
Functional Activity of Recombinant Forms of Amh and Synergistic Action with Fsh in European Sea Bass Ovary

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
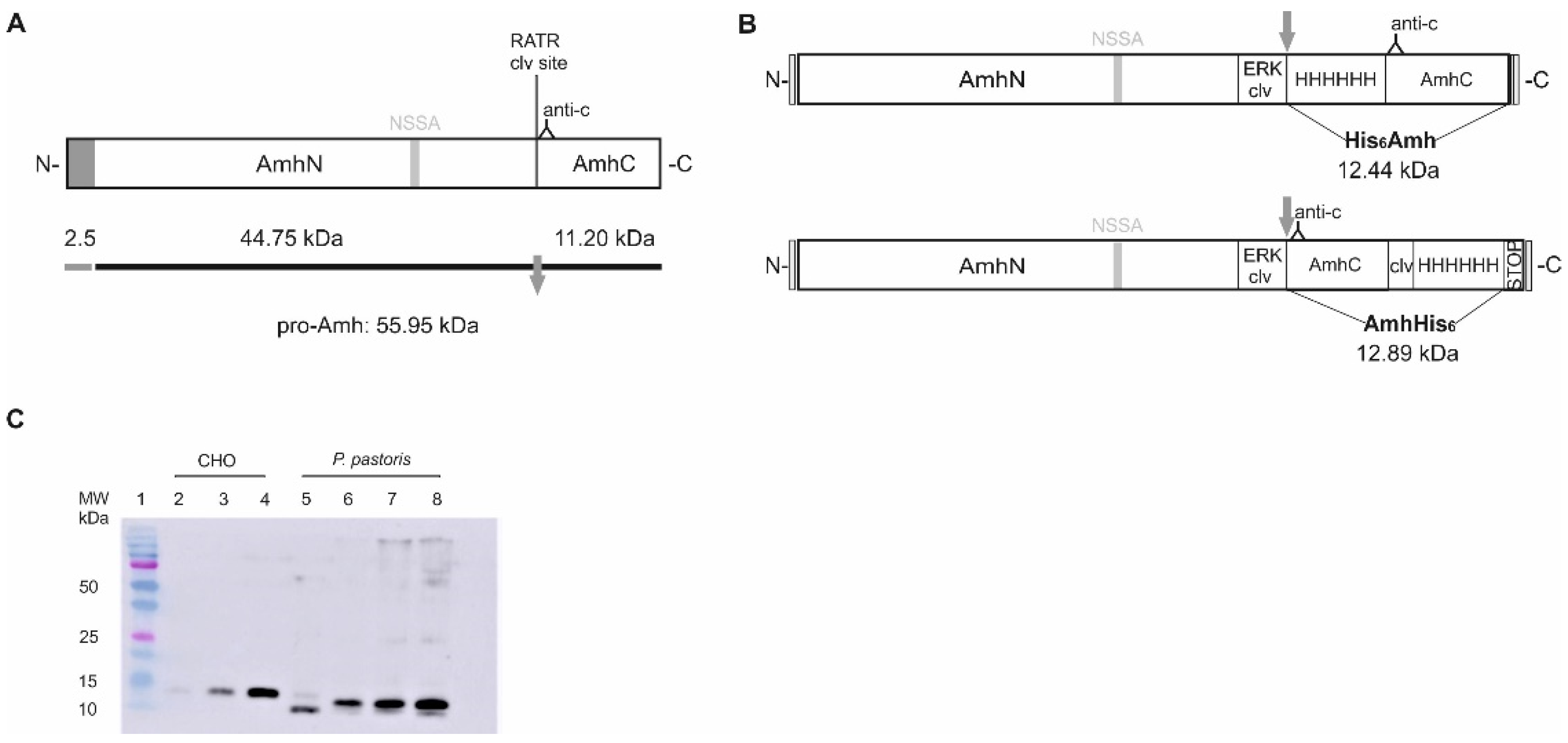
2.1. Production of Recombinant Sea Bass Amh in the Yeast P. pastoris
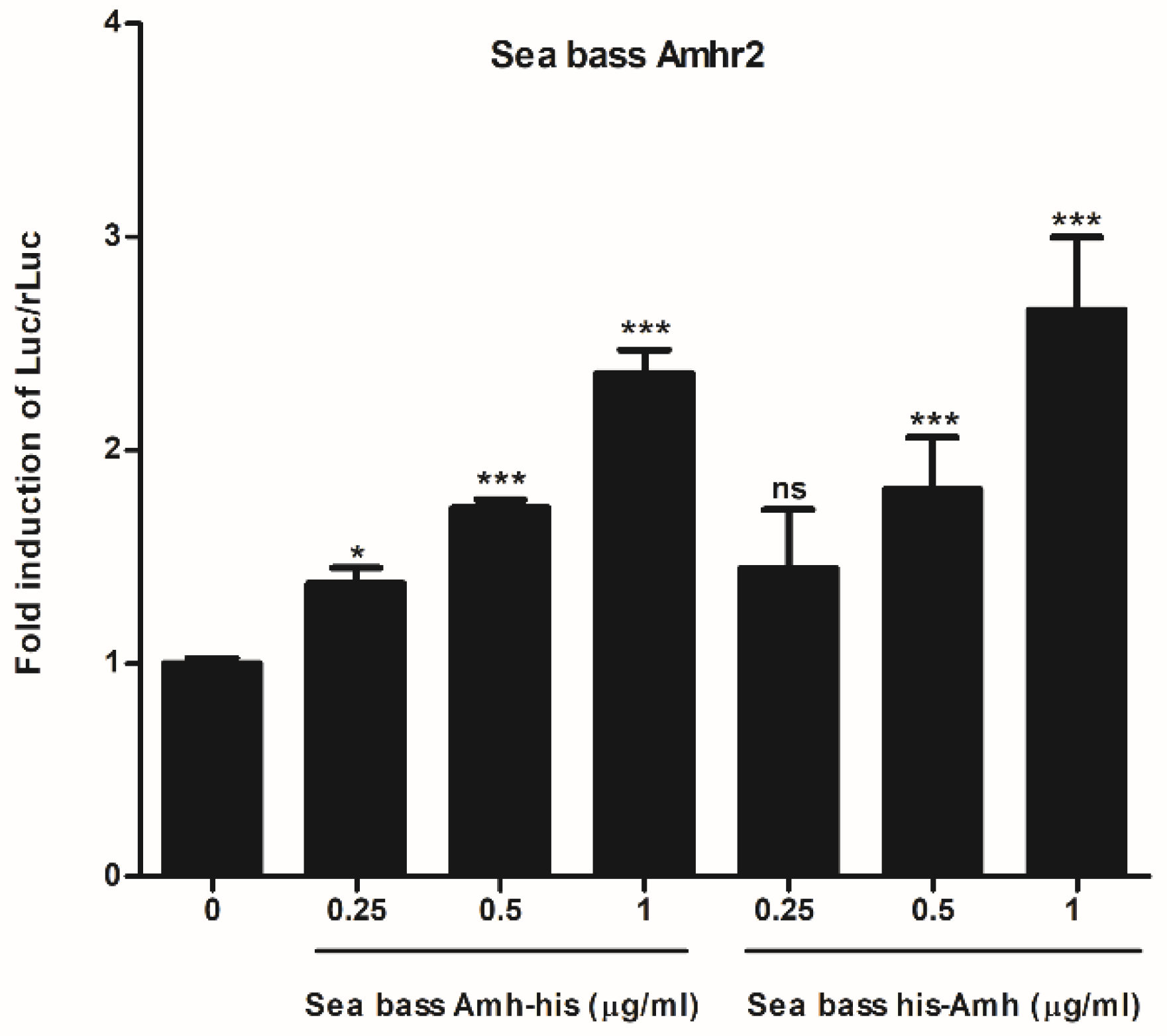
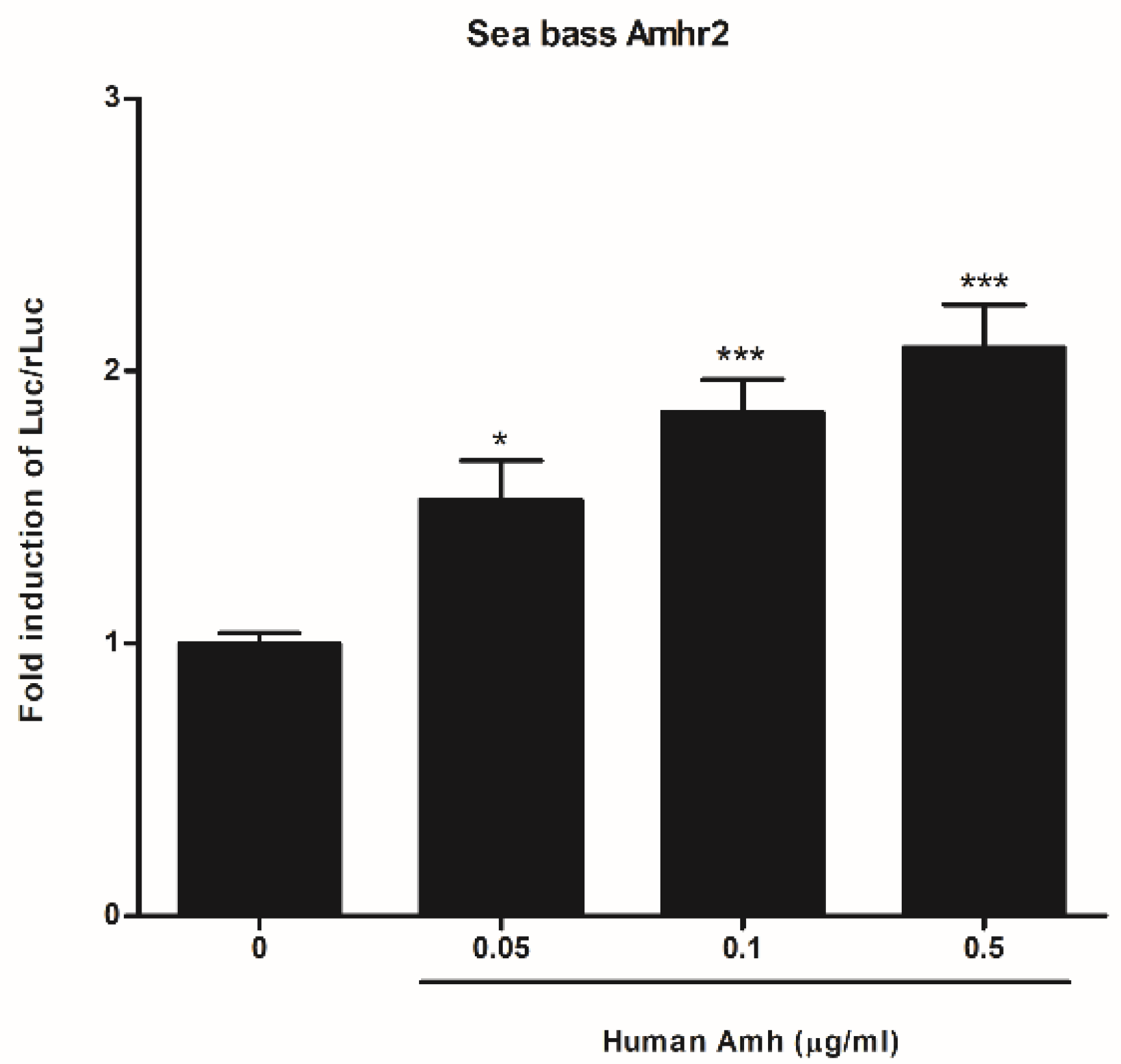
2.2. Functional Characterization of Sea Bass Amh
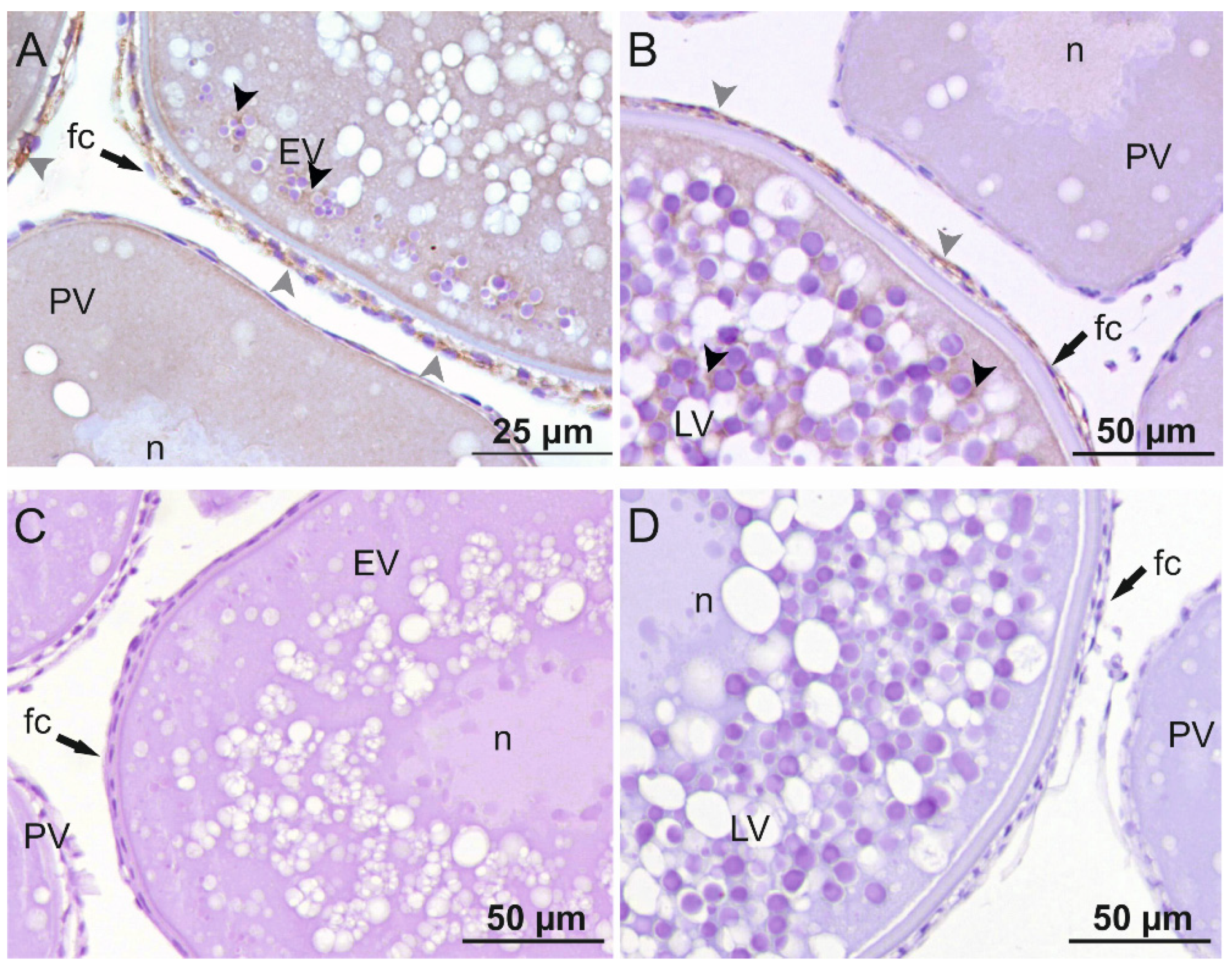
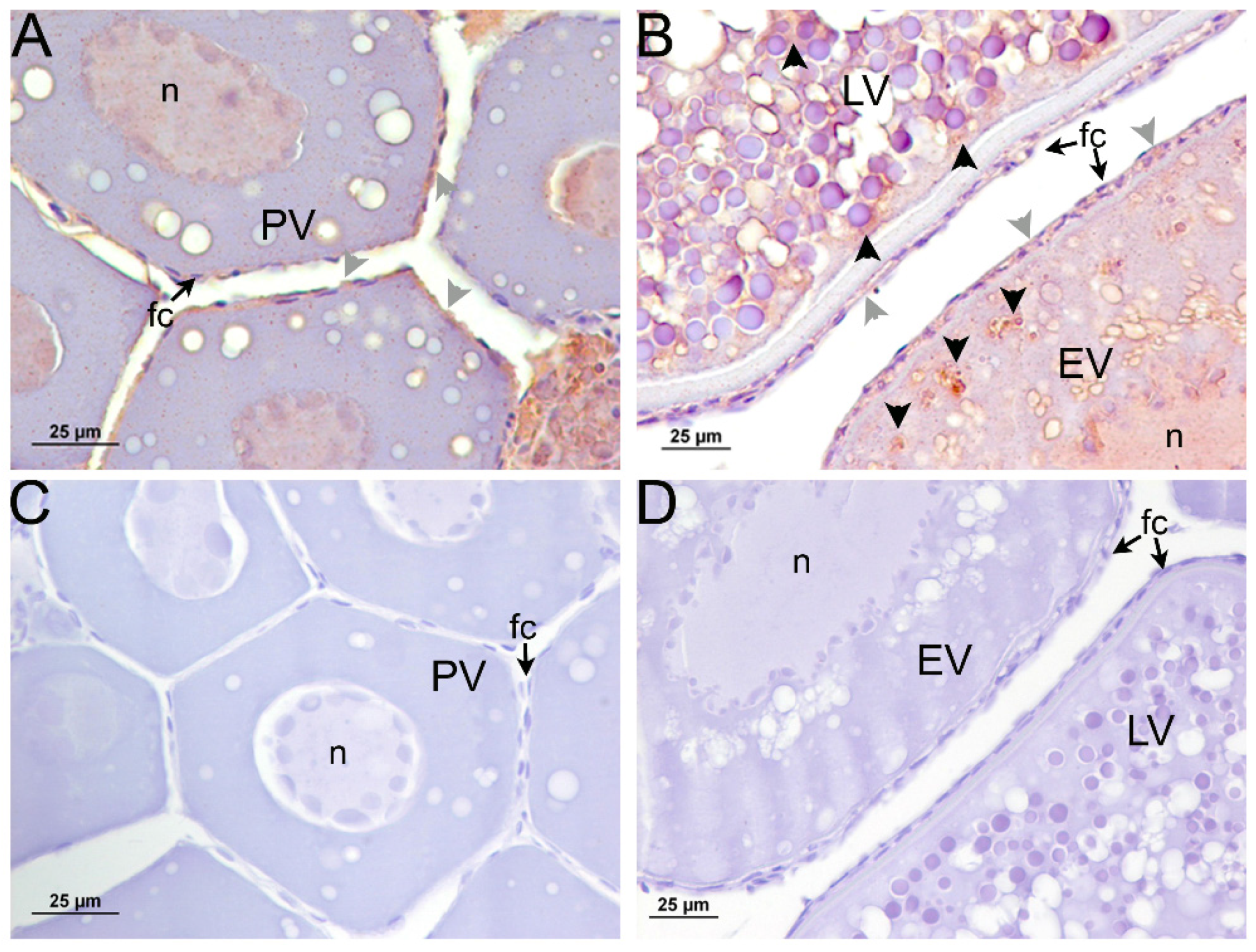
2.3. Immunolocalization of sbsAmh and sbsAmhr2 in Sea Bass Ovary
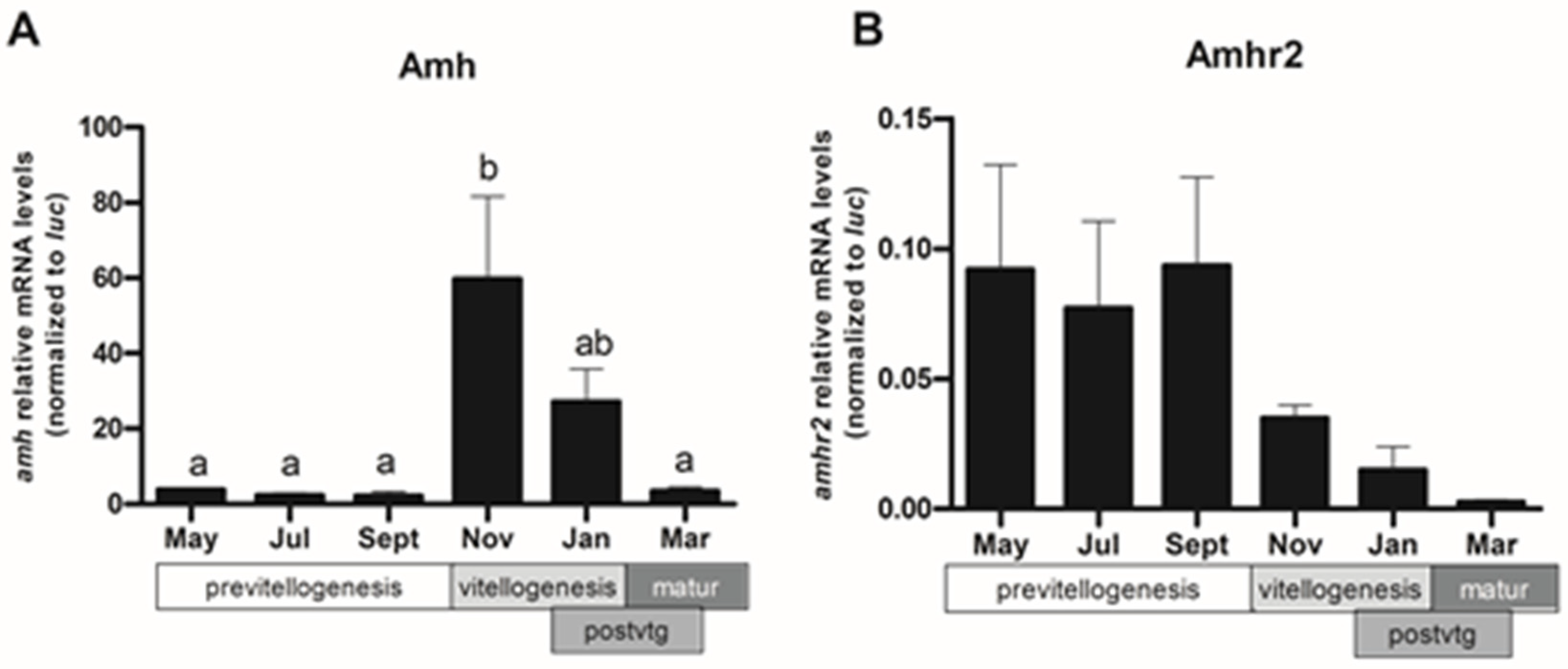
2.4. Expression Pattern of amh and amhr2 in Follicular Cells
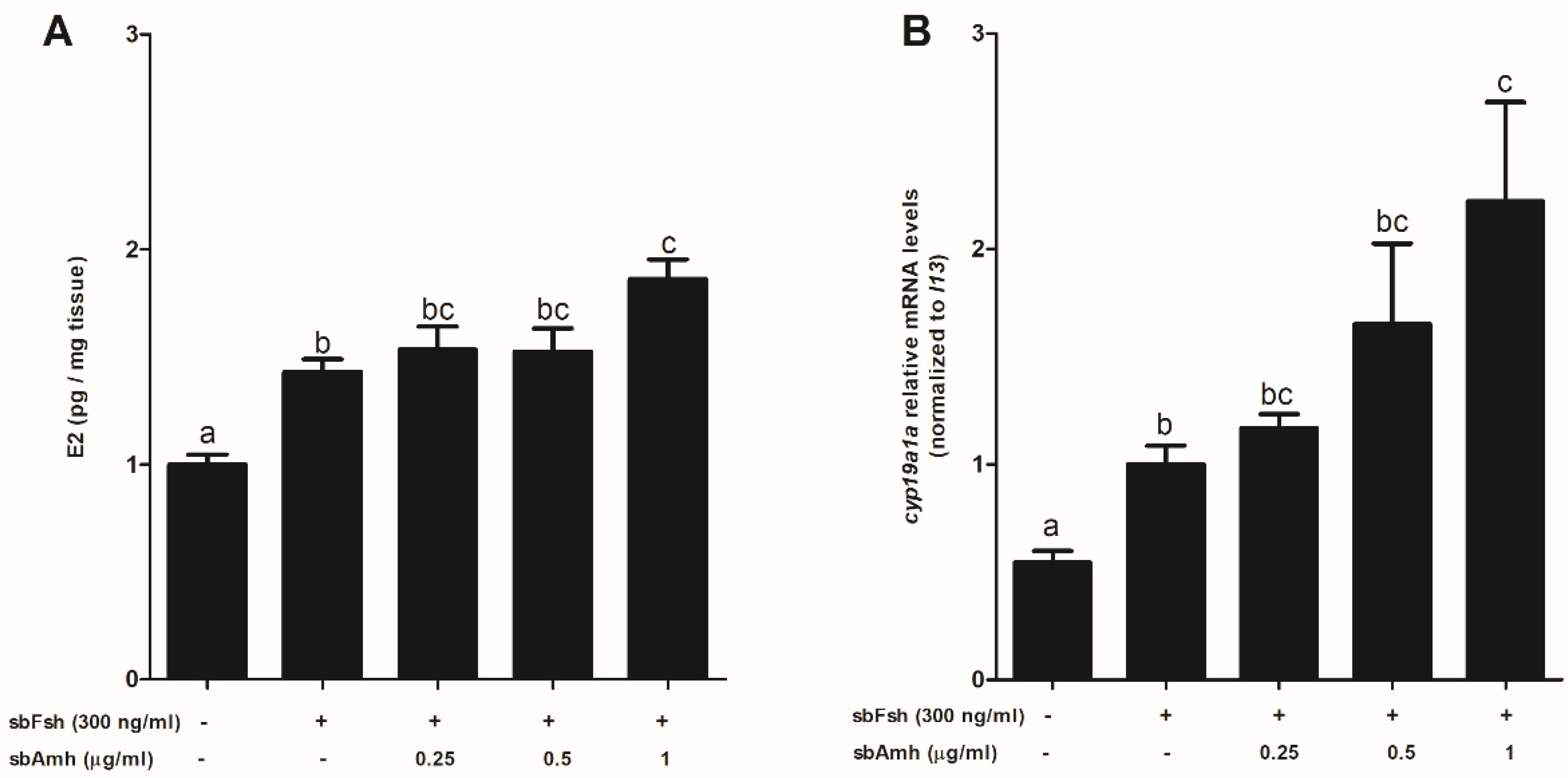
2.5. Synergistic Effect of Amh on Fsh-Induced Steroidogenesis in Previtellogeneic Ovaries
3. Discussion
4. Materials and Methods
4.1. Animal and Tissue Sampling
4.2. Expression Plasmids
4.3. Yeast Transformation and Screening for Mut+ and Muts Transformants
4.4. Large Scale Production of Recombinant Sea Bass Amh in Yeast
4.5. Sea Bass Amhr2-Directed Antibodies
4.6. Western Blot
4.7. Cell Culture, Transfection and Luciferase Assay
4.8. Detection of Endogenous Amh and Amhr2 in Adult Sea Bass Ovaries by Immunohistochemistry
4.9. Ovary Tissue Culture
4.10. Estradiol Immunoassay
4.11. Quantitative Real-Time PCR (qPCR)
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Adolfi, M.C.; Nakajima, R.T.; Nóbrega, R.H.; Schartl, M. Intersex, Hermaphroditism, and Gonadal Plasticity in Vertebrates: Evolution of the Müllerian Duct and Amh/Amhr2 Signaling. Annu. Rev. Anim. Biosci. 2019, 7, 149–172. [Google Scholar] [CrossRef] [PubMed]
- Di Clemente, N.; Jamin, S.P.; Lugovskoy, A.; Carmillo, P.; Ehrenfels, C.; Picard, J.-Y.; Whitty, A.; Josso, N.; Pepinsky, R.B.; Cate, R.L. Processing of Anti-Müllerian Hormone Regulates Receptor Activation by a Mechanism Distinct from TGF-β. Mol. Endocrinol. 2010, 24, 2193–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; He, W.W.; Shah, P.C.; Morikawa, N.; Lee, M.M.; Catlin, E.A.; Hudson, P.L.; Wing, J.; Maclaughlin, D.T.; Donahoe, P.K. Developmental expression of a candidate müllerian inhibiting substance type II receptor. Endocrinology 1996, 137, 160–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.-Y.; Protheroe, A.; Clarkson, A.N.; Imhoff, F.; Koishi, K.; McLennan, I.S. Müllerian inhibiting substance contributes to sex-linked biases in the brain and behavior. Proc. Natl. Acad. Sci. USA 2009, 106, 7203–7208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, M.; Mohapatra, B.; Urbiztondo, A.; Birusingh, R.J.; Morgado, M.; Rodriguez, M.M.; Lincoln, J.; Vatta, M. Differential Changes in TGF-β/BMP Signaling Pathway in the Right Ventricular Myocardium of Newborns with Hypoplastic Left Heart Syndrome. J. Card. Fail. 2010, 16, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Catlin, E.A.; Powell, S.M.; Manganaro, T.F.; Hudson, P.L.; Ragin, R.C.; Epstein, J.; Donahoe, P.K. Sex-specific Fetal Lung Development and Müllerian Inhibiting Substance. Am. Rev. Respir. Dis. 1990, 141, 466–470. [Google Scholar] [CrossRef]
