
Metabolic Toxification of 1,2-Unsaturated Pyrrolizidine Alkaloids Causes Human Hepatic Sinusoidal Obstruction Syndrome: The Update





Abstract
:1. Introduction
2. Literature Search and Source
3. Regulatory Approaches
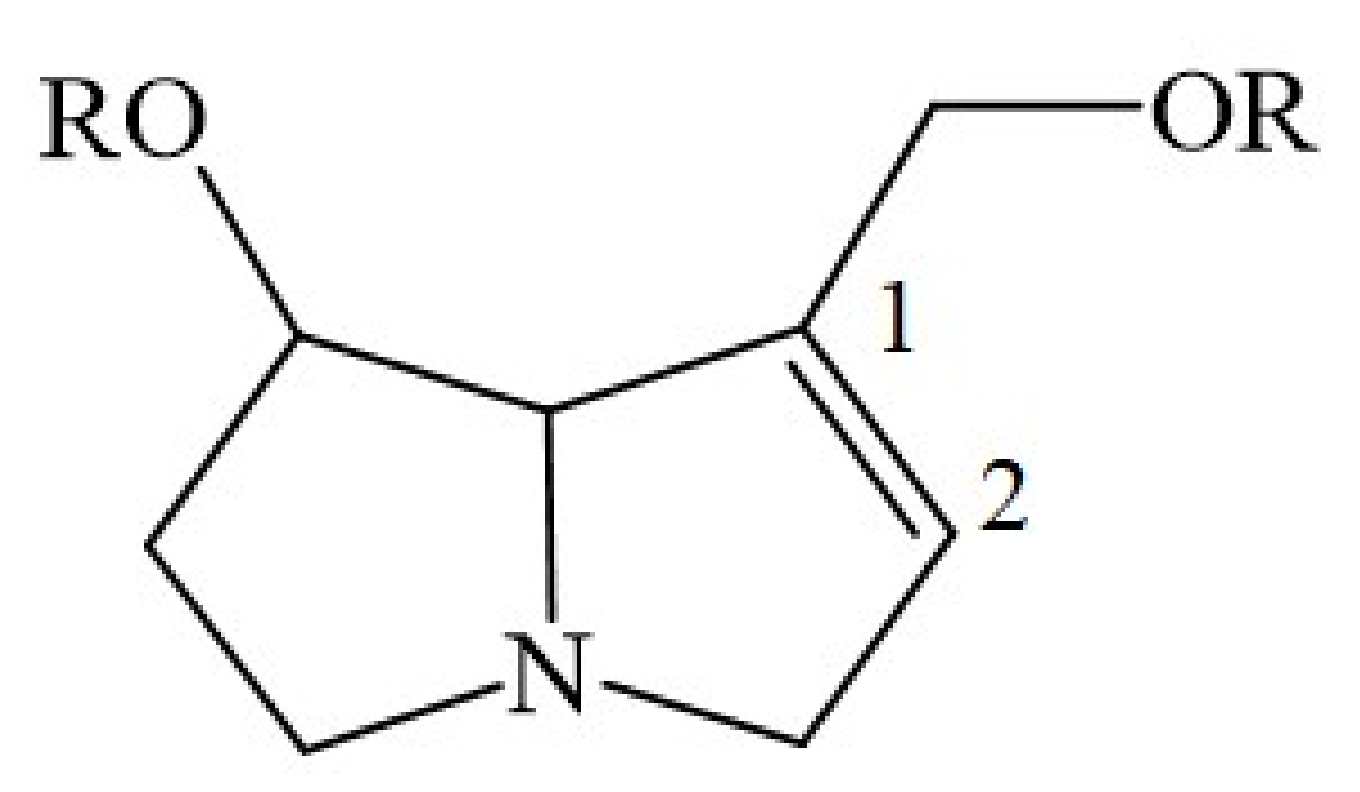
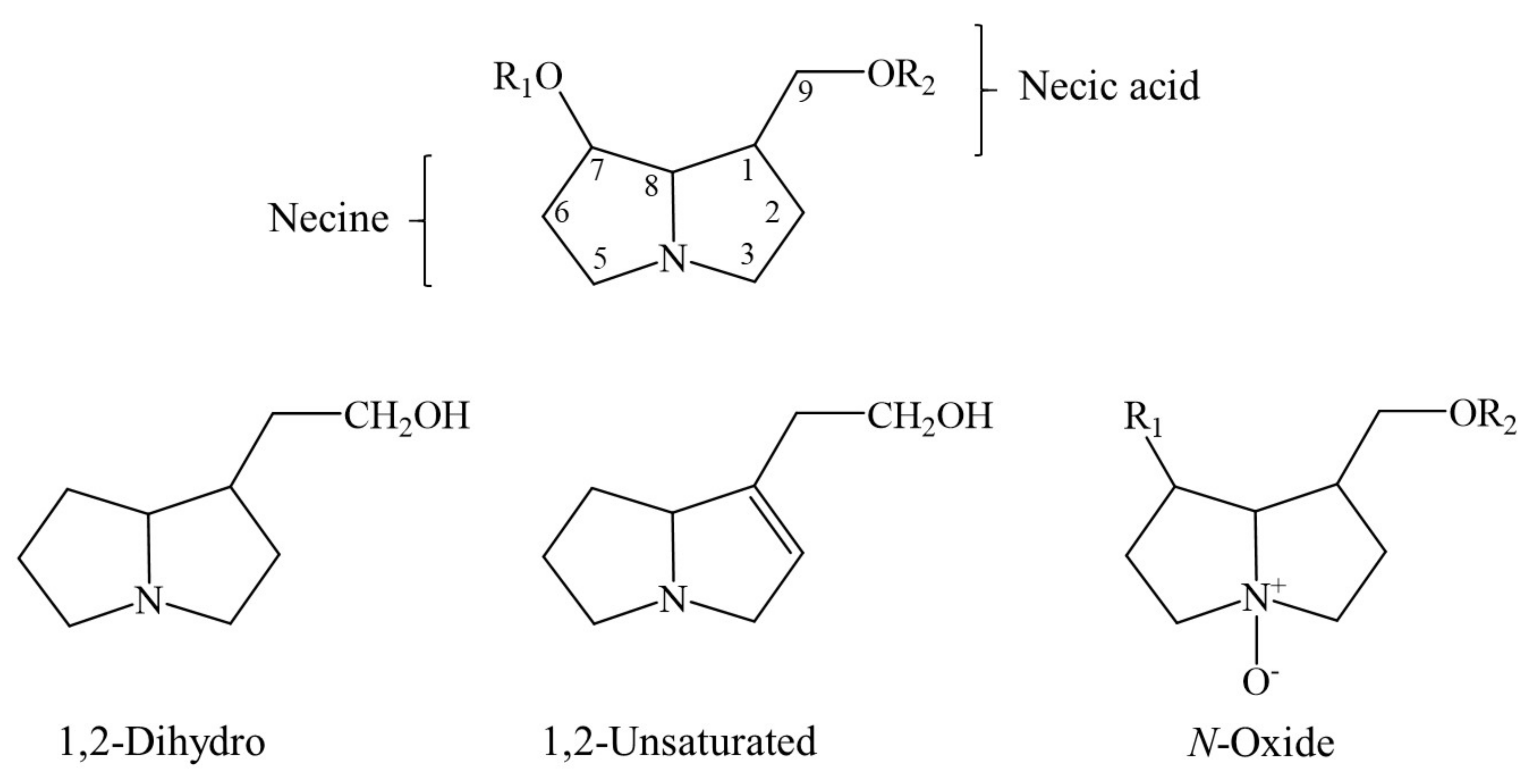
4. Chemical Structure of a Typical 1,2-Unsaturated PA
5. Global Presence and Use of Plants Containing PAs
5.1. General Aspects
5.2. Feed Products Containing PAs
5.3. Plant Families Most Involved in PA Liver Injury
5.4. Products for Human Use Containing Unsaturated PAs Derived from Plants
5.4.1. Bee Products
5.4.2. Vegetables, Including Spices
5.4.3. Bread and Grain
5.4.4. Herbal Teas, Milk, and Dairy Products
5.4.5. Herbal Medicines
6. Interactions of PAs Released from Plants into a Water Environment
7. Uptake of PAs by Plants from Contaminated Water and Soil
8. Principles of Plant PA Biosynthesis
8.1. PAs as Secondary Metabolites
8.2. Synthesis of PAs and Plant Reactive Oxygen Species with Abiotic and Biotic Oxidative Stress
8.3. PA Synthesis, Plant Circadian Clock System, and Seasonal Variation
8.4. Site of PA Biosynthesis in Plants
8.5. The Enzymatic Steps of PA Synthesis in Plants
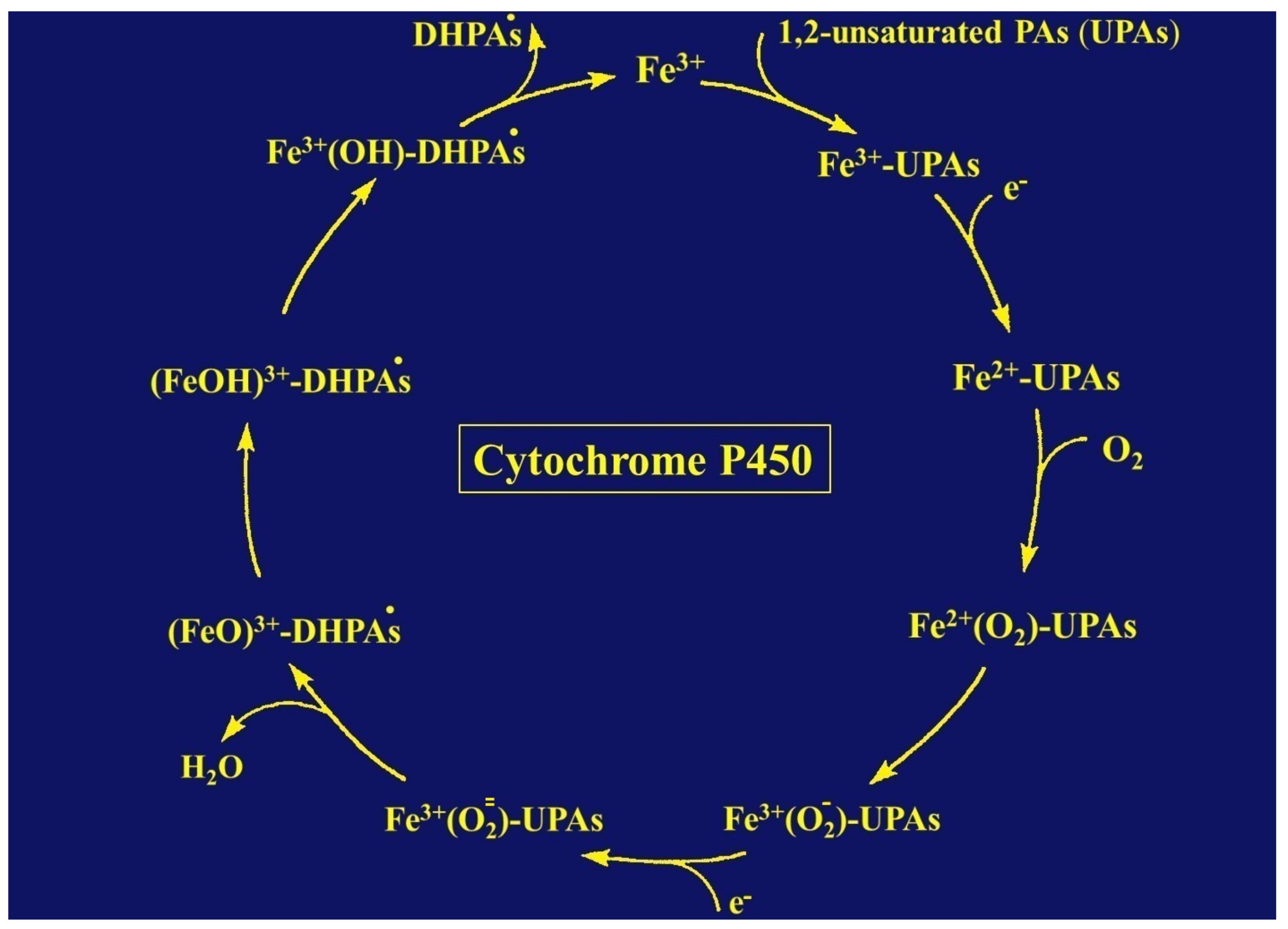
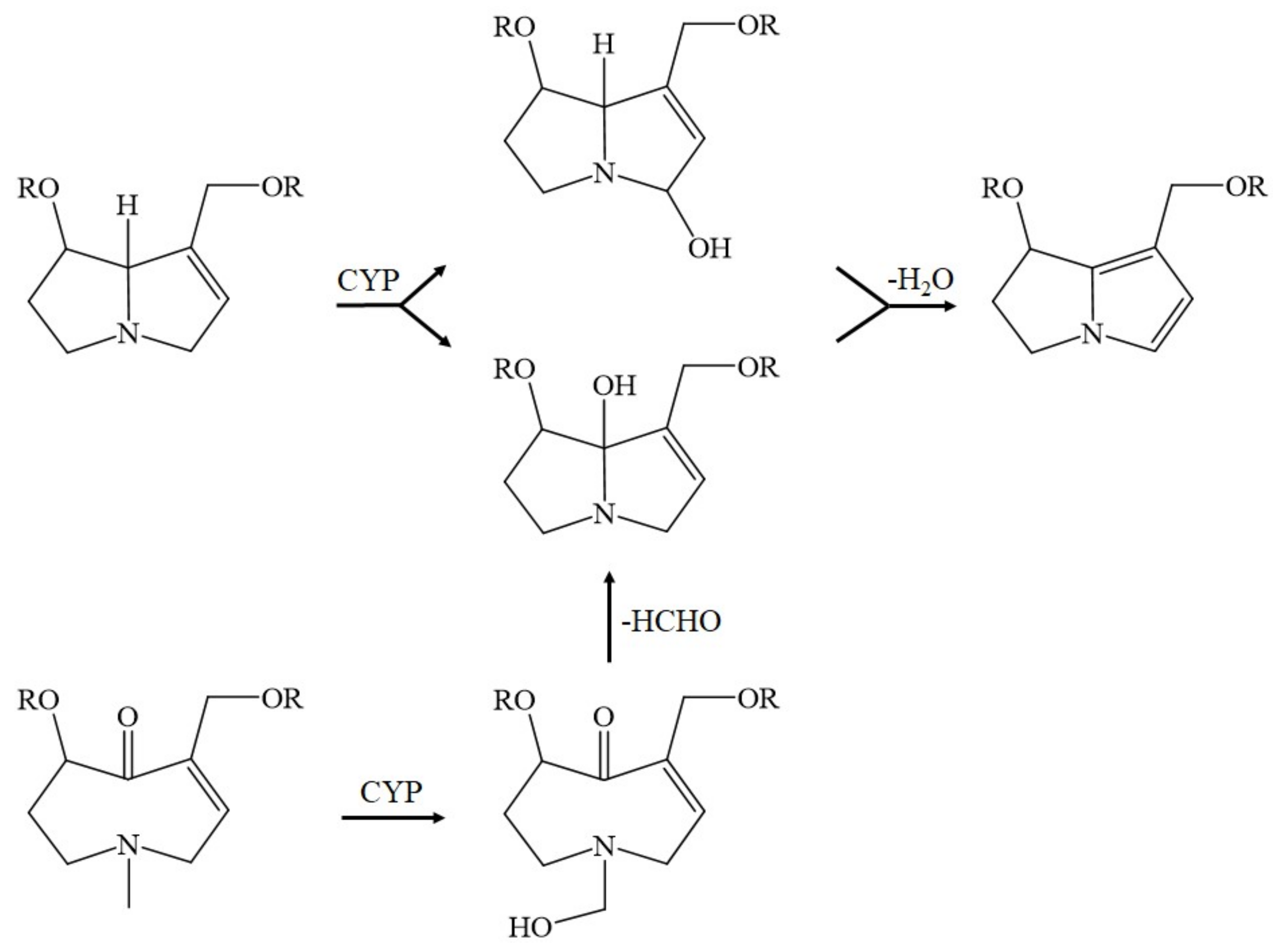
9. The Role of Cytochrome P450 in Metabolizing and Toxifying Unsaturated PAs
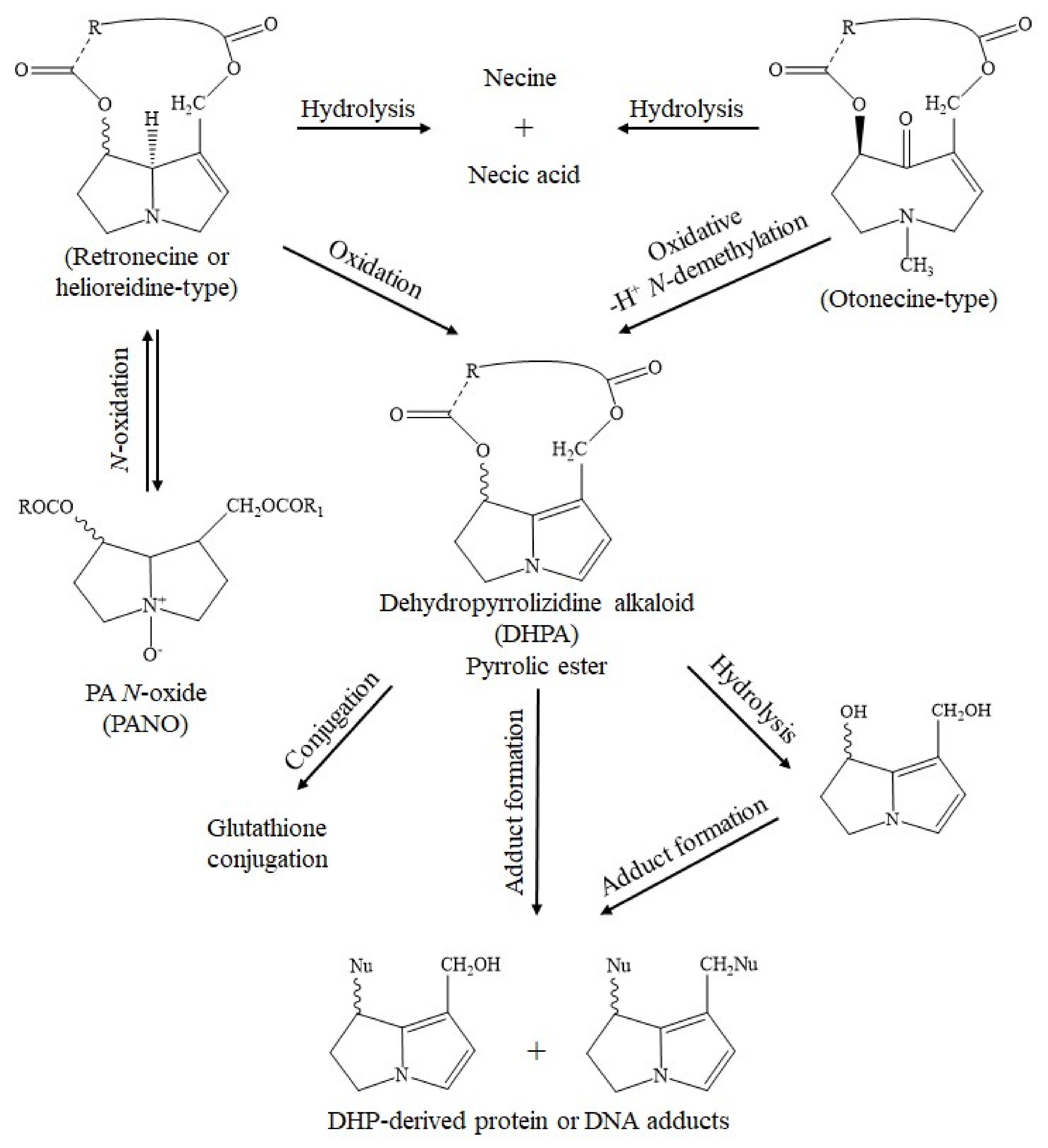
10. Overview of the Mechanistic Molecular Aspects of 1,2-Unsaturated PAs in Clinical Toxicity
11. Clinical Specifics of HSOS Caused by 1,2-Unsaturated PAs
11.1. Definition of HSOS
11.2. Classification of PA-HSOS
11.3. Epidemiology
11.4. Clinical Characteristics
11.5. Routine Laboratory Data
11.6. Specific Diagnostic Biomarkers
11.7. Imaging Features
11.8. Liver Histology
11.9. Specific Causality Assessment Using RUCAM
11.10. Treatment and Prognosis
12. Sporadic PA-HSOS Cases and Small Case Series
12.1. PA-HSOS by Senecio brasiliensis
12.2. PA-HSOS Due to Gynura segetum
12.3. PA-HSOS Related to Gynura segetum
