1. Introduction
Intracellular content is viscoelastic [
1,
2] and organized [
3]. It includes the actin cytoskeleton—an elastic polymer mesh that spans the cell volume and has a mesh size of ~50 nm [
4,
5]. This mesh is immersed in a viscoelastic and crowded medium with significant components that are gel-like and even in a solid phase [
1,
2,
6,
7].
Elasticity governs the mechanical properties of two main components of the cellular structure: the elastic cytoskeleton and the crowded viscoelastic intracellular medium [
1,
4]. Following that, elasticity is a dominant mechanical feature of living cells that enables them to maintain their shape and structure [
4]. The cell elasticity also affects the diffusion of unbound molecules in the cytosol through hydrodynamic interactions between the different cell constituents [
1].
The intracellular medium is organized in micro- and macro-compartments [
3]. While the micro-compartments consist of large functional complexes of proteins that are not membrane-engulfed [
7], the macro-compartments are typically engulfed with membranes and include structures such as the endoplasmic reticulum (ER), mitochondria, lysosomes, transport vesicles, secretory vesicles, etc. [
3]. The abundance of such membrane-engulfed structures inside the cell creates multiple and complex diffusion barriers for molecules. In that way, a significant fraction of intracellular molecules experiences confinement as they impinge on intracellular membranes.
In a living cell, the combined effects of elasticity, viscosity, structural disorder, and mechanical work govern molecular diffusion [
8,
9]. Elasticity and structural disorder tend to hinder molecular motion, thus resulting in sub-diffusion, whereas mechanical work may result in extended molecular translocations and super diffusion [
10]. Mechanical work inside living cells plays a significant role in cell physiology, utilizing molecular motors such as kinesin, dynein, and myosin II for direct and indirect transport of intracellular constituents [
11]. Indirect motor activity includes the incoherent fraction of mechanical forces that are applied by molecular motors at multiple sites on the cytoskeleton. These incoherent forces impact the elastic cytoskeleton mesh [
5], which in turn transfers those forces to the adjacent visco-elastic gel/solid macromolecular complexes. These complexes may further transfer those forces to unbound, yet hindered diffusing molecules in this crowded environment via hydrodynamic interactions [
1]. Via that mechanism, intracellular mechanical work impacts important biophysical parameters such as intracellular diffusivity and homogeneity (or entropy) [
12].
Several mathematical models are useful to describe aspects of molecular diffusion in the cell. Elasticity relates to fractional Brownian motion (fBM), which is one of the three main mathematical models that characterize intracellular random walks. A second main model is continuous time random walk (CTRW), which accounts for random walks with arbitrary distributions of translocations and waiting times. Molecular trapping and escape tend to generate such motion. Third, random walk on a fractal (RWF) accounts for molecular diffusion on a percolation structure, which has a non-Gaussian distribution of correlated translocations and relates to structural disorder and crowding [
13]. fBM is a symmetric and a Gaussian diffusion process in which the increments are not independent and are correlated due to the elasticity effect. This stands in contrast to classical CTRW, for which the particles’ motion is not correlated. Notably, fBM and RWF are ergodic, while CTRW is not [
9].
Diffusion is often characterized by its diffusion coefficient
and the power of diffusion α, according to the mean-square displacement relation:
. These parameters are determined by the underlying mechanisms of diffusion. Elasticity tends to lower α in a negative correlation to the tension of the cytoskeletal mesh [
8]. In contrast, intracellular mechanical work tends to increase diffusivity and α [
8,
12]. The contributions of elasticity, structural disorder, and mechanical work to α could be separated by inhibition of intracellular mechanical work. The summation of all of these contributions relates to the total measured α [
12].
