
Doxorubicin Paradoxically Ameliorates Tumor-Induced Inflammation in Young Mice

 , , and
, , and
Abstract
:1. Introduction
2. Results
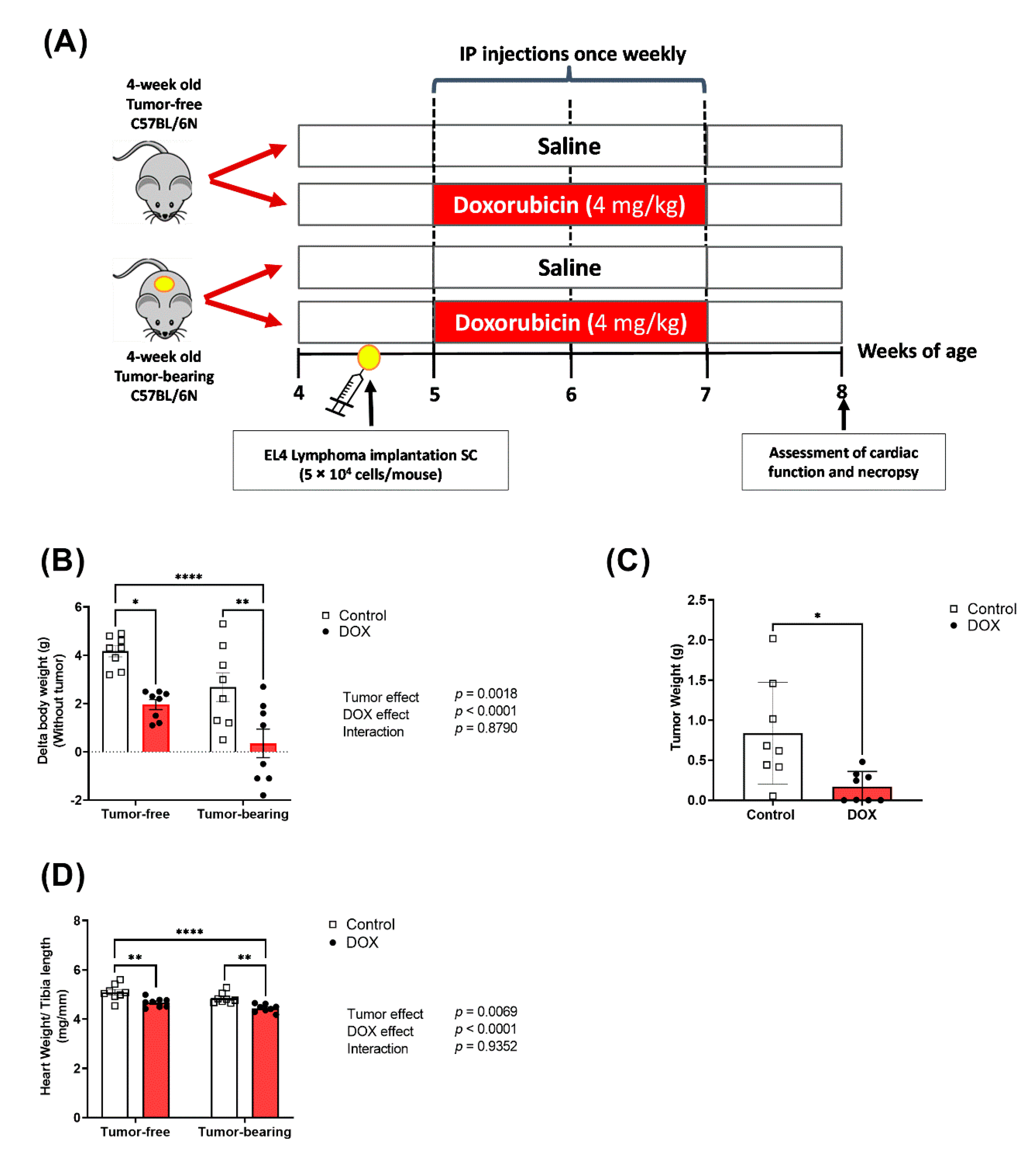
2.1. Effects of DOX on Body Weight and Tumor Growth
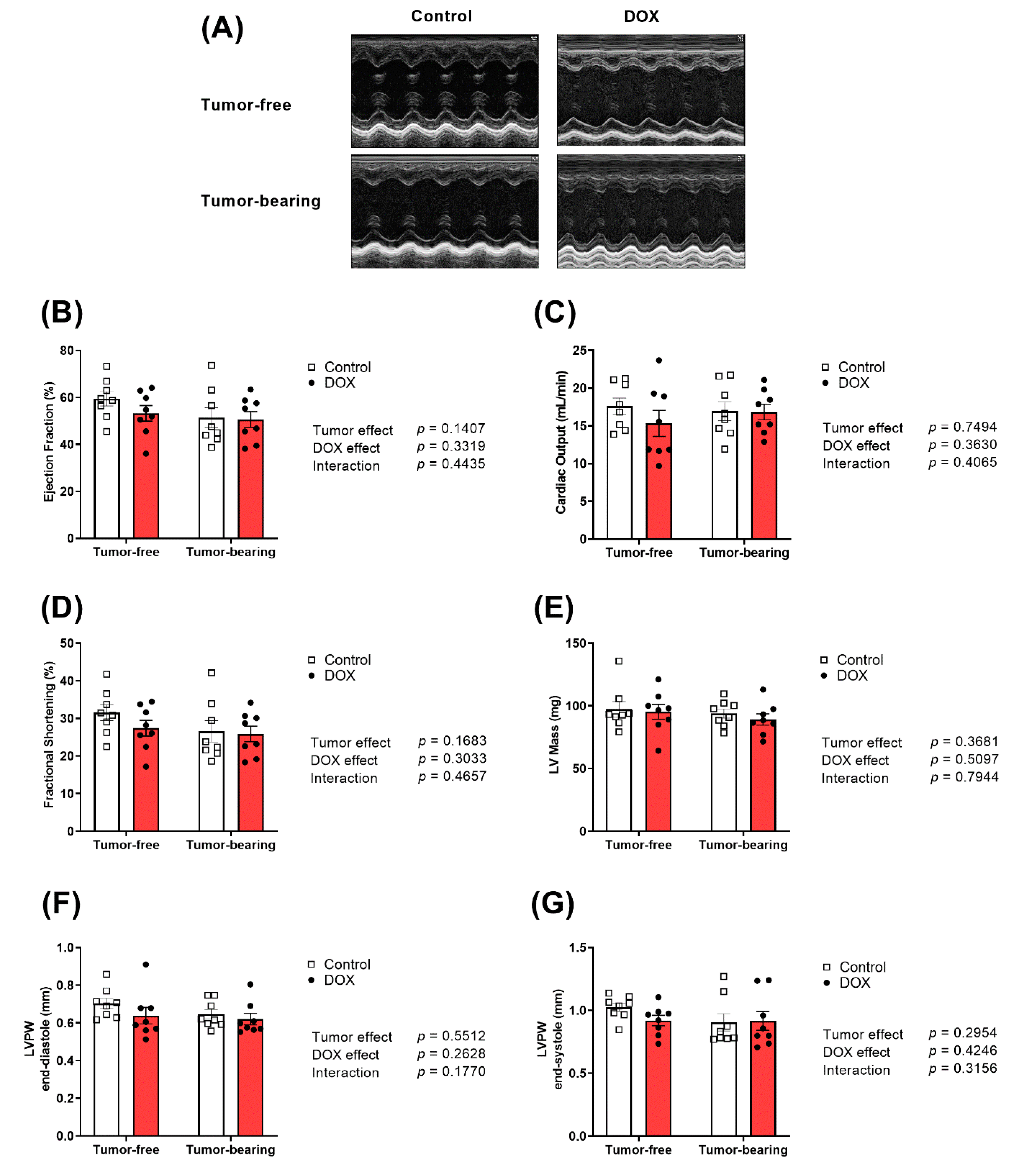
2.2. Effect of DOX Administration on Cardiac Function and Morphometry in Tumor-Free and Tumor-Bearing Mice
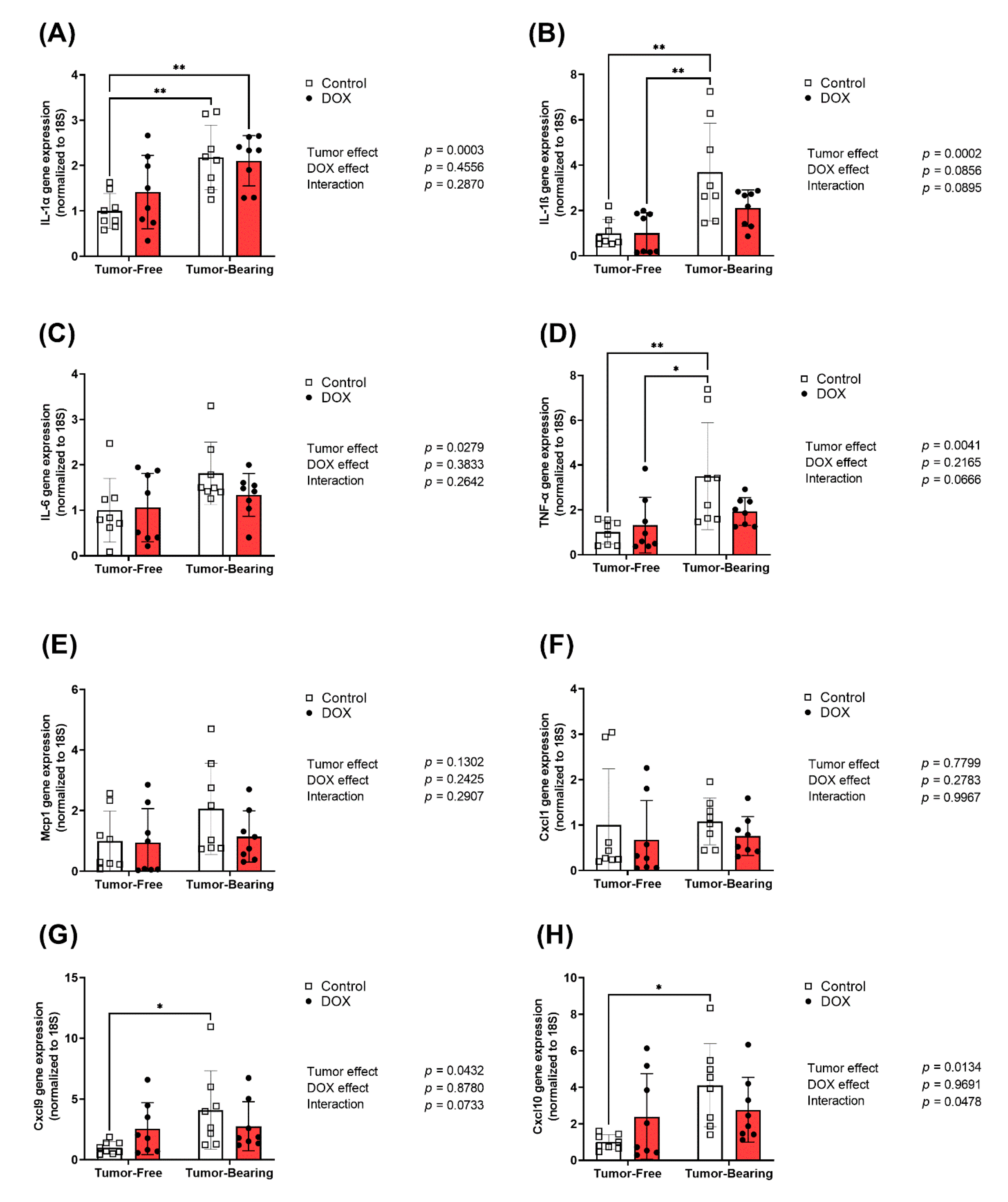
2.3. Effect of DOX Administration on Gene Expression of Inflammatory Markers in the Heart of Tumor-Bearing and Tumor-Free Mice
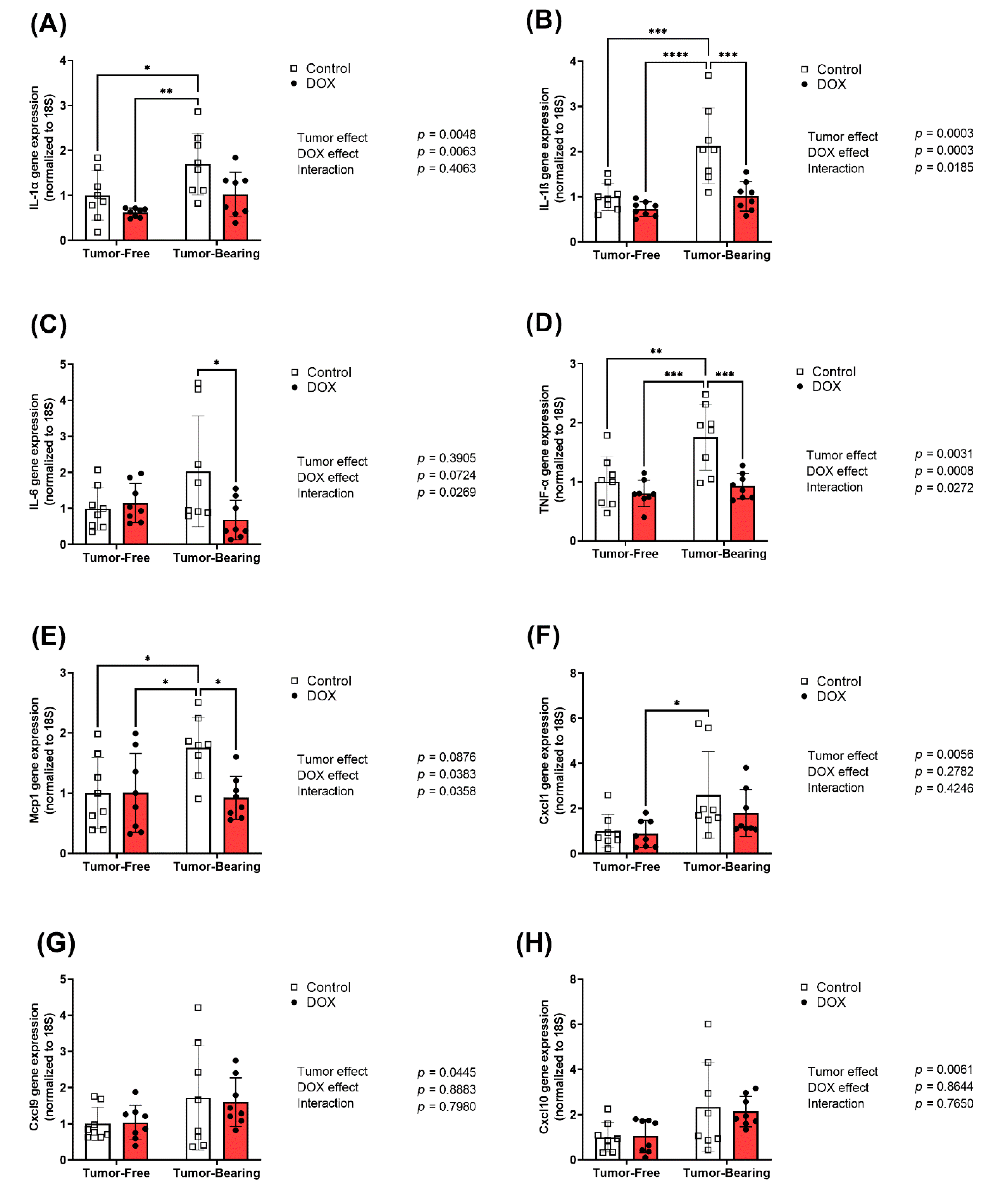
2.4. Effect of DOX Administration on Gene Expression of Inflammatory Markers in the Liver of Tumor-Free and Tumor-Bearing Mice
2.5. Effect of DOX Administration on Gene Expression of Inflammatory Markers in the Kidney of Tumor-Free and Tumor-Bearing Mice
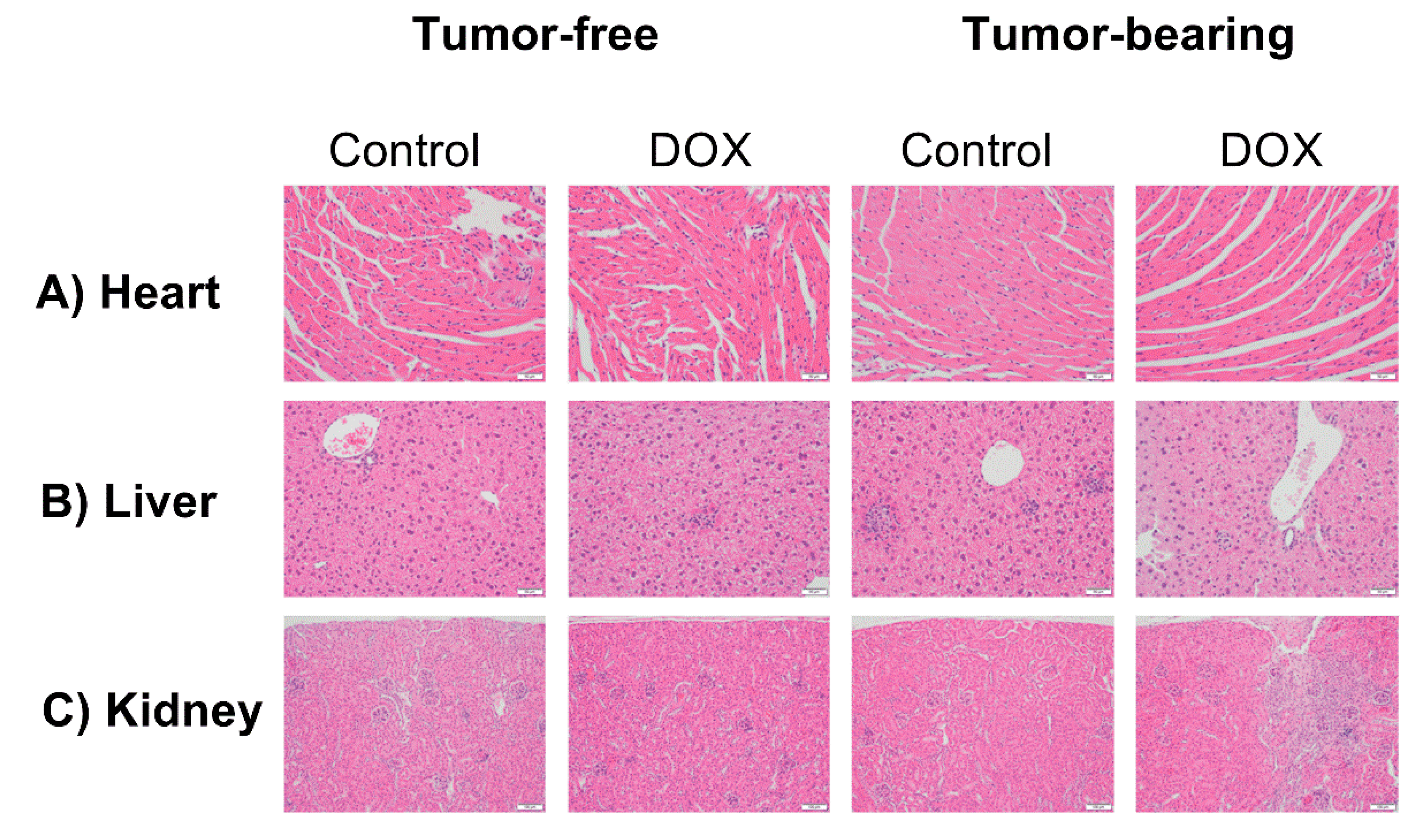
2.6. Effect of DOX Administration on Histopathological Features in the Heart, Liver, and Kidney of Tumor-Free and Tumor-Bearing Mice
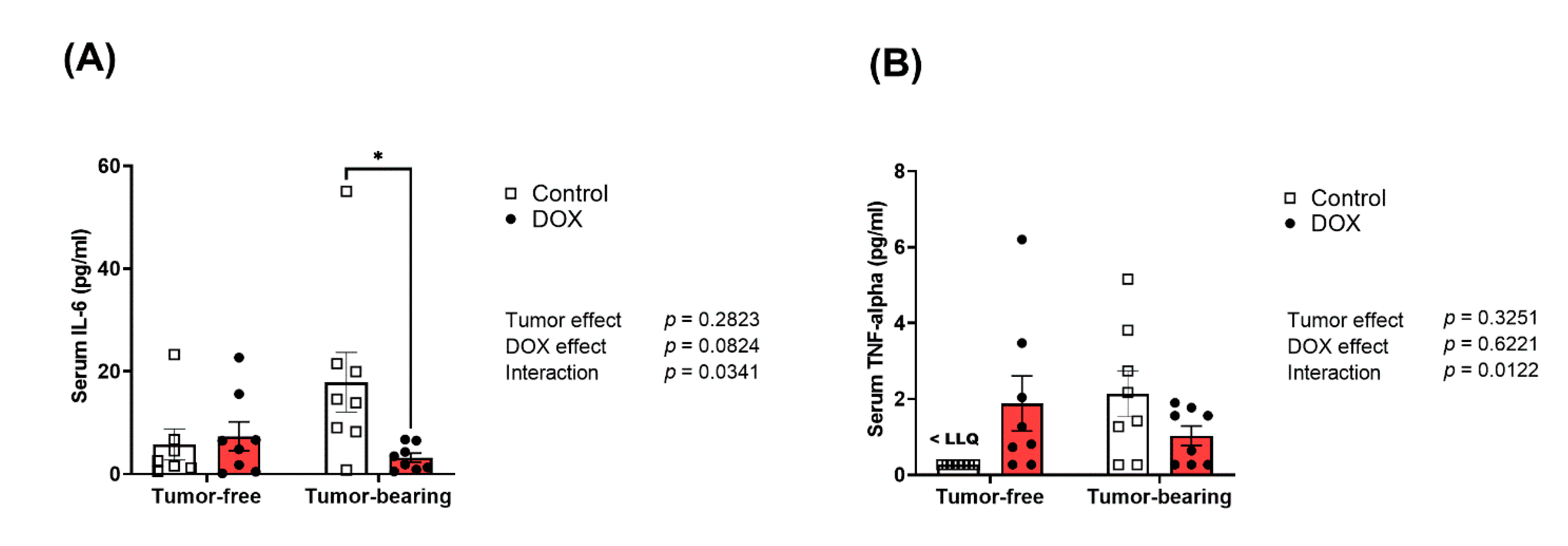
2.7. Divergent Effects of DOX on Serum Inflammatory Cytokines in Tumor-Free Versus Tumor-Bearing Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. EL4 Cell Culture and Reagents
4.3. Echocardiography
4.4. RNA Extraction and Real-Time PCR
4.5. Histopathology
4.6. Measurement of Serum Inflammatory Markers
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chatterjee, K.; Zhang, J.; Honbo, N.; Karliner, J.S. Doxorubicin cardiomyopathy. Cardiology 2010, 115, 155–162. [Google Scholar] [CrossRef]
- Mohamed, N.; Goldstein, J.; Schiff, J.; John, R. Collapsing glomerulopathy following anthracycline therapy. Am. J. Kidney Dis. 2013, 61, 778–781. [Google Scholar] [CrossRef]
- Ibrahim, K.M.; Mantawy, E.M.; Elanany, M.M.; Abdelgawad, H.S.; Khalifa, N.M.; Hussien, R.H.; El-Agroudy, N.N.; El-Demerdash, E. Protection from doxorubicin-induced nephrotoxicity by clindamycin: Novel antioxidant, anti-inflammatory and anti-apoptotic roles. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 739–748. [Google Scholar] [CrossRef]
- Zordoky, B.N.; Radin, M.J.; Heller, L.; Tobias, A.; Matise, I.; Apple, F.S.; McCune, S.A.; Sharkey, L.C. The interplay between genetic background and sexual dimorphism of doxorubicin-induced cardiotoxicity. Cardiooncology 2016, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Zordoky, B.N.; Anwar-Mohamed, A.; Aboutabl, M.E.; El-Kadi, A.O. Acute doxorubicin toxicity differentially alters cytochrome P450 expression and arachidonic acid metabolism in rat kidney and liver. Drug Metab. Dispos. 2011, 39, 1440–1450. [Google Scholar] [CrossRef] [Green Version]