12.4. PA-HSOS Caused by Gynura segetum
12.5. PA-HSOS by Gynura segetum
12.6. PA-HSOS Attributed to Gynura segetum
12.7. PA-HSOS Due to Gynura japonica
12.8. Misdiagnosed PA-HSOS by Petasites hybridus
13. Subtropical and Tropical PA-HSOS Outbreaks
13.1. Afghanistan
13.2. India
13.3. Tadjistan
13.4. Ethiopia
13.5. Iraq
13.6. Uganda
14. Photography of PA-Containing Plants
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Culvenor, C.C.J. Alkaloids and human disease. In Toxicology in the Tropics; Smith, R.L., Bababunmi, E.A., Eds.; Taylor and Francis: London, UK, 1980; pp. 124–141. [Google Scholar]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in chemical structures and biological properties of plant alkaloids. Molecules 2021, 26, 3374. [Google Scholar] [CrossRef]
- Neuman, M.G.; Cohen, L.; Opris, M.; Nanau, R.M.; Hyunjin, J. Hepatotoxicity of pyrrolizidine alkaloids. J. Pharm. Pharm. Sci. 2015, 18, 825–843. [Google Scholar] [CrossRef]
- Teschke, R.; Qi, X.; Xuan, T.D.; Eickhoff, A. Tropical herb induced liver injury by pyrrolizidine alkaloids. In Diagnosis and Management of Tropical Liver Diseases; Debes, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2022; in press. [Google Scholar]
- Wang, X.; Qi, X.; Guo, X. Tusanqi-related sinusoidal obstruction syndrome in China: A systematic review of the literatures. Medicine 2015, 94, e942. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Q.; Ye, J.; Li, X.; Li, Q.; Song, Y.H. Pyrrolizidine alkaloids-induced hepatic sinusoidal obstruction syndrome: Pathogenesis, clinical manifestations, diagnosis, treatment, and outcomes. World J. Gastroent. 2019, 25, 3753–3763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ye, Y.; Yang, X.; Jiao, Y. Systematic review on Chinese herbal medicine induced liver injury. Evid. Based Complement. Alternat. Med. 2016, 2016, 3560812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Qi, X.; Yoshida, E.M.; Méndez-Sánchez, N.; Teschke, R.; Sun, M.; Liu, X.; Su, C.; Deng, J.; Deng, H.; et al. Clinical characteristics and outcomes of traditional Chinese medicine-induced liver injury: A systematic review. Exp. Rev. Gastroenterol. Hepatol. 