
Selection of Endophytic Strains for Enhanced Bacteria-Assisted Phytoremediation of Organic Pollutants Posing a Public Health Hazard

Abstract
:1. Introduction
2. Evidence of Benefits from the Plant-Endophyte Partnership in Proximity of Xenobiotics
2.1. Removal of Hydrocarbons
2.2. Decontamination of Textile Dyes
2.3. Bioremediation of Polyhalogenated Organic Compounds—Biphenyls and Dibenzodioxins
2.4. Removal of Agrochemicals—Pesticides/Herbicides/Insecticides/Fertilizers
2.5. Removal of Pharmaceutical and Personal Care Products (PPCPs)
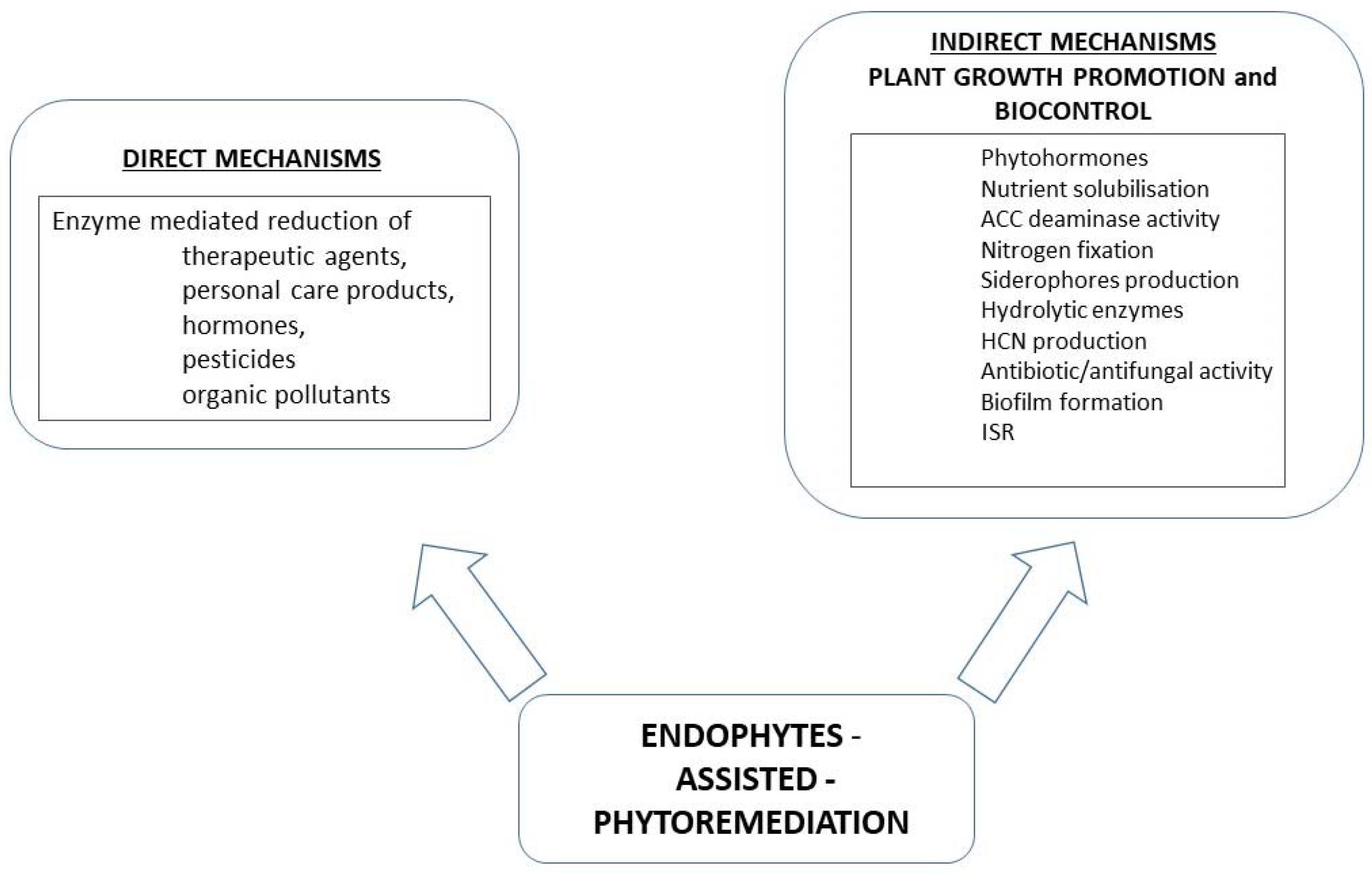
3. Indirect Methods Improving the Efficiency of Bacteria-Assisted Phytoremediation
Mechanisms of Reduction of the Impact of Xenobiotic-Induced Stress Conditions by Bacterial Endophytes
4. Methods of Selecting Endophytic Strains with Biodegradation Potential
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Air Quality in Europe—2020 Report—European Environment Agency. Available online: https://www.eea.europa.eu/publications/air-quality-in-europe-2020-report (accessed on 14 August 2021).
