
Novel Nitrogen-Based Chalcone Analogs Provoke Substantial Apoptosis in HER2-Positive Human Breast Cancer Cells via JNK and ERK1/ERK2 Signaling Pathways

 , ,
, , 
 and
and 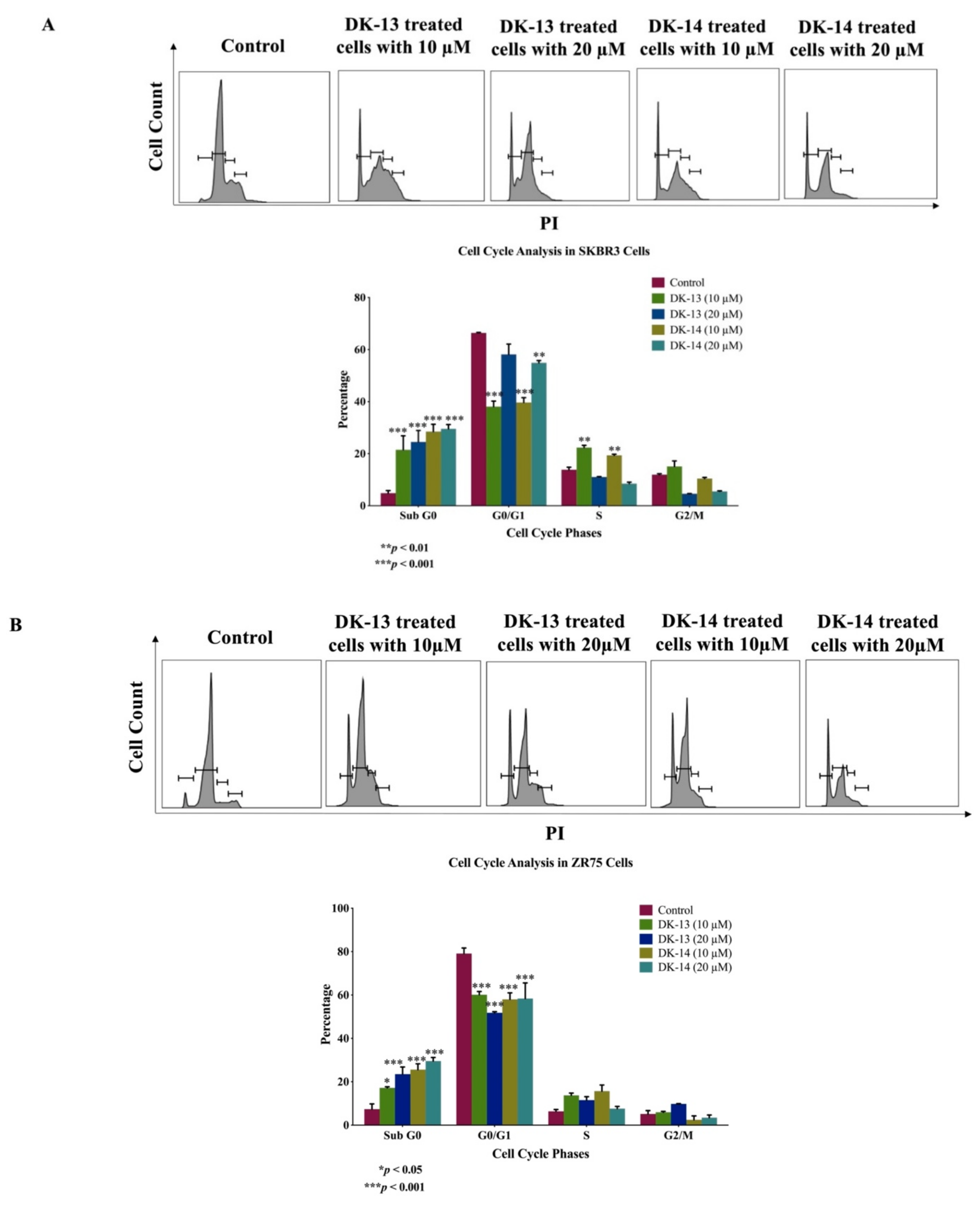{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
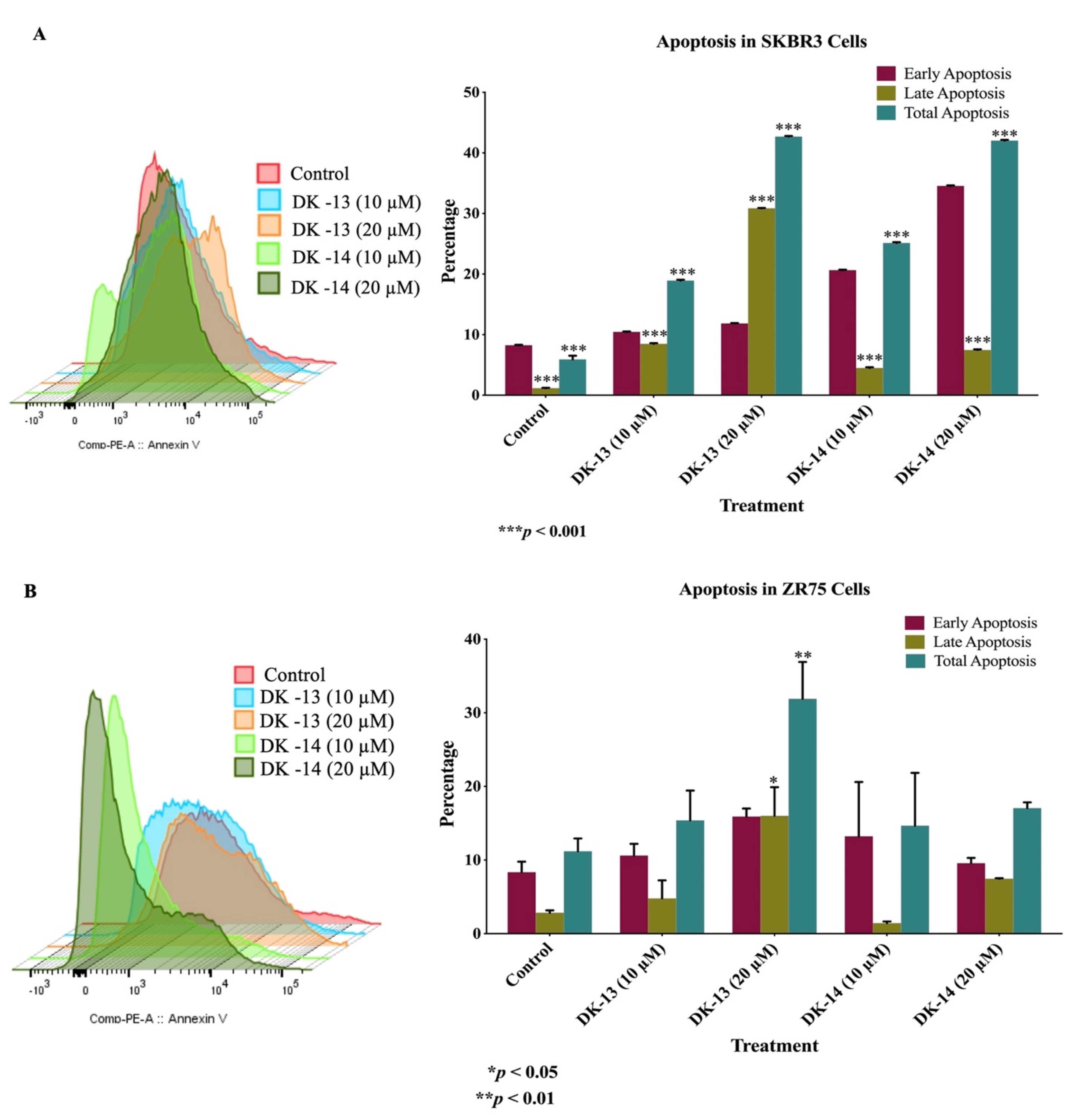
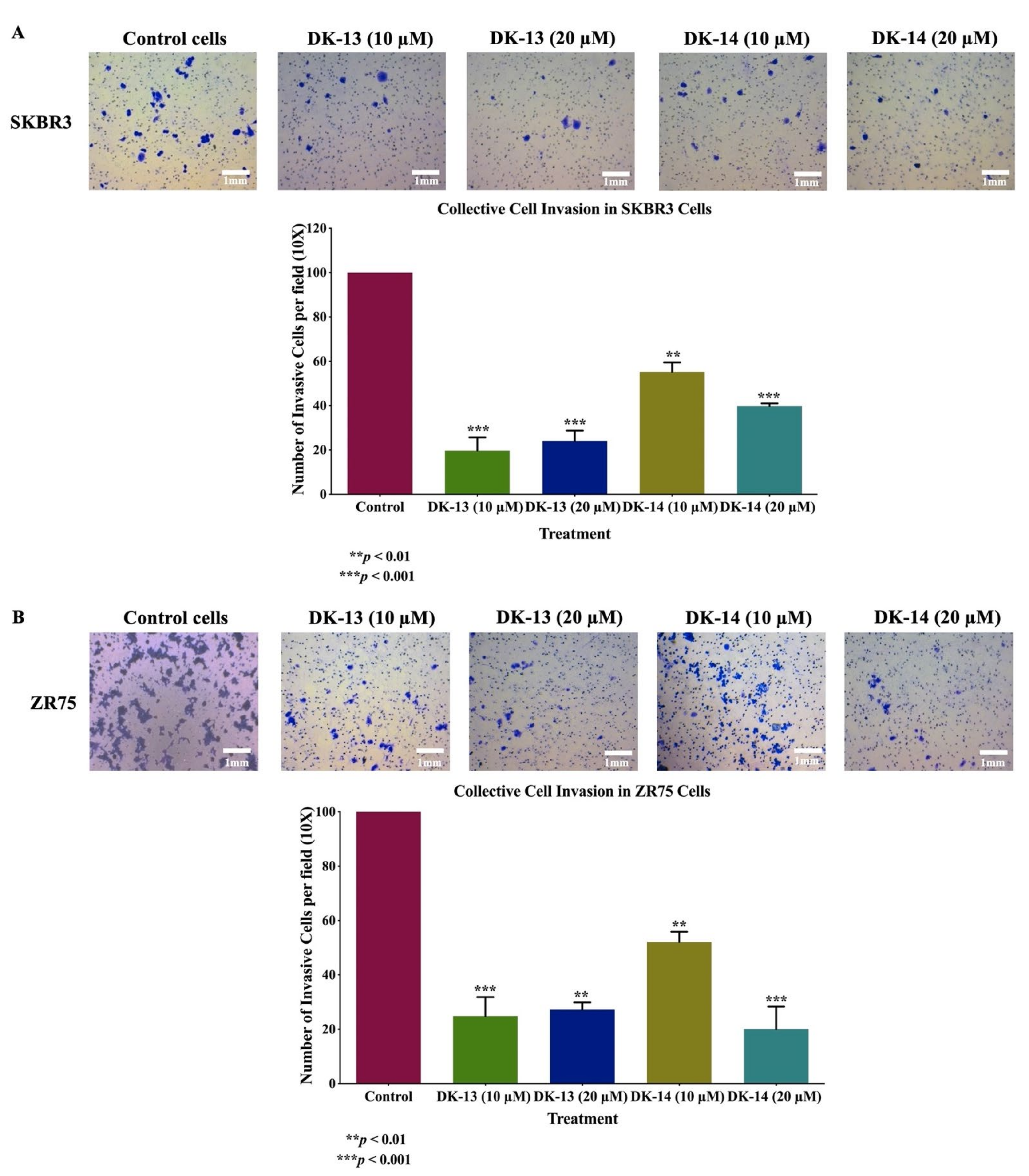
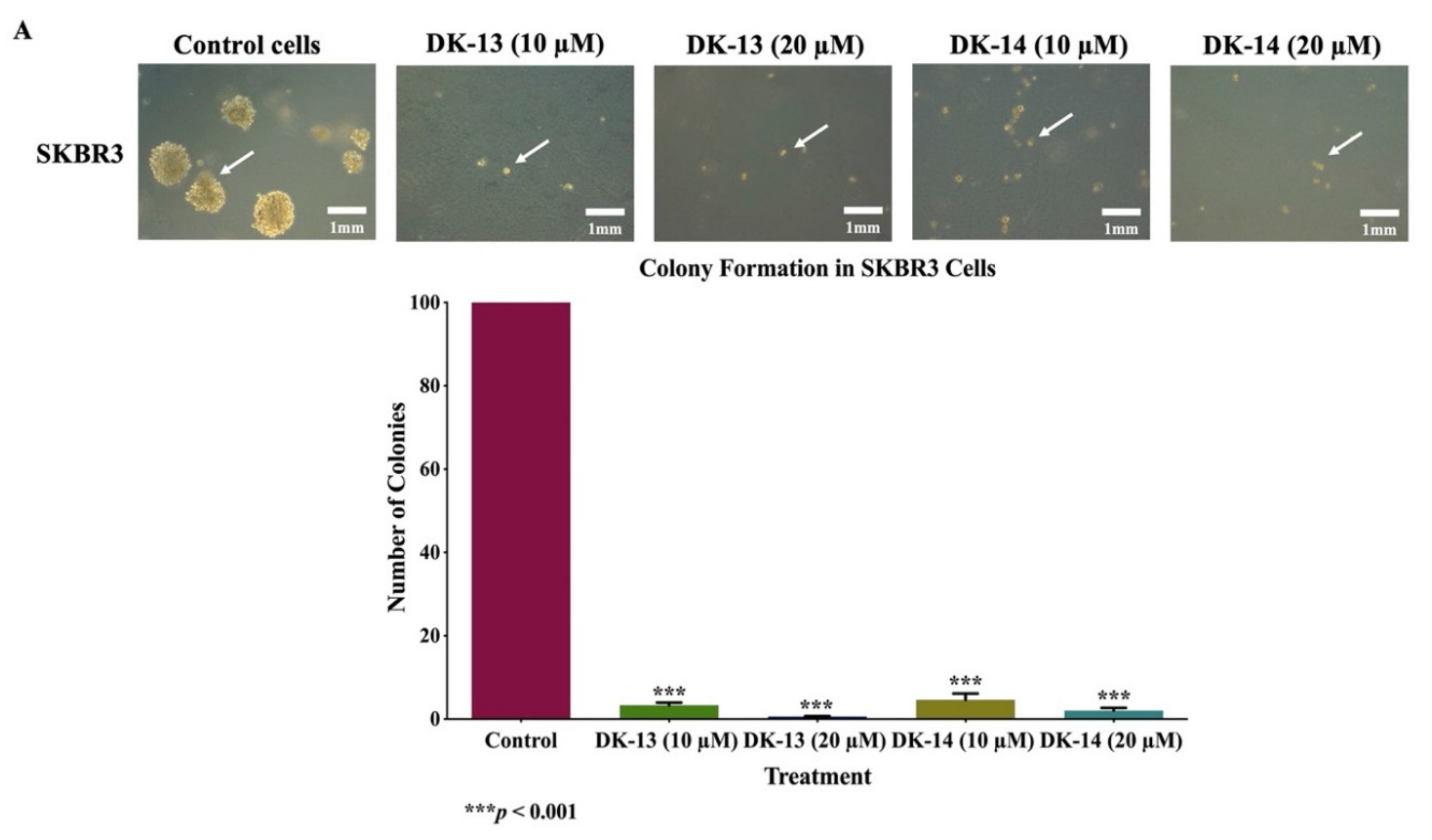
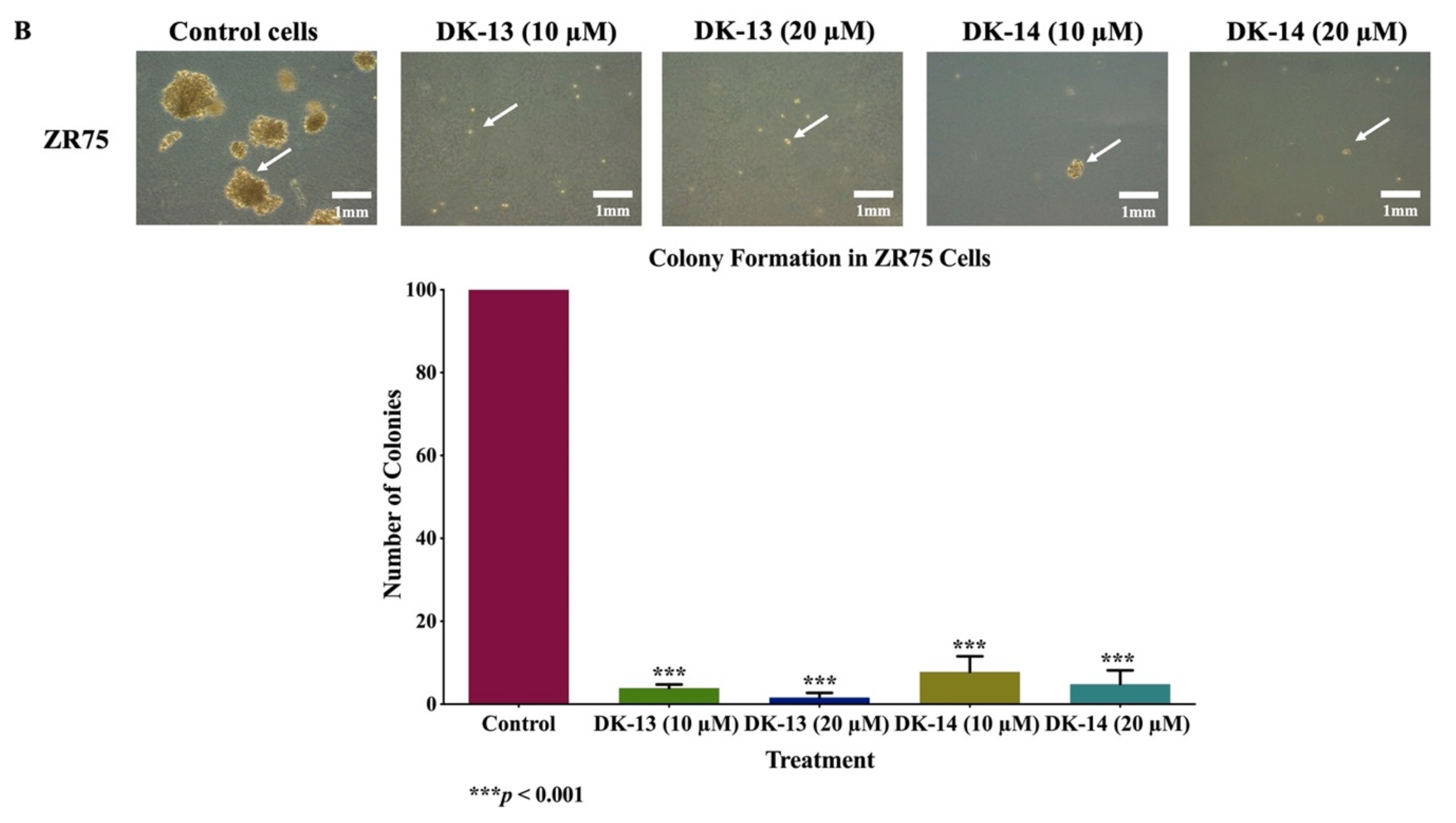
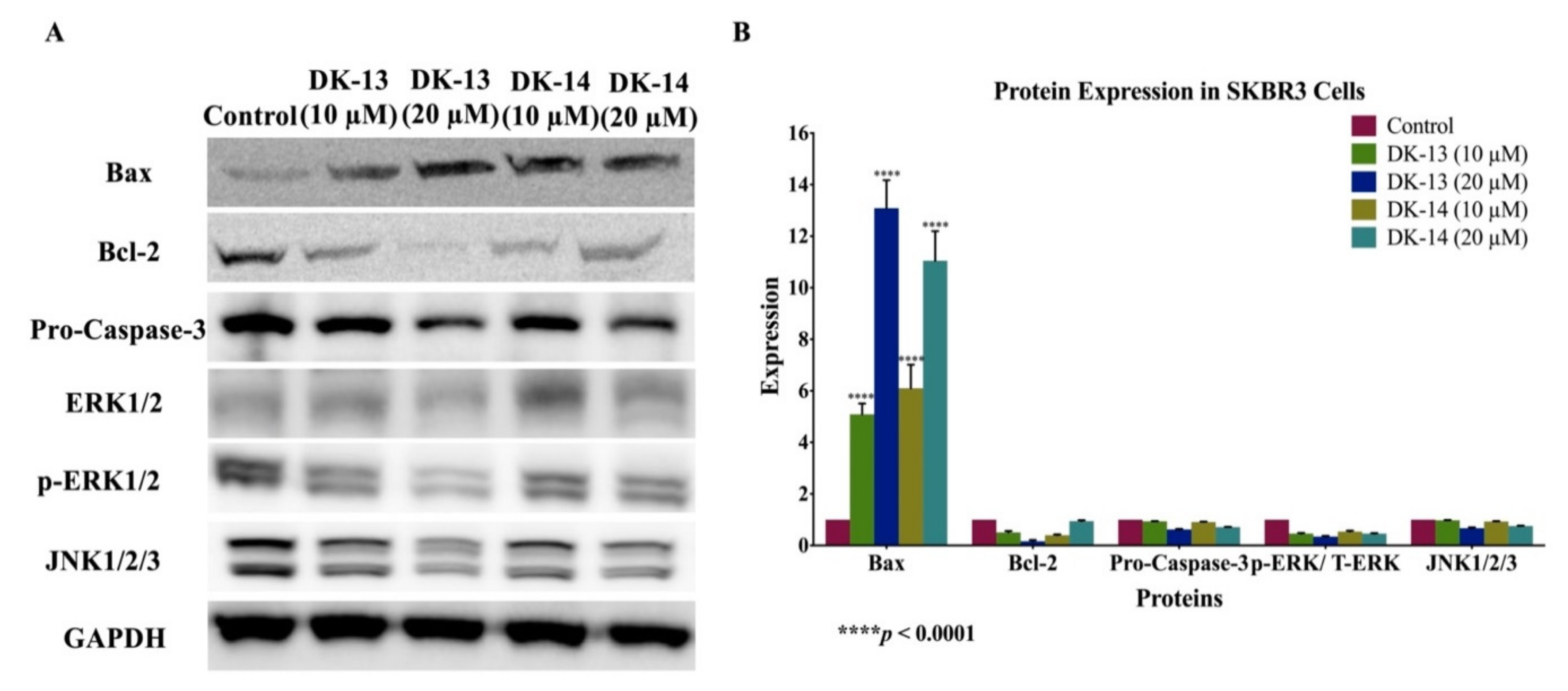
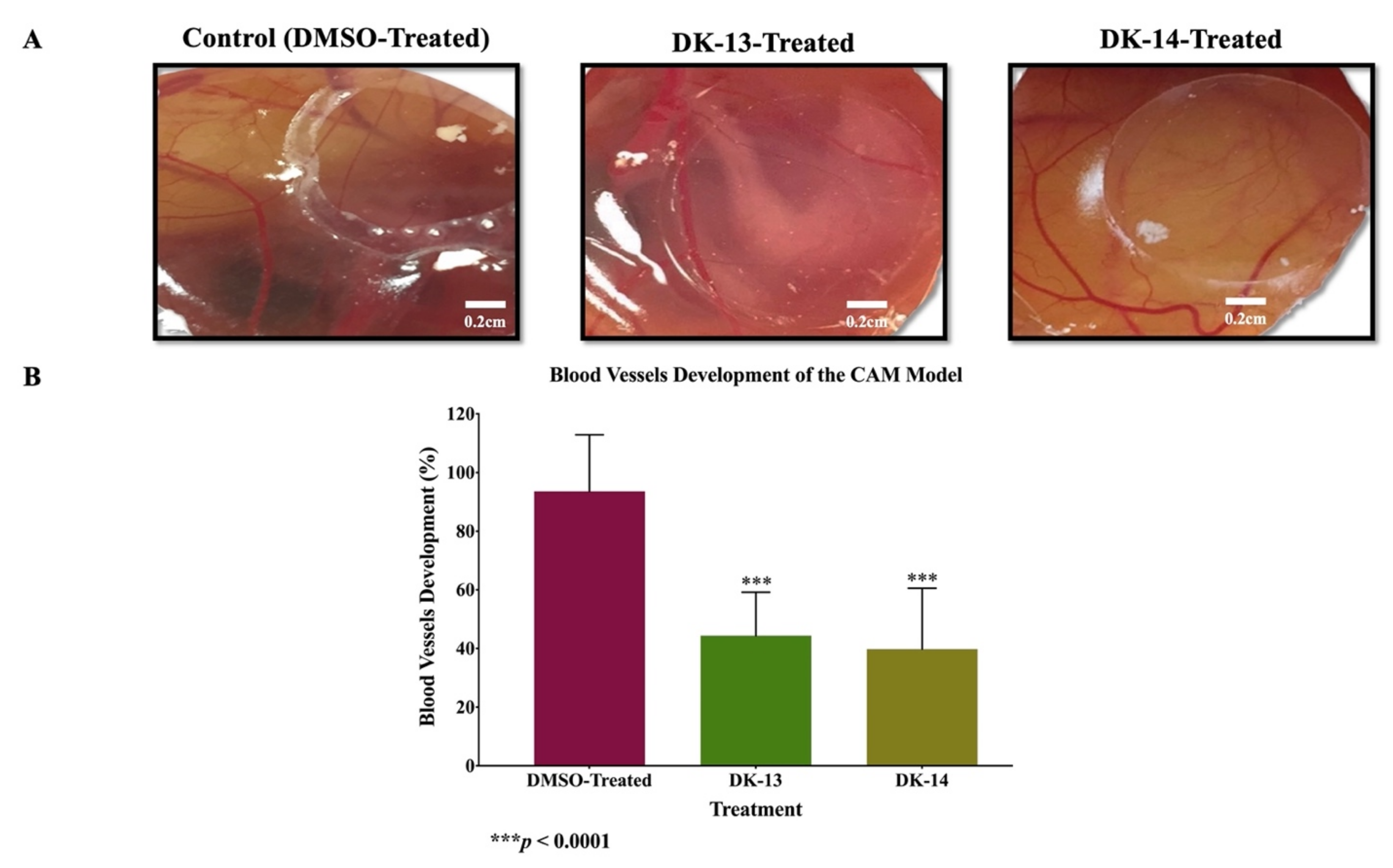
2. Results
3. Discussion
4. Materials and Methods
4.1. Compounds Preparation
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Cycle
4.5. Annexin V Apoptosis Assay
4.6. Cell Invasion Assay
4.7. Soft Agar Colony Formation Assay
4.8. Western Blotting
4.9. Angiogenesis Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Slamon, D.J.; Godolphin, W.; Jones, L.A.; Holt, J.A.; Wong, S.G.; Keith, D.E.; Levin, W.J.; Stuart, S.G.; Udove, J.; Ullrich, A.; et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science 1989, 244, 707–712. [Google Scholar] [CrossRef]
- Schettini, F.; Buono, G.; Cardalesi, C.; Desideri, I.; De Placido, S.; Del Mastro, L. Hormone receptor/human epidermal growth factor receptor 2-positive breast cancer: Where we are now and where we are going. Cancer Treat. Rev. 2016, 46, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Payne, C.M.; Bernstein, C.; Bernstein, H. Apoptosis overview emphasizing the role of oxidative stress, DNA damage and signal-transduction pathways. Leuk. Lymphoma 1995, 19, 43–93. [Google Scholar] [CrossRef]
- Wang, J.; Xu, B. Targeted therapeutic options and future perspectives for HER2-positive breast cancer. Signal Transduct. Target. Ther. 2019, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Gottesman, M.M.; Lavi, O.; Hall, M.D.; Gillet, J.-P. Toward a better understanding of the complexity of cancer drug resistance. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 85–102. [Google Scholar] [CrossRef]
- Pisha, E.; Chai, H.; Lee, I.-S.; Chagwedera, T.E.; Farnsworth, N.R.; Cordell, G.A.; Beecher, C.W.; Fong, H.H.; Kinghorn, A.D.; Brown, D.M. Discovery of betulinic acid as a selective inhibitor of human melanoma that functions by induction of apoptosis. Nat. Med. 1995, 1, 1046–1051. [Google Scholar] [CrossRef]