Recently, Guggenberger et al. [
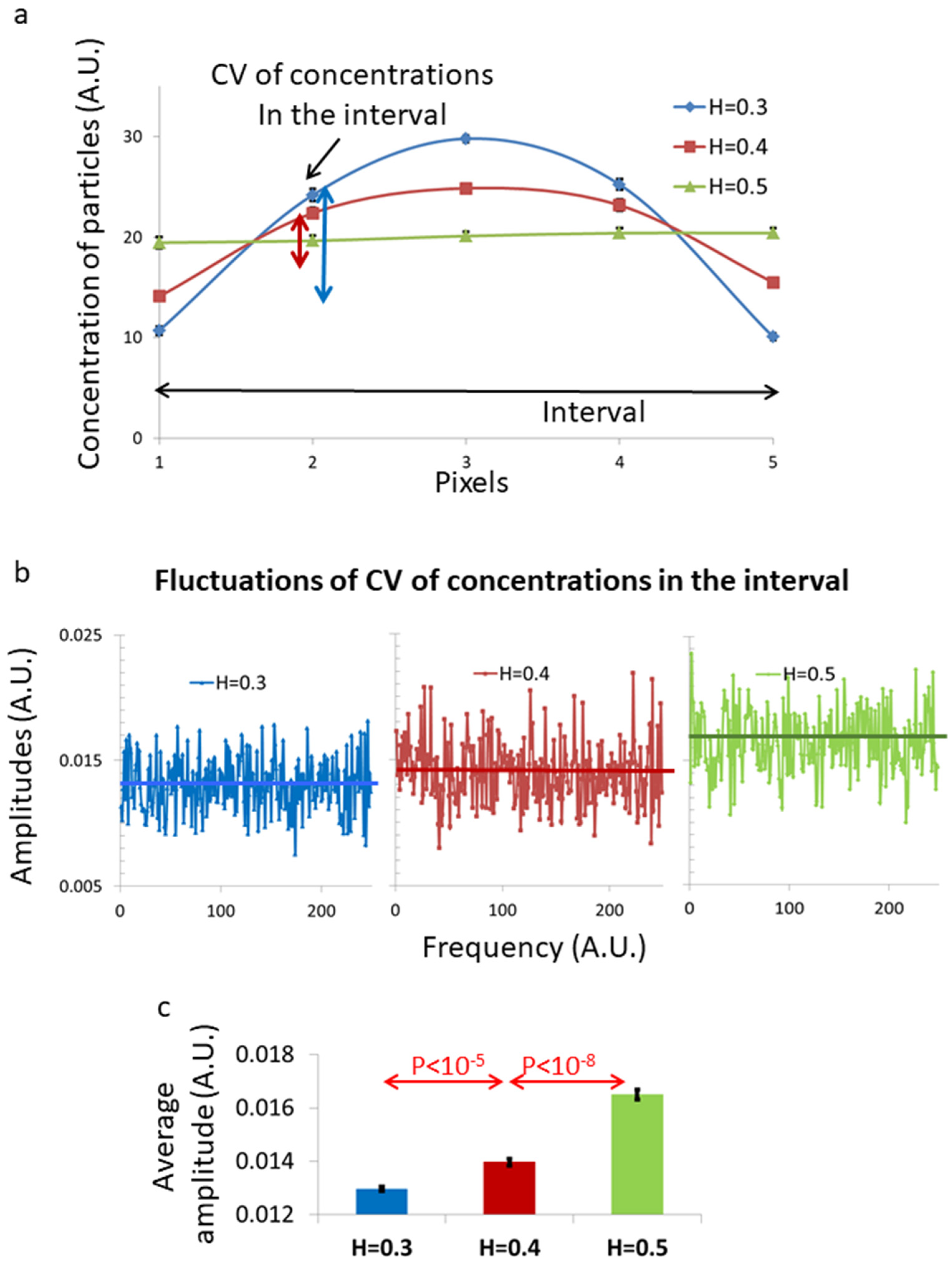
14] suggested theoretically that fBM in a one-dimensional finite interval may create inhomogeneity in the spatial distribution of the diffusing material inside that interval under steady-state conditions. The steady-state patterns of the spatial distribution of the diffusing particles inside that interval depend on the value of the Hurst index (H) of the fBM diffusing process. As shown by Guggenberger et al. [
14], for fBM with H < 0.5, the diffusing particles tend to occupy the space near the interval borders less than at the center of the interval. The opposite occurs for H > 0.5, while for H = 0.5 (Brownian motion), the spatial distribution of the diffusing particles inside the interval is homogenous.
Ions and relatively small molecules undergo diffusion in the cell cytosol and organelles and may serve as mobile tracers for studying diffusion in this complex medium. Ca
++ is an important intracellular messenger and is responsible for multiple cell functions [
15]. Ca
++ is enriched in the endoplasmic reticulum (ER) [
16] and mitochondria [
17]. Fluo-4 AM (ester form) is an advanced fluorescence Ca
++ indicator that enables the simple and sensitive imaging of intracellular and intra-organelle Ca
++ levels in live cells [
18]. Its molecular weight is 1097 g/mol and it can readily cross the cell or an organelle membrane via its ester moiety. Afterwards, intracellular esterases remove the ester moiety, and the Fluo-4 that now binds Ca
++ can no longer cross membranes or exit the cell or the organelles [
19]. As such, Ca
++-Fluo-4 complexes may serve as an important example of intracellular molecules that organize inside multiple membrane-bound compartments and organelles.
