
Analyses of the Mode of Action of an Alpha-Adrenoceptor Blocker in Substantia Gelatinosa Neurons in Rats


Abstract
:1. Introduction
2. Results
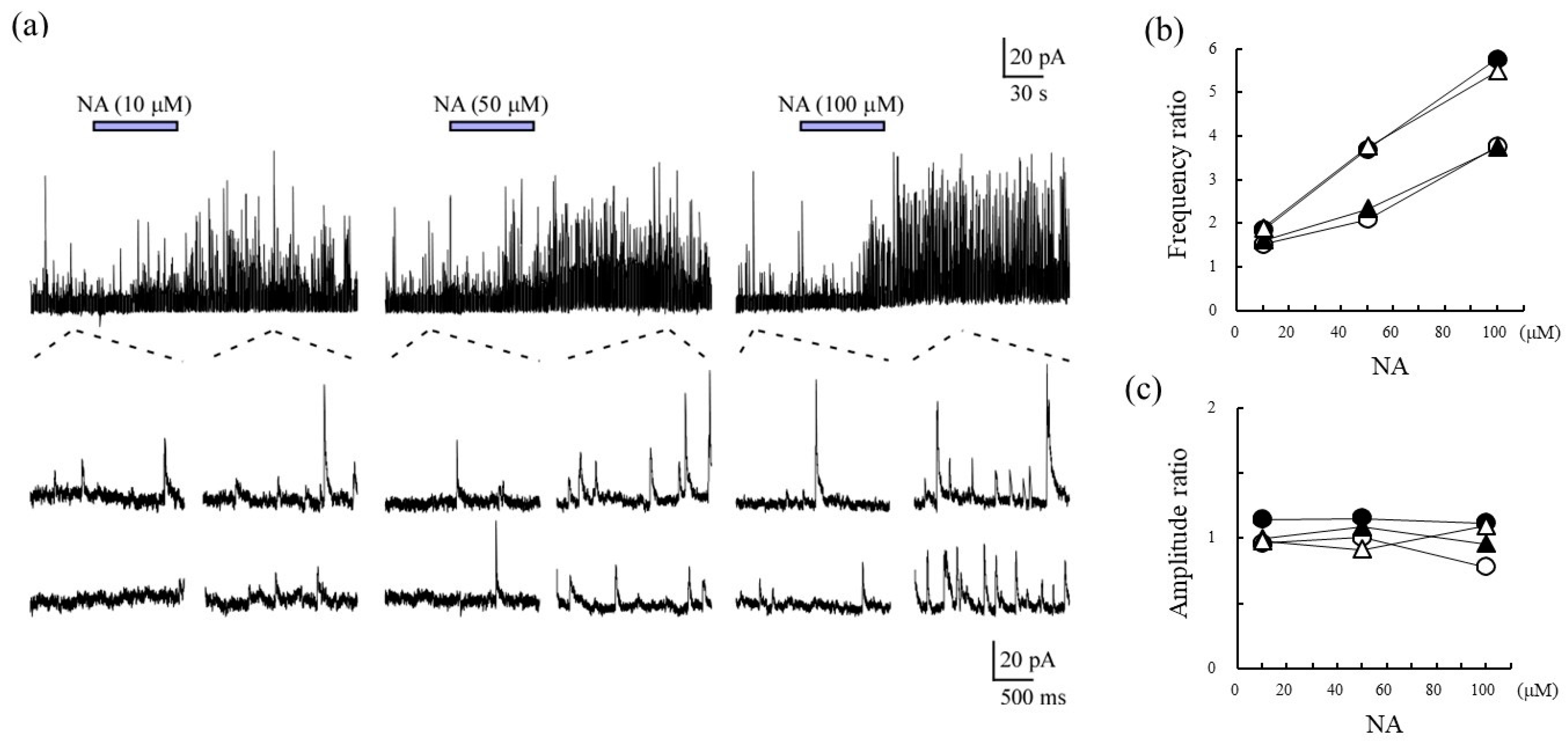
2.1. Characterization of SG Neurons for sIPSCs Responding to Noradrenalin
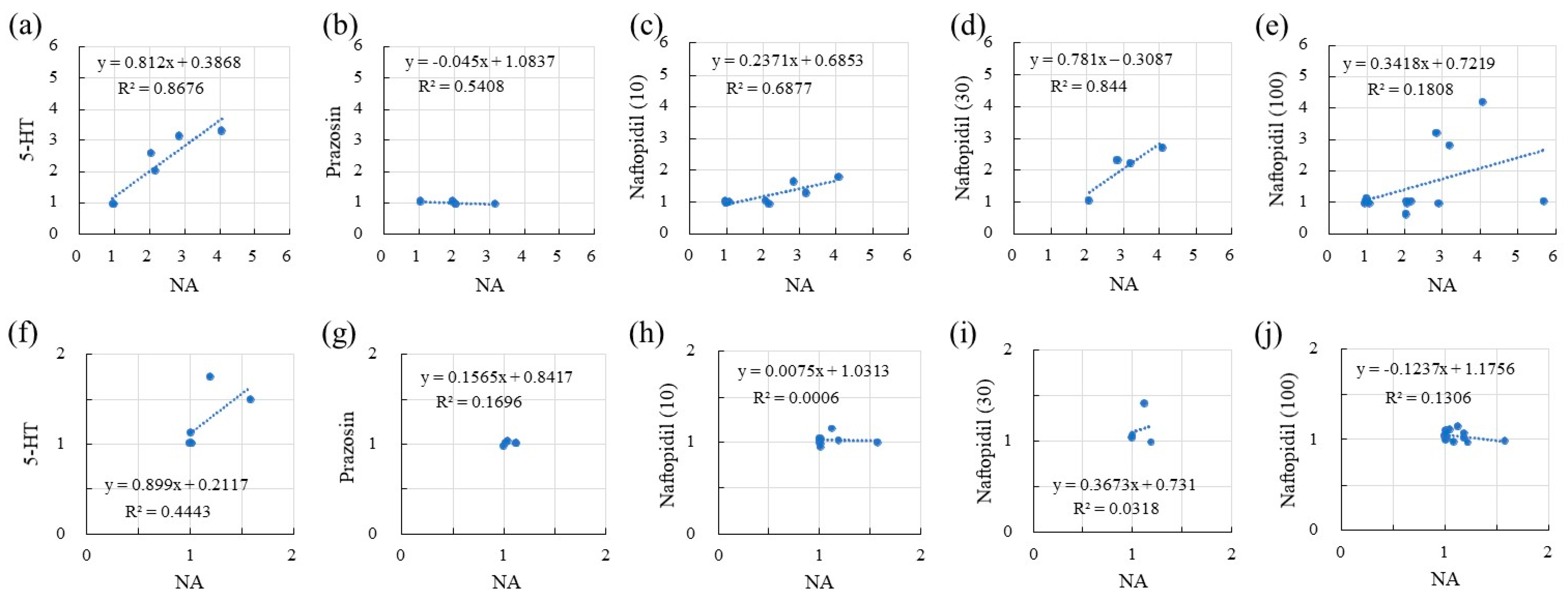
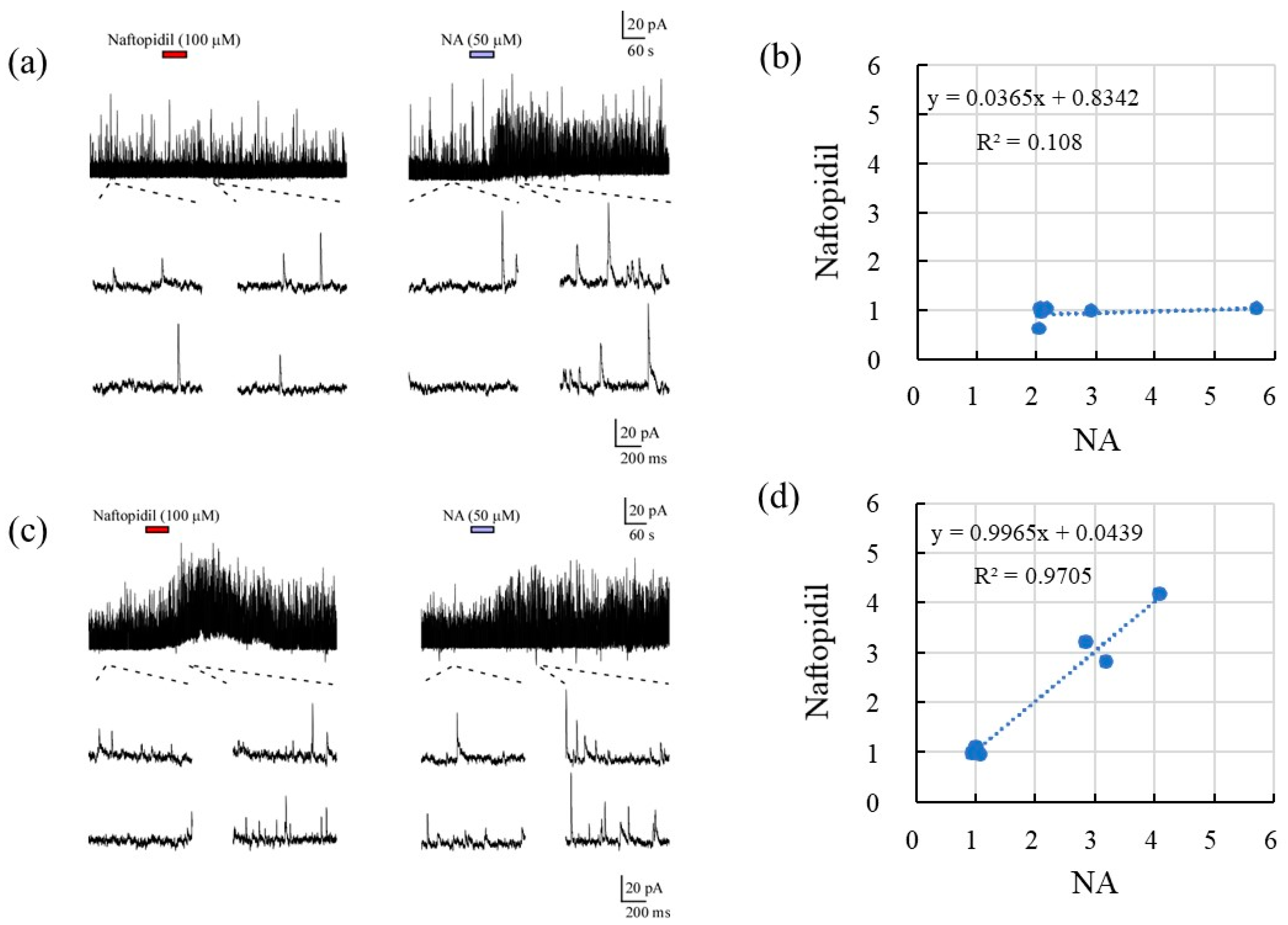
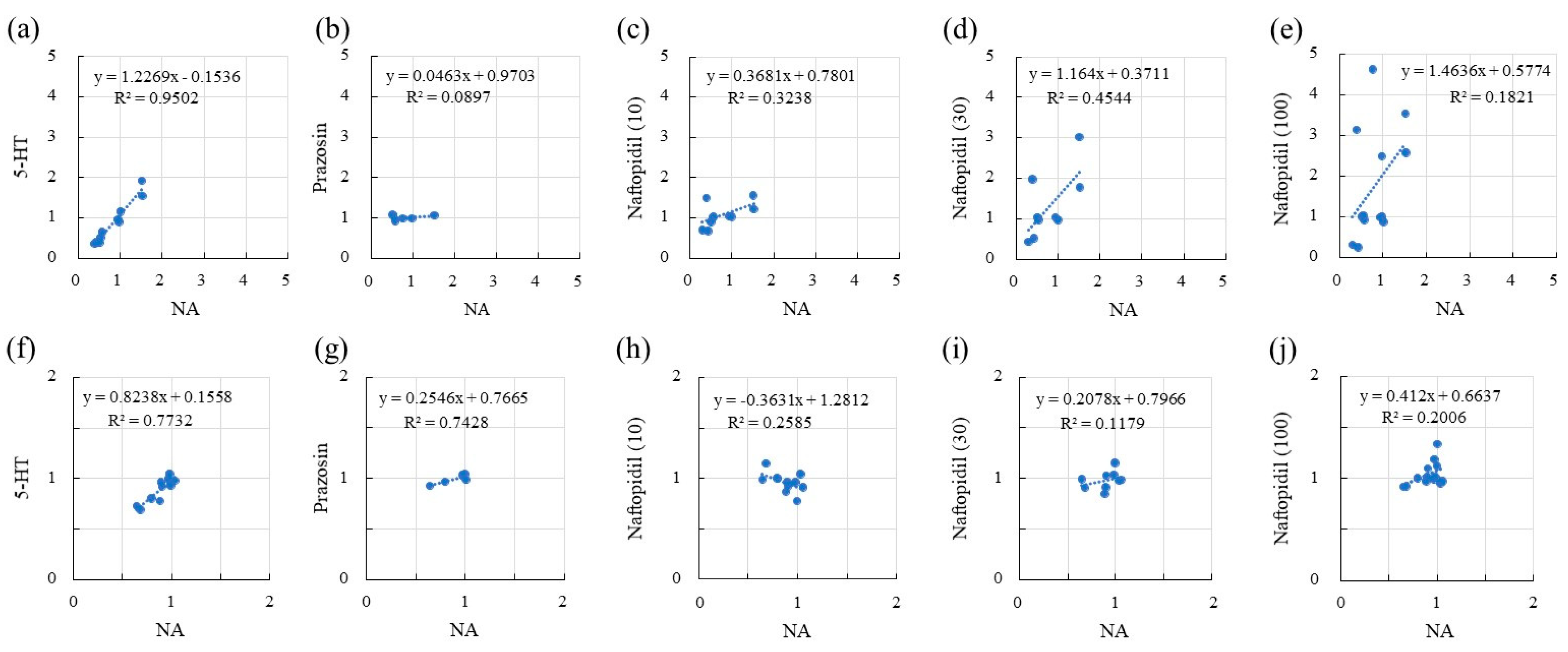
2.2. Correlation of sIPSCs Following Exposure of SGs to NA and Seratonin, Prazosin, and Naftopidil
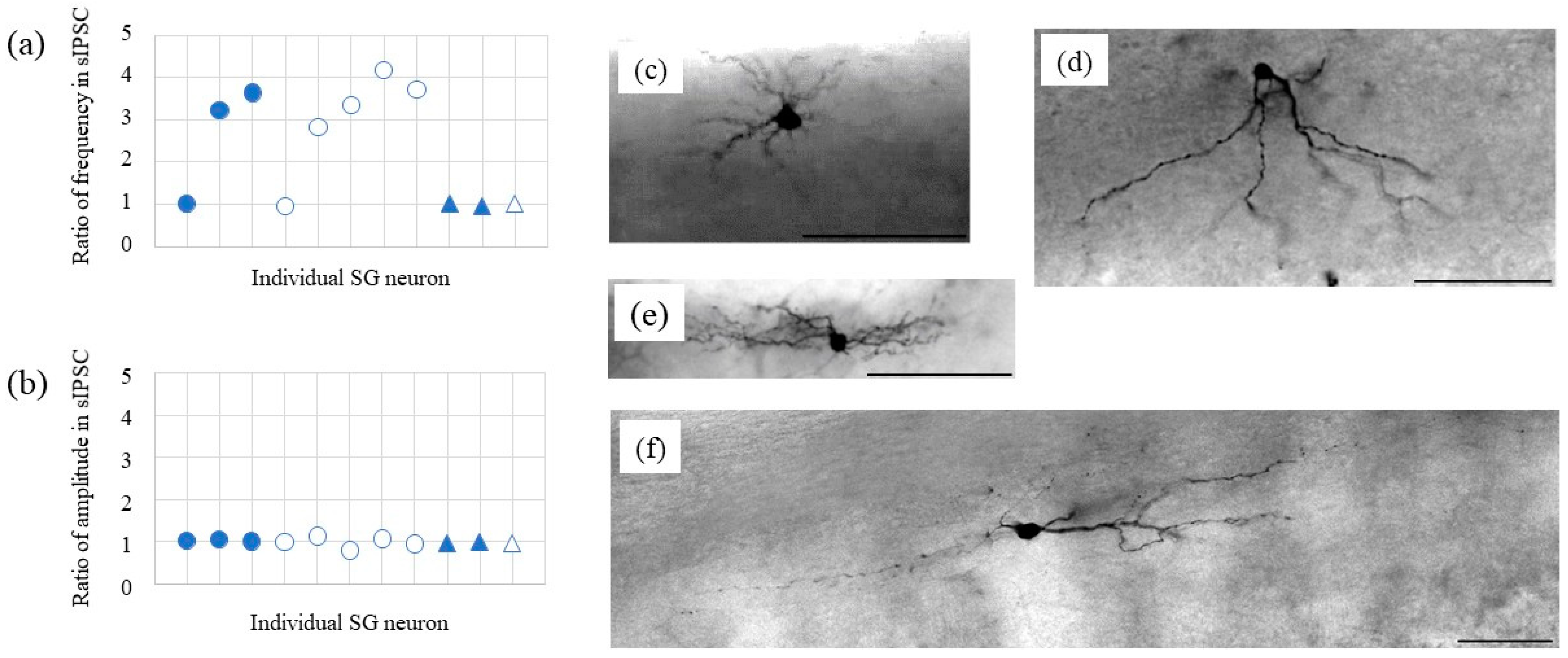
2.3. Morphological Analysis and sIPSCs
2.4. Correlation of sEPSCs between NA and 5-HT, Prazosin, and Naftopidil
3. Discussion
4. Materials and Methods
4.1. Animals and Slice Preparation
4.2. Blind Patch-Clamp Recording
4.3. Morphological Analysis
4.4. Drug Application
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Population Prospects: Ten Key Findings. Available online: https://population.un.org/wpp/Publications/Files/WPP2019_10KeyFindings.pdf (accessed on 17 May 2021).
- Kupelian, V.; Wei, J.T.; O’Leary, M.P.; Kusek, J.W.; Litman, H.J.; Link, C.L.; McKinlay, J.B. BACH Survery Investigators Prevalence of Lower Urinary Tract Symptoms and Effect on Quality of Life in a Racially and Ethnically Diverse Random Sample: The Boston Area Community Health (BACH) Survey. Arch. Intern. Med. 2006, 166, 2381–2387. [Google Scholar] [CrossRef] [Green Version]