- Sporn, M.B.; Dunlop, N.; Newton, D.; Smith, J. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs (retinoids). Fed. Proc. 1976, 35, 1332–1338. [Google Scholar]
- Rizeq, B.; Gupta, I.; Ilesanmi, J.; AlSafran, M.; Rahman, M.M.; Ouhtit, A. The Power of Phytochemicals Combination in Cancer Chemoprevention. J. Cancer 2020, 11, 4521. [Google Scholar] [CrossRef]
- Chavan, B.; Gadekar, A.; Mehta, P.; Vawhal, P.; Kolsure, A.; Chabukswar, A. Synthesis & Medicinal Significance of Chalcones-A Review. Asian J. Biomed. Pharm. Sci. 2016, 6, 1. [Google Scholar]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef]
- Carroll, A.E.; Bauer, N.S.; Dugan, T.M.; Anand, V.; Saha, C.; Downs, S.M. Use of a computerized decision aid for developmental surveillance and screening: A randomized clinical trial. JAMA Pediatrics 2014, 168, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Bonakdar, A.P.S.; Vafaei, F.; Farokhpour, M.; Esfahani, M.H.N.; Massah, A.R. Synthesis and anticancer activity assay of novel chalcone-sulfonamide derivatives. Iran. J. Pharm. Res. IJPR 2017, 16, 565. [Google Scholar]
- Osório, T.M.; Delle Monache, F.; Chiaradia, L.D.; Mascarello, A.; Stumpf, T.R.; Zanetti, C.R.; Silveira, D.B.; Barardi, C.R.M.; Smânia, E.d.F.A.; Viancelli, A. Antibacterial activity of chalcones, hydrazones and oxadiazoles against methicillin-resistant Staphylococcus aureus. Bioorganic Med. Chem. Lett. 2012, 22, 225–230. [Google Scholar] [CrossRef]
- Kuete, V.; Sandjo, L.P. Isobavachalcone: An overview. Chin. J. Integr. Med. 2012, 18, 543–547. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Zhou, D.-G.; He, F.-C.; Chen, J.-X.; Chen, Y.-Z.; Gan, X.-H.; Hu, D.-Y.; Song, B.-A. Synthesis and antiviral bioactivity of novel chalcone derivatives containing purine moiety. Chin. Chem. Lett. 2018, 29, 127–130. [Google Scholar] [CrossRef]
- Sivakumar, P.; Prabhakar, P.; Doble, M. Synthesis, antioxidant evaluation, and quantitative structure–activity relationship studies of chalcones. Med. Chem. Res. 2011, 20, 482–492. [Google Scholar] [CrossRef]
- Huang, Z.-H.; Yin, L.-Q.; Guan, L.-P.; Li, Z.-H.; Tan, C. Screening of Chalcone Analogs with Anti-depressant, Anti-inflammatory, Analgesic, and COX-2–inhibiting Effects. Bioorganic Med. Chem. Lett. 2020, 30, 127173. [Google Scholar] [CrossRef]
- Gómez-Rivera, A.; Aguilar-Mariscal, H.; Romero-Ceronio, N.; Roa-de la Fuente, L.F.; Lobato-García, C.E. Synthesis and anti-inflammatory activity of three nitro chalcones. Bioorganic Med. Chem. Lett. 2013, 23, 5519–5522. [Google Scholar] [CrossRef] [Green Version]