- Jamin, S.P.; Arango, N.A.; Mishina, Y.; Hanks, M.C.; Behringer, R.R. Requirement of Bmpr1a for Müllerian duct regression during male sexual development. Nat. Genet. 2002, 32, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Di Clemente, N.; Josso, N.; Gouédard, L.; Belville, C. Components of the anti-Müllerian hormone signaling pathway in gonads. Mol. Cell. Endocrinol. 2003, 211, 9–14. [Google Scholar] [CrossRef]
- McLennan, I.S.; Pankhurst, M.W. Anti-Müllerian hormone is a gonadal cytokine with two circulating forms and cryptic actions. J. Endocrinol. 2015, 226, R45–R57. [Google Scholar] [CrossRef] [Green Version]
- Baarends, W.M.; Uilenbroek, J.T.; Kramer, P.; Hoogerbrugge, J.W.; van Leeuwen, E.C.; Themmen, A.P.; Grootegoed, J.A. Anti-müllerian hormone and anti-müllerian hormone type II receptor messenger ribonucleic acid expression in rat ovaries during postnatal development, the estrous cycle, and gonadotropin-induced follicle growth. Endocrinology 1995, 136, 4951–4962. [Google Scholar] [CrossRef]
- Baarends, W.M.; Hoogerbrugge, J.W.; Post, M.; Visser, J.A.; De Rooij, D.G.; Parvinen, M.; Themmen, A.P.; Grootegoed, J.A. Anti-müllerian hormone and anti-müllerian hormone type II receptor messenger ribonucleic acid expression during postnatal testis development and in the adult testis of the rat. Endocrinology 1995, 136, 5614–5622. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.H.; Ocón-Grove, O.M.; Johnson, A.L. Anti-Müllerian hormone (AMH) receptor type II expression and AMH activity in bovine granulosa cells. Theriogenology 2016, 86, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josso, N.; Picard, J.Y.; Rey, R.; di Clemente, N. Testicular anti-Müllerian hormone: History, genetics, regulation and clinical applications. Pediatr. Endocrinol. Rev. PER 2006, 3, 347–358. [Google Scholar]
- Visser, J.A.; Themmen, A.P.N. Anti-Müllerian hormone and folliculogenesis. Mol. Cell. Endocrinol. 2005, 234, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Vigier, B.; Forest, M.G.; Eychenne, B.; Bezard, J.; Garrigou, O.; Robel, P.; Josso, N. Anti-Mullerian hormone produces endocrine sex reversal of fetal ovaries. Proc. Natl. Acad. Sci. USA 1989, 86, 3684–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouiller-Fabre, V.; Carmona, S.; Merhi, R.A.; Cate, R.; Habert, R.; Vigier, B. Effect of anti-Mullerian hormone on Sertoli and Leydig cell functions in fetal and immature rats. Endocrinology 1998, 139, 1213–1220. [Google Scholar] [CrossRef]
