
Blockade of Serotonin 5-HT6 Receptor Constitutive Activity Alleviates Cognitive Deficits in a Preclinical Model of Neurofibromatosis Type 1

 , and
, and
Abstract
:1. Introduction
2. Results
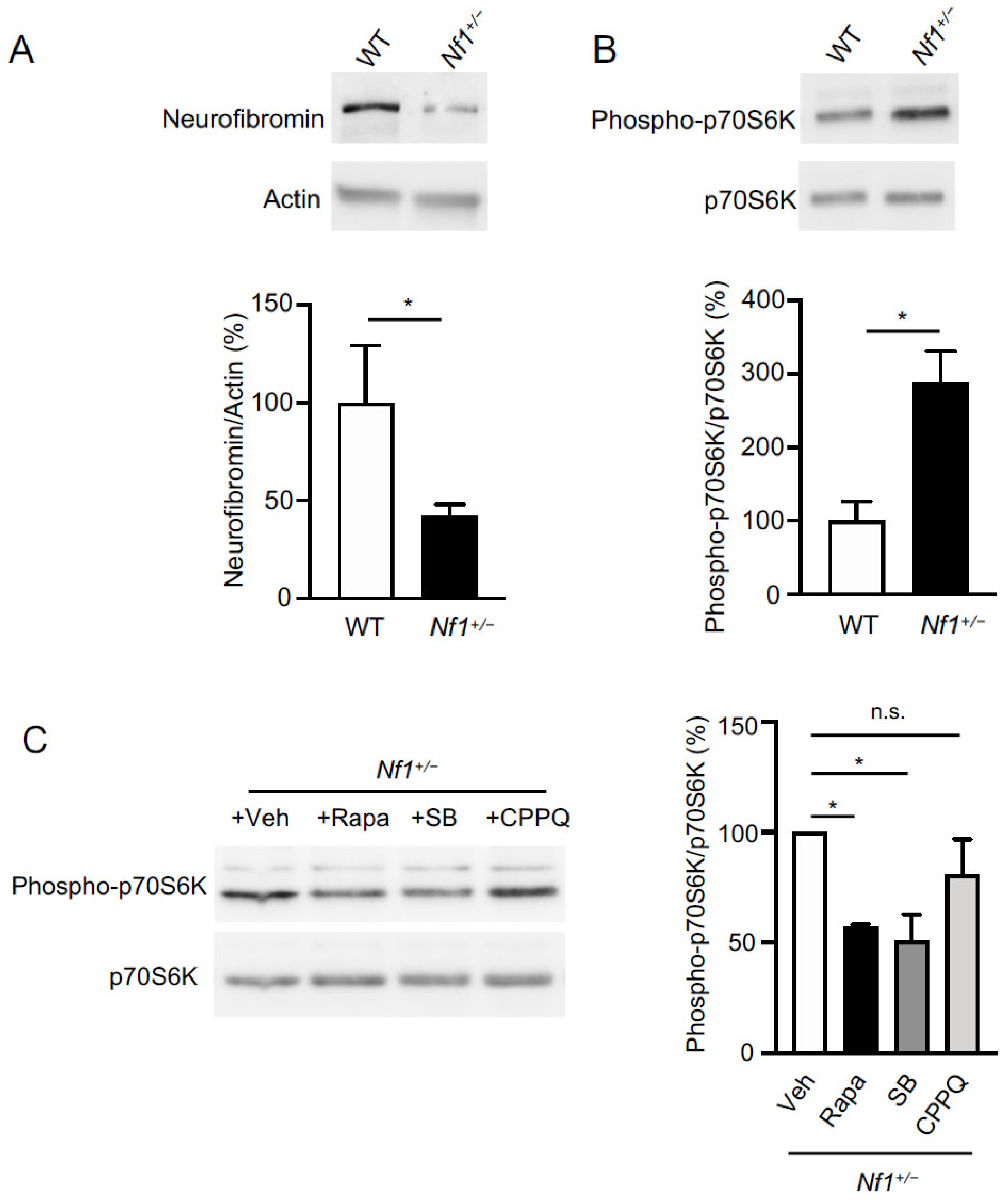
2.1. Role of 5-HT6 Receptor in mTOR Overactivation in Prefrontal Cortex of Nf1+/− Mice
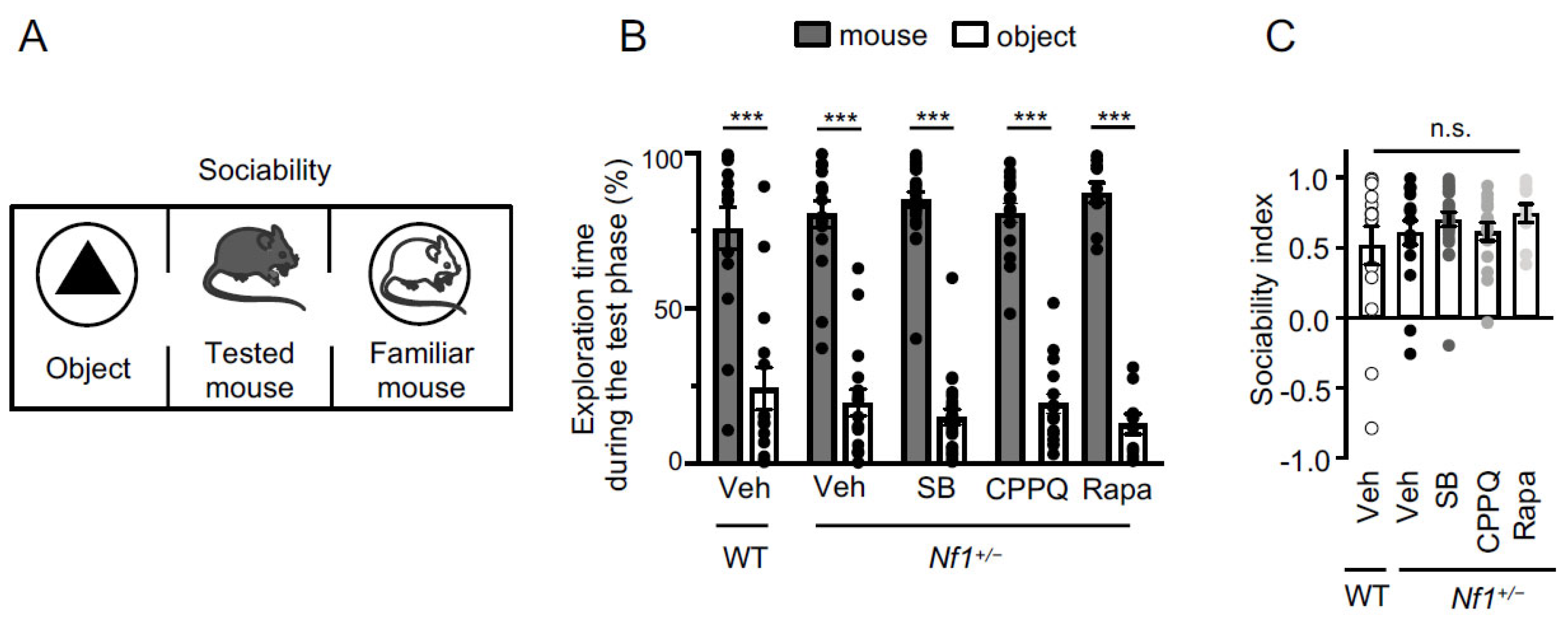
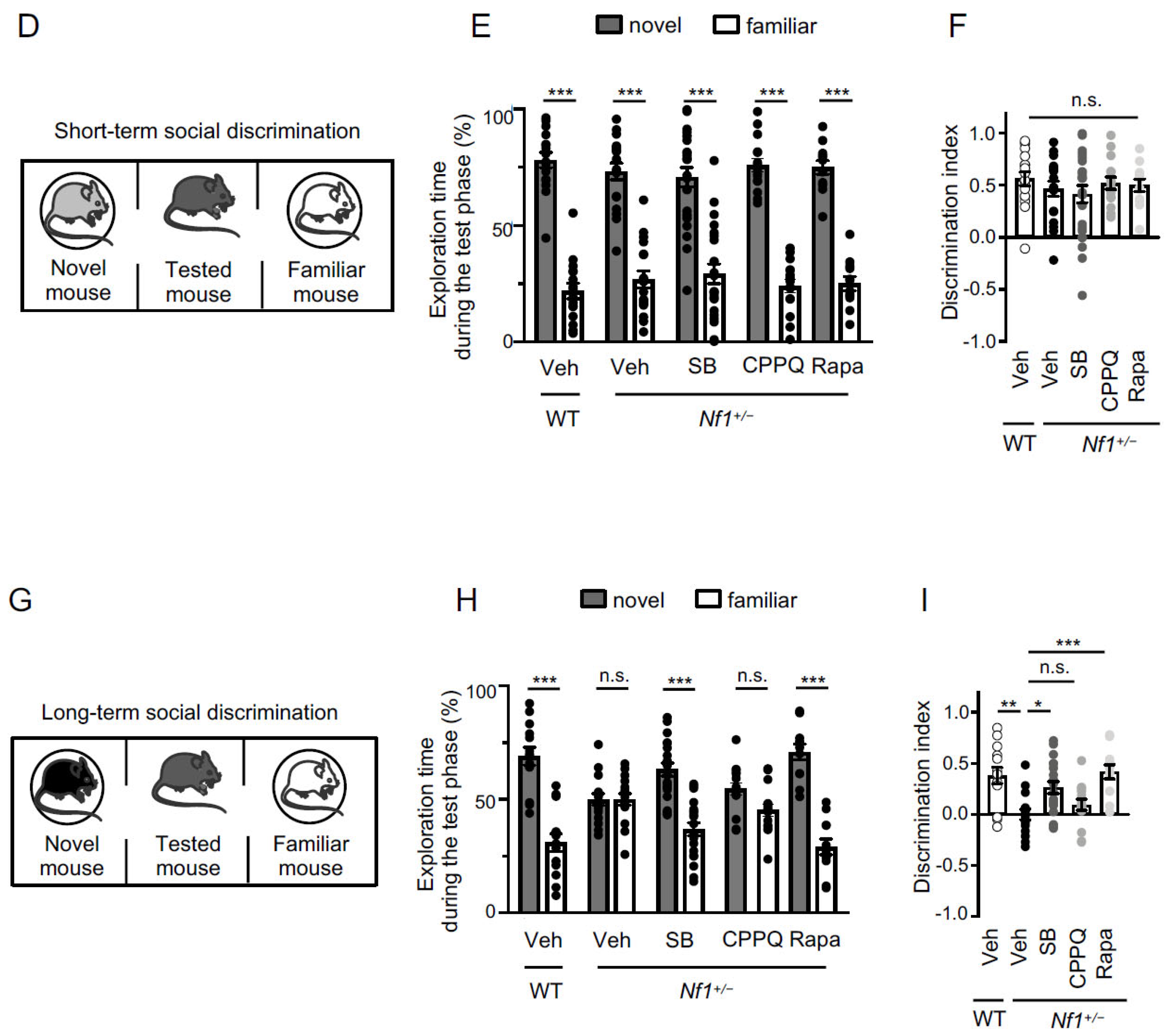
2.2. Effect of Blocking the 5-HT6 Receptor-mTOR Pathway on Sociability and Short-Term and Long-Term Social Memories of Nf1+/− Mice
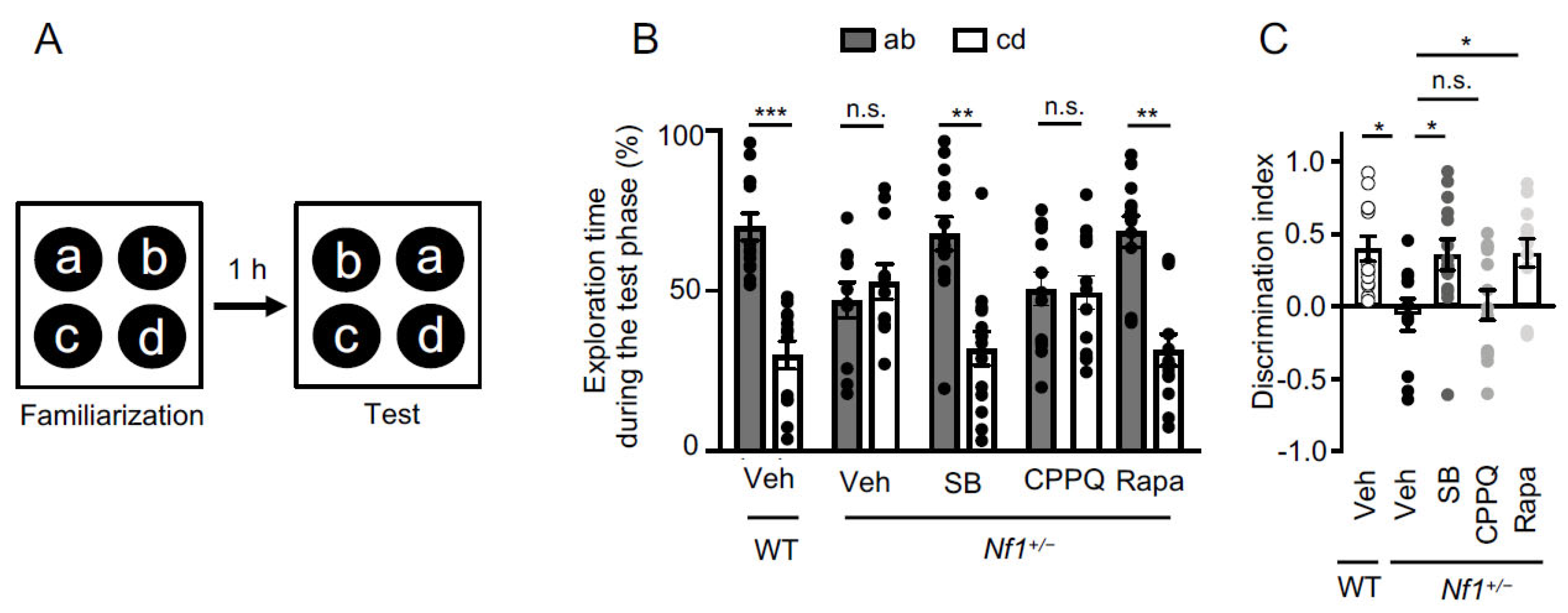
2.3. Role of 5-HT6 Receptor-Operated mTOR Signaling in Associative Memory Deficit of Nf1+/− Mice
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Drug Administration
4.3. Behavioral Tests
4.3.1. Social Behavior Tests
4.3.2. Object-in-Place Task
4.4. Western Blotting
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergqvist, C.; Hemery, F.; Ferkal, S.; Wolkenstein, P. Neurofibromatosis I and multiple sclerosis. Orphanet. J. Rare. Dis. 2020, 15, 186. [Google Scholar] [CrossRef]
- Graf, A.; Landolt, M.A.; Mori, A.C.; Boltshauser, E. Quality of life and psychological adjustment in children and adolescents with neurofibromatosis type 1. J. Pediatr. 2006, 149, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Sanagoo, A.; Jouybari, L.; Koohi, F.; Sayehmiri, F. Evaluation of QoL in neurofibromatosis patients: A systematic review and meta-analysis study. BMC Neurol. 2019, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Gipson, T.T.; Johnston, M.V. Plasticity and mTOR: Towards restoration of impaired synaptic plasticity in mTOR-related neurogenetic disorders. Neural. Plast. 2012, 2012, 486402. [Google Scholar] [CrossRef]
- Hyman, S.L.; Shores, A.; North, K.N. The nature and frequency of cognitive deficits in children with neurofibromatosis type 1. Neurology 2005, 65, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Zoller, M.E.; Rembeck, B.; Backman, L. Neuropsychological deficits in adults with neurofibromatosis type 1. Acta Neurol. Scand. 1997, 95, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.M.; Hyman, S.L.; Shores, E.A.; North, K.N. Assessment of executive function and attention in children with neurofibromatosis type 1: Relationships between cognitive measures and real-world behavior. Child. Neuropsychol. 2011, 17, 313–329. [Google Scholar] [CrossRef]
- Pride, N.A.; Crawford, H.; Payne, J.M.; North, K.N. Social functioning in adults with neurofibromatosis type 1. Res. Dev. Disabil. 2013, 34, 3393–3399. [Google Scholar] [CrossRef]