- Jandial, D.; A Blair, C.; Zhang, S.; S Krill, L.; Zhang, Y.-B.; Zi, X. Molecular targeted approaches to cancer therapy and prevention using chalcones. Curr. Cancer Drug Targets 2014, 14, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Elkhalifa, D.; Siddique, A.B.; Qusa, M.; Cyprian, F.S.; El Sayed, K.; Alali, F.; Al Moustafa, A.-E.; Khalil, A. Design, synthesis, and validation of novel nitrogen-based chalcone analogs against triple negative breast cancer. Eur. J. Med. Chem. 2020, 187, 111954. [Google Scholar] [CrossRef]
- León-González, A.J.; Acero, N.; Muñoz-Mingarro, D.; Navarro, I.; Martín-Cordero, C. Chalcones as Promising Lead Compounds on Cancer Therapy. Curr. Med. Chem. 2015, 22, 3407–3425. [Google Scholar] [CrossRef]
- Peyrot, V.; Leynadier, D.; Sarrazin, M.; Briand, C.; Rodriquez, A.; Nieto, J.M.; Andreu, J.M. Interaction of tubulin and cellular microtubules with the new antitumor drug MDL 27048. A powerful and reversible microtubule inhibitor. J. Biol. Chem. 1989, 264, 21296–21301. [Google Scholar] [CrossRef]
- Ji, T.; Lin, C.; Krill, L.S.; Eskander, R.; Guo, Y.; Zi, X.; Hoang, B.H. Flavokawain B, a kava chalcone, inhibits growth of human osteosarcoma cells through G2/M cell cycle arrest and apoptosis. Mol. Cancer 2013, 12, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Hsieh, T.C.; Guo, J.; Kunicki, J.; Lee, M.Y.; Darzynkiewicz, Z.; Wu, J.M. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2004, 322, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Lim, D.Y.; Choi, H.J.; Jung, J.I.; Chung, W.Y.; Park, J.H. Induction of cell cycle arrest in prostate cancer cells by the dietary compound isoliquiritigenin. J. Med. Food 2009, 12, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Issaenko, O.A.; Amerik, A.Y. Chalcone-based small-molecule inhibitors attenuate malignant phenotype via targeting deubiquitinating enzymes. Cell Cycle 2012, 11, 1804–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Chen, M.; Chen, W.; Hui, J.; Ji, J.; Hu, S.; Zhou, J.; Wang, Y.; Liang, G. Chemopreventive effect of chalcone derivative, L2H17, in colon cancer development. BMC Cancer 2015, 15, 870. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-L.; Ma, Y.-H.; Li, Y.-H.; Zhang, Y.-P.; Tian, H.-C.; Huang, Y.-C.; Li, Y.; Chen, W.; Yang, L.-J. Design, Synthesis, and Anticancer Activity of Novel Trimethoxyphenyl-Derived Chalcone-Benzimidazolium Salts. ACS Omega 2019, 4, 20381–20393. [Google Scholar] [CrossRef]