2018, 12, 425–434. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Teschke, R. Potential hepatotoxins found in herbal medicinal products: A systematic review. Int. J. Mol. Sci. 2020, 21, 5011. [Google Scholar] [CrossRef]
- Teschke, R.; Zhu, Y.; Jing, J. Herb induced liver injury (HILI) in the Asian region and current role of RUCAM for causality assessment in 11,160 published cases: Analysis and outlook. J. Clin. Transl. Hepatol. 2020, 8, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Teschke, R. Traditional Chinese medicine (TCM) and herb induced liver injury: Comparison with drug induced liver injury. J. Clin. Transl. Hepatol. 2018, 6, 57–68. [Google Scholar]
- Jiang, Y.; Zhang, T.; Kusumanchi, P.; Han, S.; Yang, Z.; Liangpunsakul, S. Alcohol metabolizing enzymes, microsomal. ethanol oxidizing system, cytochrome P450 2E1, catalase, and aldehyde dehydrogenase in alcohol-associated liver disease. Biomedicines 2020, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R. Alcoholic liver disease: Alcohol metabolism, cascade of molecular mechanisms, cellular targets, and clinical aspects. Biomedicines 2018, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R. Alcoholic liver disease: Current mechanistic aspects with focus on their clinical relevance. Biomedicines 2019, 7, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Sánchez, N.; Valencia-Rodriguez, A.; Vera-Barajas, A.; Abenavoli, L.; Scarpellini, E.; Ponciano-Rodriguez, G.; Wang, D.Q. The mechanism of dysbiosis in alcoholic liver disease leading to liver cancer. Hepatoma Res. 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Lu, C.; Shen, R.; Lu, T.; Ma, B.; Hua, Y. Knowledge mapping of drug-induced liver injury: A scientometric investigation (2010–2019). Front. Pharmacol. 2020, 11, 842. [Google Scholar] [CrossRef]