- Rey, R.; Lukas-Croisier, C.; Lasala, C.; Bedecarrás, P. AMH/MIS: What we know already about the gene, the protein and its regulation. Mol. Cell. Endocrinol. 2003, 211, 21–31. [Google Scholar] [CrossRef]
- Durlinger, A.L.L.; Gruijters, M.J.G.; Kramer, P.; Karels, B.; Ingraham, H.A.; Nachtigal, M.W.; Uilenbroek, J.T.J.; Grootegoed, J.A.; Themmen, A.P.N. Anti-Müllerian hormone inhibits initiation of primordial follicle growth in the mouse ovary. Endocrinology 2002, 143, 1076–1084. [Google Scholar] [CrossRef]
- Miura, T.; Miura, C.; Konda, Y.; Yamauchi, K. Spermatogenesis-preventing substance in Japanese eel. Development 2002, 129, 2689–2697. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Marí, A.; Yan, Y.-L.; Bremiller, R.A.; Wilson, C.; Cañestro, C.; Postlethwait, J.H. Characterization and expression pattern of zebrafish Anti-Müllerian hormone (Amh) relative to sox9a, sox9b, and cyp19a1a, during gonad development. Gene Expr. Patterns GEP 2005, 5, 655–667. [Google Scholar] [CrossRef]
- Halm, S.; Rocha, A.; Miura, T.; Prat, F.; Zanuy, S. Anti-Müllerian hormone (AMH/AMH) in the European sea bass: Its gene structure, regulatory elements, and the expression of alternatively-spliced isoforms. Gene 2007, 388, 148–158. [Google Scholar] [CrossRef]
- Klüver, N.; Pfennig, F.; Pala, I.; Storch, K.; Schlieder, M.; Froschauer, A.; Gutzeit, H.O.; Schartl, M. Differential expression of anti-Müllerian hormone (amh) and anti-Müllerian hormone receptor type II (amhrII) in the teleost medaka. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 271–281. [Google Scholar] [CrossRef]
- Pfennig, F.; Standke, A.; Gutzeit, H.O. The role of Amh signaling in teleost fish--Multiple functions not restricted to the gonads. Gen. Comp. Endocrinol. 2015, 223, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, N.; Shiraishi, E.; Yamamoto, T.; Iguchi, T.; Abe, S.; Kitano, T. Sexually dimorphic expression of a teleost homologue of Müllerian inhibiting substance during gonadal sex differentiation in Japanese flounder, Paralichthys olivaceus. Biochem. Biophys. Res. Commun. 2004, 322, 508–513. [Google Scholar] [CrossRef]
- Wang, X.G.; Orban, L. Anti-Müllerian hormone and 11 beta-hydroxylase show reciprocal expression to that of aromatase in the transforming gonad of zebrafish males. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 1329–1338. [Google Scholar] [CrossRef]