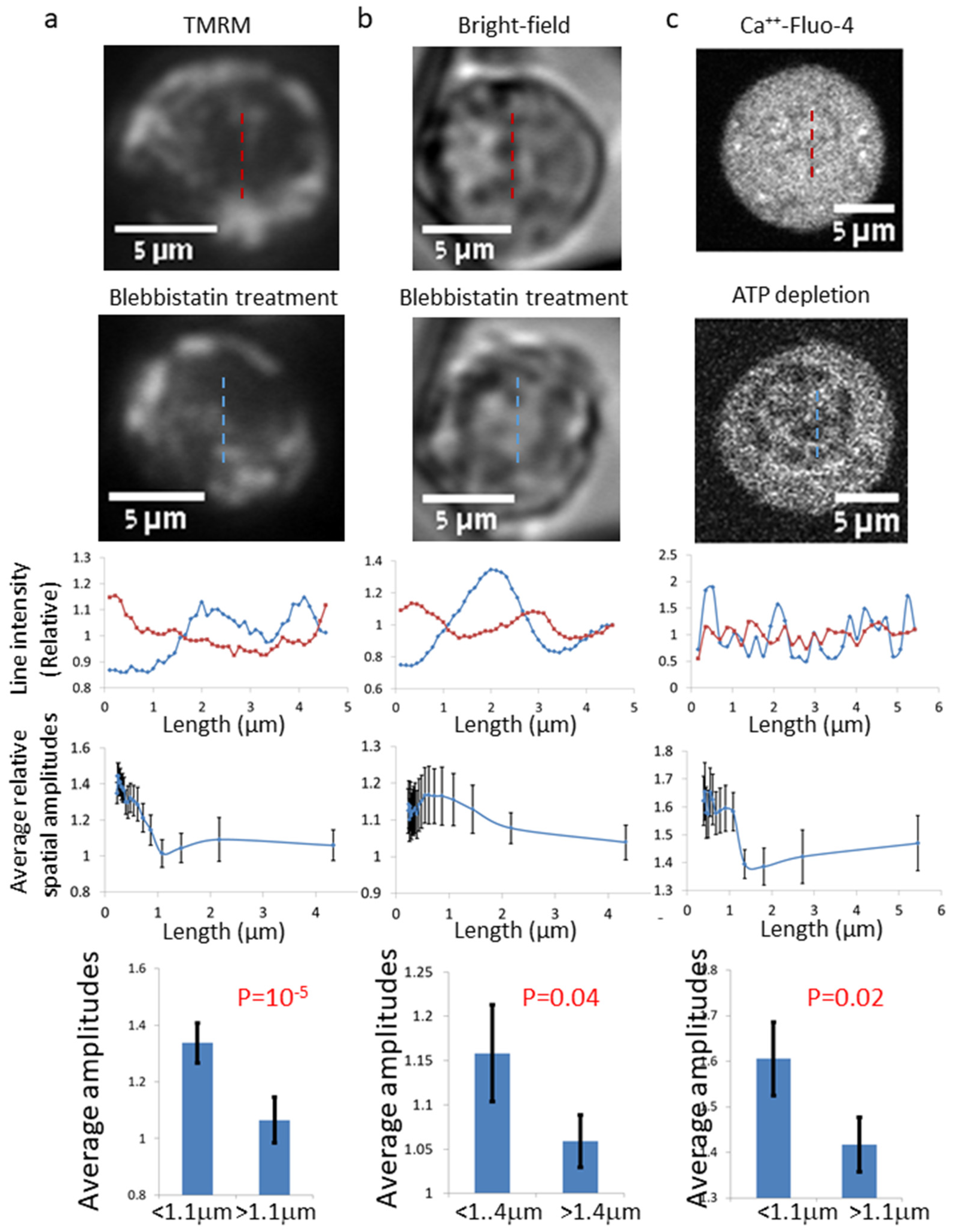
Here, we aimed to study the dynamic organization of intracellular content in relation to intracellular organelles, especially in relation to the ER, which is the largest organelle and spans most of the cytoplasmic volume. For that, we studied the spatio-temporal organization of Ca++-Fluo-4 in Jurkat cells. We initially tested the effect of inhibition of intracellular mechanical work on the spatial and temporal distributions of imaged intracellular Ca++-Fluo-4 in living cells. In order to inhibit intracellular mechanical work, we utilized blebbistatin. This drug is an inhibitor of myosin II motors, which interact with the actin cytoskeletal network. We found that cell treatment with blebbistatin tends to reduce the Ca++-Fluo-4 homogeneity while reducing intracellular dynamics. We found similar effects in intracellular features, identified in label-free bright-field (BF) images of living cells before and after blebbistatin treatment, in mitochondria-highlighted cells and in ATP-depleted cells. Finally, we suggest a mechanism related to intracellular elasticity and fBM that could become detectable after blebbistatin treatment of the cells. Specifically, fBM may significantly affect the organization of material content in intracellular compartments, and thus determine overall intracellular homogeneity, and possibly the functional state of the cell and its physiology.
