
Endothelium-Derived Hyperpolarizing Factor (EDHF) Mediates Acetylsalicylic Acid (Aspirin) Vasodilation of Pregnant Rat Mesenteric Arteries


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
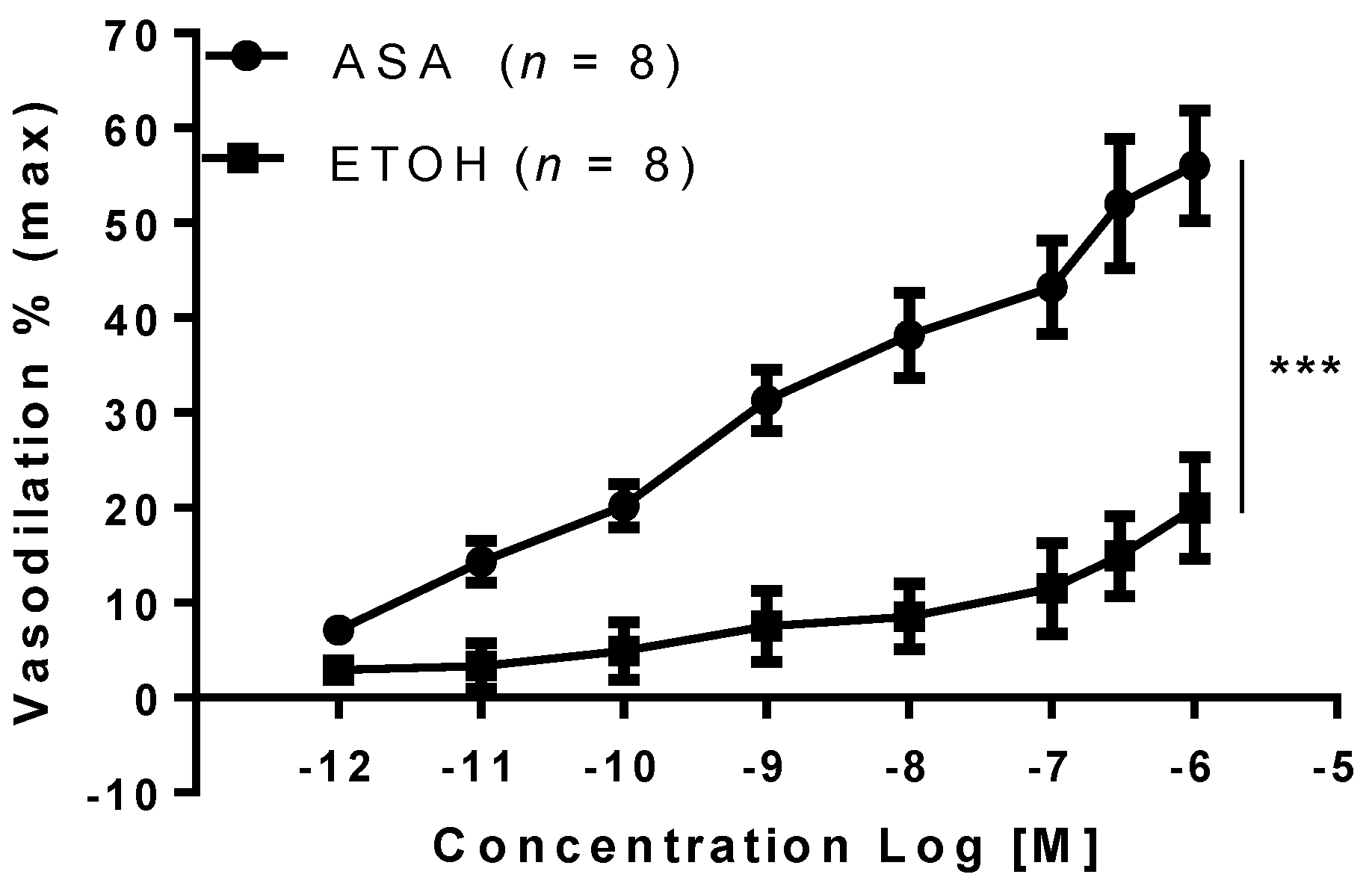
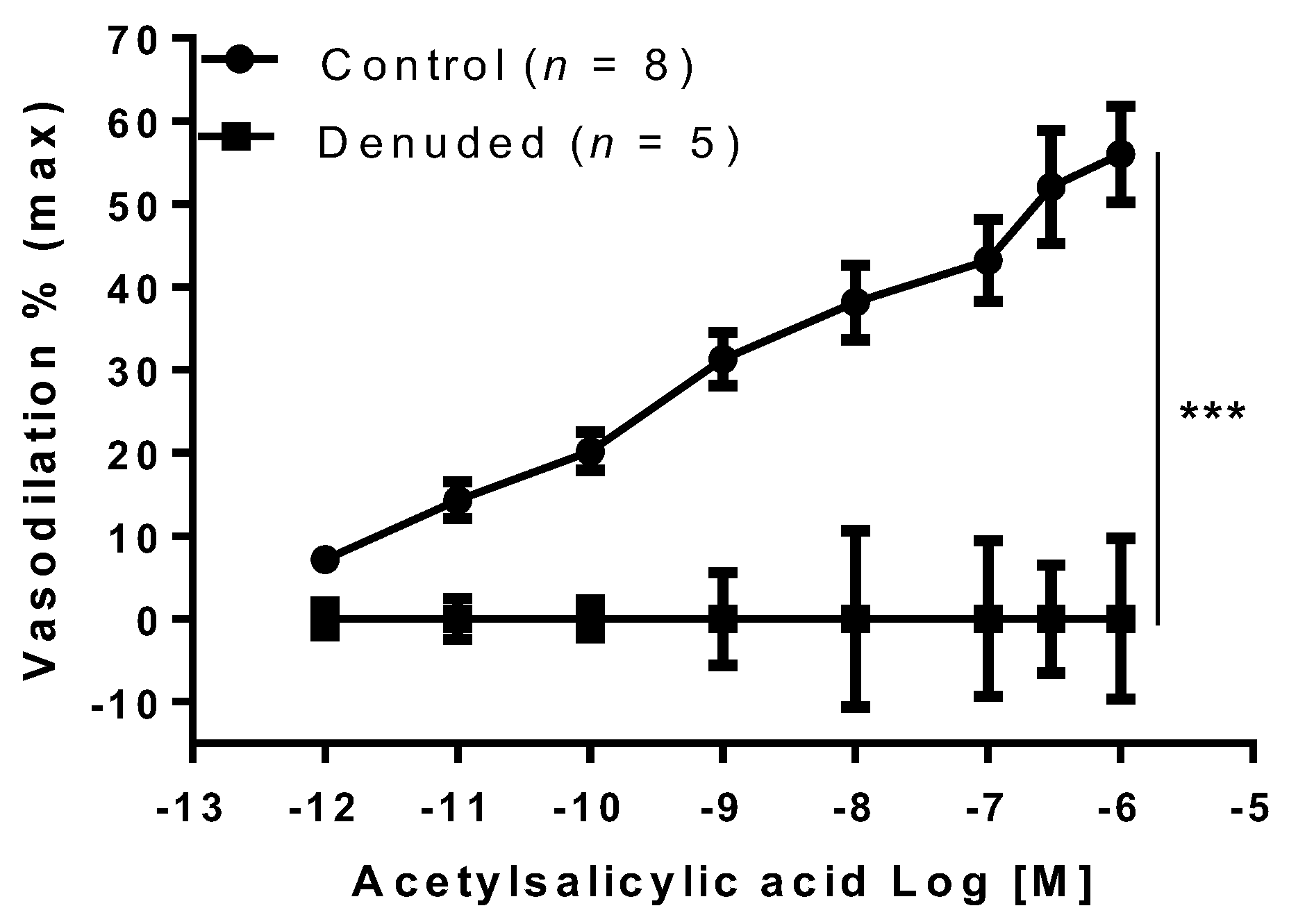
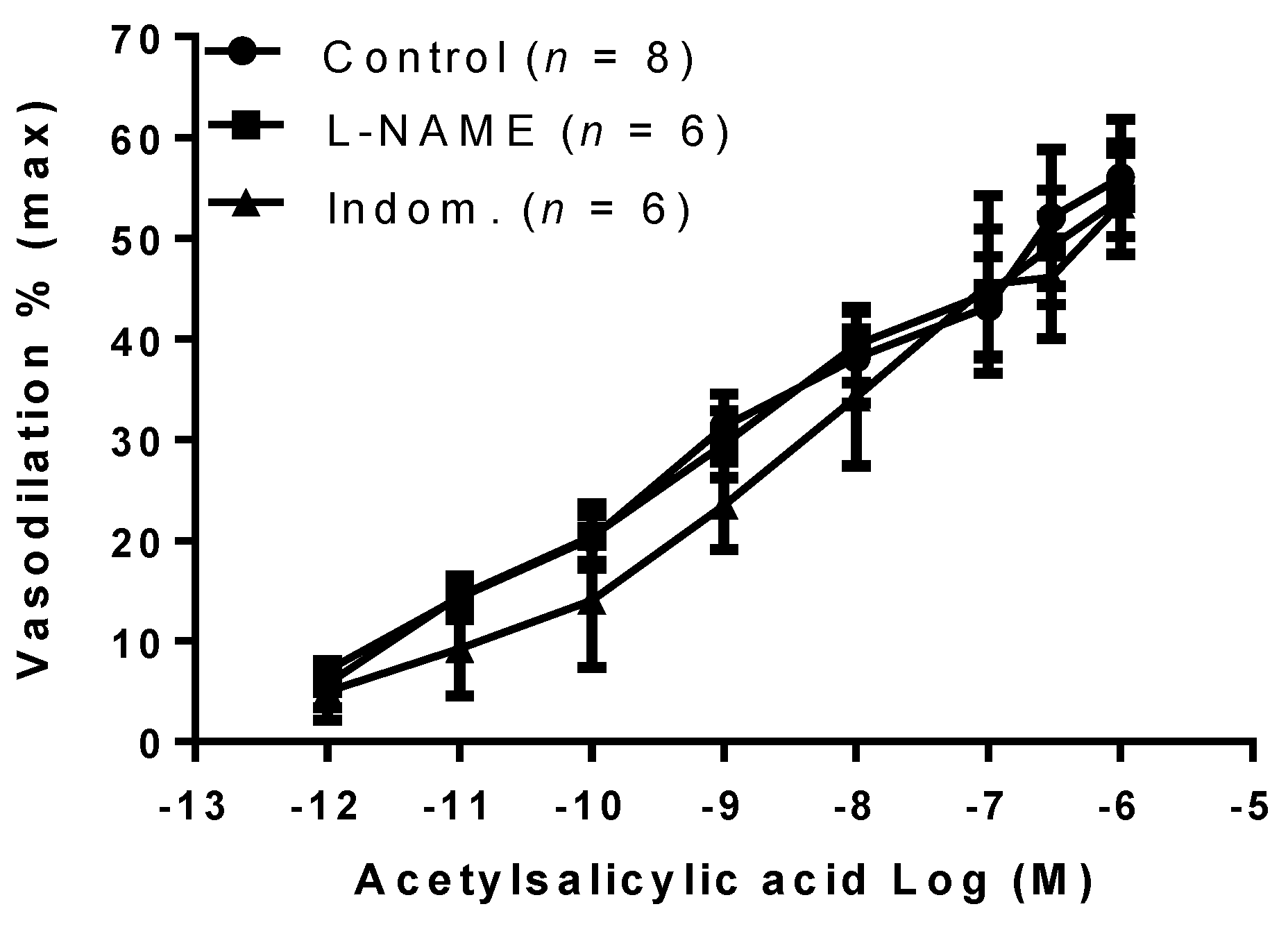
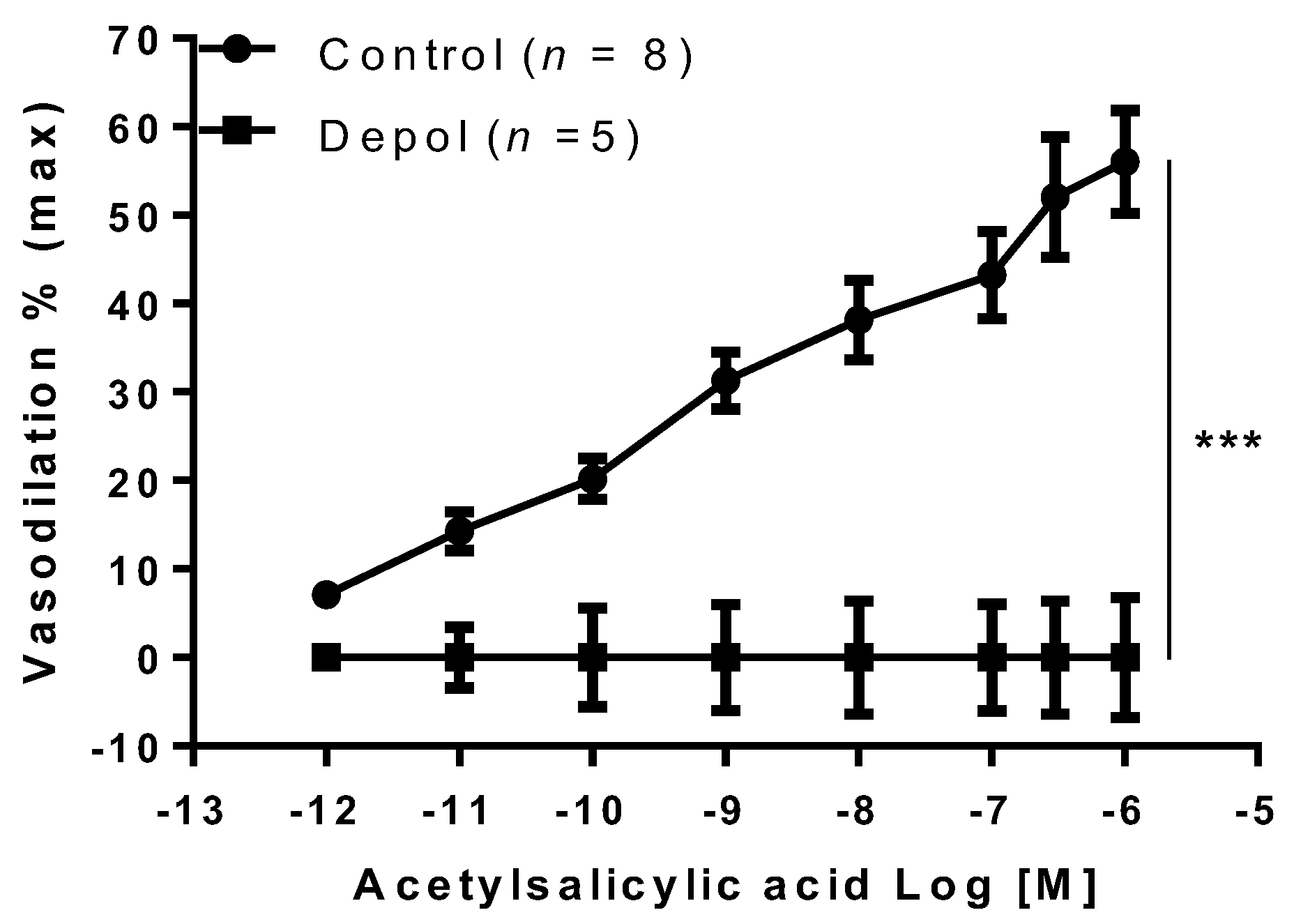
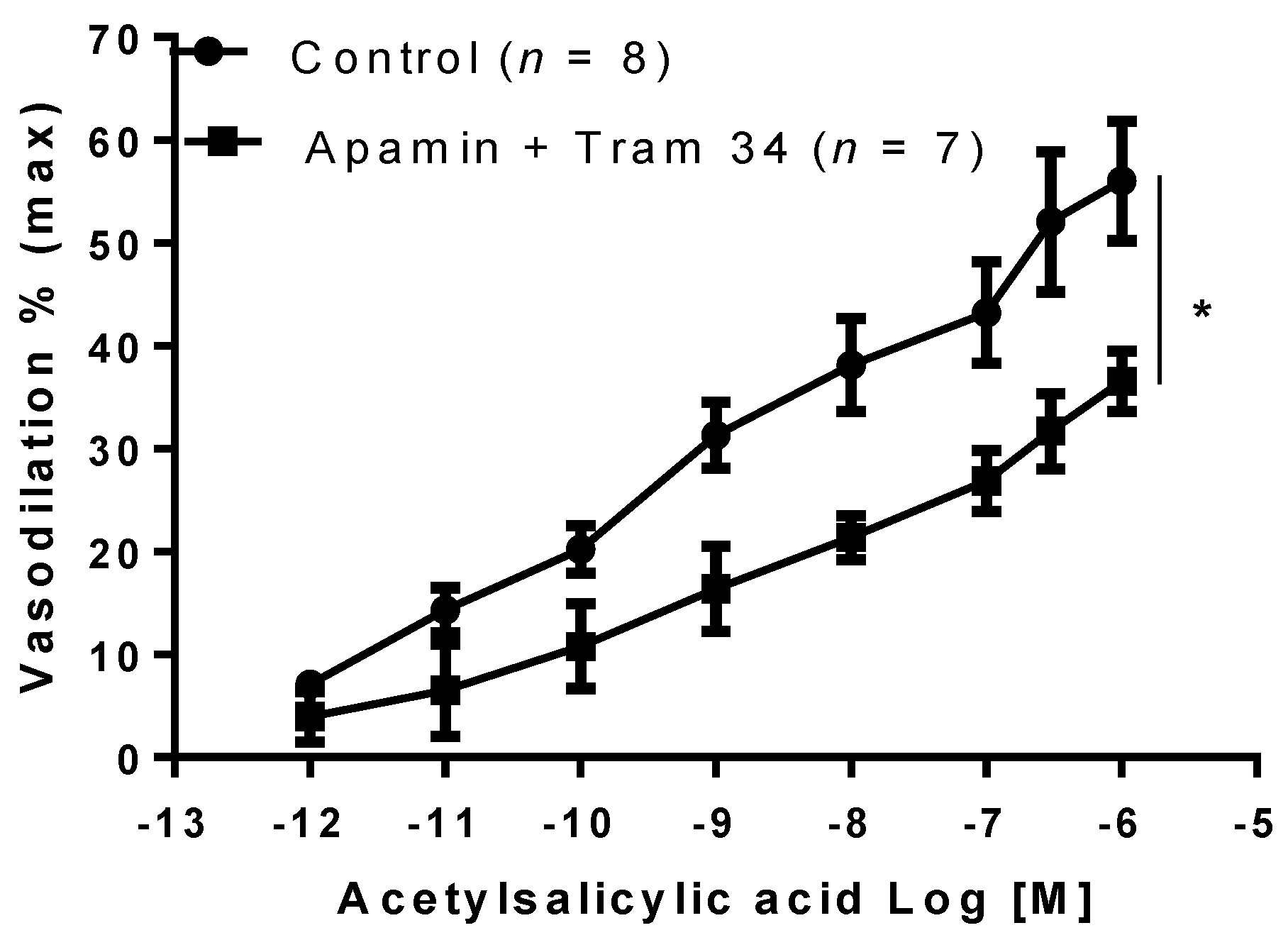
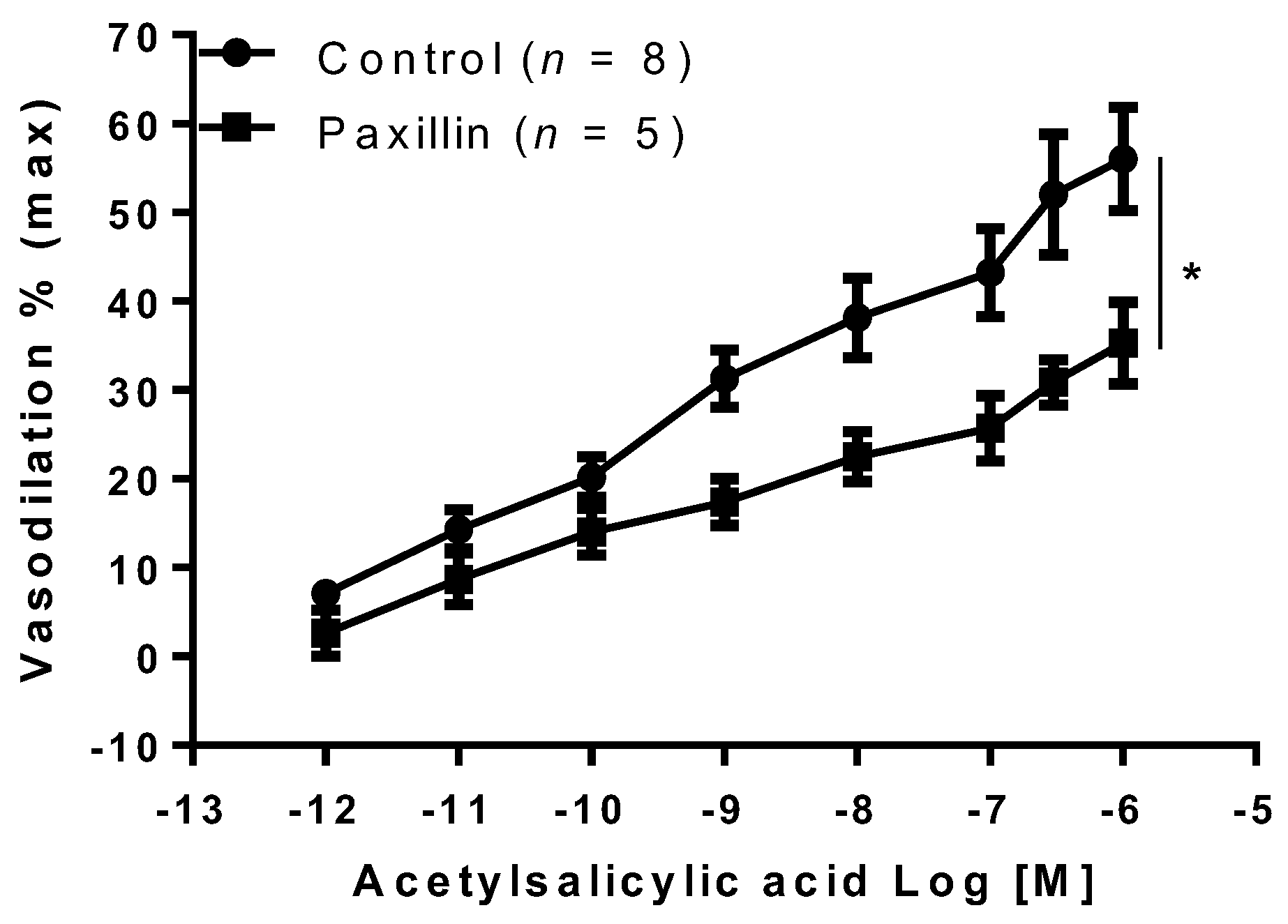
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Vessel Preparation
4.3. Experimental Protocol
4.4. Materials
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schrör, K. General aspects (History and Chemistry). In Acetylsalicylic Acid; Wiley-VCH: Weinheim, Germany, 2009; pp. 3–32. [Google Scholar]
- Zhou, B.; Wang, Q.; Zhao, Z.; Feng, X. A Systematic Review of the Efficacy and Safety of Aspirin When Delivered at Different Medication Times for the Primary and Secondary Prevention of Cardiovascular and Cerebrovascular Diseases. Innov. Regul. Sci. 2020, 54, 1339–1348. [Google Scholar] [CrossRef]
- Rolnik, D.L.; Wright, D.; Poon, L.C.; O’Gorman, N.; Syngelaki, A.; de Paco Matallana, C.; Akolekar, R.; Cicero, S.; Janga, D.; Singh, M.; et al. Aspirin versus Placebo in Pregnancies at High Risk for Preterm Preeclampsia. N. Engl. J. Med. 2017, 377, 613–622. [Google Scholar] [CrossRef]