- Ashraf, M.A. Persistent Organic Pollutants (POPs): A Global Issue, a Global Challenge. Environ. Sci. Pollut. Res. Int. 2017, 24, 4223–4227. [Google Scholar] [CrossRef]
- Bartrons, M.; Catalan, J.; Penuelas, J. Spatial and Temporal Trends of Organic Pollutants in Vegetation from Remote and Rural Areas. Sci. Rep. 2016, 6, 25446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui, H.; Wakai, T.; Gion, K.; Kim, Y.-S.; Eun, H. Differential Uptake for Dioxin-like Compounds by Zucchini Subspecies. Chemosphere 2008, 73, 1602–1607. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Feng, Y.; Liu, Y.; Chang, H.; Li, Z.; Xue, J. Uptake and Translocation of Organic Pollutants in Plants: A Review. J. Integr. Agric. 2017, 16, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, A.; Gupta, A.; Sar, P.; Maiti, M.K. Metagenomics of Two Gnotobiotically Grown Aromatic Rice Cultivars Reveals Genotype-Dependent and Tissue-Specific Colonization of Endophytic Bacterial Communities Attributing Multiple Plant Growth Promoting Traits. World J. Microbiol. Biotechnol. 2021, 37, 59. [Google Scholar] [CrossRef]
- Zhu, X.; Ni, X.; Waigi, M.G.; Liu, J.; Sun, K.; Gao, Y. Biodegradation of Mixed PAHs by PAH-Degrading Endophytic Bacteria. Int. J. Environ. Res. Public Health 2016, 13, 805. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant Growth-Promoting Bacterial Endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Sharma, S.; Roy, S. Biodegradation of Dye Reactive Black-5 by a Novel Bacterial Endophyte. Int. Res. J. Environ. Sci. 2015, 4, 44–53. [Google Scholar]
- Siciliano, S.D.; Fortin, N.; Mihoc, A.; Wisse, G.; Labelle, S.; Beaumier, D.; Ouellette, D.; Roy, R.; Whyte, L.G.; Banks, M.K.; et al. Selection of Specific Endophytic Bacterial Genotypes by Plants in Response to Soil Contamination. Appl. Environ. Microbiol. 2001, 67, 2469–2475. [Google Scholar] [CrossRef] [Green Version]
- Taghavi, S.; Barac, T.; Greenberg, B.; Borremans, B.; Vangronsveld, J.; van der Lelie, D. Horizontal Gene Transfer to Endogenous Endophytic Bacteria from Poplar Improves Phytoremediation of Toluene. Appl. Environ. Microbiol. 2005, 71, 8500–8505. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P.; Bae, H. Horizontal Gene Transfer and Endophytes: An Implication for the Acquisition of Novel Traits. Plants 2020, 9, 305. [Google Scholar] [CrossRef] [Green Version]
- Pinski, A.; Betekhtin, A.; Hupert-Kocurek, K.; Mur, L.A.J.; Hasterok, R. Defining the Genetic Basis of Plant−Endophytic Bacteria Interactions. Int. J. Mol. Sci. 2019, 20, 1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant Beneficial Endophytic Bacteria: Mechanisms, Diversity, Host Range and Genetic Determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Anandan, S.; Kumar Ponnusamy, V.; Ashokkumar, M. A Review on Hybrid Techniques for the Degradation of Organic Pollutants in Aqueous Environment. Ultrason. Sonochem. 2020, 67, 105130. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, S.; Sun, K.; Sheng, Y.; Gu, Y.; Gao, Y. Colonization on Root Surface by a Phenanthrene-Degrading Endophytic Bacterium and Its Application for Reducing Plant Phenanthrene Contamination. PLoS ONE 2014, 9, e108249. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Liu, J.; Guo, W.-H.; Li, X.-B.; Xia, J.-B.; Xie, W.-J.; Yao, Z.-G.; Zhang, Y.-M.; Wang, R.-Q. Characterization and Initial Application of Endophytic Bacillus Safensis Strain ZY16 for Improving Phytoremediation of Oil-Contaminated Saline Soils. Front. Microbiol. 2019, 10, 991. [Google Scholar] [CrossRef] [Green Version]
- Andreolli, M.; Lampis, S.; Poli, M.; Gullner, G.; Biró, B.; Vallini, G. Endophytic Burkholderia Fungorum DBT1 Can Improve Phytoremediation Efficiency of Polycyclic Aromatic Hydrocarbons. Chemosphere 2013, 92, 688–694. [Google Scholar] [CrossRef]
- Mitter, E.K.; Kataoka, R.; de Freitas, J.R.; Germida, J.J. Potential Use of Endophytic Root Bacteria and Host Plants to Degrade Hydrocarbons. Int. J. Phytoremediat. 2019, 21, 928–938. [Google Scholar] [CrossRef]
- Bisht, S.; Pandey, P.; Kaur, G.; Aggarwal, H.; Sood, A.; Sharma, S.; Kumar, V.; Bisht, N.S. Utilization of Endophytic Strain Bacillus sp. SBER3 for Biodegradation of Polyaromatic Hydrocarbons (PAH) in Soil Model System. Eur. J. Soil Biol. 2014, 60, 67–76. [Google Scholar] [CrossRef]
- Furtak, K.; Gawryjołek, K.; Gałązka, A.; Grządziel, J. The Response of Red Clover (Trifolium Pratense L.) to Separate and Mixed Inoculations with Rhizobium Leguminosarum and Azospirillum Brasilense in Presence of Polycyclic Aromatic Hydrocarbons. Int. J. Environ. Res. Public Health 2020, 17, 5751. [Google Scholar] [CrossRef]