3. Discussion
Here, we studied the effect of myosin II and viscoelastic diffusion (especially via fBM) on the organization of intracellular content. For that, we visualized intracellular Ca
++-Fluo-4, TMRM, and label-free brightfield images in live Jurkat T cells before and after mechanical work inhibition by blebbistatin. Ca
++, highlighted by Fluo-4, is localized in intracellular organelles such as ER and mitochondria, while TMRM is localized inside active mitochondria. The relation of Ca
++ repositories to the ER was confirmed by double staining of live cells with Fluo-4 and ER tracker red (
Supplementary Figure S3). We found that inhibiting intracellular mechanical work by myosin II through blebbistatin treatment increased cellular dis-homogeneity of Ca
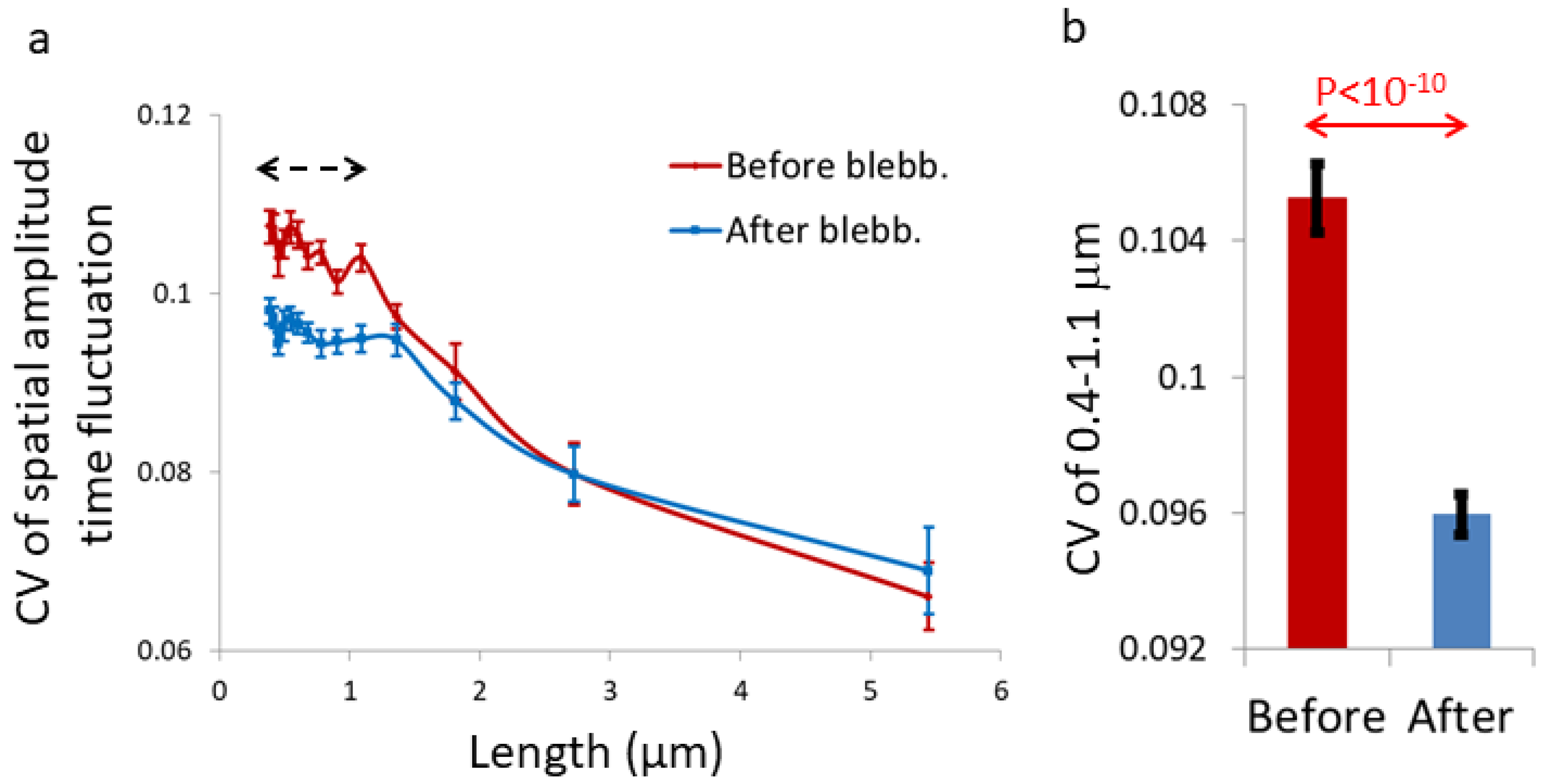
++-rich features in length scale < 1.1 μm. We detected a similar effect in TMRM and label-free BF cell images.
Notably, ER structural support and dynamic growth is mainly related to the microtubules network along with its molecular motors: kinesin 1 and dynein, and not to myosin 2 [
20,
21]. Accordingly, inhibiting myosin II by blebbistatin is not expected to impact the ER structure and dynamics. To confirm that, we first analyzed the spatial DFT amplitudes in the ER-stained cells and found no difference in those amplitudes before and after blebbistatin treatment (
Supplementary Figure S3f). Next, we analyzed the TICS results in the ER-stained cells and we found that the slopes to the TICS decays were not changed in those cells after blebbistatin treatment (
Supplementary Figure S3c). On the other hand, TICS results in those cells utilizing the Ca
++-Fluo-4 staining revealed that the correlation decays are smaller in the blebbistatin-treated cells (
Supplementary Figure S3b, in accordance with results in
Figure 3). Following that, we conclude that although blebbistatin treatment reduces Ca
++-Fluo-4 homogeneity and dynamics, it has no significant impact on the ER homogeneity and dynamics. Furthermore, blebbistatin did not change the size of the treated cells (
Supplementary Figure S7). Mitochondrial structure may also be affected by myosin II activity that relates to the mitochondrial fission process [
22]. Still, the time scale of that process is long (about 2 h) [
23], such that it is not expected to significantly impact our results, which were obtained over much shorter time scales.