- Rosenberg, J.J.; Higley, C.; Shabazi, S.; Lewis, J.H. Selected highlights and controversies of drug-induced liver injury from the recent literature. World J. Gastroenterol. Hepatol. Endosc. Res. 2020, 1, 1–16. [Google Scholar]
- Teschke, R.; Danan, G. Idiosyncratic drug induced liver injury, cytochrome P450, metabolic risk factors, and lipophilicity: Highlights and controversies. Int. J. Mol. Sci. 2021, 22, 3441. [Google Scholar] [CrossRef]
- Teschke, R.; Danan, G. Idiosyncratic drug-induced liver injury (DILI) and herb-induced liver injury (HILI): Diagnostic algorithm based on the quantitative Roussel Uclaf Causality Assessment Method (RUCAM). Diagnostics 2021, 11, 458. [Google Scholar] [CrossRef]
- Teschke, R. Aliphatic halogenated hydrocarbons: Liver injury in 60 patients. J. Clin. Translat. Hepatol. 2018, 28, 350–361. [Google Scholar]
- BfR. Federal institute for Risk Assessment (Bundesinstitut für Risikobewertung, BfR in short). Pyrrolizidinalkaloid: Gehalte in Lebensmitteln Sollen Nach Wie VOR so weit Wie Möglich Gesenkt Werden. Available online: https://www.bfr.bund.de/cm/343/pyrrolizidinalkaloide-gehalte-in-lebensmitteln-sollen-nach-wie-vor-so-weit-wie-moeglich-gesenkt-werden.pdf (accessed on 8 July 2021).
- EMA (European Medicines Agency). Public Statement on the Use of Herbal Medicinal Products Containing Toxic, Unsaturated Pyrrolizidine Alkaloids (PAs) Including Recommendations Regarding Contamination of Herbal Medicinal Products with Pyrrolizidine Alkaloids. Available online: https://www.ema.europa.eu/en/documents/public-statement/public-statement-use-herbal-medicinal-products-containing-toxic-unsaturated-pyrrolizidine-alkaloids_en.pdf (accessed on 8 July 2020).
- EFSA (European Food Safety Authority) Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for human health related to the presence of pyrrolizidine alkaloids in honey, tea, herbal infusions and food supplements. EFSA J. 2017, 15, e04908. [Google Scholar] [CrossRef] [Green Version]