- Hunt, L.J.; Duca, D.; Dan, T.; Knopper, L.D. Petroleum Hydrocarbon (PHC) Uptake in Plants: A Literature Review. Environ. Pollut. 2019, 245, 472–484. [Google Scholar] [CrossRef]
- Hussein, R.A.; Al-Ghanim, K.; Abd-El-Atty, M.; Mohamed, L. Contamination of Red Sea Shrimp (Palaemon Serratus) with Polycyclic Aromatic Hydrocarbons: A Health Risk Assessment Study. Pol. J. Environ. Stud. 2016, 25, 615–620. [Google Scholar] [CrossRef]
- Pawlik, M.; Cania, B.; Thijs, S.; Vangronsveld, J.; Piotrowska-Seget, Z. Hydrocarbon Degradation Potential and Plant Growth-Promoting Activity of Culturable Endophytic Bacteria of Lotus Corniculatus and Oenothera Biennis from a Long-Term Polluted Site. Environ. Sci. Pollut. Res. 2017, 24, 19640–19652. [Google Scholar] [CrossRef]
- Zhu, X.; Jin, L.; Sun, K.; Li, S.; Li, X.; Ling, W. Phenanthrene and Pyrene Modify the Composition and Structure of the Cultivable Endophytic Bacterial Community in Ryegrass (Lolium Multiflorum Lam). Int. J. Environ. Res. Public Health 2016, 13, 1081. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xiang, Y.; Zhang, Z.; Ling, W.; Gao, Y. Inoculation of a Phenanthrene-Degrading Endophytic Bacterium Reduces the Phenanthrene Level and Alters the Bacterial Community Structure in Wheat. Appl. Microbiol. Biotechnol. 2017, 101, 5199–5212. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas Aeruginosa L10: A Hydrocarbon-Degrading, Biosurfactant-Producing, and Plant-Growth-Promoting Endophytic Bacterium Isolated From a Reed (Phragmites Australis). Front. Microbiol. 2018, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Kukla, M.; Płociniczak, T.; Piotrowska-Seget, Z. Diversity of Endophytic Bacteria in Lolium Perenne and Their Potential to Degrade Petroleum Hydrocarbons and Promote Plant Growth. Chemosphere 2014, 117, 40–46. [Google Scholar] [CrossRef]
- Yousaf, S.; Afzal, M.; Reichenauer, T.G.; Brady, C.L.; Sessitsch, A. Hydrocarbon Degradation, Plant Colonization and Gene Expression of Alkane Degradation Genes by Endophytic Enterobacter Ludwigii Strains. Environ. Pollut. 2011, 159, 2675–2683. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Chen, X.; He, L. Characteristics of an Endophytic Pyrene-Degrading Bacterium of Enterobacter sp. 12J1 from Allium Macrostemon Bunge. Int. Biodeterior. Biodegrad. 2008, 2, 88–95. [Google Scholar] [CrossRef]
- Zhu, X.; Jin, L.; Sun, K.; Li, S.; Ling, W.; Li, X. Potential of Endophytic Bacterium Paenibacillus sp. PHE-3 Isolated from Plantago Asiatica L. for Reduction of PAH Contamination in Plant Tissues. Int. J. Environ. Res. Public Health 2016, 13, 633. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Liu, J.; Gao, Y.; Jin, L.; Gu, Y.; Wang, W. Isolation, Plant Colonization Potential and Phenanthrene Degradation Performance of the Endophytic Bacterium Pseudomonas sp. Ph6-Gfp. Sci. Rep. 2014, 4, 5462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Habteselassie, M.Y.; Liu, J.; Li, S.; Gao, Y. Subcellular Distribution and Biotransformation of Phenanthrene in Pakchoi after Inoculation with Endophytic Pseudomonas Sp. as Probed Using HRMS Coupled with Isotope-Labeling. Environ. Pollut. 2018, 237, 858–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Ma, Z.; Li, S.; Waigi, M.G.; Jiang, J.; Liu, J.; Ling, W. Colonization of Polycyclic Aromatic Hydrocarbon-Degrading Bacteria on Roots Reduces the Risk of PAH Contamination in Vegetables. Environ. Int. 2019, 132, 105081. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, J.; Ling, W.; Huang, Q.; Gao, Y. Composite of PAH-Degrading Endophytic Bacteria Reduces Contamination and Health Risks Caused by PAHs in Vegetables. Sci. Total Environ. 2017, 598, 471–478. [Google Scholar] [CrossRef]
- Davin, M.; Colinet, G.; Fauconnier, M.-L. Targeting the Right Parameters in PAH Remediation Studies. Environ. Pollut. 2021, 278, 116857. [Google Scholar] [CrossRef]
- Pretorius, T.R.; Charest, C.; Kimpe, L.E.; Blais, J.M. The Accumulation of Metals, PAHs and Alkyl PAHs in the Roots of Echinacea Purpurea. PLoS ONE 2018, 13, e0208325. [Google Scholar] [CrossRef]
- Ishii, Y.; Konishi, J.; Okada, H.; Hirasawa, K.; Onaka, T.; Suzuki, M. Operon Structure and Functional Analysis of the Genes Encoding Thermophilic Desulfurizing Enzymes of Paenibacillus sp. A11-2. Biochem. Biophys. Res. Commun. 2000, 270, 81–88. [Google Scholar] [CrossRef]