- Bulucu, F.; Ocal, R.; Karadurmus, N.; Sahin, M.; Kenar, L.; Aydin, A.; Oktenli, C.; Koc, B.; Inal, V.; Yamanel, L.; et al. Effects of N-acetylcysteine, deferoxamine and selenium on doxorubicin-induced hepatotoxicity. Biol. Trace Elem. Res. 2009, 132, 184–196. [Google Scholar] [CrossRef]
- van Dalen, E.C.; Raphael, M.F.; Caron, H.N.; Kremer, L.C. Treatment including anthracyclines versus treatment not including anthracyclines for childhood cancer. Cochrane Database Syst. Rev. 2009, CD006647. [Google Scholar] [CrossRef] [Green Version]
- Lipshultz, S.E.; Sambatakos, P.; Maguire, M.; Karnik, R.; Ross, S.W.; Franco, V.I.; Miller, T.L. Cardiotoxicity and cardioprotection in childhood cancer. Acta Haematol. 2014, 132, 391–399. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer Facts & Figures 2021; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- Sauter, K.A.; Wood, L.J.; Wong, J.; Iordanov, M.; Magun, B.E. Doxorubicin and daunorubicin induce processing and release of interleukin-1beta through activation of the NLRP3 inflammasome. Cancer Biol. Ther. 2011, 11, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, M.K.; Abdelgawad, I.Y.; Lewis, C.A.; Zordoky, B.N. Sexual Dimorphism in Doxorubicin-induced Systemic Inflammation: Implications for Hepatic Cytochrome P450 Regulation. Int. J. Mol. Sci. 2020, 21, 1279. [Google Scholar] [CrossRef] [Green Version]
- Maayah, Z.H.; Alam, A.S.; Takahara, S.; Soni, S.; Ferdaoussi, M.; Matsumura, N.; Zordoky, B.N.; Eisenstat, D.D.; Dyck, J.R.B. Resveratrol reduces cardiac NLRP3-inflammasome activation and systemic inflammation to lessen doxorubicin-induced cardiotoxicity in juvenile mice. FEBS Lett. 2021. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, M.; Del Pizzo, M.; Marzocco, S.; Sorrentino, R.; Ciccarelli, M.; Iaccarino, G.; Pinto, A.; Popolo, A. Inflammatory mediators in a short-time mouse model of doxorubicin-induced cardiotoxicity. Toxicol. Appl. Pharmacol. 2016, 293, 44–52. [Google Scholar] [CrossRef]
- Wu, Q.; Li, W.; Zhao, J.; Sun, W.; Yang, Q.; Chen, C.; Xia, P.; Zhu, J.; Zhou, Y.; Huang, G.; et al. Apigenin ameliorates doxorubicin-induced renal injury via inhibition of oxidative stress and inflammation. Biomed. Pharmacother. 2021, 137, 111308. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Chu, L.; Liang, H.; Chen, J.; Liang, J.; Huang, Z.; Zhang, B.; Chen, X. Protective Effects of Dioscin Against Doxorubicin-Induced Hepatotoxicity Via Regulation of Sirt1/FOXO1/NF-κb Signal. Front. Pharmacol. 2019, 10, 1030. [Google Scholar] [CrossRef]