- Maugars, G.; Schmitz, M. Gene expression profiling during spermatogenesis in early maturing male Atlantic salmon parr testes. Gen. Comp. Endocrinol. 2008, 159, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Morinaga, C.; Saito, D.; Nakamura, S.; Sasaki, T.; Asakawa, S.; Shimizu, N.; Mitani, H.; Furutani-Seiki, M.; Tanaka, M.; Kondoh, H. The hotei mutation of medaka in the anti-Mullerian hormone receptor causes the dysregulation of germ cell and sexual development. Proc. Natl. Acad. Sci. USA 2007, 104, 9691–9696. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Watakabe, I.; Nishimura, T.; Picard, J.-Y.; Toyoda, A.; Taniguchi, Y.; di Clemente, N.; Tanaka, M. Hyperproliferation of mitotically active germ cells due to defective anti-Müllerian hormone signaling mediates sex reversal in medaka. Development 2012, 139, 2283–2287. [Google Scholar] [CrossRef] [Green Version]
- Pala, I.; Klüver, N.; Thorsteinsdóttir, S.; Schartl, M.; Coelho, M.M. Expression pattern of anti-Müllerian hormone (amh) in the hybrid fish complex of Squalius alburnoides. Gene 2008, 410, 249–258. [Google Scholar] [CrossRef]
- Rocha, A.; Zanuy, S.; Gómez, A. Conserved anti-müllerian hormone: Anti-müllerian hormone type-2 receptor specific interaction and intracellular signaling in teleosts. Biol. Reprod. 2016, 94, 141-1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, W.; Su, M.; Zhang, J. Effects of recombinant gonadotropin hormones on the gonadal maturation in the spotted scat, Scatophagus argus. Aquaculture 2018, 483, 263–272. [Google Scholar] [CrossRef]
- Skaar, K.S.; Nóbrega, R.H.; Magaraki, A.; Olsen, L.C.; Schulz, R.W.; Male, R. Proteolytically activated, recombinant anti-mullerian hormone inhibits androgen secretion, proliferation, and differentiation of spermatogonia in adult zebrafish testis organ cultures. Endocrinology 2011, 152, 3527–3540. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Mei, J.; Li, Z.; Zhang, X.; Zhou, L.; Gui, J.-F. Distinct and Cooperative Roles of amh and dmrt1 in Self-Renewal and Differentiation of Male Germ Cells in Zebrafish. Genetics 2017, 207, 1007–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.-L.; Batzel, P.; Titus, T.; Sydes, J.; Desvignes, T.; BreMiller, R.; Draper, B.; Postlethwait, J.H. A Hormone That Lost Its Receptor: Anti-Müllerian Hormone (AMH) in Zebrafish Gonad Development and Sex Determination. Genetics 2019, 213, 529–553. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xiao, H.; Jie, M.; Dai, S.; Wu, X.; Li, M.; Wang, D. Amh regulate female folliculogenesis and fertility in a dose-dependent manner through Amhr2 in Nile tilapia. Mol. Cell. Endocrinol. 2020, 499, 110593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, B.; Chen, W.; Ge, W. Anti-Müllerian hormone (Amh/amh) plays dual roles in maintaining gonadal homeostasis and gametogenesis in zebrafish. Mol. Cell. Endocrinol. 2020, 517, 110963. [Google Scholar] [CrossRef]
- Fairlie, W.D.; Zhang, H.; Brown, P.K.; Russell, P.K.; Bauskin, A.R.; Breit, S.N. Expression of a TGF-beta superfamily protein, macrophage inhibitory cytokine-1, in the yeast Pichia pastoris. Gene 2000, 254, 67–76. [Google Scholar] [CrossRef]