- Tseng, C.H.; Chen, Y.L.; Hsu, C.Y.; Chen, T.C.; Cheng, C.M.; Tso, H.C.; Lu, Y.J.; Tzeng, C.C. Synthesis and antiproliferative evaluation of 3-phenylquinolinylchalcone derivatives against non-small cell lung cancers and breast cancers. Eur. J. Med. Chem. 2013, 59, 274–282. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Singh, A.K.; Meena, S.; Lohani, M.; Singh, A.; Arya, R.K.; Cheruvu, S.H.; Sarkar, J.; Gayen, J.R.; Datta, D.; et al. Synthesis of novel β-carboline based chalcones with high cytotoxic activity against breast cancer cells. Bioorganic Med. Chem. Lett. 2014, 24, 2820–2824. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, M.S.; Koh, D.; Lee, Y.H.; Lim, Y.; Shin, S.Y. A new synthetic 2’-hydroxy-2,4,6-trimethoxy-5’,6’-naphthochalcone induces G2/M cell cycle arrest and apoptosis by disrupting the microtubular network of human colon cancer cells. Cancer Lett. 2014, 354, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sarkar, J.; Sashidhara, K.V.; Ali, S.; Sinha, S. Anti-tumour activity of a novel coumarin-chalcone hybrid is mediated through intrinsic apoptotic pathway by inducing PUMA and altering Bax/Bcl-2 ratio. Apoptosis 2014, 19, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gwak, J.; Han, S.J.; Oh, S. Cardamonin suppresses the proliferation of colon cancer cells by promoting β-catenin degradation. Biol. Pharm. Bull. 2013, 36, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.; Xu, L.; Hua, L.; Li, A.; Li, S.; Lu, W.; Pang, Y.; Cao, C.; Liu, X.; Jiao, P. Synthesis and evaluation of novel isoxazolyl chalcones as potential anticancer agents. Bioorganic Chem. 2014, 54, 38–43. [Google Scholar] [CrossRef]
- Zhang, E.-H.; Wang, R.-F.; Guo, S.-Z.; Liu, B. An Update on Antitumor Activity of Naturally Occurring Chalcones. Evid. Based Complementary Altern. Med. 2013, 2013, 815621. [Google Scholar] [CrossRef]
- Chen, J.; Kang, C.Y.; Niu, Z.X.; Zhou, H.C.; Yang, H.M. A chalcone inhibits the growth and metastasis of KYSE-4 esophageal cancer cells. J. Int. Med. Res. 2020, 48, 300060520928831. [Google Scholar] [CrossRef]
