
Multiplication of ampC upon Exposure to a Beta-Lactam Antibiotic Results in a Transferable Transposon in Escherichia coli

 , and
, and
Abstract
:1. Introduction
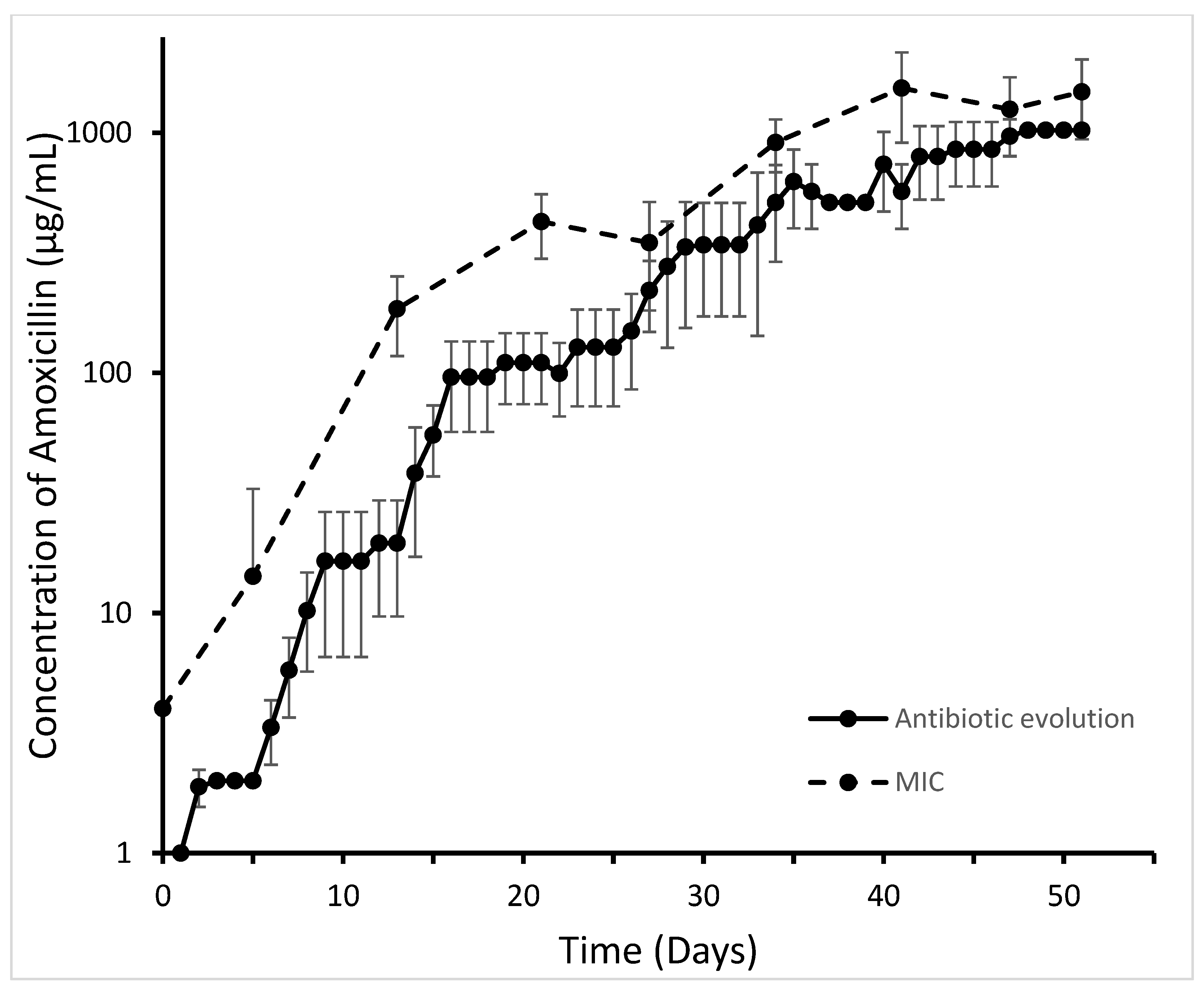
2. Results
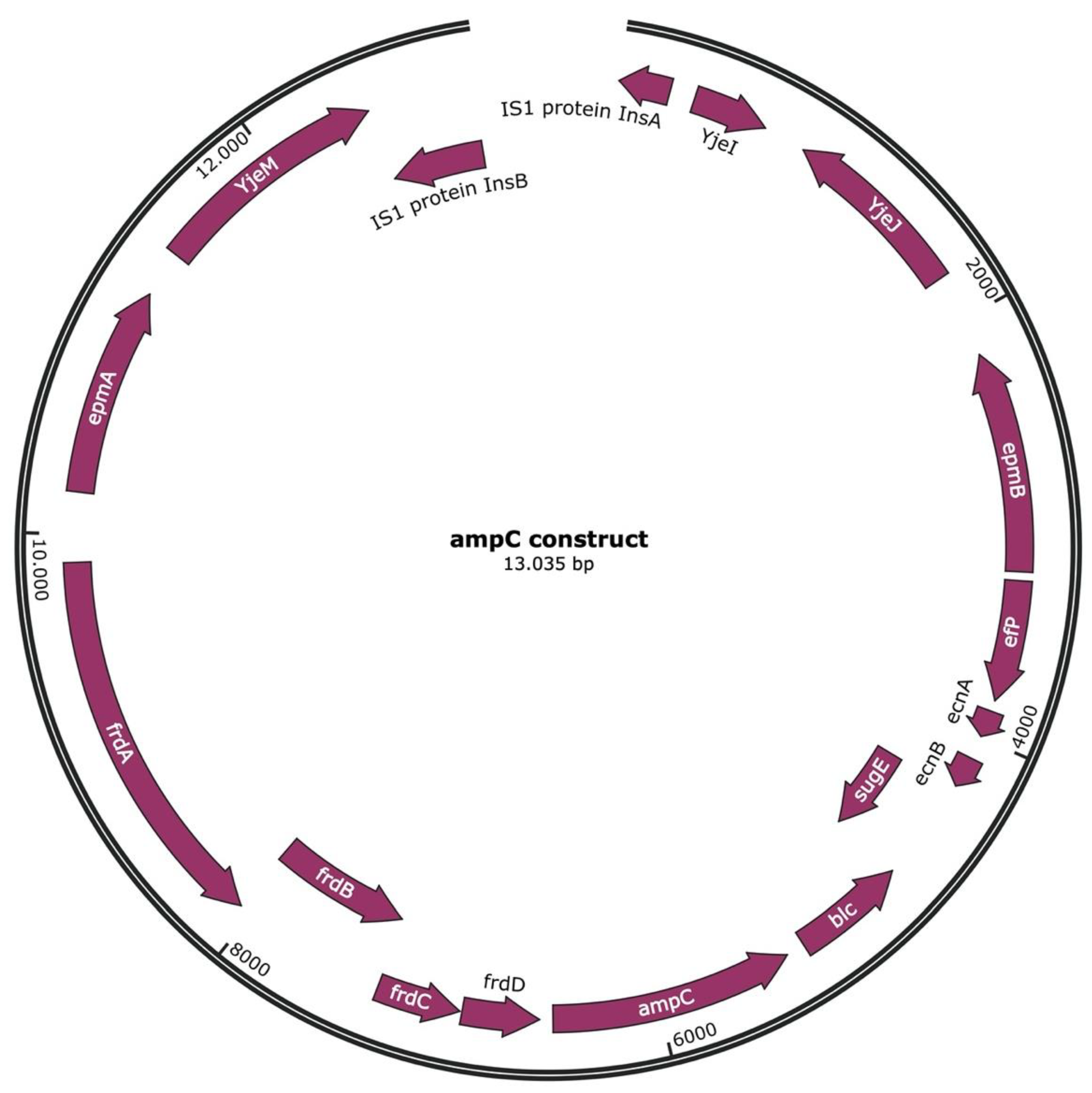
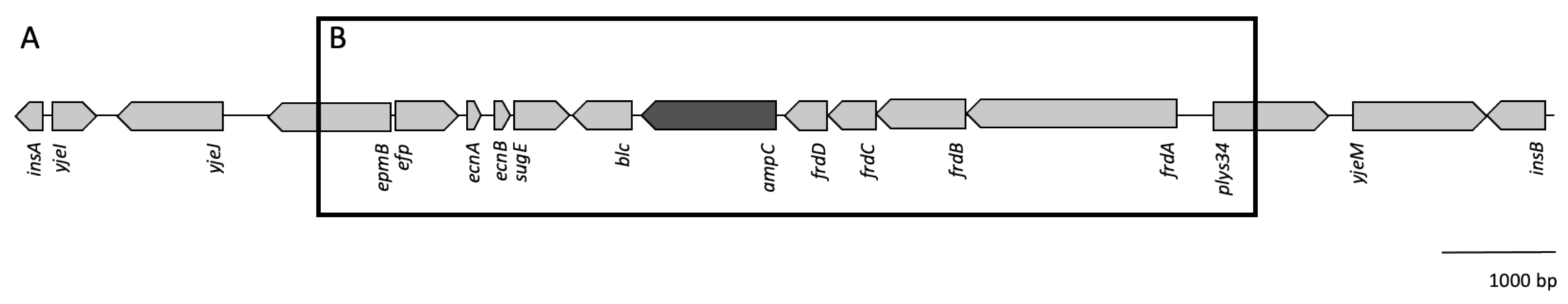
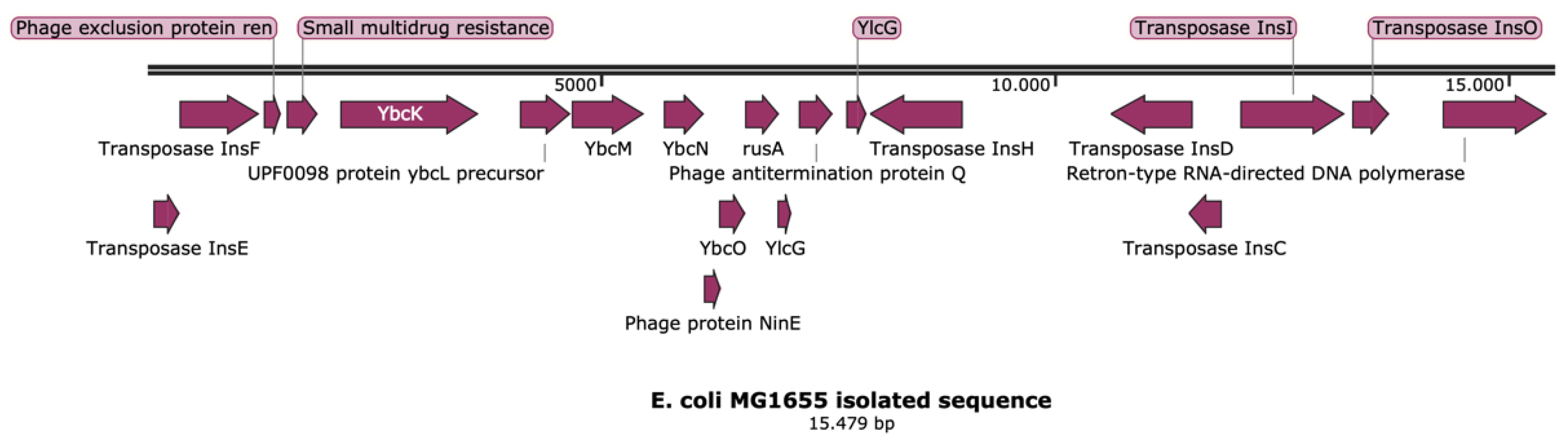
2.1. Transposon Containing ampC
2.2. Transfer of the ampC Transposon
3. Discussion
4. Material and Methods
4.1. Bacterial Strains, Evolution Experiment, and MICs
4.2. Transfer Experiments
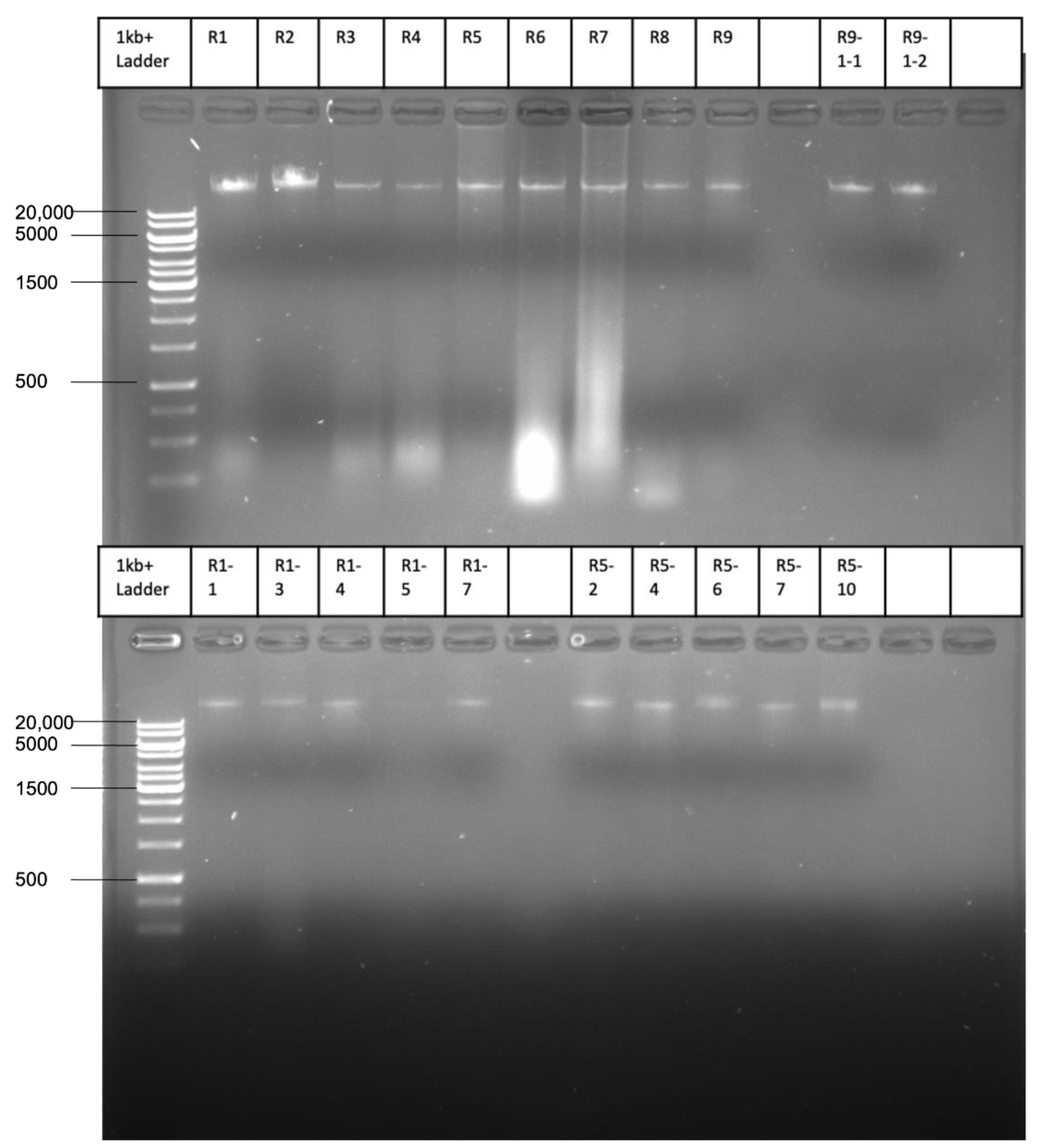
4.3. DNA Isolation, Sequencing, and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, M.; Jonker, M.J.; Bel, K.; Brul, S.; Ter Kuile, B.H. Genome rearrangements in Escherichia coli during de novo acquisition of resistance to a single antibiotic or two antibiotics successively. BMC Genom. 2018, 19, 973. [Google Scholar] [CrossRef]
- Barlow, M.; Hall, B.G. Origin and evolution of the AmpC beta-lactamases of Citrobacter freundii. Antimicrob. Agents Chemother. 2002, 46, 1190–1198. [Google Scholar] [CrossRef] [Green Version]
- Rizi, K.S.; Mosavat, A.; Youssefi, M.; Jamehdar, S.A.; Ghazvini, K.; Safdari, H.; Amini, Y.; Farsiani, H. High prevalence of blaCMY AmpC beta-lactamase in ESBL co-producing Escherichia coli and Klebsiella spp. clinical isolates in the northeast of Iran. J. Glob. Antimicrob. Resist. 2020, 22, 477–482. [Google Scholar] [CrossRef]
- Jacoby, G.A. AmpC beta-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caroff, N.; Espaze, E.; Bérard, I.; Richet, H.; Reynaud, A. Mutations in the ampC promoter of Escherichia coli isolates resistant to oxyiminocephalosporins without extended spectrum B-lactamase production. FEMS Microbiol. Lett. 1999, 173, 459–465. [Google Scholar] [CrossRef]