- Plasschaert, E.; Van Eylen, L.; Descheemaeker, M.J.; Noens, I.; Legius, E.; Steyaert, J. Executive functioning deficits in children with neurofibromatosis type 1: The influence of intellectual and social functioning. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171B, 348–362. [Google Scholar] [CrossRef]
- Chisholm, A.K.; Anderson, V.A.; Pride, N.A.; Malarbi, S.; North, K.N.; Payne, J.M. Social Function and Autism Spectrum Disorder in Children and Adults with Neurofibromatosis Type 1: A Systematic Review and Meta-Analysis. Neuropsychol. Rev. 2018, 28, 317–340. [Google Scholar] [CrossRef]
- Payne, J.M.; Barton, B.; Ullrich, N.J.; Cantor, A.; Hearps, S.J.; Cutter, G.; Rosser, T.; Walsh, K.S.; Gioia, G.A.; Wolters, P.L.; et al. Randomized placebo-controlled study of lovastatin in children with neurofibromatosis type 1. Neurology 2016, 87, 2575–2584. [Google Scholar] [CrossRef]
- Pride, N.A.; Barton, B.; Hutchins, P.; Coghill, D.R.; Korgaonkar, M.S.; Hearps, S.J.C.; Rouel, M.; Malarbi, S.; North, K.N.; Payne, J.M. Effects of methylphenidate on cognition and behaviour in children with neurofibromatosis type 1: A study protocol for a randomised placebo-controlled crossover trial. BMJ Open 2018, 8, e021800. [Google Scholar] [CrossRef] [PubMed]
- Stivaros, S.; Garg, S.; Tziraki, M.; Cai, Y.; Thomas, O.; Mellor, J.; Morris, A.A.; Jim, C.; Szumanska-Ryt, K.; Parkes, L.M.; et al. Randomised controlled trial of simvastatin treatment for autism in young children with neurofibromatosis type 1 (SANTA). Mol. Autism 2018, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, N.J.; Payne, J.M.; Walsh, K.S.; Cutter, G.; Packer, R.; North, K.; Rey-Casserly, C.; Consortium, N.F.C.T. Visual spatial learning outcomes for clinical trials in neurofibromatosis type 1. Ann. Clin. Transl. Neurol. 2020, 7, 245–249. [Google Scholar] [CrossRef]
- Payne, J.M.; Hearps, S.J.C.; Walsh, K.S.; Paltin, I.; Barton, B.; Ullrich, N.J.; Haebich, K.M.; Coghill, D.; Gioia, G.A.; Cantor, A.; et al. Reproducibility of cognitive endpoints in clinical trials: Lessons from neurofibromatosis type 1. Ann. Clin. Transl. Neurol. 2019, 6, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Ballester, R.; Marchuk, D.; Boguski, M.; Saulino, A.; Letcher, R.; Wigler, M.; Collins, F. The NF1 locus encodes a protein functionally related to mammalian GAP and yeast IRA proteins. Cell 1990, 63, 851–859. [Google Scholar] [CrossRef]