- Gunam, I.B.W.; Yaku, Y.; Hirano, M.; Yamamura, K.; Tomita, F.; Sone, T.; Asano, K. Biodesulfurization of Alkylated Forms of Dibenzothiophene and Benzothiophene by Sphingomonas Subarctica T7b. J. Biosci. Bioeng. 2006, 101, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khedkar, S.; Shanker, R. Isolation and Classification of a Soil Actinomycete Capable of Sulphur-Specific Biotransformation of Dibenzothiophene, Benzothiophene and Thianthrene. J. Appl. Microbiol. 2015, 118, 62–74. [Google Scholar] [CrossRef]
- Del Olmo, C.H.; Santos, V.E.; Alcon, A.; Garcia-Ochoa, F. Production of a Rhodococcus Erythropolis IGTS8 Biocatalyst for DBT Biodesulfurization: Influence of Operational Conditions. Biochem. Eng. J. 2005, 3, 229–237. [Google Scholar] [CrossRef]
- Kaczorek, E.; Sałek, K.; Guzik, U.; Jesionowski, T.; Cybulski, Z. Biodegradation of Alkyl Derivatives of Aromatic Hydrocarbons and Cell Surface Properties of a Strain of Pseudomonas Stutzeri. Chemosphere 2013, 90, 471–478. [Google Scholar] [CrossRef]
- Lellis, B.; Fávaro-Polonio, C.Z.; Pamphile, J.A.; Polonio, J.C. Effects of Textile Dyes on Health and the Environment and Bioremediation Potential of Living Organisms. Biotechnol. Res. Innov. 2019, 3, 275–290. [Google Scholar] [CrossRef]
- Shang, N.; Ding, M.; Dai, M.; Si, H.; Li, S.; Zhao, G. Biodegradation of Malachite Green by an Endophytic Bacterium Klebsiella Aerogenes S27 Involving a Novel Oxidoreductase. Appl. Microbiol. Biotechnol. 2019, 103, 2141–2153. [Google Scholar] [CrossRef]
- Neetha, J.N.; Sandesh, K.; Girish Kumar, K.; Chidananda, B.; Ujwal, P. Optimization of Direct Blue-14 Dye Degradation by Bacillus Fermus (Kx898362) an Alkaliphilic Plant Endophyte and Assessment of Degraded Metabolite Toxicity. J. Hazard. Mater. 2019, 364, 742–751. [Google Scholar] [CrossRef]
- Zucca, P.; Cocco, G.; Sollai, F.; Sanjust, E. Fungal Laccases as Tools for Biodegradation of Industrial Dyes. Biocatalysis 2016, 1, 82–108. [Google Scholar] [CrossRef] [Green Version]
- Khandare, R.V.; Kabra, A.N.; Awate, A.V.; Govindwar, S.P. Synergistic Degradation of Diazo Dye Direct Red 5B by Portulaca Grandiflora and Pseudomonas Putida. Int. J. Environ. Sci. Technol. 2013, 10, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Shehzadi, M.; Afzal, M.; Khan, M.U.; Islam, E.; Mobin, A.; Anwar, S.; Khan, Q.M. Enhanced Degradation of Textile Effluent in Constructed Wetland System Using Typha Domingensis and Textile Effluent-Degrading Endophytic Bacteria. Water Res. 2014, 58, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, N.; Ali, S.; Shabir, G.; Rizwan, M.; Shakoor, M.B.; Shahid, M.J.; Afzal, M.; Arslan, M.; Hashem, A.; Abd_Allah, E.F.; et al. Bacterial Augmented Floating Treatment Wetlands for Efficient Treatment of Synthetic Textile Dye Wastewater. Sustainability 2020, 12, 3731. [Google Scholar] [CrossRef]
- Riva, V.; Mapelli, F.; Syranidou, E.; Crotti, E.; Choukrallah, R.; Kalogerakis, N.; Borin, S. Root Bacteria Recruited by Phragmites Australis in Constructed Wetlands Have the Potential to Enhance Azo-Dye Phytodepuration. Microorganisms 2019, 7, 384. [Google Scholar] [CrossRef] [Green Version]
- Goud, B.S.; Cha, H.L.; Koyyada, G.; Kim, J.H. Augmented Biodegradation of Textile Azo Dye Effluents by Plant Endophytes: A Sustainable, Eco-Friendly Alternative. Curr. Microbiol. 2020, 77, 3240–3255. [Google Scholar] [CrossRef] [PubMed]
- Ausec, L.; Zakrzewski, M.; Goesmann, A.; Schlüter, A.; Mandic-Mulec, I. Correction: Bioinformatic Analysis Reveals High Diversity of Bacterial Genes for Laccase-Like Enzymes. PLoS ONE 2012, 7, e25724. [Google Scholar] [CrossRef]
- Shi, X.; Liu, Q.; Ma, J.; Liao, H.; Xiong, X.; Zhang, K.; Wang, T.; Liu, X.; Xu, T.; Yuan, S.; et al. An Acid-Stable Bacterial Laccase Identified from the Endophyte Pantoea Ananatis Sd-1 Genome Exhibiting Lignin Degradation and Dye Decolorization Abilities. Biotechnol. Lett. 2015, 37. [Google Scholar] [CrossRef]
- Pawlik, A.; Wójcik, M.; Rułka, K.; Motyl-Gorzel, K.; Osińska-Jaroszuk, M.; Wielbo, J.; Marek-Kozaczuk, M.; Skorupska, A.; Rogalski, J.; Janusz, G. Purification and Characterization of Laccase from Sinorhizobium Meliloti and Analysis of the Lacc Gene. Int. J. Biol. Macromol. 2016, 92, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Suyamud, B.; Thiravetyan, P.; Gadd, G.M.; Panyapinyopol, B.; Inthorn, D. Bisphenol A Removal from a Plastic Industry Wastewater by Dracaena Sanderiana Endophytic Bacteria and Bacillus Cereus NI. Int. J. Phytoremediat. 2020, 22, 167–175. [Google Scholar] [CrossRef]
- Priority Substances and Certain Other Pollutants According to Annex II of Directive 2008/105/EC—Environment—European Commission. Available online: https://ec.europa.eu/environment/water/water-framework/priority_substances.htm (accessed on 14 August 2021).