- Händel, N.; Schuurmans, J.M.; Feng, Y.; Brul, S.; ter Kuile, B.H. Interaction between mutations and regulation of gene expression during development of de novo antibiotic resistance. Antimicrob. Agents Chemother. 2014, 58, 4371–4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeksema, M.; Jonker, M.J.; Brul, S.; Ter Kuile, B.H. Effects of a previously selected antibiotic resistance on mutations acquired during development of a second resistance in Escherichia coli. BMC Genom. 2019, 20, 284. [Google Scholar] [CrossRef] [Green Version]
- Jaurin, B.; Grundström, T.; Normark, S. Sequence elements determining ampC promoter strength in E. coli. EMBO J. 1982, 1, 875–881. [Google Scholar] [CrossRef]
- Tracz, D.M.; Boyd, D.A.; Hizon, R.; Bryce, E.; McGeer, A.; Ofner-Agostini, M.; Simor, A.E.; Paton, S.; Mulvey, M.R.; Canadian Nosocomial Infection Surveillance Program. ampC gene expression in promoter mutants of cefoxitin-resistant Escherichia coli clinical isolates. FEMS Microbiol. Lett. 2007, 270, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Bing, L.; Zhenhua, L. AmpC promoter and attenuator mutations affect function of three Escherichia coli strains. Curr. Microbiol. 2009, 59, 244–247. [Google Scholar] [CrossRef]
- Olsson, O.; Bergström, S.; Lindberg, F.P.; Normark, S. ampC B-lactamase hyperproduction in Escherichia coli: Natural ampicillin resistance generated by horizontal chromosomal DNA transfer from Shigella. Proc. Natl. Acad. Sci. USA 1983, 80, 7556–7560. [Google Scholar] [CrossRef] [Green Version]