- Walsh, S.W.; Strauss, J.F., 3rd. The Road to Low-Dose Aspirin Therapy for the Prevention of Preeclampsia Began with the Placenta. Int. J. Mol. Sci. 2021, 22, 6985. [Google Scholar] [CrossRef] [PubMed]
- Backes, C.H.; Markham, K.; Moorehead, P.; Cordero, L.; Nankervis, C.A.; Giannone, P.J. Maternal preeclampsia and neonatal outcomes. J. Pregnancy 2011, 2011, 214365. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.M.; Wallace, K.; Owens, M.; LaMarca, B. Pathophysiology and Current Clinical Management of Preeclampsia. Curr. Hypertens Rep. 2017, 19, 61. [Google Scholar] [CrossRef] [Green Version]
- Bujold, E.; Roberge, S.; Lacasse, Y.; Bureau, M.; Audibert, F.; Marcoux, S.; Forest, J.C.; Giguere, Y. Prevention of Preeclampsia and Intrauterine Growth Restriction with Aspirin Started in Early Pregnancy: A Meta-Analysis. Obstet. Gynecol. 2010, 116, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Roberge, S.; Bujold, E.; Nicolaides, K.H. Aspirin for the prevention of preterm and term preeclampsia: Systematic review and metaanalysis. Am. J. Obs. Gynecol. 2018, 218, 287–293.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haapsamo, M.; Martikainen, H.; Rasanen, J. Low-dose aspirin reduces uteroplacental vascular impedance in early and mid gestation in IVF and ICSI patients: A randomized, placebo-controlled double-blind study. Ultrasound Obstet. Gynecol. 