- Suyamud, B.; Thiravetyan, P.; Panyapinyopol, B.; Inthorn, D. Dracaena Sanderiana Endophytic Bacteria Interactions: Effect of Endophyte Inoculation on Bisphenol A Removal. Ecotoxicol. Environ. Saf. 2018, 157, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.-A.T.; Hsieh, J.-L.; Lo, S.-C.; Wang, S.-Y.; Hung, C.-H.; Huang, E.; Hung, S.-H.; Chin, W.-C.; Huang, C.-C. Biodegradation of Dioxins by Burkholderia Cenocepacia Strain 869T2: Role of 2-Haloacid Dehalogenase. J. Hazard. Mater. 2021, 401, 123347. [Google Scholar] [CrossRef] [PubMed]
- Pileggi, M.; Pileggi, S.A.V.; Sadowsky, M.J. Herbicide Bioremediation: From Strains to Bacterial Communities. Heliyon 2020, 6, e05767. [Google Scholar] [CrossRef]
- Liu, M.; Luo, K.; Wang, Y.; Zeng, A.; Zhou, X.; Luo, F.; Bai, L. Isolation, Identification and Characteristics of an Endophytic Quinclorac Degrading Bacterium Bacillus Megaterium Q3. PLoS ONE 2014, 9, e108012. [Google Scholar] [CrossRef] [Green Version]
- Mesquini, J.A.; Sawaya, A.C.H.F.; López, B.G.C.; Oliveira, V.M.; Miyasaka, N.R.S. Detoxification of Atrazine by Endophytic Streptomyces Sp. Isolated from Sugarcane and Detection of Nontoxic Metabolite. Bull. Environ. Contam. Toxicol. 2015, 95, 803–809. [Google Scholar] [CrossRef]
- Germaine, K.J.; Liu, X.; Cabellos, G.G.; Hogan, J.P.; Ryan, D.; Dowling, D.N. Bacterial Endophyte-Enhanced Phytoremediation of the Organochlorine Herbicide 2,4-Dichlorophenoxyacetic Acid. FEMS Microbiol. Ecol. 2006, 57, 302–310. [Google Scholar] [CrossRef]
- Jabeen, H.; Iqbal, S.; Ahmad, F.; Afzal, M.; Firdous, S. Enhanced Remediation of Chlorpyrifos by Ryegrass (Lolium Multiflorum) and a Chlorpyrifos Degrading Bacterial Endophyte Mezorhizobium sp. HN3. Int. J. Phytoremediat. 2016, 18, 126–133. [Google Scholar] [CrossRef]
- Feng, F.; Li, Y.; Ge, J.; Chen, J.; Jiang, W.; He, S.; Liu, X.; Yu, X. Degradation of Chlorpyrifos by an Endophytic Bacterium of the Sphingomonas Genus (Strain HJY) Isolated from Chinese Chives (Allium Tuberosum). J. Environ. Sci. Health B 2017, 52, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Ge, J.; Li, Y.; Cheng, J.; Zhong, J.; Yu, X. Isolation, Colonization, and Chlorpyrifos Degradation Mediation of the Endophytic Bacterium Sphingomonas Strain HJY in Chinese Chives (Allium Tuberosum). J. Agric. Food Chem. 2017, 65, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, Y.; Zang, P.; Zhao, Y.; He, Z.; Zhu, H.; Song, S.; Zhang, L. Study on the Simultaneous Degradation of Five Pesticides by Paenibacillus Polymyxa from Panax Ginseng and the Characteristics of Their Products. Ecotoxicol. Environ. Saf. 2019, 168, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Ge, J.; Li, Y.; He, S.; Zhong, J.; Liu, X.; Yu, X. Enhanced Degradation of Chlorpyrifos in Rice (Oryza Sativa L.) by Five Strains of Endophytic Bacteria and Their Plant Growth Promotional Ability. Chemosphere 2017, 184, 505–513. [Google Scholar] [CrossRef]
- Barman, D.N.; Haque, M.A.; Islam, S.M.A.; Yun, H.D.; Kim, M.K. Cloning and Expression of OphB Gene Encoding Organophosphorus Hydrolase from Endophytic Pseudomonas Sp. BF1-3 Degrades Organophosphorus Pesticide Chlorpyrifos. Ecotoxicol. Environ. Saf. 2014, 108, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Sauka, D. Bacillus Thuringiensis and Their Endophytic Capabilities. EC Microbiol. 2017, 9, 170–171. [Google Scholar]
- Chen, S.; Deng, Y.; Chang, C.; Lee, J.; Cheng, Y.; Cui, Z.; Zhou, J.; He, F.; Hu, M.; Zhang, L.-H. Pathway and Kinetics of Cyhalothrin Biodegradation by Bacillus Thuringiensis Strain ZS-19. Sci. Rep. 2015, 5, 8784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birolli, W.G.; Dos Santos, A.; Pilau, E.; Rodrigues-Filho, E. New Role for a Commercially Available Bioinsecticide: Bacillus Thuringiensis Berliner Biodegrades the Pyrethroid Cypermethrin. Environ. Sci. Technol. 2021, 55, 4792–4803. [Google Scholar] [CrossRef]
- Żur, J.; Piński, A.; Marchlewicz, A.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic Micropollutants Paracetamol and Ibuprofen-Toxicity, Biodegradation, and Genetic Background of Their Utilization by Bacteria. Environ. Sci. Pollut. Res. Int. 2018, 25, 21498–21524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Sabourin, L.; Renaud, J.; Halloran, S.; Singh, A.; Sumarah, M.; Dagnew, M.; Ray, M.B. Simultaneous Quantification of Five Pharmaceuticals and Personal Care Products in Biosolids and Their Fate in Thermo-Alkaline Treatment. J. Environ. Manage. 2021, 278, 111404. [Google Scholar] [CrossRef] [PubMed]
- Sauvêtre, A.; Węgrzyn, A.; Yang, L.; Vestergaard, G.; Miksch, K.; Schröder, P.; Radl, V. Enrichment of Endophytic Actinobacteria in Roots and Rhizomes of Miscanthus × Giganteus Plants Exposed to Diclofenac and Sulfamethoxazole. Environ. Sci. Pollut. Res. Int. 2020, 27, 11892–11904. [Google Scholar] [CrossRef] [PubMed]
