrossRef] [Green Version]
- Higashikuni, Y.; Sainz, J.; Nakamura, K.; Takaoka, M.; Enomoto, S.; Iwata, H.; Sahara, M.; Tanaka, K.; Koibuchi, N.; Ito, S.; et al. The ATP-Binding Cassette Transporter BCRP1/ABCG2 Plays a Pivotal Role in Cardiac Repair After Myocardial Infarction Via Modulation of Microvascular Endothelial Cell Survival and Function. Arter. Thromb. Vasc. Biol. 2010, 30, 2128–2135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Mojsilovic-Petrovic, J.; Andrade, M.F.; Zhang, H.; Ball, M.; Stanimirovic, D.B. The expression and functional characterization of ABCG2 in brain endothelial cells and vessels. FASEB J. 2003, 17, 2085–2087. [Google Scholar] [CrossRef] [Green Version]
- Oosterhoff, L.A.; Kruitwagen, H.S.; Van Wolferen, M.E.; Van Balkom, B.W.; Mokry, M.; Lansu, N.; Dungen, N.A.V.D.; Penning, L.C.; Spanjersberg, T.C.; De Graaf, J.W.; et al. Characterization of Endothelial and Smooth Muscle Cells From Different Canine Vessels. Front. Physiol. 2019, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, S.; Baluk, P.; Kaidoh, T.; Haskell, A.; Jain, R.K.; McDonald, D.M. Abnormalities in Pericytes on Blood Vessels and Endothelial Sprouts in Tumors. Am. J. Pathol. 2002, 160, 985–1000. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Dunn, J.; Dickinson, A.M.; Gillespie, J.I.; Baudouin, S.V. Smooth Muscle α-Actin Expression in Endothelial Cells Derived from CD34+ Human Cord Blood Cells. Stem Cells Dev. 2004, 13, 521–527. [Google Scholar] [CrossRef]
- Spillmann, F.; Miteva, K.; Pieske, B.; Tschope, C.; Van Linthout, S. High-density lipoproteins reduce endothelial-to-mesenchymal transition. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1774–1777. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Brindley, D.N. Lipid Phosphate Phosphatases and Cancer. Biomolecules 2020, 10, 1263. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.P.; Xu, M.; Chen, Y.; Shao, W.W. Expression of MAGE-A11 in breast cancer tissues and its effects on the proliferation of breast cancer cells. Mol. Med. Rep. 2013, 7, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Kuo, W.H.; Lin, C.C.; Oyang, Y.J.; Huang, H.C.; Juan, H.F. MicroRNA-regulated protein-protein interaction networks and their functions in breast cancer. Int. J. Mol. Sci. 2013, 14, 11560–11606. [Google Scholar] [CrossRef] [Green Version]
- Phillips, L.; Gill, A.J.; Baxter, R.C. Novel Prognostic Markers in Triple-Negative Breast Cancer Discovered by MALDI-Mass Spectrometry Imaging. Front. Oncol. 2019, 9, 379. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Matsushita, Y.; Suzuki, H.; Komatsu, M.; Yoshimaru, T.; Kimura, R.; Yanai, A.; Honda, J.; Tangoku, A.; Sasa, M.; et al. Frequent downregulation of LRRC26 by epigenetic alterations is involved in the malignant progression of triple-negative breast cancer. Int. J. Oncol. 2018, 52, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhu, S.; Li, L.; Zhang, L.; Xian, S.; Wang, Y.; Cheng, Y. Identification of differentially expressed genes and signaling pathways in ovarian cancer by integrated bioinformatics analysis. OncoTargets Ther. 2018, 11, 1457–1474. [Google Scholar] [CrossRef] [Green Version]
- Murthy, S.R.; Dupart, E.; Al-Sweel, N.; Chen, A.; Cawley, N.X.; Loh, Y.P. Carboxypeptidase E promotes cancer cell survival, but inhibits migration and invasion. Cancer Lett. 2013, 341, 204–213. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkus, K.; Brodaczewska, K.; Kajdasz, A.; Kieda, C. Distinctive Properties of Endothelial Cells from Tumor and Normal Tissue in Human Breast Cancer. Int. J. Mol. Sci. 2021, 22, 8862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168862
Wilkus K, Brodaczewska K, Kajdasz A, Kieda C. Distinctive Properties of Endothelial Cells from Tumor and Normal Tissue in Human Breast Cancer. International Journal of Molecular Sciences. 2021; 22(16):8862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168862
Chicago/Turabian StyleWilkus, Kinga, Klaudia Brodaczewska, Arkadiusz Kajdasz, and Claudine Kieda. 2021. "Distinctive Properties of Endothelial Cells from Tumor and Normal Tissue in Human Breast Cancer" International Journal of Molecular Sciences 22, no. 16: 8862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168862