- Xu, G.F.; O’Connell, P.; Viskochil, D.; Cawthon, R.; Robertson, M.; Culver, M.; Dunn, D.; Stevens, J.; Gesteland, R.; White, R.; et al. The neurofibromatosis type 1 gene encodes a protein related to GAP. Cell 1990, 62, 599–608. [Google Scholar] [CrossRef]
- Hennig, A.; Markwart, R.; Wolff, K.; Schubert, K.; Cui, Y.; Prior, I.A.; Esparza-Franco, M.A.; Ladds, G.; Rubio, I. Feedback activation of neurofibromin terminates growth factor-induced Ras activation. Cell Commun. Signal. 2016, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.F.; Yasuda, R. Neurofibromin is the major ras inactivator in dendritic spines. J. Neurosci. 2014, 34, 776–783. [Google Scholar] [CrossRef]
- Meffre, J.; Chaumont-Dubel, S.; Mannoury la Cour, C.; Loiseau, F.; Watson, D.J.; Dekeyne, A.; Seveno, M.; Rivet, J.M.; Gaven, F.; Deleris, P.; et al. 5-HT(6) receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol. Med. 2012, 4, 1043–1056. [Google Scholar] [CrossRef]
- Deraredj Nadim, W.; Chaumont-Dubel, S.; Madouri, F.; Cobret, L.; De Tauzia, M.L.; Zajdel, P.; Benedetti, H.; Marin, P.; Morisset-Lopez, S. Physical interaction between neurofibromin and serotonin 5-HT6 receptor promotes receptor constitutive activity. Proc. Natl. Acad. Sci. USA 2016, 113, 12310–12315. [Google Scholar] [CrossRef]
- Berthoux, C.; Hamieh, A.M.; Rogliardo, A.; Doucet, E.L.; Coudert, C.; Ango, F.; Grychowska, K.; Chaumont-Dubel, S.; Zajdel, P.; Maldonado, R.; et al. Early 5-HT6 receptor blockade prevents symptom onset in a model of adolescent cannabis abuse. EMBO Mol. Med. 2020, 12, e10605. [Google Scholar] [CrossRef]
- Martin, P.Y.; Doly, S.; Hamieh, A.M.; Chapuy, E.; Canale, V.; Drop, M.; Chaumont-Dubel, S.; Bantreil, X.; Lamaty, F.; Bojarski, A.J.; et al. mTOR activation by constitutively active serotonin6 receptors as new paradigm in neuropathic pain and its treatment. Prog. Neurobiol. 2020, 193, 101846. [Google Scholar] [CrossRef]
- Teng, L.L.; Lu, G.L.; Chiou, L.C.; Lin, W.S.; Cheng, Y.Y.; Hsueh, T.E.; Huang, Y.C.; Hwang, N.H.; Yeh, J.W.; Liao, R.M.; et al. Serotonin receptor HTR6-mediated mTORC1 signaling regulates dietary restriction-induced memory enhancement. PLoS Biol. 2019, 17, e2007097. [Google Scholar] [CrossRef]
- Yun, H.M.; Rhim, H. The serotonin-6 receptor as a novel therapeutic target. Exp. Neurobiol. 2011, 20, 159–168. [Google Scholar] [CrossRef]
- Codony, X.; Vela, J.M.; Ramirez, M.J. 5-HT(6) receptor and cognition. Curr. Opin. Pharmacol. 2011, 11, 94–100. [Google Scholar] [CrossRef]