- Abrams, P.; Cardozo, L.; Fall, M.; Griffiths, D.; Rosier, P.; Ulmsten, U.; van Kerrebroeck, P.; Victor, A.; Wein, A. Standardisation Sub-Committee of the International Continence Society The Standardisation of Terminology of Lower Urinary Tract Function: Report from the Standardisation Sub-Committee of the International Continence Society. Neurourol. Urodyn. 2002, 21, 167–178. [Google Scholar] [CrossRef] [PubMed]
- EAU Guidelines on Non-Neurogenic Male LUTS, Incl. BPO 2020. Available online: https://uroweb.org/wp-content/uploads/EAU-Guidelines-on-Non-Neurogenic-Male-LUTS-incl.-BPO-2020.pdf (accessed on 17 May 2021).
- Gotoh, M.; Kamihira, O.; Kinukawa, T.; Ono, Y.; Ohshima, S.; Origasa, H.; Tokai Urological Clinical Trial Group. Comparison of Tamsulosin and Naftopidil for Efficacy and Safety in the Treatment of Benign Prostatic Hyperplasia: A Randomized Controlled Trial. BJU Int. 2005, 96, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Takei, R.; Ikegaki, I.; Shibata, K.; Tsujimoto, G.; Asano, T. Naftopidil, a Novel alpha1-adrenoceptor Antagonist, Displays Selective Inhibition of Canine Prostatic Pressure and High Affinity Binding to Cloned Human Alpha1-Adrenoceptors. Jpn. J. Pharmacol. 1999, 79, 447–454. [Google Scholar] [PubMed] [Green Version]