- Vadlamani, G.; Thomas, M.D.; Patel, T.R.; Donald, L.J.; Reeve, T.M.; Stetefeld, J.; Standing, K.G.; Vocadlo, D.J.; Mark, B.L. The beta-lactamase gene regulator AmpR is a tetramer that recognizes and binds the D-Ala-D-Ala motif of its repressor UDP-N-acetylmuramic acid (MurNAc)-pentapeptide. J. Biol. Chem. 2015, 290, 2630–2643. [Google Scholar] [CrossRef] [Green Version]
- Edlund, T.; Grundström, T.; Björk, G.R.; Normark, S. Tandem Duplication Induced by an Unusual ampAl-, ampC Transducing Lambda Phage: A Probe to Initiate GeneAmplification. Mol. Gen. Genet. 1980, 180, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Reisbig, M.D.; Hossain, A.; Hanson, N.D. Factors influencing gene expression and resistance for Gram-negative organisms expressing plasmid-encoded ampC genes of Enterobacter origin. J. Antimicrob. Chemother. 2003, 51, 1141–1151. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, M.; Brul, S.; ter Kuile, B.H. Influence of Reactive Oxygen Species on De Novo Acquisition of Resistance to Bactericidal Antibiotics. Antimicrob. Agents Chemother. 2018, 62, e02354-17. [Google Scholar] [CrossRef] [Green Version]
- Darphorn, T.S.; Bel, K.; Koenders-van Sint Anneland, B.B.; Brul, S.; Ter Kuile, B.H. Antibiotic resistance plasmid composition and architecture in Escherichia coli isolates from meat. Sci. Rep. 2021, 11, 2136. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Händel, N.; Otte, S.; Jonker, M.; Brul, S.; ter Kuile, B.H. Factors that affect transfer of the IncI1 beta-lactam resistance plasmid pESBL-283 between E. coli strains. PLoS ONE 2015, 10, e0123039. [Google Scholar] [CrossRef]