- Albini, A.; Dell’Eva, R.; Vené, R.; Ferrari, N.; Buhler, D.R.; Noonan, D.M.; Fassina, G. Mechanisms of the antiangiogenic activity by the hop flavonoid xanthohumol: NF-kappaB and Akt as targets. Faseb J. 2006, 20, 527–529. [Google Scholar] [CrossRef]
- Dell’Eva, R.; Ambrosini, C.; Vannini, N.; Piaggio, G.; Albini, A.; Ferrari, N. AKT/NF-kappaB inhibitor xanthohumol targets cell growth and angiogenesis in hematologic malignancies. Cancer 2007, 110, 2007–2011. [Google Scholar] [CrossRef]
- Pilatova, M.; Varinska, L.; Perjesi, P.; Sarissky, M.; Mirossay, L.; Solar, P.; Ostro, A.; Mojzis, J. In vitro antiproliferative and antiangiogenic effects of synthetic chalcone analogues. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2010, 24, 1347–1355. [Google Scholar] [CrossRef]
- Nam, N.H.; Kim, Y.; You, Y.J.; Hong, D.H.; Kim, H.M.; Ahn, B.Z. Cytotoxic 2′,5′-dihydroxychalcones with unexpected antiangiogenic activity. Eur. J. Med. Chem. 2003, 38, 179–187. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lim, S.S.; Shin, K.H.; Kim, Y.S.; Ohuchi, K.; Jung, S.H. Anti-angiogenic and anti-tumor activities of 2′-hydroxy-4′-methoxychalcone. Biol. Pharm. Bull. 2006, 29, 1028–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.F.; Xie, B.F.; Zhou, J.M.; Feng, G.K.; Liu, Z.C.; Wei, X.Y.; Zhang, F.X.; Liu, M.F.; Zeng, Y.X. Blockade of vascular endothelial growth factor receptor signal pathway and antitumor activity of ON-III (2′,4′-dihydroxy-6′-methoxy-3′,5′-dimethylchalcone), a component from Chinese herbal medicine. Mol. Pharmacol. 2005, 67, 1444–1450. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.U.; Siddiqui, H.L.; Nisar, M.; Khan, N.; Khan, I. Discovery and molecular docking of quinolyl-thienyl chalcones as anti-angiogenic agents targeting VEGFR-2 tyrosine kinase. Bioorganic Med. Chem. Lett. 2012, 22, 942–944. [Google Scholar] [CrossRef]
- Mojzis, J.; Varinska, L.; Mojzisova, G.; Kostova, I.; Mirossay, L. Antiangiogenic effects of flavonoids and chalcones. Pharmacol. Res. 2008, 57, 259–265. [Google Scholar] [CrossRef]
- Negrão, R.; Incio, J.; Lopes, R.; Azevedo, I.; Soares, R. Evidence for the Effects of Xanthohumol in Disrupting Angiogenic, but not Stable Vessels. Int. J. Biomed. Sci 2007, 3, 279–286. [Google Scholar]