- Węgrzyn, A.; Felis, E. Isolation of Bacterial Endophytes from Phalaris Arundinacea and Their Potential in Diclofenac and Sulfamethoxazole Degradation. Pol. J. Microbiol. 2018, 67, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchlewicz, A.; Guzik, U.; Smułek, W.; Wojcieszyńska, D. Exploring the Degradation of Ibuprofen by Bacillus Thuringiensis B1(2015b): The New Pathway and Factors Affecting Degradation. Molecules 2017, 22, 1676. [Google Scholar] [CrossRef]
- Wojcieszyńska, D.; Guzik, U. Naproxen in the Environment: Its Occurrence, Toxicity to Nontarget Organisms and Biodegradation. Appl. Microbiol. Biotechnol. 2020, 104, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Matta, M.K.; Zusterzeel, R.; Pilli, N.R.; Patel, V.; Volpe, D.A.; Florian, J.; Oh, L.; Bashaw, E.; Zineh, I.; Sanabria, C.; et al. Effect of Sunscreen Application Under Maximal Use Conditions on Plasma Concentration of Sunscreen Active Ingredients: A Randomized Clinical Trial. JAMA 2019, 321, 2082–2091. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Huber, C.; May, R.; Schröder, P. Metabolism of Oxybenzone in a Hairy Root Culture: Perspectives for Phytoremediation of a Widely Used Sunscreen Agent. J. Hazard. Mater. 2016, 306, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Huber, C.; Schröder, P. Fate of the Sunscreen Compound Oxybenzone in Cyperus Alternifolius Based Hydroponic Culture: Uptake, Biotransformation and Phytotoxicity. Chemosphere 2017, 182, 638–646. [Google Scholar] [CrossRef]
- Fagervold, S.K.; Rohée, C.; Rodrigues, A.M.S.; Stien, D.; Lebaron, P. Efficient Degradation of the Organic UV Filter Benzophenone-3 by Sphingomonas Wittichii Strain BP14P Isolated from WWTP Sludge. Sci. Total Environ. 2021, 758, 143674. [Google Scholar] [CrossRef]
- Li, Z.; Nandakumar, R.; Madayiputhiya, N.; Li, X. Proteomic Analysis of 17β-Estradiol Degradation by Stenotrophomonas Maltophilia. Environ. Sci. Technol. 2012, 46, 5947–5955. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Sun, M.; Ling, W.; Zhu, X. Isolation, Characterization, and Degradation Performance of the 17β-Estradiol-Degrading Bacterium Novosphingobium sp. E2S. Int. J. Environ. Res. Public Health 2017, 14, 115. [Google Scholar] [CrossRef] [Green Version]
- Sauvêtre, A.; May, R.; Harpaintner, R.; Poschenrieder, C.; Schröder, P. Metabolism of Carbamazepine in Plant Roots and Endophytic Rhizobacteria Isolated from Phragmites Australis. J. Hazard. Mater. 2018, 342, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Sauvêtre, A.; Schröder, P. Uptake of Carbamazepine by Rhizomes and Endophytic Bacteria of Phragmites Australis. Front. Plant Sci. 2015, 6, 83. [Google Scholar] [CrossRef]
- Ramel, F.; Sulmon, C.; Serra, A.-A.; Gouesbet, G.; Couée, I. Xenobiotic Sensing and Signalling in Higher Plants. J. Exp. Bot. 2012, 63, 3999–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, P.K. Impacts of Particulate Matter Pollution on Plants: Implications for Environmental Biomonitoring. Ecotoxicol. Environ. Saf. 2016, 129, 120–136. [Google Scholar] [CrossRef]
- Afzal, M.; Shabir, G.; Tahseen, R.; Islam, E.; Iqbal, S.; Khan, Q.M.; Khalid, Z.M. Endophytic Burkholderia Sp. Strain PsJN Improves Plant Growth and Phytoremediation of Soil Irrigated with Textile Effluent. CLEAN–Soil Air Water 2013, 42, 1304–1310. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of Action of Plant Growth Promoting Bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duca, D.R.; Glick, B.R. Indole-3-Acetic Acid Biosynthesis and Its Regulation in Plant-Associated Bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC Deaminase Can Promote Plant Growth and Help to Feed the World. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Raghuwanshi, R.; Prasad, J.K. Perspectives of Rhizobacteria with ACC Deaminase Activity in Plant Growth Under Abiotic Stress. In Root Biology; Giri, B., Prasad, R., Varma, A., Eds.; Soil Biology; Springer International Publishing: Cham, Switzerland, 2018; pp. 303–321. ISBN 978-3-319-75910-4. [Google Scholar]
- Del Carmen Orozco-Mosqueda, M.; Glick, B.R.; Santoyo, G. ACC Deaminase in Plant Growth-Promoting Bacteria (PGPB): An Efficient Mechanism to Counter Salt Stress in Crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Gulati, A.; Sharma, N.; Vyas, P.; Sood, S.; Rahi, P.; Pathania, V.; Prasad, R. Organic Acid Production and Plant Growth Promotion as a Function of Phosphate Solubilization by Acinetobacter Rhizosphaerae Strain BIHB 723 Isolated from the Cold Deserts of the Trans-Himalayas. Arch. Microbiol. 2010, 192, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate Solubilizing Bacteria with Glucose Dehydrogenase Gene for Phosphorus Uptake and Beneficial Effects on Wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef] [Green Version]
- De Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant Growth-Promoting Bacteria as Inoculants in Agricultural Soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Bolivar-Anillo, H.J.; González-Rodríguez, V.E.; Cantoral, J.M.; García-Sánchez, D.; Collado, I.G.; Garrido, C. Endophytic Bacteria Bacillus Subtilis, Isolated from Zea Mays, as Potential Biocontrol Agent against Botrytis Cinerea. Biology 2021, 10, 492. [Google Scholar] [CrossRef]