- Maayah, Z.H.; Takahara, S.; Dyck, J.R.B. The beneficial effects of reducing NLRP3 inflammasome activation in the cardiotoxicity and the anti-cancer effects of doxorubicin. Arch. Toxicol. 2021, 95, 1–9. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, J.; Zhang, L.; Du, R.; Xiang, D.; Wu, M.; Zhang, R.; Han, W. Interleukin-1 signaling mediates acute doxorubicin-induced cardiotoxicity. Biomed. Pharmacother. 2011, 65, 481–485. [Google Scholar] [CrossRef]
- Belloum, Y.; Rannou-Bekono, F.; Favier, F.B. Cancer-induced cardiac cachexia: Pathogenesis and impact of physical activity (Review). Oncol. Rep. 2017, 37, 2543–2552. [Google Scholar] [CrossRef]
- Todorova, V.K.; Hsu, P.C.; Wei, J.Y.; Lopez-Candales, A.; Chen, J.Z.; Su, L.J.; Makhoul, I. Biomarkers of inflammation, hypercoagulability and endothelial injury predict early asymptomatic doxorubicin-induced cardiotoxicity in breast cancer patients. Am. J. Cancer Res. 2020, 10, 2933–2945. [Google Scholar]
- Zhu, J.; He, S.; Du, J.; Wang, Z.; Li, W.; Chen, X.; Jiang, W.; Zheng, D.; Jin, G. Local administration of a novel Toll-like receptor 7 agonist in combination with doxorubicin induces durable tumouricidal effects in a murine model of T cell lymphoma. J. Hematol. Oncol. 2015, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches Towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Kelly, K.M. Management of children with high-risk Hodgkin lymphoma. Br. J. Haematol. 2012, 157, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, N.; Zordoky, B.N.; Robertson, I.M.; Hamza, S.M.; Parajuli, N.; Soltys, C.-L.M.; Beker, D.L.; Grant, M.K.; Razzoli, M.; Bartolomucci, A.; et al. Co-administration of resveratrol with doxorubicin in young mice attenuates detrimental late-occurring cardiovascular changes. Cardiovasc. Res. 2018, 114, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Faber, J.; Wingerter, A.; Neu, M.A.; Henninger, N.; Eckerle, S.; Munzel, T.; Lackner, K.J.; Beutel, M.E.; Blettner, M.; Rathmann, W.; et al. Burden of cardiovascular risk factors and cardiovascular disease in childhood cancer survivors: Data from the German CVSS-study. Eur. Heart J. 2018, 39, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Feleszko, W.; Młynarczuk, I.; Bałkowiec-Iskra, E.Z.; Czajka, A.; Świtaj, T.; Stokłosa, T.; Giermasz, A.; Jakóbisiak, M. Lovastatin Potentiates Antitumor Activity and Attenuates Cardiotoxicity of Doxorubicin in Three Tumor Models in Mice. Clin. Cancer Res. 2000, 6, 2044. [Google Scholar]