2008, 32, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Kovo, M.; Rubinchik-Stern, M.; Miremberg, H.; Luria, O.; Bliecher, I.; Mizrachi, Y.; Bar, J. The effect of aspirin on placental vessels reactivity using the ex-vivo placental perfusion model. Thromb. Res. 2018, 170, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Ying, Z.K.; Giachini, F.R.C.; Tostes, R.C.; Webb, R.C. Salicylates dilate blood vessels through inhibiting PYK2-mediated RhoA/Rho-kinase activation. Cardiovasc. Res. 2009, 83, 155–162. [Google Scholar] [CrossRef]
- von der Weid, P.Y.; Hollenberg, M.D.; Fiorucci, S.; Wallace, J.L. Aspirin-triggered, cyclooxygenase-2-dependent lipoxin synthesis modulates vascular tone. Circulation 2004, 110, 1320–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helgadottir, H.; Tropea, T.; Gizurarson, S.; Mandala, M. Aspirin causes endothelium-dependent vasodilation of resistance arteries from non-gravid and gravid rats. Pregnancy Hypertens 2019, 15, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Ayala, D.E.; Calvo, C.; Lopez, J.E. Aspirin administered at bedtime, but not on awakening, has an effect on ambulatory blood pressure in hypertensive patients. J. Am. Coll. Cardiol. 