- Khoury, A.; Runnstrom, M.; Ebied, A.; Penny, E.S. Linezolid-associated serotonin toxicity after escitalopram discontinuation: Concomitant drug considerations. BMJ Case Rep. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Brannan, C.I.; Perkins, A.S.; Vogel, K.S.; Ratner, N.; Nordlund, M.L.; Reid, S.W.; Buchberg, A.M.; Jenkins, N.A.; Parada, L.F.; Copeland, N.G. Targeted disruption of the neurofibromatosis type-1 gene leads to developmental abnormalities in heart and various neural crest-derived tissues. Genes Dev. 1994, 8, 1019–1029. [Google Scholar] [CrossRef]
- Silva, A.J.; Frankland, P.W.; Marowitz, Z.; Friedman, E.; Laszlo, G.S.; Cioffi, D.; Jacks, T.; Bourtchuladze, R. A mouse model for the learning and memory deficits associated with neurofibromatosis type I. Nat. Genet. 1997, 15, 281–284. [Google Scholar] [CrossRef]
- Molosh, A.I.; Johnson, P.L.; Spence, J.P.; Arendt, D.; Federici, L.M.; Bernabe, C.; Janasik, S.P.; Segu, Z.M.; Khanna, R.; Goswami, C.; et al. Social learning and amygdala disruptions in Nf1 mice are rescued by blocking p21-activated kinase. Nat. Neurosci. 2014, 17, 1583–1590. [Google Scholar] [CrossRef]
- Petrella, L.I.; Cai, Y.; Sereno, J.V.; Goncalves, S.I.; Silva, A.J.; Castelo-Branco, M. Brain and behaviour phenotyping of a mouse model of neurofibromatosis type-1: An MRI/DTI study on social cognition. Genes Brain Behav. 2016, 15, 637–646. [Google Scholar] [CrossRef]
- Lukkes, J.L.; Drozd, H.P.; Fitz, S.D.; Molosh, A.I.; Clapp, D.W.; Shekhar, A. Guanfacine treatment improves ADHD phenotypes of impulsivity and hyperactivity in a neurofibromatosis type 1 mouse model. J. Neurodev. Disord. 2020, 12, 2. [Google Scholar] [CrossRef]
- Grychowska, K.; Satala, G.; Kos, T.; Partyka, A.; Colacino, E.; Chaumont-Dubel, S.; Bantreil, X.; Wesolowska, A.; Pawlowski, M.; Martinez, J.; et al. Novel 1H-Pyrrolo[3,2-c]quinoline Based 5-HT6 Receptor Antagonists with Potential Application for the Treatment of Cognitive Disorders Associated with Alzheimer’s Disease. ACS Chem. Neurosci. 2016, 7, 972–983. [Google Scholar] [CrossRef]
- Grychowska, K.; Chaumont-Dubel, S.; Kurczab, R.; Koczurkiewicz, P.; Deville, C.; Krawczyk, M.; Pietrus, W.; Satala, G.; Buda, S.; Piska, K.; et al. Dual 5-HT6 and D3 Receptor Antagonists in a Group of 1H-Pyrrolo[3,2-c]quinolines with Neuroprotective and Procognitive Activity. ACS Chem. Neurosci. 2019, 10, 3183–3196. [Google Scholar] [CrossRef]
- Shilyansky, C.; Karlsgodt, K.H.; Cummings, D.M.; Sidiropoulou, K.; Hardt, M.; James, A.S.; Ehninger, D.; Bearden, C.E.; Poirazi, P.; Jentsch, J.D.; et al. Neurofibromin regulates corticostriatal inhibitory networks during working memory performance. Proc. Natl. Acad. Sci. USA 2010, 107, 13141–13146. [Google Scholar] [CrossRef]