- Schuurmans, J.M.; van Hijum, S.A.; Piet, J.R.; Händel, N.; Smelt, J.; Brul, S.; ter Kuile, B.H. Effect of growth rate and selection pressure on rates of transfer of an antibiotic resistance plasmid between E. coli strains. Plasmid 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Sekino, N.; Sekine, Y.; Ohtsubo, E. IS1-encoded proteins, InsA and the InsA-B’-InsB transframe protein (transposase): Functions deduced from their DNA-binding ability. Adv. Biophys. 1995, 31, 209–222. [Google Scholar] [CrossRef]
- Sandegren, L.; Andersson, D.I. Bacterial gene amplification: Implications for the evolution of antibiotic resistance. Nat. Rev. Microbiol. 2009, 7, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Moller, H.D.; Larsen, C.E.; Parsons, L.; Hansen, A.J.; Regenberg, B.; Mourier, T. Formation of Extrachromosomal Circular DNA from Long Terminal Repeats of Retrotransposons in Saccharomyces cerevisiae. G3 (Bethesda) 2015, 6, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Hickman, A.B.; Chandler, M.; Dyda, F. Integrating prokaryotes and eukaryotes: DNA transposases in light of structure. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 50–69. [Google Scholar] [CrossRef] [Green Version]
- Shiga, Y.; Sekine, Y.; Ohtsubo, E. Transposition of ISI circles. Genes Cells 1999, 4, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Harms, K.; Fricke, W.F.; Johnsen, P.J.; da Silva, G.J.; Nielsen, K.M. Natural transformation facilitates transfer of transposons, integrons and gene cassettes between bacterial species. PLoS Pathog. 2012, 8, e1002837. [Google Scholar] [CrossRef] [Green Version]