- Kolundžija, B.; Marković, V.; Stanojković, T.; Joksović, L.; Matić, I.; Todorović, N.; Nikolić, M.; Joksović, M.D. Novel anthraquinone based chalcone analogues containing an imine fragment: Synthesis, cytotoxicity and anti-angiogenic activity. Bioorganic Med. Chem. Lett. 2014, 24, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Ducki, S.; Mackenzie, G.; Greedy, B.; Armitage, S.; Chabert, J.F.; Bennett, E.; Nettles, J.; Snyder, J.P.; Lawrence, N.J. Combretastatin-like chalcones as inhibitors of microtubule polymerisation. Part 2: Structure-based discovery of alpha-aryl chalcones. Bioorganic Med. Chem. 2009, 17, 7711–7722. [Google Scholar] [CrossRef]
- Ducki, S.; Rennison, D.; Woo, M.; Kendall, A.; Chabert, J.F.; McGown, A.T.; Lawrence, N.J. Combretastatin-like chalcones as inhibitors of microtubule polymerization. Part 1: Synthesis and biological evaluation of antivascular activity. Bioorganic Med. Chem. 2009, 17, 7698–7710. [Google Scholar] [CrossRef]
- Lei, K.; Nimnual, A.; Zong, W.X.; Kennedy, N.J.; Flavell, R.A.; Thompson, C.B.; Bar-Sagi, D.; Davis, R.J. The Bax subfamily of Bcl2-related proteins is essential for apoptotic signal transduction by c-Jun NH(2)-terminal kinase. Mol. Cell. Biol. 2002, 22, 4929–4942. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.L.; Chia, C.C.; Chen, P.J.; Huang, S.E.; Huang, S.C.; Kuo, P.L. Shallot and licorice constituent isoliquiritigenin arrests cell cycle progression and induces apoptosis through the induction of ATM/p53 and initiation of the mitochondrial system in human cervical carcinoma HeLa cells. Mol. Nutr. Food Res. 2009, 53, 826–835. [Google Scholar] [CrossRef]
- Shen, K.H.; Chang, J.K.; Hsu, Y.L.; Kuo, P.L. Chalcone arrests cell cycle progression and induces apoptosis through induction of mitochondrial pathway and inhibition of nuclear factor kappa B signalling in human bladder cancer cells. Basic Clin. Pharmacol. Toxicol. 2007, 101, 254–261. [Google Scholar] [CrossRef]
- Yun, J.M.; Kweon, M.H.; Kwon, H.; Hwang, J.K.; Mukhtar, H. Induction of apoptosis and cell cycle arrest by a chalcone panduratin A isolated from Kaempferia pandurata in androgen-independent human prostate cancer cells PC3 and DU145. Carcinogenesis 2006, 27, 1454–1464. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.L.; Kuo, P.L.; Lin, C.C. Isoliquiritigenin induces apoptosis and cell cycle arrest through p53-dependent pathway in Hep G2 cells. Life Sci. 2005, 77, 279–292. [Google Scholar] [CrossRef]