- Papakonstantinou, T.; Harris, S.J.; Fredericks, D.; Harrison, C.; Wallace, E.M.; Hearn, M.T.W. Synthesis, purification and bioactivity of recombinant human activin A expressed in the yeast Pichia pastoris. Protein Expr. Purif. 2009, 64, 131–138. [Google Scholar] [CrossRef]
- Fredericks, D.; Clay, R.; Warner, T.; O’Connor, A.; de Kretser, D.M.; Hearn, M.T.W. Optimization of the expression of recombinant human activin A in the yeast Pichia pastoris. Biotechnol. Prog. 2010, 26, 372–383. [Google Scholar] [CrossRef]
- Pulkki, M.M.; Myllymaa, S.; Pasternack, A.; Lun, S.; Ludlow, H.; Al-Qahtani, A.; Korchynskyi, O.; Groome, N.; Juengel, J.L.; Kalkkinen, N.; et al. The bioactivity of human bone morphogenetic protein-15 is sensitive to C-terminal modification: Characterization of the purified untagged processed mature region. Mol. Cell. Endocrinol. 2011, 332, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Pépin, D.; Hoang, M.; Nicolaou, F.; Hendren, K.; Benedict, L.A.; Al-Moujahed, A.; Sosulski, A.; Marmalidou, A.; Vavvas, D.; Donahoe, P.K. An albumin leader sequence coupled with a cleavage site modification enhances the yield of recombinant C-terminal Mullerian Inhibiting Substance. Technology 2013, 1, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.-C.; Li, H.-W.; Luo, J.-W.; Chen, C.; Chang, C.-F. The Potential Role of Amh to Prevent Ectopic Female Development in Testicular Tissue of the Protandrous Black Porgy, Acanthopagrus schlegelii. Biol. Reprod. 2015, 92, 158. [Google Scholar] [CrossRef] [PubMed]
- Macauley-Patrick, S.; Fazenda, M.L.; McNeil, B.; Harvey, L.M. Heterologous protein production using the Pichia pastoris expression system. Yeast 2005, 22, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.B.; Ng, S.K. Impact of host cell line choice on glycan profile. Crit. Rev. Biotechnol. 2018, 38, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Bézard, J.; Vigier, B.; Tran, D.; Mauléon, P.; Josso, N. Immunocytochemical study of anti-Müllerian hormone in sheep ovarian follicles during fetal and post-natal development. J. Reprod. Fertil. 1987, 80, 509–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weenen, C.; Laven, J.S.E.; Von Bergh, A.R.M.; Cranfield, M.; Groome, N.P.; Visser, J.A.; Kramer, P.; Fauser, B.C.J.M.; Themmen, A.P.N. Anti-Müllerian hormone expression pattern in the human ovary: Potential implications for initial and cyclic follicle recruitment. Mol. Hum. Reprod. 2004, 10, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, S.A.; Hardy, K.; Da Silva-Buttkus, P.; Stark, J.; Webber, L.J.; Flanagan, A.M.; Themmen, A.P.N.; Visser, J.A.; Groome, N.P.; Franks, S. Anti-müllerian hormone protein expression is reduced during the initial stages of follicle development in human polycystic ovaries. J. Clin. Endocrinol. Metab. 2005, 90, 5536–5543. [Google Scholar] [CrossRef] [Green Version]