It is suggested that the morphological changes that we observed in the cells after blebbistatin treatment are related to the effect of blebbistatin because each of the cells was observed before and after blebbistatin treatment. In this way, the individual effect on each cell could be measured and the net effect of blebbistatin (as presented by the relative amplitudes or entropy) could be obtained. Although the cells were observed in a single cell resolution and the results were relative to the effect of blebbistatin, we cannot completely exclude other effects. To further relate the increase in cellular dis-homogeny to the effect of mechanical work inhibition by blebbistatin, we studied Ca
++-Fluo-4-highlighted cells where their mechanical work was inhibited by ATP depletion. The obtained results (
Figure 2c) were highly similar to the results obtained by blebbistatin (
Figure 1).
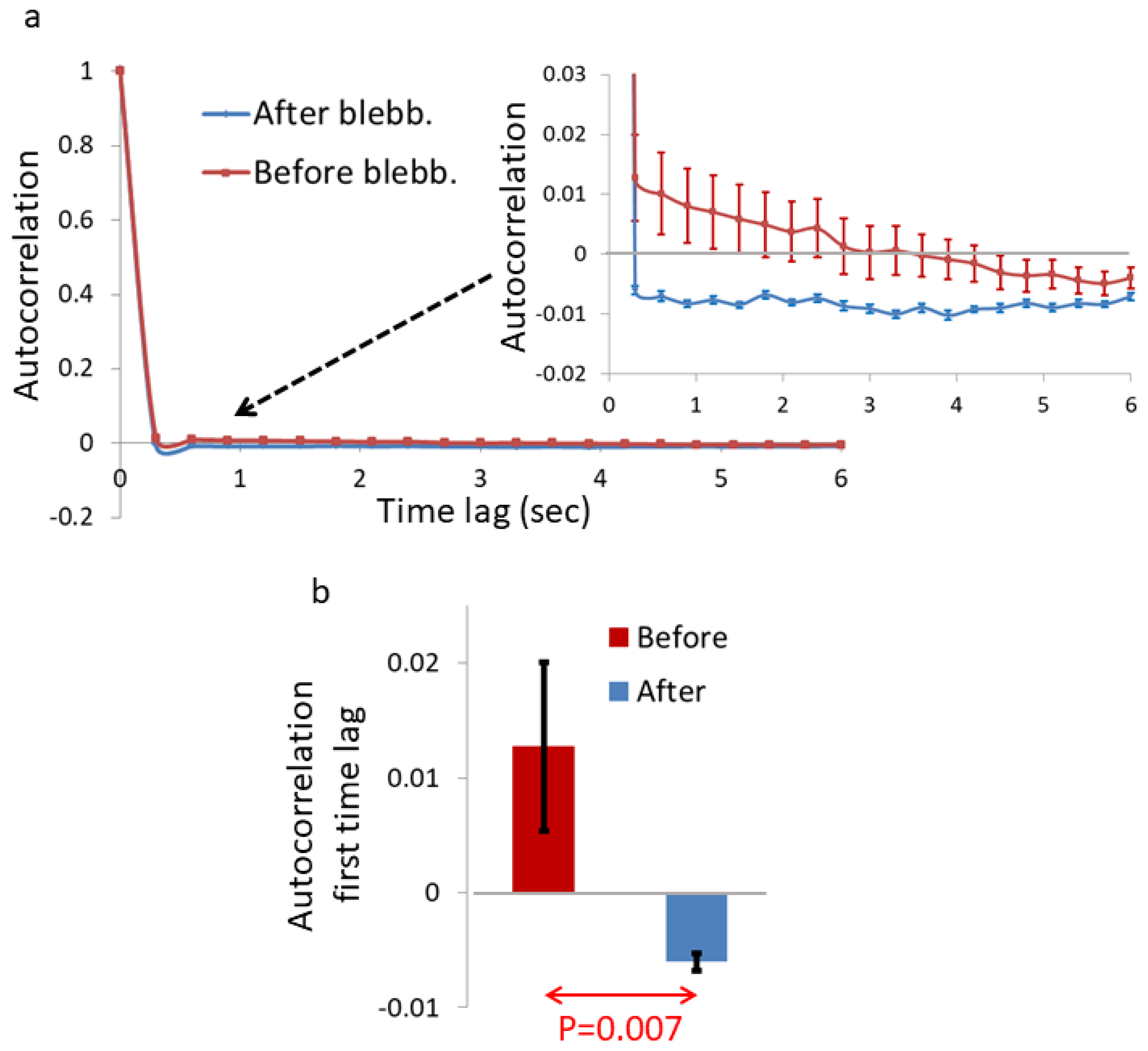
Blebbistatin treatment reduced the dynamics of the Ca++-rich features and generated prominent negative temporal correlations in their signals. We further wanted to account for the apparent changes in Ca++-highlighted complexes and their dynamics via consideration of possible diffusion processes in the cell.