- Baidya, A.K.; Rosenshine, I.; Ben-Yehuda, S. Donor-delivered cell wall hydrolases facilitate nanotube penetration into recipient bacteria. Nat. Commun. 2020, 11, 1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capy, P.; Gasperi, G.; Biémont, C.; Bazin, C. Stress and transposable elements: Co-evolution or useful parasites? Heredity 2000, 85, 101–106. [Google Scholar] [CrossRef]
- Horvath, V.; Merenciano, M.; Gonzalez, J. Revisiting the Relationship between Transposable Elements and the Eukaryotic Stress Response. Trends Genet. 2017, 33, 832–841. [Google Scholar] [CrossRef]
- Gutierrez, A.; Laureti, L.; Crussard, S.; Abida, H.; Rodriguez-Rojas, A.; Blazquez, J.; Baharoglu, Z.; Mazel, D.; Darfeuille, F.; Vogel, J.; et al. beta-Lactam antibiotics promote bacterial mutagenesis via an RpoS-mediated reduction in replication fidelity. Nat. Commun. 2013, 4, 1610. [Google Scholar] [CrossRef] [Green Version]
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-determined AmpC-type beta-lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Verdet, C.; Gautier, V.; Chachaty, E.; Ronco, E.; Hidri, N.; Decre, D.; Arlet, G. Genetic context of plasmid-carried blaCMY-2-like genes in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 4002–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lartigue, M.F.; Poirel, L.; Aubert, D.; Nordmann, P. In vitro analysis of ISEcp1B-mediated mobilization of naturally occurring beta-lactamase gene blaCTX-M of Kluyvera ascorbata. Antimicrob. Agents Chemother. 