- Rocha, R.M.P.; Lima, L.F.; Carvalho, A.A.; Chaves, R.N.; Bernuci, M.P.; Rosa-e-Silva, A.C.J.S.; Rodrigues, A.P.R.; Campello, C.C.; Figueiredo, J.R. Immunolocalization of the Anti-Müllerian Hormone (AMH) in Caprine Follicles and the Effects of AMH on In Vitro Culture of Caprine Pre-antral Follicles Enclosed in Ovarian Tissue. Reprod. Domest. Anim. 2016, 51, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Modi, D.; Bhartiya, D.; Puri, C. Developmental expression and cellular distribution of Mullerian inhibiting substance in the primate ovary. Reproduction 2006, 132, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.L.T.; Kryger-Baggesen, N.; Byskov, A.G.; Andersen, C.Y. Anti-Müllerian hormone initiates growth of human primordial follicles in vitro. Mol. Cell. Endocrinol. 2005, 234, 87–93. [Google Scholar] [CrossRef]
- Han, Y.; Peng, C.; Wang, L.; Guo, J.; Lu, M.; Chen, J.; Liu, Y.; Li, S.; Zhao, M.; Zhang, Y.; et al. Female-to-male sex reversal in orange-spotted grouper (Epinephelus coioides) caused by overexpressing of Amh in vivo. Biol. Reprod. 2018, 99, 1205–1215. [Google Scholar] [CrossRef] [Green Version]
- Von Schalburg, K.R.; Gowen, B.E.; Rondeau, E.B.; Johnson, N.W.; Minkley, D.R.; Leong, J.S.; Davidson, W.S.; Koop, B.F. Sex-specific expression, synthesis and localization of aromatase regulators in one-year-old Atlantic salmon ovaries and testes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2013, 164, 236–246. [Google Scholar] [CrossRef]
- Ingraham, H.A.; Hirokawa, Y.; Roberts, L.M.; Mellon, S.H.; McGee, E.; Nachtigal, M.W.; Visser, J.A. Autocrine and paracrine Müllerian inhibiting substance hormone signaling in reproduction. Recent Prog. Horm. Res. 2000, 55, 53–68. [Google Scholar]
- McGee, E.A.; Smith, R.; Spears, N.; Nachtigal, M.W.; Ingraham, H.; Hsueh, A.J. Müllerian inhibitory substance induces growth of rat preantral ovarian follicles. Biol. Reprod. 2001, 64, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Cheon, K.Y.; Chung, Y.J.; Cho, H.H.; Kim, M.R.; Cha, J.H.; Kang, C.S.; Lee, J.Y.; Kim, J.H. Expression of Müllerian-Inhibiting Substance/Anti-Müllerian Hormone Type II Receptor in the Human Theca Cells. J. Clin. Endocrinol. Metab. 2018, 103, 3376–3385. [Google Scholar] [CrossRef]
- Von Hofsten, J.; Larsson, A.; Olsson, P.E. Novel steroidogenic factor-1 homolog (ff1d) is coexpressed with anti-Mullerian hormone (AMH) in zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2005, 233, 595–604. [Google Scholar] [CrossRef]