The intracellular medium is viscoelastic [
1]. The cytoplasm is filled with large functional complexes of macromolecules [
24]. Accordingly, as much as half of the cellular protein content may be in a solid phase [
7]. Under this condition, the free volume for diffusion for other macromolecules that are not organized in large complexes is very small. As a result, their motion would displace the position of some of the organized macromolecules through hydrodynamic interactions [
1]. That mechanism would account for the elasticity effect on the unbound moving molecule [
24]. The diffusion coefficient of these mobile molecules under these conditions is expected to be hindered by as much as 10-fold, depending on the size of the molecule [
25]. Even relatively small molecules such as the tracers of Ca
++-Fluo-4 (with a size of ~1 KDa) or TMRM (with a size of ~0.5 KDa) used in this study would undergo viscoelastic diffusion.
As suggested by Guggenberger et al. [
14], fBM dynamics inside a finite spatial segment (or interval) are expected to produce major inhomogeneity in the content of that interval. To complement our experimental findings, we conducted simulations of viscoelastic diffusion due to fBM in living cells. The simulations were conducted over a length scale of 0.8 μm, which is compatible with intracellular organelles such as ER segments or mitochondria. Under these conditions and for H = 0.3, the fBM process produced a three-fold concentration difference between the concentration of the interval content at the center of the interval relative to the concentration close to the borders.
The dynamics of the cell content are influenced by three main factors: intracellular elasticity which relates to fBM, intracellular mechanical work, and intracellular structural disorder [
13]. The first two factors could be more readily controlled by the cell as cross-linking and myosin II interactions determine the actin cytoskeleton elasticity [
26], and since the activity of molecular motors (such as myosin II) determines intracellular mechanical work [
5]. The dynamics of a particular constituent in a particular section of the cell are determined by the combined contributions of fBM controllable dynamics and mechanical work controllable dynamics. These contributions are on top of the contribution of diffusion within a percolation structure. The cell can readily control the extent of contributions from fBM and mechanical work, and the summation of these contributions relates to the total dynamics. As demonstrated in previous work [
12], the power of diffusion α could be separated into two components of α: one that relates to mechanical work and the other that relates to other mechanisms that include the controllable elasticity. Inhibition of mechanical work reduces the total α, which under this condition reflects mainly elasticity and fBM. The α value under this sub-diffusion condition is less than 1 and H < 0.5. In this situation, this sub-diffusive fBM process is expected to induce a significant inhomogeneity in the distribution of the intracellular content due to multiple finite diffusion intervals that are created by the membranes of intracellular organelles.
Following these considerations, we analyzed images of living cells, where their intracellular Ca
++ repositories (mainly in the ER and mitochondria) were labeled with Flu-4 AM fluorescence dye. The Ca
++-Fluo-4 complexes that are created inside the ER or mitochondria cannot cross the organelle membrane and exit the interval due to the removal of the ester moiety of the Fluo-4 by intracellular esterases. Hence, those complexes diffuse in finite intra-organelle intervals while embedded in a crowded viscoelastic gel/solid surrounding medium. Under this condition, the diffusion is inhibited and is most consistent with fBM [
1].
Analyzing the dynamics of these intra-organelle Ca
++-Fluo-4 complexes by TICS revealed that their dynamic is reduced after inhibition of intracellular mechanical work (
Figure 3). We also found that these dynamics are characterized by strong negative autocorrelation values (
Figure 4), which are expected in sub-diffusion fBM dynamics. These results support the assumption that blebbistatin treatment and mechanical work inhibition result in the dominancy of fBM as a mechanism of molecular sub-diffusion. Realizing that, the increase in spatial inhomogeneity after blebbistatin treatment in
Figure 1 is assumed to be related to the increase in inhomogeneity that is caused by a sub-diffusion fBM process in a finite diffusion interval when H < 0.5. The expected stabilization of the dis-homogeneity morphological structures in a sub-diffusion fBM process in a finite interval that was demonstrated in simulations (
Figure 5) was also demonstrated in live Ca
++-Fluo-4-highlighted cells (
Figure 6).
The results of double ER and Ca
++ staining revealed that apart from the significant (and expected) spatial correlation between those highlighted features, the spatial cross-correlation between them decayed more steeply after blebbistatin treatment (
Supplementary Figure S3d,e). These cross-correlation results suggest that blebbistatin affects the combined morphological organization of those two entities. Blebbistatin treatment only affects the morphological organization or dis-homogeneity of Ca
++-Fluo-4 and not the ER (see
Supplementary Figure S3f). Accordingly, it may be assumed that the change in Ca
++-Fluo-4 homogeneity by blebbistatin interferes with the Ca
++-Fluo-4 and ER cross-correlations. Ca
++-Fluo-4 that becomes less homogeneous inside the ER lumen may explain the increased decay of the cross-correlation results that is dominant in length < 1.1 μm.