- Buysens, S.; Heungens, K.; Poppe, J.; Hofte, M. Involvement of Pyochelin and Pyoverdin in Suppression of Pythium-Induced Damping-Off of Tomato by Pseudomonas Aeruginosa 7NSK2. Appl. Environ. Microbiol. 1996, 62, 865–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migunova, V.D.; Sasanelli, N. Bacteria as Biocontrol Tool against Phytoparasitic Nematodes. Plants 2021, 10, 389. [Google Scholar] [CrossRef]
- Trapet, P.; Avoscan, L.; Klinguer, A.; Pateyron, S.; Citerne, S.; Chervin, C.; Mazurier, S.; Lemanceau, P.; Wendehenne, D.; Besson-Bard, A. The Pseudomonas Fluorescens Siderophore Pyoverdine Weakens Arabidopsis Thaliana Defense in Favor of Growth in Iron-Deficient Conditions. Plant Physiol. 2016, 171, 675–693. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The Role of Plant Growth Promoting Bacteria in Improving Nitrogen Use Efficiency for Sustainable Crop Production: A Focus on Wheat. AIMS Microbiol. 2017, 3, 413–434. [Google Scholar] [CrossRef]
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 2021, 12, 354. [Google Scholar] [CrossRef]
- Del Carmen Orozco-Mosqueda, M.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant Growth-Promoting Bacteria as Bioinoculants: Attributes and Challenges for Sustainable Crop Improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Nandi, M.; Selin, C.; Brawerman, G.; Fernando, W.G.D.; de Kievit, T. Hydrogen Cyanide, Which Contributes to Pseudomonas Chlororaphis Strain PA23 Biocontrol, Is Upregulated in the Presence of Glycine. Biol. Control 2017, 108, 47–54. [Google Scholar]
- Galeano Vanegas, N.F.; Marulanda Moreno, S.M.; Padilla Hurtado, B.E.; Mantilla Afanador, J.G.; Ceballos Aguirre, N.; Restrepo Franco, G.M.; Galeano Vanegas, N.F.; Marulanda Moreno, S.M.; Padilla Hurtado, B.E.; Mantilla Afanador, J.G.; et al. Antagonism of Plant Growth Promoting Rhizobacteria against the Causal Agent of the Vascular Wilting of Tomato. Rev. Colomb. Biotechnol. 2020, 22, 35–43. [Google Scholar] [CrossRef]
- Ngalimat, M.S.; Mohd Hata, E.; Zulperi, D.; Ismail, S.I.; Ismail, M.R.; Mohd Zainudin, N.A.I.; Saidi, N.B.; Yusof, M.T. Plant Growth-Promoting Bacteria as an Emerging Tool to Manage Bacterial Rice Pathogens. Microorganisms 2021, 9, 682. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C. Plant Responses to Plant Growth-Promoting Rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Zebelo, S.; Song, Y.; Kloepper, J.W.; Fadamiro, H. Rhizobacteria Activates (+)-δ-Cadinene Synthase Genes and Induces Systemic Resistance in Cotton against Beet Armyworm (Spodoptera Exigua). Plant Cell Environ. 2016, 39, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Rokem, J.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.; Amstutz, J.; Patrick, J.E. Biofilm Formation by Bacillus Subtilis Is Altered in the Presence of Pesticides. Access Microbiol. 2020, 2, acmi000175. [Google Scholar] [CrossRef]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus Subtilis against Infection of Arabidopsis Roots by Pseudomonas Syringae Is Facilitated by Biofilm Formation and Surfactin Production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Triveni, S.; Prasanna, R.; Saxena, A.K. Optimization of Conditions for In Vitro Development of Trichoderma Viride-Based Biofilms as Potential Inoculants. Folia Microbiol. 2012, 57, 431–437. [Google Scholar] [CrossRef]
- Kasim, W.A.; Gaafar, R.M.; Abou-Ali, R.M.; Omar, M.N.; Hewait, H.M. Effect of Biofilm Forming Plant Growth Promoting Rhizobacteria on Salinity Tolerance in Barley. Ann. Agric. Sci. 2016, 61, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, G.; Boll, M.; Heider, J. Microbial Degradation of Aromatic Compounds–from One Strategy to Four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef]
- Aliferis, K. Metabolomics in plant protection product research and development: Discovering the mode(s)-of-action and mechanisms of toxicity. In Environmental Metabolomics; Álvarez-Muñoz, D., Farré, M., Eds.; Elsevier: Amsterdam, The Netherland, 2020; pp. 163–194. ISBN 978-0-12-818196-6. [Google Scholar]
- Marchut-Mikołajczyk, O.; Drożdżyński, P.; Polewczyk, A.; Smułek, W.; Antczak, T. Biosurfactant from Endophytic Bacillus Pumilus 2A: Physicochemical Characterization, Production and Optimization and Potential for Plant Growth Promotion. Microb. Cell Factories 2021, 20, 40. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. MMBR 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Lin, Z.; Pang, S.; Zhang, W.; Bhatt, P.; Chen, S. Recent Advanced Technologies for the Characterization of Xenobiotic-Degrading Microorganisms and Microbial Communities. Front. Bioeng. Biotechnol. 2021, 9, 31. [Google Scholar] [CrossRef]