- Morais, R.D.V.S.; Crespo, D.; Nóbrega, R.H.; Lemos, M.S.; van de Kant, H.J.G.; de França, L.R.; Male, R.; Bogerd, J.; Schulz, R.W. Antagonistic regulation of spermatogonial differentiation in zebrafish (Danio rerio) by Igf3 and Amh. Mol. Cell. Endocrinol. 2017, 454, 112–124. [Google Scholar] [CrossRef] [Green Version]
- Fernandino, J.I.; Hattori, R.S.; Kimura, H.; Strüssmann, C.A.; Somoza, G.M. Expression profile and estrogenic regulation of anti-Müllerian hormone during gonadal development in pejerrey Odontesthes bonariensis, a teleost fish with strong temperature-dependent sex determination. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2008, 237, 3192–3199. [Google Scholar] [CrossRef]
- Johnsen, H.; Tveiten, H.; Torgersen, J.S.; Andersen, Ø. Divergent and sex-dimorphic expression of the paralogs of the Sox9-Amh-Cyp19a1 regulatory cascade in developing and adult atlantic cod (Gadus morhua L.). Mol. Reprod. Dev. 2013, 80, 358–370. [Google Scholar] [CrossRef]
- Luckenbach, J.A.; Iliev, D.B.; Goetz, F.W.; Swanson, P. Identification of differentially expressed ovarian genes during primary and early secondary oocyte growth in coho salmon, Oncorhynchus kisutch. Reprod. Biol. Endocrinol. RBE 2008, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Rocha, A.; Zanuy, S.; Carrillo, M.; Gómez, A. Seasonal changes in gonadal expression of gonadotropin receptors, steroidogenic acute regulatory protein and steroidogenic enzymes in the European sea bass. Gen. Comp. Endocrinol. 2009, 162, 265–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molés, G.; Gómez, A.; Carrillo, M.; Zanuy, S. Development of a homologous enzyme-linked immunosorbent assay for European sea bass FSH. Reproductive cycle plasma levels in both sexes and in yearling precocious and non-precocious males. Gen. Comp. Endocrinol. 2012, 176, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Molés, G.; Zanuy, S.; Muñoz, I.; Crespo, B.; Martínez, I.; Mañanós, E.; Gómez, A. Receptor Specificity and Functional Comparison of Recombinant Sea Bass (Dicentrarchus labrax) Gonadotropins (Fsh and Lh) Produced in Different Host Systems1. Biol. Reprod. 2011, 84, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durlinger, A.L.; Gruijters, M.J.; Kramer, P.; Karels, B.; Kumar, T.R.; Matzuk, M.M.; Rose, U.M.; de Jong, F.H.; Uilenbroek, J.T.; Grootegoed, J.A.; et al. Anti-Müllerian hormone attenuates the effects of FSH on follicle development in the mouse ovary. Endocrinology 2001, 142, 4891–4899. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Kramer, P.; Karels, B.; de Jong, F.H.; Uilenbroek, J.T.; Grootegoed, J.A.; Themmen, A.P. Control of primordial follicle recruitment by anti-Müllerian hormone in the mouse ovary. Endocrinology 1999, 140, 5789–5796. [Google Scholar] [CrossRef]