- Pokrzywinski, K.L.; Biel, T.G.; Rosen, E.T.; Bonanno, J.L.; Aryal, B.; Mascia, F.; Moshkelani, D.; Mog, S.; Rao, V.A. Doxorubicin-induced cardiotoxicity is suppressed by estrous-staged treatment and exogenous 17β-estradiol in female tumor-bearing spontaneously hypertensive rats. Biol. Sex Differ. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Lamas, D.; Nicoud, M.B.; Sterle, H.A.; Carabajal, E.; Tesan, F.; Perazzo, J.C.; Cremaschi, G.A.; Rivera, E.S.; Medina, V.A. Selective cytoprotective effect of histamine on doxorubicin-induced hepatic and cardiac toxicity in animal models. Cell Death Discov. 2015, 1, 15059. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Shou, W.; Payne, R.M.; Caldwell, R.; Field, L.J. A mouse model for juvenile doxorubicin-induced cardiac dysfunction. Pediatr. Res. 2008, 64, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Hayward, R.; Lien, C.Y.; Jensen, B.T.; Hydock, D.S.; Schneider, C.M. Exercise training mitigates anthracycline-induced chronic cardiotoxicity in a juvenile rat model. Pediatr. Blood Cancer 2012, 59, 149–154. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, X.; Ramil, J.M.; Rikka, S.; Kim, L.; Lee, Y.; Gude, N.A.; Thistlethwaite, P.A.; Sussman, M.A.; Gottlieb, R.A.; et al. Juvenile Exposure to Anthracyclines Impairs Cardiac Progenitor Cell Function and Vascularization Resulting in Greater Susceptibility to Stress-Induced Myocardial Injury in Adult Mice. Circulation 2010, 121, 675–683. [Google Scholar] [CrossRef]
- Wang, F.; Iskra, B.; Kleinerman, E.; Alvarez-Florez, C.; Andrews, T.; Shaw, A.; Chandra, J.; Schadler, K.; Aune, G.J. Aerobic Exercise During Early Murine Doxorubicin Exposure Mitigates Cardiac Toxicity. J. Pediatr. Hematol. Oncol. 2018, 40, 208–215. [Google Scholar] [CrossRef]
- Zhang, J.; Knapton, A.; Lipshultz, S.E.; Cochran, T.R.; Hiraragi, H.; Herman, E.H. Sex-related differences in mast cell activity and doxorubicin toxicity: A study in spontaneously hypertensive rats. Toxicol. Pathol. 2014, 42, 361–375. [Google Scholar] [CrossRef]
- Rose-Felker, K.; Border, W.L.; Hong, B.J.; Chow, E.J. Cardio-oncology Related to Heart Failure: Pediatric Considerations for Cardiac Dysfunction. Heart Fail. Clin. 2017, 13, 311–325. [Google Scholar] [CrossRef]