- EU. Commission Regulation (EU) 2020/2040 of 11 December 2020 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Pyrrolizidine Alkaloids in Certain Foodstuffs (Text with EEA Relevance), ELI. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32020R2040 (accessed on 8 July 2021).
- Martins, C.H.Z.; Cunha, B.P.; Solferini, V.N.; Trigo, J.R. Feeding on host plants with different concentrations and structures of pyrrolizidine alkaloids impacts the chemical-defense effectiveness of a specialist herbivore. PLoS ONE 2015, 10, e0141480. [Google Scholar] [CrossRef]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, propolis, and royal jelly: A comprehensive review of their biological actions and health benefits. Oxid. Med. Cell Longev. 2017, 2017, 1259510. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, E.; Sieradzki, Z.; Kwiatek, K. Determination of pyrrolizidine alkaloids in honey with sensitive gas chromatography-mass spectrometry method. Food Anal. Meth. 2018, 11, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Dübecke, A.; Beckh, G.; Lüllmann, C. Pyrrolizidine alkaloids in honey and bee pollen. Food Addit. Contam. 2011, 28, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Letsyo, E.; Jerz, G.; Winterhalter, P.; Dübecke, A.; von der Ohe, W.; von der Ohe, K.; Beuerle, T. Pyrrolizidine alkaloids in floral honey of tropical Ghana: A health risk assessment. Food Addit. Contam. 2017, 10, 300–310. [Google Scholar] [CrossRef]
- Gottschalk, C.; Kaltner, F.; Zimmermann, M.; Korten, R.; Morris, O.; Schwaiger, K.; Gareis, M. Spread of Jacobaea vulgaris and occurrence of pyrrolizidine alkaloids in regionally produced honeys from Northern Germany: Inter- and intra-site variations and risk assessment for special consumer groups. Toxins 2020, 12, 441. [Google Scholar] [CrossRef]
- CBI (Centre for the Promotion of Imports). What is the Demand for Honey in Europe? Available online: https://www.cbi.eu/market-information/honey-sweeteners/trade-statistics (accessed on 23 July 2021).