- Sugaya, K.; Nishijima, S.; Miyazato, M.; Ashitomi, K.; Hatano, T.; Ogawa, Y. Effects of Intrathecal Injection of Tamsulosin and Naftopidil, Alpha-1A and -1D Adrenergic Receptor Antagonists, on Bladder Activity in Rats. Neurosci. Lett. 2002, 328, 74–76. [Google Scholar] [CrossRef]
- Chen, Z.; Ishizuka, O.; Imamura, T.; Aizawa, N.; Igawa, Y.; Nishizawa, O.; Andersson, K.E. Role of Alpha1-Adrenergic Receptors in Detrusor Overactivity Induced by Cold Stress in Conscious Rats. Neurourol. Urodyn. 2009, 28, 251–256. [Google Scholar] [CrossRef]
- Majima, T.; Yamamoto, T.; Funahashi, Y.; Takai, S.; Matsukawa, Y.; Yoshida, M.; Gotoh, M. Effect of Naftopidil on Bladder Microcirculation in a Rat Model of Bladder Outlet Obstruction. Low. Urin. Tract Symptoms 2017, 9, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Mine, S.; Yamamoto, T.; Mizuno, H.; Endo, K.; Matsukawa, Y.; Funahashi, Y.; Kato, M.; Hattori, R.; Gotoh, M. Effect of Tamsulosin on Bladder Microcirculation in Rat Model of Bladder Outlet Obstruction Using Pencil Lens Charge-Coupled Device Microscopy System. Urology 2013, 81, 155–159. [Google Scholar] [CrossRef]
- Nishino, Y.; Masue, T.; Miwa, K.; Takahashi, Y.; Ishihara, S.; Deguchi, T. Comparison of Two Alpha1-Adrenoceptor Antagonists, Naftopidil and Tamsulosin Hydrochloride, in the Treatment of Lower Urinary Tract Symptoms with Benign Prostatic Hyperplasia: A Randomized Crossover Study. BJU Int. 2006, 97, 747–751; discussion 751. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, K.; Nishijima, S.; Kadekawa, K.; Ashitomi, K.; Ueda, T.; Yamamoto, H. Spinal Mechanism of Micturition Reflex Inhibition by Naftopidil in Rats. Life Sci. 2014, 116, 106–111. [Google Scholar] [CrossRef]
- de Groat, W.C. Integrative Control of the Lower Urinary Tract: Preclinical Perspective. Br. J. Pharmacol. 2006, 147, S25–S40. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Akimoto, N.; Hakozaki, A.; Noma, T.; Nakamura, A.; Hayashi, Y.; Sasaki, E.; Ozaki, N.; Furue, H. Responsiveness of Lumbosacral Superficial Dorsal Horn Neurons During the Voiding Reflex and Functional Loss of Spinal Urethral-Responsive Neurons in Streptozotocin-Induced Diabetic Rats. Neurourol. Urodyn. 2020, 39, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Uta, D.; Xie, D.J.; Hattori, T.; Kasahara, K.I.; Yoshimura, M. Effects of Naftopidil on Inhibitory Transmission in Substantia gelatinosa Neurons of the Rat Spinal Dorsal Horn In Vitro. J. Neurol. Sci. 2017, 380, 205–211. [Google Scholar] [CrossRef]
- Uta, D.; Hattori, T.; Yoshimura, M. Differential Effects of Alpha 1-Adrenoceptor Antagonists on the Postsynaptic Sensitivity: Using Slice Patch-Clamp Technique for Inhibitory Postsynaptic Current in Substantia Gelatinosa Neurons from Lumbosacral Spinal Cord in Rats. Int. Neurourol. J. 2020, 24, 127–134. [Google Scholar] [CrossRef]