- Winter, E.; Chiaradia, L.D.; de Cordova, C.A.; Nunes, R.J.; Yunes, R.A.; Creczynski-Pasa, T.B. Naphthylchalcones induce apoptosis and caspase activation in a leukemia cell line: The relationship between mitochondrial damage, oxidative stress, and cell death. Bioorganic Med. Chem. 2010, 18, 8026–8034. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Chiaradia-Delatorre, L.D.; dos Santos Bubniak, L.; Mascarello, A.; Marzarotto, M.A.L.; de Moraes, A.C.R.; Stumpf, T.R.; Cordeiro, M.N.S.; Yunes, R.A.; Nunes, R.J.; et al. Apoptotic effect of synthetic 2′,4′,5′-trimethoxychalcones in human K562 and Jurkat leukemia cells. Med. Chem. Res. 2014, 23, 4301–4319. [Google Scholar] [CrossRef]
- Wada, T.; Penninger, J.M. Mitogen-activated protein kinases in apoptosis regulation. Oncogene 2004, 23, 2838–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.K. MAP kinase pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a011254. [Google Scholar] [CrossRef]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death—Apoptosis, autophagy and senescence. Febs J. 2010, 277, 2–21. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, J.; Xiao, H.; Wu, W.; Wang, Y.; Liu, X. MAPK signaling pathways regulate mitochondrial-mediated apoptosis induced by isoorientin in human hepatoblastoma cancer cells. Food Chem. Toxicol. 2013, 53, 62–68. [Google Scholar] [CrossRef]
- Kuruc, T.; Kello, M.; Petrova, K.; Kudlickova, Z.; Kubatka, P.; Mojzis, J. The Newly Synthetized Chalcone L1 Is Involved in the Cell Growth Inhibition, Induction of Apoptosis and Suppression of Epithelial-to-Mesenchymal Transition of HeLa Cells. Molecules 2021, 26, 1356. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK signaling in apoptosis. Oncogene 2008, 27, 6245–6251. [Google Scholar] [CrossRef] [Green Version]
- Warmka, J.K.; Solberg, E.L.; Zeliadt, N.A.; Srinivasan, B.; Charlson, A.T.; Xing, C.; Wattenberg, E.V. Inhibition of mitogen activated protein kinases increases the sensitivity of A549 lung cancer cells to the cytotoxicity induced by a kava chalcone analog. Biochem. Biophys. Res. Commun. 2012, 424, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Sian, L.J.; Jasamai, M.; Yaakob, N.S.; Fauzi, N.M. Synthetic chalcone derivatives inhibit cytokine secretion via inhibition of ERK and JNK pathways in human U937 macrophage. Trop. J. Pharm. Res. 2019, 18, 753–759. [Google Scholar] [CrossRef]