- Lumactud, R.; Shen, S.Y.; Lau, M.; Fulthorpe, R. Bacterial Endophytes Isolated from Plants in Natural Oil Seep Soils with Chronic Hydrocarbon Contamination. Front. Microbiol. 2016, 7, 755. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Li, X.; Xu, J.; Liu, L.; Ren, L.; Dong, B.; Li, W.; Xie, W.; Yao, Z.; Chen, Q.; et al. Diversity and Functional Characteristics of Endophytic Bacteria from Two Grass Species Growing on an Oil-Contaminated Site in the Yellow River Delta, China. Sci. Total Environ. 2021, 767, 144340. [Google Scholar] [CrossRef] [PubMed]
- Lumactud, R.; Fulthorpe, R.R. Endophytic Bacterial Community Structure and Function of Herbaceous Plants From Petroleum Hydrocarbon Contaminated and Non-Contaminated Sites. Front. Microbiol. 2018, 9, 1926. [Google Scholar] [CrossRef] [Green Version]
- White, J.F.; Kingsley, K.L.; Zhang, Q.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K.P. Review: Endophytic Microbes and Their Potential Applications in Crop Management. Pest Manag. Sci. 2019, 75, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Macchi, M.; Festa, S.; Nieto, E.; Irazoqui, J.M.; Vega-Vela, N.E.; Junca, H.; Valacco, M.P.; Amadio, A.F.; Morelli, I.S.; Coppotelli, B.M. Design and Evaluation of Synthetic Bacterial Consortia for Optimized Phenanthrene Degradation through the Integration of Genomics and Shotgun Proteomics. Biotechnol. Rep. 2021, 29, e00588. [Google Scholar] [CrossRef]
- Gonzalez, E.; Pitre, F.E.; Pagé, A.P.; Marleau, J.; Guidi Nissim, W.; St-Arnaud, M.; Labrecque, M.; Joly, S.; Yergeau, E.; Brereton, N.J.B. Trees, Fungi and Bacteria: Tripartite Metatranscriptomics of a Root Microbiome Responding to Soil Contamination. Microbiome 2018, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Endophytic Strain | Host Plant | Degraded Xenobiotic | PGP-Activity | References |
|---|---|---|---|---|
| Pseudomonas aeruginosa L10 | roots Phragmites australis | C10—C26 n-alkanes, naphthalene, phenanthrene, pyrene | Siderophore, biosurfactant and IAA production, ACC activity | [27] |
| Bacillus safensis ZY16 | roots Chloris virgate Sw | C12—C32 n-alkanes, naphthalene, phenanthrene, pyrene | Siderophore, biosurfactant and IAA production, ACC activityphosphate solubilization | [17] |
| Enterobacter sp. 12J1 | Allium macrostemon Bunge | pyrene | Siderophore, IAA production, phosphate solubilization | [30] |
| Bacillus sp. SBER3 | Populus deltoides | anthracene, naphthalene, benzene, toluene and xylene | Siderophore, IAA production, ACC activity, phosphate solubilization, biocontrol activity | [20] |
| Burkholderia sp. PsJN | Onion roots | Textile effluent | ACC activity | [89] |
| Microbacterium arborescens TYSI04 | shoot T. domingensis | [49] | ||
| Bacillus pumilus PIRI30 | roots Pistia | [49] | ||
| Pseudomonas aeruginosa RRA, Bacillus megaterium RRB, Sphingobacterium siyangensis RSA, Stenotrophomonas pavanii RSB and Curtobacterium plantarum RSC | Oryza sativa L. | chlorpyrifos | Siderophore, IAA production, ACC activity, phosphate solubilization | [68] |
| Pantoea dispersa | roots Dracena sanderiana | Bisphenol A | IAA production, ROS scavenging | [58] |
| Streptomyces griseorubiginosus strains DS24 and DS4 | roots Miscanthus × giganteus | diclofenac and sulfamethoxazole | Siderophore, IAA production | [75] |
| “Omics”—Based System | Used Technics | Source of Bacteria | Pollutant for Degradation | References |
|---|---|---|---|---|
| metagenomics | TRFLP of PCR-amplified 16S rRNA gene fragments obtained with restriction enzymes PvuII and MscI | Achillea millefolium Dactylis glomerata Trifolium aureum Solidago canadensis | PHCs | [122] |
| 16S rRNA gene amplicon sequencing | Phragmites australis Chloris virgata | PHCs with saline | [123] | |
| functional metagenomics | alkB, ndoB, and ntdA genes probe analysis | Festuca arundinacea Trifolium fragiferum | nitroaromatics | [10] |
| proteomics | nanoLC-MS/MS with XCalibur 3.0.63 software | synthetic consortium | phenanthrene | [126] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaś, M.A.; Wdowiak-Wróbel, S.; Sokołowski, W. Selection of Endophytic Strains for Enhanced Bacteria-Assisted Phytoremediation of Organic Pollutants Posing a Public Health Hazard. Int. J. Mol. Sci. 2021, 22, 9557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179557
Karaś MA, Wdowiak-Wróbel S, Sokołowski W. Selection of Endophytic Strains for Enhanced Bacteria-Assisted Phytoremediation of Organic Pollutants Posing a Public Health Hazard. International Journal of Molecular Sciences. 2021; 22(17):9557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179557
Chicago/Turabian StyleKaraś, Magdalena Anna, Sylwia Wdowiak-Wróbel, and Wojciech Sokołowski. 2021. "Selection of Endophytic Strains for Enhanced Bacteria-Assisted Phytoremediation of Organic Pollutants Posing a Public Health Hazard" International Journal of Molecular Sciences 22, no. 17: 9557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179557