- Cross, L.; Brown, M.W.; Aggleton, J.P.; Warburton, E.C. The medial dorsal thalamic nucleus and the medial prefrontal cortex of the rat function together to support associative recognition and recency but not item recognition. Learn. Mem. 2012, 20, 41–50. [Google Scholar] [CrossRef]
- Chen, P.C.; Chang, Y.L. Associative memory and underlying brain correlates in older adults with mild cognitive impairment. Neuropsychologia 2016, 85, 216–225. [Google Scholar] [CrossRef]
- Johannessen, C.M.; Johnson, B.W.; Williams, S.M.; Chan, A.W.; Reczek, E.E.; Lynch, R.C.; Rioth, M.J.; McClatchey, A.; Ryeom, S.; Cichowski, K. TORC1 is essential for NF1-associated malignancies. Curr. Biol. 2008, 18, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, C.M.; Reczek, E.E.; James, M.F.; Brems, H.; Legius, E.; Cichowski, K. The NF1 tumor suppressor critically regulates TSC2 and mTOR. Proc. Natl. Acad. Sci. USA 2005, 102, 8573–8578. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, B.; Yi, Y.; Chen, D.Y.; Weber, J.D.; Gutmann, D.H. Proteomic analysis reveals hyperactivation of the mammalian target of rapamycin pathway in neurofibromatosis 1-associated human and mouse brain tumors. Cancer Res. 2005, 65, 2755–2760. [Google Scholar] [CrossRef] [PubMed]
- Bockaert, J.; Marin, P. mTOR in Brain Physiology and Pathologies. Physiol. Rev. 2015, 95, 1157–1187. [Google Scholar] [CrossRef]
- Maloney, S.E.; Chandler, K.C.; Anastasaki, C.; Rieger, M.A.; Gutmann, D.H.; Dougherty, J.D. Characterization of early communicative behavior in mouse models of neurofibromatosis type 1. Autism Res. 2018, 11, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Pujol, C.N.; Dupuy, V.; Seveno, M.; Runtz, L.; Bockaert, J.; Marin, P.; Chaumont-Dubel, S. Dynamic interactions of the 5-HT6 receptor with protein partners control dendritic tree morphogenesis. Sci. Signal. 2020, 13. [Google Scholar] [CrossRef]
- Hegedus, B.; Banerjee, D.; Yeh, T.H.; Rothermich, S.; Perry, A.; Rubin, J.B.; Garbow, J.R.; Gutmann, D.H. Preclinical cancer therapy in a mouse model of neurofibromatosis-1 optic glioma. Cancer Res. 2008, 68, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Johansson, G.; Mahller, Y.Y.; Collins, M.H.; Kim, M.O.; Nobukuni, T.; Perentesis, J.; Cripe, T.P.; Lane, H.A.; Kozma, S.C.; Thomas, G.; et al. Effective in vivo targeting of the mammalian target of rapamycin pathway in malignant peripheral nerve sheath tumors. Mol. Cancer Ther. 2008, 7, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Bhola, P.; Banerjee, S.; Mukherjee, J.; Balasubramanium, A.; Arun, V.; Karim, Z.; Burrell, K.; Croul, S.; Gutmann, D.H.; Guha, A. Preclinical in vivo evaluation of rapamycin in human malignant peripheral nerve sheath explant xenograft. Int. J. Cancer 2010, 126, 563–571. [Google Scholar] [CrossRef]