- Yasmeen, A.; Alachkar, A.; Dekhil, H.; Gambacorti-Passerini, C.; Al Moustafa, A.E. Locking Src/Abl Tyrosine Kinase Activities Regulate Cell Differentiation and Invasion of Human Cervical Cancer Cells Expressing E6/E7 Oncoproteins of High-Risk HPV. J. Oncol. 2010, 2010. [Google Scholar] [CrossRef]
- Jabeen, A.; Sharma, A.; Gupta, I.; Kheraldine, H.; Vranic, S.; Al Moustafa, A.-E.; Al Farsi, H.F. Elaeagnus angustifolia Plant Extract Inhibits Epithelial-Mesenchymal Transition and Induces Apoptosis via HER2 Inactivation and JNK Pathway in HER2-Positive Breast Cancer Cells. Molecules 2020, 25, 4240. [Google Scholar] [CrossRef]
- Alhussain, H.; Augustine, R.; Hussein, E.A.; Gupta, I.; Hasan, A.; Al Moustafa, A.E.; Elzatahry, A. MXene Nanosheets May Induce Toxic Effect on the Early Stage of Embryogenesis. J. Biomed. Nanotechnol. 2020, 16, 364–372. [Google Scholar] [CrossRef]
- Abdo, G.G.; Kheraldine, H.; Gupta, I.; Rizeq, B.; Elzatahry, A.; Al Moustafa, A.E.; Khalil, A. Significant Toxic Effect of Carbon Nanofibers at the Early Stage of Embryogenesis. J. Biomed. Nanotechnol. 2020, in press. [Google Scholar] [CrossRef]
- Zudaire, E.; Gambardella, L.; Kurcz, C.; Vermeren, S. A Computational Tool for Quantitative Analysis of Vascular Networks. PLoS ONE 2011, 6, e27385. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizeq, B.; Gupta, I.; Kheraldine, H.; Elkhalifa, D.; Al-Farsi, H.F.; Moustafa, A.-E.A.; Khalil, A. Novel Nitrogen-Based Chalcone Analogs Provoke Substantial Apoptosis in HER2-Positive Human Breast Cancer Cells via JNK and ERK1/ERK2 Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 9621. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179621
Rizeq B, Gupta I, Kheraldine H, Elkhalifa D, Al-Farsi HF, Moustafa A-EA, Khalil A. Novel Nitrogen-Based Chalcone Analogs Provoke Substantial Apoptosis in HER2-Positive Human Breast Cancer Cells via JNK and ERK1/ERK2 Signaling Pathways. International Journal of Molecular Sciences. 2021; 22(17):9621. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179621
Chicago/Turabian StyleRizeq, Balsam, Ishita Gupta, Hadeel Kheraldine, Dana Elkhalifa, Halema F. Al-Farsi, Ala-Eddin Al Moustafa, and Ashraf Khalil. 2021. "Novel Nitrogen-Based Chalcone Analogs Provoke Substantial Apoptosis in HER2-Positive Human Breast Cancer Cells via JNK and ERK1/ERK2 Signaling Pathways" International Journal of Molecular Sciences 22, no. 17: 9621. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179621