- De Jesus Inacio, L.; Lanza, I.; Merlanti, R.; Contiero, B.; Lucatello, L.; Serva, L.; Bisutti, V.; Mirisola, M.; Tenti, S.; Segato, S.; et al. Discriminant analysis of pyrrolizidine alkaloid contamination in bee pollen based on near-infrared data from lab-stationary and portable spectrometers. Eur. Food Res. Technol. 2020, 246, 2471–2483. [Google Scholar] [CrossRef]
- Mulder, P.P.J.; López, P.; Castellari, M.; Bodi, D.; Ronczka, S.; Preiss-Weigert, A.; These, A. Occurrence of pyrrolizidine alkaloids in animal- and plant-derived food: Results of a survey across Europe. Food Addit. Contam. Part A Chem. Anal. Control Expo Risk Assess. 2018, 35, 118–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugnerotto, P.; Seraglio, S.K.T.; Schulz, M.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Pyrrolizidine alkaloids and beehive products: A review. Food Chem. 2021, 342, 128384. [Google Scholar] [CrossRef]
- Kast, C.; Kilchenmann, V.; Reinhard, H.; Bieri, K.; Zoller, O. Pyrrolizidine alkaloids: The botanical origin of pollen collected during the flowering period of Echium vulgare and the stability of pyrrolizidine alkaloids in bee bread. Molecules 2019, 24, 2214. [Google Scholar] [CrossRef] [Green Version]
- Schramm, S.; Köhler, N.; Rozhon, W. Pyrrolizidine alkaloids: Biosynthesis, biological activities and occurrence in crop plants. Molecules 2019, 24, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegemann, T.; Kruse, L.H.; Brütt, M.; Ober, D. Specific Distribution of pyrrolizidine alkaloids in floral parts of Comfrey (Symphytum officinale) and its implications for flower ecology. J. Chem. Ecology 2019, 45, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Flade, J.; Beschow, H.; Wensch-Dorendorf, M.; Plescher, A.; Wätjen, W. Occurrence of nine pyrrolizidine alkaloids in Senecio vulgaris L. depending on developmental stage and season. Plants 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA (European Food Safety Authority) Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on pyrrolizidine alkaloids in food and feed. EFSA J. 2011, 9, 2406. Available online: https://efsa.onlinelibrary.wiley (accessed on 8 July 2021). [CrossRef]
- Chen, L.; Mulder, P.P.J.; Peijnenburg, A.; Rietjens, I.M.C.M. Risk assessment of intake of pyrrolizidine alkaloids from herbal teas and medicines following realistic exposure scenarios. Food Chem. Toxicol. 2019, 130, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Xuan, T.D. Suspected herb induced liver injury by green tea extracts: Critical review and case analysis applying RUCAM for causality assessment. Jap. J. Gastroenterol. Hepatol. 2019, 1, 1–16. [Google Scholar]
- Mulder, P.P.J.; Klijnstra, M.D.; Goselink, R.M.A.; van Vuuren, A.M.; Cone, J.W.; Stoopen, G.; Hoogenboom, R.L.A.P. Transfer of pyrrolizidine alkaloids from ragwort, common groundsel and viper’s bugloss to milk from dairy cows. Food Addit. Contam. Part A Chem. Anal. Control Expo Risk Assess. 2020, 37, 1906–1921. [Google Scholar] [CrossRef] [PubMed]
- Nardin, T.; Piasentier, E.; Barnaba, C.; Romanzin, A.; Larcher, R. Targeted and untargeted alkaloid characterisation of pasture herbs and milk from eastern Italian Alps using high resolution mass spectrometry. In Mountain Pastures and Livestock Farming Facing Uncertainty: Environmental, Technical and Socio-Economic Challenges, Proceedings of the 19th Meeting of the FAO-CIHEAM Mountain Pastures Subnetwork, Zaragoza, Spain, 14–16 June 2016; Casasús, I., Lombardi, G., Eds.; CIHEAM: Zaragoza, Spain, 2016; pp. 223–226. Available online: http://om.ciheam.org/om/pdf/a116/00007449.pdf (accessed on 8 July 2021).
- De Nijs, M.; Mulder, P.P.J.; Klijnstra, M.D.; Driehuis, F.; Hoogenboom, R.L.A.P. Fate of pyrrolizidine alkaloids during processing of milk of cows treated with ragwort. Food Addit. Contam. Part A 2017, 34, 2212–2219. [Google Scholar] [CrossRef]
- Msomi, N.Z.; Simelane, M.B.C. Herbal Medicine; InTech: Rijeka, Croatia, 2019; pp. 215–227. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.W.; Wu, T.X.; Shang, H.C.; Li, Y.P.; Altman, D.G.; Moher, D.; Bian, Z.X.; CONSORT-CHM Formulas 2017 Group. CONSORT Extension for Chinese Herbal Medicine Formulas 2017: Recommendations, Explanation, and Elaboration. Ann. Intern. Med. 2017, 167, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Melchart, D.; Hager, S.; Albrecht, S.; Dai, J.; Weidenhammer, W.; Teschke, R. Herbal Traditional Chinese Medicine and suspected liver injury: A prospective study. World J. Hepatol. 2017, 18, 1141–1157. [Google Scholar] [CrossRef]
- Roeder, E. Medicinal plants in Europe containing pyrrolizidine alkaloids. Pharmazie 1995, 50, 83–93. [Google Scholar] [PubMed]
- Roeder, E.; Wiedenfeld, H.; Edgar, J.A. Pyrrolizidine alkaloids in medicinal plants from North America. Pharmazie 2015, 6, 357–367. [Google Scholar] [CrossRef]
- Roeder, E. Medicinal plants in China containing pyrrolizidine alkaloids. Pharmazie 2000, 55, 711–726. [Google Scholar] [PubMed]
- Roeder, E.; Wiedenfeld, H. Pyrrolizidine alkaloids in medicinal plants of Mongolia, Nepal and Tibet. Pharmazie 2009, 64, 699. [Google Scholar]
- Roeder, E.; Wiedenfeld, H. Plants containing pyrrolizidine alkaloids used in the traditional Indian medicine-including ayurveda. Pharmazie 2013, 68, 83–92. [Google Scholar] [PubMed]
- Arseculeratne, S.N.; Gunatilaka, A.A.; Panabokke, R.G. Studies on medicinal plants of Sri Lanka: Occurrence of pyrrolizidine alkaloids and hepatotoxic properties in some traditional medicinal herbs. J. Ethnopharmacol. 1981, 4, 159–177. [Google Scholar] [CrossRef]
- Ebrahimi, F.; Azadbakht, M.; Akbari, F.; Davoodi, A.M.; Godarzi, S.; Azadbakht, M. Qualitative and quantitative study of pyrrolizidine alkaloids in medicinal plants sold in herb shops. J. Maz. Univ. Med. Sci. 2020, 30, 109–114. Available online: http://jmums.mazums.ac.ir/article-1–14864-en.html (accessed on 8 July 2021).