2005, 46, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Leithäuser, B.; Mrowietz, C.; Park, J.W.; Jung, F. Influence of acetylsalicylic acid (Aspirin) on cutaneous microcirculation. Clin. Hemorheol. Microcirc. 2012, 50, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Osol, G.; Ko, N.L.; Mandalà, M. Plasticity of the Maternal Vasculature During Pregnancy. Annu. Rev. Physiol. 2019, 81, 89–111. [Google Scholar] [CrossRef]
- Lo Presti, D.; Scala, R.L.; Tiralongo, G.M.; Pisani, I.; Gagliardi, G.; Novelli, G.P.; Vasapollo, B.; Andreoli, A.; Valensise, H. Assessment of total vascular resistance and total body water in normotensive women during the first trimester of pregnancy in order to predict hypertensive complications. Pregnancy Hypertens. Int. J. Women’s Cardiovasc. Health 2013, 3, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Khalil, R.A.; Granger, J.P. Vascular mechanisms of increased arterial pressure in preeclampsia: Lessons from animal models. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R29–R45. [Google Scholar] [CrossRef] [Green Version]
- Sandoo, A.; van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Blocki, A.; Franke, R.P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M. Endothelial dysfunction in preeclampsia. Semin. Reprod. Endocrinol. 1998, 16, 5–15. [Google Scholar] [CrossRef]