- Spike, R.C.; Todd, A.J. Ultrastructural and Immunocytochemical Study of lamina II Islet Cells in Rat Spinal Dorsal Horn. J. Comp. Neurol. 1992, 323, 359–369. [Google Scholar] [CrossRef]
- Todd, A.J.; Sullivan, A.C. Light Microscope Study of the Coexistence of GABA-Like and Glycine-Like Immunoreactivities in the Spinal Cord of the Rat. J. Comp. Neurol. 1990, 296, 496–505. [Google Scholar] [CrossRef]
- Uta, D.; Hattori, T.; Yoshimura, M. Characterization on Responsiveness of Excitatory Synaptic Transmissions to α1-Adrenoceptor Blockers in Substantia Gelatinosa Neurons Isolated from Lumbo-Sacral Level in Rat Spinal Cords. Int. Neurourol. J. 2019, 23, 13–21. [Google Scholar] [CrossRef]
- Uta, D.; Hattori, T.; Yoshimura, M. Effect of Alpha 1-Adrnoceptor Antagonists on Postsynaptic Sensitivity in Substantia Gelatinosa Neurons from Lumbosacral Spinal Cord in Rats Using Slice Patch-Clamp Technique for mEPSC. Int. Neurourol. J. 2020, 24, 135–143. [Google Scholar] [CrossRef]
- Uta, D.; Hattori, T.; Yoshimura, M. Effects of High Concentrations of Naftopidil on Dorsal Root-Evoked Excitatory Synaptic Transmissions in Substantia Gelatinosa Neurons In Vitro. Int. Neurourol. J. 2018, 22, 252–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, O.; Yusup, A.; Oyama, N.; Aoki, Y.; Tanase, K.; Matsuta, Y.; Miwa, Y.; Akino, H. Improvement of Bladder Storage Function by alpha1-Blocker Depends on the Suppression of C-Fiber Afferent Activity in Rats. Neurourol. Urodyn. 