- Chakouri, N.; Farah, C.; Matecki, S.; Amedro, P.; Vincenti, M.; Saumet, L.; Vergely, L.; Sirvent, N.; Lacampagne, A.; Cazorla, O. Screening for in-vivo regional contractile defaults to predict the delayed Doxorubicin Cardiotoxicity in Juvenile Rat. Theranostics 2020, 10, 8130–8142. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Reuter, S.; Field, L.J. Targeted expression of cyclin D2 ameliorates late stage anthracycline cardiotoxicity. Cardiovasc. Res. 2019, 115, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Nagiub, M.; Filippone, S.; Durrant, D.; Das, A.; Kukreja, R.C. Long-acting PDE5 inhibitor tadalafil prevents early doxorubicin-induced left ventricle diastolic dysfunction in juvenile mice: Potential role of cytoskeletal proteins. Can. J. Physiol. Pharmacol. 2017, 95, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhang, W.; Shou, W.; Field, L.J. P53 inhibition exacerbates late-stage anthracycline cardiotoxicity. Cardiovasc. Res. 2014, 103, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Grant, M.; Razzoli, M.; Bartolomucci, A.; Zordoky, B. Psychosocial stress unmasks latent doxorubicin-induced cardiotoxicity. J. Mol. Cell. Cardiol. 2018, 124, 93–94. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Sirova, M.; Kabesova, M.; Kovar, L.; Etrych, T.; Strohalm, J.; Ulbrich, K.; Rihova, B. HPMA Copolymer-Bound Doxorubicin Induces Immunogenic Tumor Cell Death. Curr. Med. Chem. 2013, 20, 4815–4826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, Z.; Ding, H.; Zhou, Y.; Doan, H.A.; Sin, K.W.T.; Zhu, Z.J.; Flores, R.; Wen, Y.; Gong, X.; et al. Tumor induces muscle wasting in mice through releasing extracellular Hsp70 and Hsp90. Nat. Commun. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Tamta, A.K.; Sarikhani, M.; Desingu, P.A.; Kizkekra, S.M.; Pandit, A.S.; Kumar, S.; Khan, D.; Raghavan, S.C.; Sundaresan, N.R. Subcutaneous Ehrlich Ascites Carcinoma mice model for studying cancer-induced cardiomyopathy. Sci. Rep. 2018, 8, 5599. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, B.; Xu, Y.; Meng, S.; Wang, Y.; Jiang, Y. Luteolin reduces cancer-induced skeletal and cardiac muscle atrophy in a Lewis lung cancer mouse model. Oncol. Rep. 2018, 40, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Asp, M.L.; Nishijima, Y.; Belury, M.A. Evidence for cardiac atrophic remodeling in cancer-induced cachexia in mice. Int. J. Oncol. 2011, 39, 1321–1326. [Google Scholar] [CrossRef]
- Lipshultz, S.E.; Lipsitz, S.R.; Sallan, S.E.; Simbre, V.C., 2nd; Shaikh, S.L.; Mone, S.M.; Gelber, R.D.; Colan, S.D. Long-term enalapril therapy for left ventricular dysfunction in doxorubicin-treated survivors of childhood cancer. J. Clin. Oncol. 2002, 20, 4517–4522. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Miller, T.L.; Lipsitz, S.R.; Neuberg, D.S.; Dahlberg, S.E.; Colan, S.D.; Silverman, L.B.; Henkel, J.M.; Franco, V.I.; Cushman, L.L.; et al. Continuous Versus Bolus Infusion of Doxorubicin in Children With ALL: Long-term Cardiac Outcomes. Pediatrics 2012, 130, 1003–1011. [Google Scholar] [CrossRef] [Green Version]