- Ohlmann, P.; Jung, F.; Mrowietz, C.; Alt, T.; Alt, S.; Schmidt, W. Peripheral microcirculation during pregnancy and in women with pregnancy induced hypertension. Clin. Hemorheol. Microcirc. 2001, 24, 183–191. [Google Scholar]
- Pelland, A.; George, R.B.; Lehmann, C.; Coolen, J. Sidestream Dark Field Imaging of the Microcirculation to Assess Preeclampsia Microvascular Dysfunction. J. Clin. Med. Res. 2018, 10, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Klabunde, R.E. Cardiovascular Physiology Concepts, 3rd ed.; Wolters Kluwer: The Alfven am Rhein, The Netherlands, 2021; ISBN 9781975150075. [Google Scholar]
- Takala, J. Determinants of splanchnic blood flow. Br. J. Anaesth. 1996, 77, 50–58. [Google Scholar] [CrossRef]
- Kim, T.H.; Weiner, C.P.; Thompson, L.P. Effect of pregnancy on contraction and endothelium-mediated relaxation of renal and mesenteric arteries. Am. J. Physiol. 1994, 267 Pt 2, H41–H47. [Google Scholar] [CrossRef]
- Nelson, S.H.; Steinsland, O.S.; Suresh, M.S.; Lee, N.M. Pregnancy augments nitric oxide-dependent dilator response to acetylcholine in the human uterine artery. Hum. Reprod. 1998, 13, 1361–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luksha, L.; Nisell, H.; Luksha, N.; Kublickas, M.; Hultenby, K.; Kublickiene, K. Endothelium-derived hyperpolarizing factor in preeclampsia: Heterogeneous contribution, mechanisms, and morphological prerequisites. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R510–R519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barthelmes, J.; Nägele, M.P.; Ludovici, V.; Ruschitzka, F.; Sudano, I.; Flammer, A.J. Endothelial dysfunction in cardiovascular disease and Flammer syndrome-similarities and differences. EPMA J. 2017, 8, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Tuzcu, Z.B.; Asicioglu, E.; Sunbul, M.; Ozben, B.; Arikan, H.; Koc, M. Circulating endothelial cell number and markers of endothelial dysfunction in previously preeclamptic women. Am. J. Obstet. Gynecol. 2015, 213, 533.e1–533.e17. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Aranguren, L.C.; Prada, C.E.; Riaño-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: Role of oxidative stress. Front. Physiol. 2014, 5, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynard, S.; Epstein, F.H.; Karumanchi, S.A. Preeclampsia and angiogenic imbalance. Annu. Rev. Med. 2008, 59, 61–78. [Google Scholar] [CrossRef]
- Crews, J.K.; Herrington, J.N.; Granger, J.P.; Khalil, R.A. Decreased endothelium-dependent vascular relaxation during reduction of uterine perfusion pressure in pregnant rat. Hypertension 2000, 35 Pt 2, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Hecker, M.; Bara, A.T.; Bauersachs, J.; Busse, R. Characterization of EDHF as a cytochrome P450-derived arachidonic acid metabolite in mammals. J. Physiol. 