- Weiss, B.; Widemann, B.C.; Wolters, P.; Dombi, E.; Vinks, A.; Cantor, A.; Perentesis, J.; Schorry, E.; Ullrich, N.; Gutmann, D.H.; et al. Sirolimus for progressive neurofibromatosis type 1-associated plexiform neurofibromas: A neurofibromatosis Clinical Trials Consortium phase II study. Neuro. Oncol. 2015, 17, 596–603. [Google Scholar] [CrossRef]
- Ullrich, N.J.; Prabhu, S.P.; Reddy, A.T.; Fisher, M.J.; Packer, R.; Goldman, S.; Robison, N.J.; Gutmann, D.H.; Viskochil, D.H.; Allen, J.C.; et al. A phase II study of continuous oral mTOR inhibitor everolimus for recurrent, radiographic-progressive neurofibromatosis type 1-associated pediatric low-grade glioma: A Neurofibromatosis Clinical Trials Consortium study. Neuro. Oncol. 2020, 22, 1527–1535. [Google Scholar] [CrossRef]
- Ehninger, D.; Han, S.; Shilyansky, C.; Zhou, Y.; Li, W.; Kwiatkowski, D.J.; Ramesh, V.; Silva, A.J. Reversal of learning deficits in a Tsc2+/− mouse model of tuberous sclerosis. Nat. Med. 2008, 14, 843–848. [Google Scholar] [CrossRef]
- Ricciardi, S.; Boggio, E.M.; Grosso, S.; Lonetti, G.; Forlani, G.; Stefanelli, G.; Calcagno, E.; Morello, N.; Landsberger, N.; Biffo, S.; et al. Reduced AKT/mTOR signaling and protein synthesis dysregulation in a Rett syndrome animal model. Hum. Mol. Genet. 2011, 20, 1182–1196. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Hoeffer, C.A.; Takayasu, Y.; Miyawaki, T.; McBride, S.M.; Klann, E.; Zukin, R.S. Dysregulation of mTOR signaling in fragile X syndrome. J. Neurosci. 2010, 30, 694–702. [Google Scholar] [CrossRef]
- Troca-Marin, J.A.; Alves-Sampaio, A.; Montesinos, M.L. Deregulated mTOR-mediated translation in intellectual disability. Prog. Neurobiol. 2012, 96, 268–282. [Google Scholar] [CrossRef]
- Kwon, M.; Han, J.; Kim, U.J.; Cha, M.; Um, S.W.; Bai, S.J.; Hong, S.K.; Lee, B.H. Inhibition of Mammalian Target of Rapamycin (mTOR) Signaling in the Insular Cortex Alleviates Neuropathic Pain after Peripheral Nerve Injury. Front. Mol. Neurosci. 2017, 10, 79. [Google Scholar] [CrossRef]
- Puighermanal, E.; Marsicano, G.; Busquets-Garcia, A.; Lutz, B.; Maldonado, R.; Ozaita, A. Cannabinoid modulation of hippocampal long-term memory is mediated by mTOR signaling. Nat. Neurosci. 2009, 12, 1152–1158. [Google Scholar] [CrossRef]