2006, 50, 1282–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Lartigue, M.F.; Decousser, J.W.; Nordmann, P. ISEcp1B-Mediated Transposition of blaCTX-M in Escherichia coli. Antimicrob. Agents Chemother. 2005, 49, 447–550. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.X.; Li, X.P.; Li, L.; Chen, M.Y.; Wu, C.Y.; Li, L.L.; Liao, X.P.; Liu, Y.H.; Sun, J. ISEcp1-mediated transposition of chromosome-borne blaCMY-2 into an endogenous ColE1-like plasmid in Escherichia coli. Infect. Drug Resist. 2018, 11, 995–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.G.T.; Herbert, D.; Tempest, D.W. Chapter XIII—The Continuous Cultivation of Micro-organisms. Methods Microbiol. 1970, 2, 277–327. [Google Scholar]
- van der Horst, M.A.; Schuurmans, J.M.; Smid, M.C.; Koenders, B.B.; ter Kuile, B.H. De novo acquisition of resistance to three antibiotics by Escherichia coli. Microb. Drug Resist. 2011, 17, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Schuurmans, J.M.; Nuri Hayali, A.S.; Koenders, B.B.; ter Kuile, B.H. Variations in MIC value caused by differences in experimental protocol. J. Microbiol. Methods 2009, 79, 44–47. [Google Scholar] [CrossRef]
- Elowitz, M.B.; Levine, A.J.; Siggia, E.D.; Swain, P.S. Stochastic Gene Expression in a Single Cell. Science 2002, 297, 1183–1186. [Google Scholar] [CrossRef] [Green Version]
- Nikolenko, S.I.; Korobeynikov, A.I.; Alekseyev, M.A. BayesHammer: Bayesian clustering for error correction in single-cell sequencing. BMC Genom. 2013, 14, S7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boetzer, M.; Henkel, C.V.; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding pre-assembled contigs using SSPACE. Bioinformatics 2011, 27, 578–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boetzer, M.; Pirovano, W. Toward almost closed genomes with GapFiller. Genome Biol. 2012, 13, R56. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Replicate/Strain | Mutations in Promoter/Attenuator Region ampC | Mutations in Other Genes |
|---|---|---|
| 2 | −32, +32 | frdA |
| 3 | −32 | ecnA |
| 4 | −32, +32 | 3× frdA, 1× frdB |
| 5 | −32 | |
| 6 | −32 | 1× before epmB, 1× epmB |
| 7 | −32 | |
| 8 | −32 | 2× before yjeM |
| 9 | −32, +23 | Deletion in frdA |
| R5-6 | −32 | insB |
| R5-10 | −32 | insB, insertion in insA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darphorn, T.S.; Hu, Y.; Koenders-van Sintanneland, B.B.; Brul, S.; ter Kuile, B.H. Multiplication of ampC upon Exposure to a Beta-Lactam Antibiotic Results in a Transferable Transposon in Escherichia coli. Int. J. Mol. Sci. 2021, 22, 9230. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179230
Darphorn TS, Hu Y, Koenders-van Sintanneland BB, Brul S, ter Kuile BH. Multiplication of ampC upon Exposure to a Beta-Lactam Antibiotic Results in a Transferable Transposon in Escherichia coli. International Journal of Molecular Sciences. 2021; 22(17):9230. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179230
Chicago/Turabian StyleDarphorn, Tania S., Yuanqing Hu, Belinda B. Koenders-van Sintanneland, Stanley Brul, and Benno H. ter Kuile. 2021. "Multiplication of ampC upon Exposure to a Beta-Lactam Antibiotic Results in a Transferable Transposon in Escherichia coli" International Journal of Molecular Sciences 22, no. 17: 9230. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179230