- Kopp, T.; Abdel-Tawab, M.; Mizaikoff, B. Extracting and analyzing pyrrolizidine alkaloids in medicinal plants: A review. Toxins 2020, 12, 320. [Google Scholar] [CrossRef]
- El-Shazly, A.; El-Domiaty, M.; Witte, L.; Wink, M. Pyrrolizidine alkaloids in members of the Boraginaceae from Sinai (Egypt). Biochem. Syst. Ecol. 1998, 26, 619–636. [Google Scholar] [CrossRef]
- Roeder, E.; Wiedenfeld, H.; Schraut, R. Pyrrolizidine alkaloids from Alkanna tinctoria. Phytochemistry 1984, 23, 2125–2126. [Google Scholar] [CrossRef]
- Hendriks, H.; Bruins, A.P.; Huizing, H.J. Detection of Curassavine and some related pyrrolizidine alkaloids in an Anchusa officinalis strain by means of positive ion and negative ion chemical ionization GC/MS. Biol. Mass Spectrom. 1988, 17, 129–132. [Google Scholar] [CrossRef]
- Larson, K.M.; Roby, M.R.; Stermitz, F.R. Unsaturated pyrrolizidines from borage (Borago officinalis), a common garden herb. J. Nat. Prod. 1984, 47, 747–748. [Google Scholar] [CrossRef]
- Dodson, C.D.; Stermitz, F.R. Pyrrolizidine alkaloids from Borage (Borago officinalis) Seeds and Flowers. J. Nat. Prod. 1986, 49, 727–728. [Google Scholar] [CrossRef]
- Herrmann, M.; Joppe, H.; Schmaus, G. Thesinine-4′-O-β-D-glucoside the first glycosylated plant pyrrolizidine alkaloid from Borago officinalis. Phytochemistry 2002, 60, 399–402. [Google Scholar] [CrossRef]
- Benn, M.; Gul, W. Pyrrolizidine alkaloids in the antipodean genus Brachyglottis (Asteraceae). Biochem. Syst. Ecol. 2007, 35, 676–681. [Google Scholar] [CrossRef]
- Bai, Y.; Benn, M. The alkaloids of Brachyglottis hectorii. Arkat USA 2006, 3, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Tundis, R.; Loizzo, M.R.; Statti, G.A.; Passalacqua, N.G.; Peruzzi, L.; Menichini, F. Pyrrolizidine alkaloid profiles of the Senecio cineraria group (Asteraceae). Z. Nat. Sect. C J. Biosci. 2007, 62, 467–472. [Google Scholar] [CrossRef]
- El-Shazly, A. Pyrrolizidine alkaloid profiles of some Senecio species from Egypt. Z. Nat. Sect. C J. Biosci. 2002, 57, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedenfeld, H.; Montes, C.; Tawil, B.; Contin, A.; Wynsma, R. Pyrrolizidine alkaloid level in Senecio bicolor (Willd.) Tod, ssp. cineraria (DC.) from Middle Europe. Pharmazie 2006, 61, 559–561. [Google Scholar] [PubMed]
- Asres, K.; Sporer, F.; Wink, M. Pattern of pyrrolizidine alkaloids in 12 Ethiopian Crotalaria species. Biochem. Syst. Ecol. 2004, 32, 915–930. [Google Scholar] [CrossRef]
- El-Shazly, A.; Sarg, T.; Ateya, A.; Abdel Aziz, E.; Witte, L.; Wink, M. Pyrrolizidine alkaloids of Cynoglossum officinale and Cynoglossum amabile (Family Boraginaceae). Biochem. Syst. Ecol. 1996, 24, 415–421. [Google Scholar] [CrossRef]
- Furuya, T.; Hikichi, M. Lindelofine and supinine: Pyrrolizidine alkaloids from Eupatorium stoechadosmum. Phytochemistry 1973, 12, 225. [Google Scholar] [CrossRef]
- Edgar, J.A.; Lin, H.J.; Kumana, C.R.; Ng, M.M. Pyrrolizidine alkaloid composition of three Chinese medicinal herbs, Eupatorium cannabinum, E. japonicum and Crotalaria assamica. Am. J. Chin. Med. 1992, 20, 281–288. [Google Scholar] [CrossRef]
- Pedersen, E. Echinatine and supinine: Pyrrolizidine alkaloids from Eupatorium cannabinum. Phytochemistry 1975, 14, 2086–2087. [Google Scholar] [CrossRef]
- Hendriks, H.; Malingré, T.M.; Elema, E.T. Pyrrolizidine alkaloids, flavonoids and volatile compounds in the genus Eupatorium—Eupatorium cannabinum L., an ancient drug with new perspectives. Pharm. Weekbl. Sci. Ed. 1983, 5, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Colegate, S.M.; Upton, R.; Gardner, D.R.; Panter, K.E.; Betz, J.M. Potentially toxic pyrrolizidine alkaloids in Eupatorium perfoliatum and three related species. Implications for herbal use as boneset. Phytochem. Anal. 2018, 29, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Kast, C.; Kilchenmann, V.; Reinhard, H.; Droz, B.; Lucchetti, M.A.; Dübecke, A.; Beckh, G.; Zoller, O. Chemical fingerprinting identifies Echium vulgare, Eupatorium cannabinum and Senecio spp. as plant species mainly responsible for pyrrolizidine alkaloids in bee-collected pollen. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Lang, G.; Passreiter, C.M.; Medinilla, B.; Castillo, J.J.; Witte, L. Non-toxic pyrolizidine alkaloids from Eupatorium semialatum. Biochem. Syst. Ecol. 2001, 29, 143–147. [Google Scholar] [CrossRef]