2006, 25, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Uta, D.; Furue, H.; Pickering, A.E.; Rashid, M.H.; Mizuguchi-Takase, H.; Katafuchi, T.; Imoto, K.; Yoshimura, M. TRPA1-Expressing Primary Afferents Synapse with a Morphologically Identified Subclass of Substantia gelatinosa Neurons in the Adult Rat Spinal Cord. Eur. J. Neurosci. 2010, 31, 1960–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasaka, T.; Kato, G.; Furue, H.; Rashid, M.H.; Sonohata, M.; Tamae, A.; Murata, Y.; Masuko, S.; Yoshimura, M. Cell-Type-Specific Excitatory and Inhibitory Circuits Involving Primary Afferents in the Substantia gelatinosa of the Rat Spinal Dorsal Horn In Vitro. J. Physiol. 2007, 581, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Heinke, B.; Ruscheweyh, R.; Forsthuber, L.; Wunderbaldinger, G.; Sandkühler, J. Physiological, Neurochemical and Morphological Properties of a Subgroup of GABAergic Spinal Lamina II Neurones Identified by Expression of Green Fluorescent Protein in Mice. J. Physiol. 2004, 560, 249–266. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conc. (μM) | sIPSCs | sEPSCs | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frequency | Amplitude | Frequency | Amplitude | |||||||||
| R | R2 | Slope | R | R2 | Slope | R | R2 | Slope | R | R2 | Slope | |
| 5-HT | ||||||||||||