- Guo, J.R.; Wang, H.; Jin, X.J.; Jia, D.L.; Zhou, X.; Tao, Q. Effect and mechanism of inhibition of PI3K/Akt/mTOR signal pathway on chronic neuropathic pain and spinal microglia in a rat model of chronic constriction injury. Oncotarget 2017, 8, 52923–52934. [Google Scholar] [CrossRef]
- Cui, Y.; Costa, R.M.; Murphy, G.G.; Elgersma, Y.; Zhu, Y.; Gutmann, D.H.; Parada, L.F.; Mody, I.; Silva, A.J. Neurofibromin regulation of ERK signaling modulates GABA release and learning. Cell 2008, 135, 549–560. [Google Scholar] [CrossRef]
- Omrani, A.; van der Vaart, T.; Mientjes, E.; van Woerden, G.M.; Hojjati, M.R.; Li, K.W.; Gutmann, D.H.; Levelt, C.N.; Smit, A.B.; Silva, A.J.; et al. HCN channels are a novel therapeutic target for cognitive dysfunction in Neurofibromatosis type 1. Mol. Psychiatry 2015, 20, 1311–1321. [Google Scholar] [CrossRef]
- Shah, M.M. Cortical HCN channels: Function, trafficking and plasticity. J. Physiol. 2014, 592, 2711–2719. [Google Scholar] [CrossRef]
- He, C.; Chen, F.; Li, B.; Hu, Z. Neurophysiology of HCN channels: From cellular functions to multiple regulations. Prog. Neurobiol. 2014, 112, 1–23. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2011, 48, e2473. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Test | Experimental Conditions | Discrimination Index (mean ± sem) |
| Sociability | WT + Vehicle | 0.52 ± 0.14 |
| Nf1+/− + Vehicle | 0.61 ± 0.08 | |
| Nf1+/− + SB258585 | 0.70 ± 0.05 | |
| Nf1+/− + CPPQ | 0.61 ± 0.06 | |
| Nf1+/− + Rapamacyn | 0.75 ± 0.07 | |
| Short-term memory | WT + Vehicle | 0.56 ± 0.07 |
| Nf1+/− + Vehicle | 0.46 ± 0.07 | |
| Nf1+/− + SB258585 | 0.42 ± 0.08 | |
| Nf1+/− + CPPQ | 0.52 ± 0.06 | |
| Nf1+/− + Rapamacyn | 0.50 ± 0.06 | |
| Long-term memory | WT + Vehicle | 0.38 ± 0.08 |
| Nf1+/− + Vehicle | 0.00 ± 0.05 | |
| Nf1+/− + SB258585 | 0.26 ± 0.06 | |
| Nf1+/− + CPPQ | 0.09 ± 0.05 | |
| Nf1+/− + Rapamacyn | 0.42 ± 0.07 | |
| Object-in-place | WT + Vehicle | 0.40 ± 0.09 |
| Nf1+/− + Vehicle | -0.06 ± 0.11 | |
| Nf1+/− + SB258585 | 0.36 ± 0.11 | |
| Nf1+/− + CPPQ | 0.01 ± 0.10 | |
| Nf1+/− + Rapamacyn | 0.37 ± 0.10 | |
| Behavioral test | Experimental Conditions | Exploration Time in % (mean ± sem) |
| Sociability | WT + Vehicle—mouse | 75.81 ± 6.79 |
| WT + Vehicle—object | 24.19 ± 6.79 | |
| Nf1+/− + Vehicle—mouse | 80.42 ± 4.27 | |
| Nf1+/− + Vehicle—object | 19.58 ± 4.27 | |
| Nf1+/− + SB—mouse | 85.10 ± 2.54 | |
| Nf1+/− + SB—object | 14.90 ± 2.54 | |
| Nf1+/− + CPPQ—mouse | 80.74 ± 3.20 | |