- Lipshultz, S.E.; Lipsitz, S.R.; Kutok, J.L.; Miller, T.L.; Colan, S.D.; Neuberg, D.S.; Stevenson, K.E.; Fleming, M.D.; Sallan, S.E.; Franco, V.I.; et al. Impact of hemochromatosis gene mutations on cardiac status in doxorubicin-treated survivors of childhood high-risk leukemia. Cancer 2013, 119, 3555–3562. [Google Scholar] [CrossRef]
- Mawad, W.; Mertens, L.; Pagano, J.J.; Riesenkampff, E.; Reichert, M.J.E.; Mital, S.; Kantor, P.F.; Greenberg, M.; Liu, P.; Nathan, P.C.; et al. Effect of anthracycline therapy on myocardial function and markers of fibrotic remodelling in childhood cancer survivors. Eur. Heart J. Cardiovasc. Imaging 2020. [Google Scholar] [CrossRef]
- Willis, M.S.; Parry, T.L.; Brown, D.I.; Mota, R.I.; Huang, W.; Beak, J.Y.; Sola, M.; Zhou, C.; Hicks, S.T.; Caughey, M.C.; et al. Doxorubicin Exposure Causes Subacute Cardiac Atrophy Dependent on the Striated Muscle-Specific Ubiquitin Ligase MuRF1. Circ. Heart Fail. 2019, 12, e005234. [Google Scholar] [CrossRef]
- Li, J.; Wang, P.Y.; Long, N.A.; Zhuang, J.; Springer, D.A.; Zou, J.; Lin, Y.; Bleck, C.K.E.; Park, J.H.; Kang, J.G.; et al. p53 prevents doxorubicin cardiotoxicity independently of its prototypical tumor suppressor activities. Proc. Natl. Acad. Sci. USA 2019, 116, 19626–19634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira de Souza, T.; Quinaglia, A.C.S.T.; Osorio Costa, F.; Shah, R.; Neilan, T.G.; Velloso, L.; Nadruz, W.; Brenelli, F.; Sposito, A.C.; Matos-Souza, J.R.; et al. Anthracycline Therapy Is Associated With Cardiomyocyte Atrophy and Preclinical Manifestations of Heart Disease. JACC Cardiovasc. Imaging 2018, 11, 1045–1055. [Google Scholar] [CrossRef]
- Dhingra, R.; Margulets, V.; Chowdhury, S.R.; Thliveris, J.; Jassal, D.; Fernyhough, P.; Dorn, G.W., 2nd; Kirshenbaum, L.A. Bnip3 mediates doxorubicin-induced cardiac myocyte necrosis and mortality through changes in mitochondrial signaling. Proc. Natl. Acad. Sci. USA 2014, 111, E5537–E5544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoli, M.; Robertson, G. Cancer cachexia: Malignant inflammation, tumorkines, and metabolic mayhem. Trends Endocrinol. Metab. 