The similar results that were obtained for TMRM and BF label-free images of cells before and after blebbistatin treatment suggest that this phenomenon that was initially demonstrated for intracellular Ca++ repositories may reflect additional and more general cellular consequences of this mechanism. The ER spans most of the cytoplasm volume. Thus, most of the cytoplasmic materials face a nearby limiting membrane, even if this material is outside the lumen of the ER. Accordingly, it seems that most of the cytoplasmic material, which is predominantly viscoelastic, may be significantly influenced by this mechanism of fBM-induced inhomogeneity. This may explain the similar and general results that we found by BF label-free cell imaging.
It seems that this mechanism that may broadly impact the cytoplasmic degree of inhomogeneity and entropy may also significantly affect the cell physiology. The addition of intracellular mechanical work increases α and counteracts the effect of fBM that induces inhomogeneity. In contrast, increasing the cytoskeleton tension (and elasticity) facilitates this effect that induces inhomogeneity and reduces entropy. In that state of reduced entropy due to sub-diffusive fBM, the creation of functional complexes and even new compounds may be enhanced. The creation of those complexes and new compounds should increase the cell organization and should considerably impact its physiological state.
The cell can control the extent of mechanical work production and also the degree of its cytoskeleton elasticity by actin–myosin interactions and by controlling the degree of cross-linking [
26]. In this way, it may readily control the resultant inhomogeneity and organization. As a considerable fraction of the cytoplasmic content is organized in functional multi-protein complexes, the degree of stability of these complexes is physiologically important. This stability would be directly influenced by the fBM process and facilitated by lower H values. On the other hand, too much inhomogeneity and stabilization will reduce the occurrence of dynamic interactions. In order for the cell to reach its preferable state of functional organization versus dynamics and flexibility, the degree of fBM-induced inhomogeneity or entropy reduction may be tuned. For instance, the amount of mechanical work in malignant cells is higher [
5,
27], while the intracellular tension [
28,
29] and dis-homogeneity [
30] are lower. This may represent a special functional state of high dynamics, entropy and flexibility, along with their consequences. Thus, our suggested mechanism of fBM-induced inhomogeneity may be an important mechanism that affects normal and pathological cells’ physiology.
To conclude, we showed a relation between inhibition of intracellular mechanical work to the organization of intracellular content with possible (patho) physiological implications.
4. Materials and Methods
4.1. Materials
Complete Medium (medium): RPMI-1640, heat-inactivated fetal calf serum (FCS), penicillin, streptomycin, glutamine, sodium pyruvate, and HEPES were from Biological Industries (Kibbutz Beit Haemek, Israel). Blebbistatin, tetramethylrhodamine methyl ester (TMRM), rotenone, and 2-Deoxy-D-Glucose were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fluo-4 and ER tracker red were purchased from ThermoFisher scientific (Waltham, MA, USA).
4.2. Cell Line
Jurkat (human leukemic) E6.1 (CD4+) T cells were available to this study from previous research in the lab. Jurkat cells were maintained in RPMI-1640 medium supplemented with 10% FCS, 100 U/mL penicillin, 100 μg/mL streptomycin, 2% glutamine, 2% sodium pyruvate and 2% HEPES. Cells were maintained in a humidified atmosphere with a 5% CO2/air mixture at 37 °C.
4.3. Sample Preparation
For imaging, coverslip preparation was as follows: coverslips (#1.5 glass chambers, iBidi) were washed with acidic ethanol at room temperature (RT) for 10 min and dried at 37 °C for 1 h. Coverslips were then incubated at RT for 15 min with 0.01% poly-L-lysine (Sigma) diluted in water. This was followed by vacuum aspiration of the poly-L-lysine solution and drying of the coverslips at 37 °C for 1 h. Finally, cells were suspended in imaging buffer (composed of RPMI without phenol red + 10% serum + 25 mM HEPES) at a concentration of 1 million. Subsequently, 100,000–500,000 cells were applied onto the coverslips.
4.4. Treatment of Jurkat Cells with Blebbistatin or Rotenone and 2-Deoxy-D-Glucose for ATP Depletion
Upon completion of measurements in all the cells and after recording the location of each cell, 10 µM of blebbistatin or Rotenone 0.2 mM and 10 mM of 2-deoxy-d-glucose were added to the cell medium. The samples were then incubated for 30 min on the microscope stage. At the end of incubation, each cell was measured again according to its recorded location.