| Nf1+/− + CPPQ—object | 19.26 ± 3.20 | |
| Nf1+/− + Rapa—mouse | 87.27 ± 3.27 | |
| Nf1+/− + Rapa—object | 12.73 ± 3.27 | |
| Short-term memory | WT + Vehicle—novel | 78.12 ± 3.33 |
| WT + Vehicle—familiar | 21.88 ± 3.33 | |
| Nf1+/− + Vehicle—novel | 73.24 ± 3.61 | |
| Nf1+/− + Vehicle—familiar | 26.76 ± 3.61 | |
| Nf1+/− + SB—novel | 70.78 ± 4.16 | |
| Nf1+/− + SB—familiar | 29.22 ± 4.16 | |
| Nf1+/− + CPPQ—novel | 75.98 ± 2.95 | |
| Nf1+/− + CPPQ—familiar | 24.02 ± 2.95 | |
| Nf1+/− + Rapa—novel | 74.97 ± 2.99 | |
| Nf1+/− + Rapa—familiar | 25.03 ± 2.99 | |
| Long-term memory | WT + Vehicle—novel | 69.01 ± 3.96 |
| WT + Vehicle—familiar | 30.99 ± 3.96 | |
| Nf1+/− + Vehicle—novel | 49.96 ± 2.58 | |
| Nf1+/− + Vehicle—familiar | 50.02 ± 2.58 | |
| Nf1+/− + SB—novel | 63.18 ± 2.89 | |
| Nf1+/− + SB—familiar | 36.82 ± 2.88 | |
| Nf1+/− + CPPQ—novel | 54.72 ± 2.74 | |
| Nf1+/− + CPPQ—familiar | 45.28 ± 2.74 | |
| Nf1+/− + Rapa—novel | 70.85 ± 3.45 | |
| Nf1+/− + Rapa—familiar | 29.15 ± 3.45 | |
| Object-in-place | WT + Vehicle—« ab » | 69.98 ± 4.31 |
| WT + Vehicle—« cd » | 30.02 ± 4.31 | |
| Nf1+/− + Vehicle—« ab » | 47.13 ± 5.49 | |
| Nf1+/− + Vehicle—« cd » | 52.87 ± 5.49 | |
| Nf1+/− + SB—« ab » | 67.92 ± 5.32 | |
| Nf1+/− + SB—« cd » | 32.08 ± 5.32 | |
| Nf1+/− + CPPQ—« ab » | 50.54 ± 5.18 | |
| Nf1+/− + CPPQ—« cd » | 49.46 ± 5.18 | |
| Nf1+/− + Rapa—« ab » | 68.47 ± 4.89 | |
| Nf1+/− + Rapa—« cd » | 31.53 ± 4.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doucet, E.; Grychowska, K.; Zajdel, P.; Bockaert, J.; Marin, P.; Bécamel, C. Blockade of Serotonin 5-HT6 Receptor Constitutive Activity Alleviates Cognitive Deficits in a Preclinical Model of Neurofibromatosis Type 1. Int. J. Mol. Sci. 2021, 22, 10178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810178
Doucet E, Grychowska K, Zajdel P, Bockaert J, Marin P, Bécamel C. Blockade of Serotonin 5-HT6 Receptor Constitutive Activity Alleviates Cognitive Deficits in a Preclinical Model of Neurofibromatosis Type 1. International Journal of Molecular Sciences. 2021; 22(18):10178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810178
Chicago/Turabian StyleDoucet, Emilie, Katarzyna Grychowska, Pawel Zajdel, Joël Bockaert, Philippe Marin, and Carine Bécamel. 2021. "Blockade of Serotonin 5-HT6 Receptor Constitutive Activity Alleviates Cognitive Deficits in a Preclinical Model of Neurofibromatosis Type 1" International Journal of Molecular Sciences 22, no. 18: 10178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810178