2013, 24, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Cedervall, J.; Zhang, Y.; Huang, H.; Zhang, L.; Femel, J.; Dimberg, A.; Olsson, A.K. Neutrophil Extracellular Traps Accumulate in Peripheral Blood Vessels and Compromise Organ Function in Tumor-Bearing Animals. Cancer Res. 2015, 75, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariffin, H.; Azanan, M.S.; Abd Ghafar, S.S.; Oh, L.; Lau, K.H.; Thirunavakarasu, T.; Sedan, A.; Ibrahim, K.; Chan, A.; Chin, T.F.; et al. Young adult survivors of childhood acute lymphoblastic leukemia show evidence of chronic inflammation and cellular aging. Cancer 2017, 123, 4207–4214. [Google Scholar] [CrossRef] [Green Version]
- Labib, D.; Satriano, A.; Dykstra, S.; Hansen, R.; Mikami, Y.; Guzzardi, D.G.; Slavikova, Z.; Feuchter, P.; Flewitt, J.; Rivest, S.; et al. Effect of Active Cancer on the Cardiac Phenotype: A Cardiac Magnetic Resonance Imaging-Based Study of Myocardial Tissue Health and Deformation in Patients with Chemotherapy-Naïve Cancer. J. Am. Heart Assoc. 2021, 10, e019811. [Google Scholar] [CrossRef]
- Buoncervello, M.; Maccari, S.; Ascione, B.; Gambardella, L.; Marconi, M.; Spada, M.; Macchia, D.; Stati, T.; Patrizio, M.; Malorni, W.; et al. Inflammatory cytokines associated with cancer growth induce mitochondria and cytoskeleton alterations in cardiomyocytes. J. Cell. Physiol. 2019, 234, 20453–20468. [Google Scholar] [CrossRef] [Green Version]
- Eggert, T.; Medina-Echeverz, J.; Kapanadze, T.; Kruhlak, M.J.; Korangy, F.; Greten, T.F. Tumor induced hepatic myeloid derived suppressor cells can cause moderate liver damage. PLoS ONE 2014, 9, e112717. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, Q.; Qi, H.; Wang, C.; Wang, C.; Zhang, J.; Dong, L. Doxorubicin-Induced Systemic Inflammation Is Driven by Upregulation of Toll-Like Receptor TLR4 and Endotoxin Leakage. Cancer Res. 2016, 76, 6631–6642. [Google Scholar] [CrossRef] [Green Version]
- Akbaribazm, M.; Khazaei, M.R.; Khazaei, F.; Khazaei, M. Doxorubicin and Trifolium pratense L. (Red clover) extract synergistically inhibits brain and lung metastases in 4T1 tumor-bearing BALB/c mice. Food Sci. Nutr. 2020, 8, 5557–5570. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.K.O.; Abdelgawad, I.Y.; Lewis, C.A.; Seelig, D.; Zordoky, B.N. Lack of sexual dimorphism in a mouse model of isoproterenol-induced cardiac dysfunction. PLoS ONE 2020, 15, e0232507. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gualda, E.; Baker, A.G.; Fruk, L.; Munoz-Espin, D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2021, 288, 56–80. [Google Scholar] [CrossRef]
- He, C.; Medley, S.C.; Hu, T.; Hinsdale, M.E.; Lupu, F.; Virmani, R.; Olson, L.E. PDGFRbeta signalling regulates local inflammation and synergizes with hypercholesterolaemia to promote atherosclerosis. Nat. Commun. 2015, 6, 7770. [Google Scholar] [CrossRef] [Green Version]