- Grossman, M.P.; Nakajima, S.T.; Fallat, M.E.; Siow, Y. Müllerian-inhibiting substance inhibits cytochrome P450 aromatase activity in human granulosa lutein cell culture. Fertil. Steril. 2008, 89, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Crespo, B.; Zanuy, S.; Gómez, A. Development of an in vitro system for functional studies of ovarian follicular cells in European sea bass (Dicentrarchus labrax). Cytotechnology 2013, 65, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Asturiano, J.F.; Sorbera, L.A.; Ramos, J.; Kime, D.E.; Carrilo, M.; Zanuy, S. Hormonal regulation of the European sea bass reproductive cycle: An individualized female approach. J. Fish Biol. 2000, 56, 1155–1172. [Google Scholar] [CrossRef]
- Don, R.H.; Cox, P.T.; Wainwright, B.J.; Baker, K.; Mattick, J.S. “Touchdown” PCR to circumvent spurious priming during gene amplification. Nucleic Acids Res. 1991, 19, 4008. [Google Scholar] [CrossRef] [Green Version]
- Korchynskyi, O.; ten Dijke, P. Identification and functional characterization of distinct critically important bone morphogenetic protein-specific response elements in the Id1 promoter. J. Biol. Chem. 2002, 277, 4883–4891. [Google Scholar] [CrossRef] [Green Version]
- Prat, F.; Zanuy, S.; Carrillo, M.; de Mones, A.; Fostier, A. Seasonal changes in plasma levels of gonadal steroids of sea bass, Dicentrarchus labrax L. Gen. Comp. Endocrinol. 1990, 78, 361–373. [Google Scholar] [CrossRef]
- Mitter, K.; Kotoulas, G.; Magoulas, A.; Mulero, V.; Sepulcre, P.; Figueras, A.; Novoa, B.; Sarropoulou, E. Evaluation of candidate reference genes for QPCR during ontogenesis and of immune-relevant tissues of European seabass (Dicentrarchus labrax). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 153, 340–347. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zapater, C.; Rocha, A.; Molés, G.; Mascoli, A.; Ibañez, S.; Zanuy, S.; Gómez, A. Functional Activity of Recombinant Forms of Amh and Synergistic Action with Fsh in European Sea Bass Ovary. Int. J. Mol. Sci. 2021, 22, 10092. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810092
Zapater C, Rocha A, Molés G, Mascoli A, Ibañez S, Zanuy S, Gómez A. Functional Activity of Recombinant Forms of Amh and Synergistic Action with Fsh in European Sea Bass Ovary. International Journal of Molecular Sciences. 2021; 22(18):10092. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810092
Chicago/Turabian StyleZapater, Cinta, Ana Rocha, Gregorio Molés, Alessia Mascoli, Soledad Ibañez, Silvia Zanuy, and Ana Gómez. 2021. "Functional Activity of Recombinant Forms of Amh and Synergistic Action with Fsh in European Sea Bass Ovary" International Journal of Molecular Sciences 22, no. 18: 10092. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810092