| 50 | 0.931 | 0.868 | 0.812 | 0.667 | 0.444 | 0.899 | 0.975 | 0.950 | 1.227 | 0.879 | 0.773 | 0.824 |
| Prazosin | ||||||||||||
| 10 | 0.735 | 0.541 | −0.045 | 0.412 | 0.170 | 0.157 | 0.299 | 0.090 | 0.049 | 0.862 | 0.743 | 0.255 |
| Naftopidil | ||||||||||||
| 10 | 0.829 | 0.688 | 0.237 | 0.024 | 0.001 | −0.008 | 0.569 | 0.324 | 0.368 | 0.508 | 0.259 | −0.363 |
| 30 | 0.919 | 0.844 | 0.781 | 0.178 | 0.032 | 0.367 | 0.674 | 0.454 | 1.164 | 0.343 | 0.118 | 0.208 |
| 100 | 0.425 | 0.181 | 0.342 | 0.361 | 0.131 | −0.124 | 0.427 | 0.182 | 1.464 | 0.448 | 0.201 | 0.412 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uta, D.; Hattori, T.; Yoshimura, M. Analyses of the Mode of Action of an Alpha-Adrenoceptor Blocker in Substantia Gelatinosa Neurons in Rats. Int. J. Mol. Sci. 2021, 22, 9636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179636
Uta D, Hattori T, Yoshimura M. Analyses of the Mode of Action of an Alpha-Adrenoceptor Blocker in Substantia Gelatinosa Neurons in Rats. International Journal of Molecular Sciences. 2021; 22(17):9636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179636
Chicago/Turabian StyleUta, Daisuke, Tsuyoshi Hattori, and Megumu Yoshimura. 2021. "Analyses of the Mode of Action of an Alpha-Adrenoceptor Blocker in Substantia Gelatinosa Neurons in Rats" International Journal of Molecular Sciences 22, no. 17: 9636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179636