1994, 481, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Sandow, S.L.; Tare, M.; Coleman, H.A.; Hill, C.E.; Parkington, H.C. Involvement of myoendothelial gap junctions in the actions of endothelium-derived hyperpolarizing factor. Circ. Res. 2002, 90, 1108–1113. [Google Scholar] [CrossRef] [Green Version]
- Edward, G.; Dora, K.A.; Gardener, M.J.; Garland, C.J.; Weston, A.H. K+ is an endothelium-derived hyperpolarizing factor in rat arteries. Nature 1998, 396, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Grgic, I.; Kaistha, B.P.; Hoyer, J.; Kohler, R. Endothelial Ca2+ -activated K+ channels in normal and impaired EDHF-dilator responses-relevance to cardiovascular pathologies and drug discovery. Br. J. Pharmacol. 2009, 157, 509–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilgers, R.H.; Todd, J.J.R.; Webb, R.C. Regional heterogeneity in acetylcholine-induced relaxation in rat vascular bed: Role of calcium-activated K+ channels. Am. J. Physiol. Heart Cir. Physiol. 2006, 291, H216–H222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandalà, M.; Gokina, N.; Barron, C.; Osol, G. Endothelial-derived hyperpolarization factor (EDHF) contributes to PLGF-induced dilation of mesenteric resistance arteries from pregnant rats. J. Vasc. Res. 