4.5. Fluo-4, ER Tracker Red and Tetramethylrhodamine Methyl Ester (TMRM) Staining
Intracellular Ca++ was imaged using Fluo-4 fluorescence staining. From a working solution of 1 mM Fluo-4 in DMSO, 0.5 μL is added to 106 Jurkat cells in Ca++-enriched imaging buffer solution and 15 μL of probenecid. Ca++-enriched imaging buffer solution has: 125 mM NaCl, 2 mM MgCl2, 4.5 mM KCl and 2 mM CaCl2. Cells were incubated for 1 h in the Fluo-4 solution and afterward, the cells were washed with PBS and then loaded on the coverslip in Ca++-enriched imaging buffer solution. Intracellular Ca++ and ER were both imaged using Fluo-4 and ER tracker red fluorescent staining. The Fluo-4 staining followed the previously described procedure. For ER staining, we added ER tracker red in a final concentration of 1 μM from a working solution of 1 mM in DMSO. The cells were then incubated for 30 min in PBS with calcium, magnesium and dextrose. Afterwards, the cells were washed with PBS and then loaded on the coverslip in Ca++-enriched imaging buffer solution. From a working solution of 100 μM in DMSO, TMRM was added to the cells’ imaging buffer medium at a final concentration of 50 nM. The cells were then incubated for 30 min and then washed with PBS and loaded on the coverslip in imaging buffer solution.
4.6. Microscopy
Bright-field (BF) microscopy:
We conducted BF microscopy of live T cells using a motorized Olympus inverted IX81 microscope (Tokyo, Japan) with a halogen 12 V/100 W lamp light source. The microscope was equipped with a sub-micron Marzhauser-Wetzlar motorized stage type SCAN-IM, with an Lstep controller (Wetzlar-Steindorf, Germany). The condenser NA was 0.55 and the type of illumination was Koehler. A 60× air objective with NA = 0.70 was used for the BF images. Images were collected using a 12-bit cooled, highly sensitive ORCA II C4742-98 camera (Hamamatsu, Japan). Image acquisition time was 1 ms, in 16-bit TIF format, each having 1344 × 1024 pixels, and a physical dimension for each pixel of 110 nm at 60× magnification on the microscope working plane. Image stacks were generated by taking 200 serial images with an acquisition time of 0.5 s per frame. Image binning of 10 sequential gray values was used for the spatial entropy analysis.
Confocal microscopy:
We conducted confocal microscopy of live T cells that were stained with the Fluo-4 fluorescent Ca++ indicator. Cells were scanned using the FV-1200 confocal microscope (Olympus, Japan) equipped with an environmental incubator (temperature and CO2). An image scan was performed using a 60×/1.42 NA oil objective, and for the Flou-4 imaging, excitation was at 488 nm and emission at 505–550 nm. Stacks of images were generated by taking 100 serial images with an acquisition time of 0.3 s for each frame of 140 × 140 pixels (with a 176 nm pixel size).
4.7. Data Analysis
ROIs of 30 × 30 pixels for the confocal microscopy experiment and 40 × 40 pixels for the BF microscopy experiment were determined at the center of each analyzed cell. The ROI size and its location were kept unchanged during the repeated acquisitions. The analyses that are described in the sections below were conducted utilizing those ROIs.
4.8. Image Correlation Spectroscopy (ICS) Calculations
The temporal correlations (TICS) and cross-correlations were calculated for each time-dependent region of interest (ROI) according to a generalized spatiotemporal correlation function [
31]:
where
are the spatial and time coordinates,
are the spatial (horizontal and vertical) and temporal lags respectively, and
stands for light intensity. All TICS correlation values were normalized using the corresponding zero correlations. For each stack of 100 images, a ROI of 30 × 30 pixels was defined at the center of each cell. Then, the normalized temporal correlation values were defined for each ROI in each cell and condition.
4.9. Image Spatial Discrete Fourier Transform (DFT) Analysis
Image spatial discrete Fourier transform (DFT) analysis was conducted for each ROI in each of the 100 time-dependent images. In each dimension, the sequence of pixels intensities along the ROI was analyzed by DFT and the average results of the vertical and horizontal pixels’ sequences of each time-dependent ROI were further averaged to obtain the final spatial DFT of that cell and condition.
4.10. Information Entropy Calculations
For each stack of images, the ROI at the center of each cell was used for the information entropy calculations. The information entropy was calculated for all of the ROI pixels at once or for 36 units of 5 × 5 pixels, which the ROI was divided into, and that were then averaged. Information entropy values were calculated as follows: , where is the probability to obtain intensity i and summation is overall intensity values.
DFT and correlation analyses were carried out using Matlab R2017b (MathWorks, Natick, MA, USA).
4.11. Simulation of fBM
For the simulation of fBM, we used the Matlab script for: Fast (exact) fractional Gaussian noise and Brownian motion generator by B. Scott Jackson (2005).
4.12. Statistical Analyses
The acquired data were exported to Excel spreadsheets (Microsoft Office Professional plus 2010, Microsoft Inc., Redmond, WA, USA) for graph presentation and for statistical analysis with the Real Statistic Resource pack. Significance of differences between groups was calculated using the analysis of variance (ANOVA) single-factor function or t-test for paired samples, with statistical significance set at p < 0.05. The magnitude of deviations from the mean are represented as standard errors of the mean (SEM): .



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