- Falkenham, A.; de Antueno, R.; Rosin, N.; Betsch, D.; Lee, T.D.; Duncan, R.; Legare, J.F. Nonclassical resident macrophages are important determinants in the development of myocardial fibrosis. Am. J. Pathol. 2015, 185, 927–942. [Google Scholar] [CrossRef]
- Nteeba, J.; Ortinau, L.C.; Perfield, J.W., 2nd; Keating, A.F. Diet-induced obesity alters immune cell infiltration and expression of inflammatory cytokine genes in mouse ovarian and peri-ovarian adipose depot tissues. Mol. Reprod. Dev. 2013, 80, 948–958. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Zhao, J.; Bukata, C.; Wade, E.A.; McGowan, S.J.; Angelini, L.A.; Bank, M.P.; Gurkar, A.U.; McGuckian, C.A.; Calubag, M.F.; et al. Tissue specificity of senescent cell accumulation during physiologic and accelerated aging of mice. Aging Cell 2020, 19, e13094. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (3′–5′) | Ref. |
|---|---|---|---|
| Cxcl1 | CTGGGATTCACCTCAAGAACATC | CAGGGTCAAGGCAAGCCTC | [64] |
| Cxcl9 | ATCTTCCTGGAGCAGTGTGGAGTT | AGGGATTTGTAGTGGATCGTGCCT | [65] |
| Cxcl10 | ATATCGATGACGGGCCAGTGAGAA | AATGATCTCAACACGTGGGCAGGA | [65] |
| IL-1α | CGCTTGAGTCGGCAAAGAAAT | TGGCAGAACTGTAGTCTTCGT | [64] |
| IL-1β | TCCTCGGCCAAGACAGGTCGCT | CCCCCACACGTTGACAGCTAGGT | [66] |
| IL-6 | CCAGAGATACAAAGAAATGATGG | ACTCCAGAAGACCAGAGGAAAT | [67] |
| MCP-1 | GCATCCACGTGTTGGCTCA | CTCCAGCCTACTCATTGGGATCA | [68] |
| TNF alpha | CCAGACCCTCACACTCAGATCA | CACTTGGTGGTTTGCTACGAC | [67] |
| r18S | GTAACCCGTTGAACCCCATT | CCATCCAATCGGTAGTAGCG | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelgawad, I.Y.; Grant, M.K.O.; Popescu, F.E.; Largaespada, D.A.; Zordoky, B.N. Doxorubicin Paradoxically Ameliorates Tumor-Induced Inflammation in Young Mice. Int. J. Mol. Sci. 2021, 22, 9023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169023
Abdelgawad IY, Grant MKO, Popescu FE, Largaespada DA, Zordoky BN. Doxorubicin Paradoxically Ameliorates Tumor-Induced Inflammation in Young Mice. International Journal of Molecular Sciences. 2021; 22(16):9023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169023
Chicago/Turabian StyleAbdelgawad, Ibrahim Y., Marianne K. O. Grant, Flavia E. Popescu, David A. Largaespada, and Beshay N. Zordoky. 2021. "Doxorubicin Paradoxically Ameliorates Tumor-Induced Inflammation in Young Mice" International Journal of Molecular Sciences 22, no. 16: 9023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169023