2012, 49, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colton, I.; Mandalà, M.; Morton, J.; Davidge, S.T.; Osol, G. Influence of constriction, wall tension, smooth muscle activation and cellular deformation on rat resistance artery vasodilator reactivity. Cell. Physiol. Biochem. 2012, 29, 883–892. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helgadóttir, H.; Tropea, T.; Gizurarson, S.; Mandalà, M. Endothelium-Derived Hyperpolarizing Factor (EDHF) Mediates Acetylsalicylic Acid (Aspirin) Vasodilation of Pregnant Rat Mesenteric Arteries. Int. J. Mol. Sci. 2021, 22, 10162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810162
Helgadóttir H, Tropea T, Gizurarson S, Mandalà M. Endothelium-Derived Hyperpolarizing Factor (EDHF) Mediates Acetylsalicylic Acid (Aspirin) Vasodilation of Pregnant Rat Mesenteric Arteries. International Journal of Molecular Sciences. 2021; 22(18):10162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810162
Chicago/Turabian StyleHelgadóttir, Helga, Teresa Tropea, Sveinbjörn Gizurarson, and Maurizio Mandalà. 2021. "Endothelium-Derived Hyperpolarizing Factor (EDHF) Mediates Acetylsalicylic Acid (Aspirin) Vasodilation of Pregnant Rat Mesenteric Arteries" International Journal of Molecular Sciences 22, no. 18: 10162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810162