- Schulz, M.; Meins, J.; Diemert, S.; Zagermann-Muncke, P.; Goebel, R.; Schrenk, D.; Schubert-Zsilavecz, M.; Abdel-Tawab, M. Detection of pyrrolizidine alkaloids in German licensed herbal medicinal teas. Phytomedicine 2015, 22, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Reina, M.; Gonzalez-Coloma, A.; Gutierrez, C.; Cabrera, R.; Henriquez, J.; Villarroel, L. Pyrrolizidine alkaloids from Heliotropium megalanthum. J. Nat. Prod. 1998, 61, 1418–1420. [Google Scholar] [CrossRef]
- Roeder, E.; Breitmaier, E.; Birecka, H.; Frohlicht, M.W.; Badzies-Crombach, A. Pyrrolizidine alkaloids of Heliotropium spathulatum. Phytochemistry 1991, 30, 1703–1706. [Google Scholar] [CrossRef]
- Ravi, S.; Lakshmanan, A.J.; Herz, W. Iso-lycopsamine, a pyrrolizidine alkaloid from Heliotropium keralense. Phytochemistry 1990, 29, 361–364. [Google Scholar] [CrossRef]
- Mohanraj, S.; Subramanian, P.S.; Herz, W. Minor alkaloids of Heliotropium curassavicum. Phytochemistry 1982, 21, 1775–1779. [Google Scholar] [CrossRef]
- Farsam, H.; Yassa, N.; Sarkhail, P.; Shafiee, A. New pyrrolizidine alkaloids from Heliotropium crassifolium. Planta Med. 2000, 66, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Yassa, N.; Farsam, H.; Shafiee, A.; Rustaiyan, A. Pyrrolizidine alkaloids from Heliotropium esfandiarii. Planta Med. 1996, 62, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, A.; Salimi, M.; Farsam, H.; Yassa, N. Pyrrolizidine alkaloids from Heliotropium dissitiflorum Boiss. Daru 2002, 10, 168–170. [Google Scholar]
- Farrag, N.M.; Abdel-Aziz, E.M.; El-Shafae, A.M.; Ateya, A.M.; El Domiaty, M.M. Pyrrolizidine alkaloids of Heliotropium bacciferum Forssk from Egypt. Int. J. Pharm. 1996, 34, 374–377. [Google Scholar] [CrossRef]
- Zalkow, L.H.; Boxetti, S.; Gelbaum, L.; Gordon, M.; Patil, B.B.; Shaxi, A.; Van Derveer, D. Pyrrolizidine alkaloids from Middle Eastern plants. J. Nat. Prod. 1979, 42, 603–614. [Google Scholar] [CrossRef]
- Lakshmanan, A.J.; Shanmugasundaram, S. Helibractinecine, a pyrrolizidine alkaloid from Heliotropium bracteatum. Phytochemistry 1994, 36, 245–248. [Google Scholar] [CrossRef]
- Mohanraj, S.; Kulanthaivel, P.; Subramanian, P.S.; Herz, W. Helifoline, a pyrrolizidine alkaloid from Heliotropium ovalifolium. Phytochemistry 1981, 20, 1991–1995. [Google Scholar] [CrossRef]
- Lakshmanan, A.J.; Shanmugasundaram, S. Heliscabine, a pyrrolizidine ester alkaloid from Heliotropium scabrum. Phytochemistry 1995, 39, 473–475. [Google Scholar] [CrossRef]
- Krenn, L.; Wiedenfeld, H.; Roeder, E. Pyrrolizidine alkaloids from Lithospermum officinale. Phytochemistry 1994, 37, 275–277. [Google Scholar] [CrossRef]
- Wiedenfeld, H.; Pietrosiuk, A.; Furmanowa, M.; Roeder, E. Pyrrolizidine alkaloids from Lithospermum canescens Lehm. Z. Nat. Sect. C J. Biosci. 2003, 58, 173–176. [Google Scholar] [CrossRef]
- Pietrosiuk, A.; Sykłowska-Baranek, K.; Wiedenfeld, H.; Wolinowska, R.; Furmanowa, M.; Jaroszyk, E. The shikonin derivatives and pyrrolizidine alkaloids in hairy root cultures of Lithospermum canescens (Michx.) Lehm. Plant Cell Rep. 2006, 25, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Roeder, E.; Rengel, B. Pyrrolizidine alkaloids from Lithospermum Erythrorhizon. Phytochemistry 1990, 29, 690–693. [Google Scholar] [CrossRef]
- Mroczek, T.; Glowniak, K.; Wlaszczyk, A. Simultaneous determination of N-oxides and free bases of pyrrolizidine alkaloids by cation-exchange solid-phase extraction and ion-pair high-performance liquid chromatography. J. Chromatogr. A 2002, 949, 249–262. [Google Scholar] [CrossRef]
- Niwa, H.; Ishiwata, H.; Yamada, K. Separation and determination of macrocyclic pyrrolizidine alkaloids of the otonecine type present in the edible plant Petasites japonicus by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1983, 257, 146–150. [Google Scholar] [CrossRef]
- Chizzola, R.; Ozelsberger, B.; Langer, T. Variability in chemical constituents in Petasites hybridus from Austria. Biochem. Syst. Ecol. 2000, 28, 421–432. [Google Scholar] [CrossRef]
- Segall, H.J. Reverse phase isolation of pyrrolizidine alkaloids. J. Liq. Chromatogr. 1979, 2, 429–436. [Google Scholar] [CrossRef]
- Yang, X.; Yang, L.; Xiong, A.; Li, D.; Wang, Z. Authentication of Senecio scandens and S. vulgaris based on the comprehensive secondary metabolic patterns gained by UPLC-DAD/ESI-MS. J. Pharm. Biomed. Anal. 2011, 56, 165–172. [Google Scholar] [CrossRef]
- Borstel, K.V.; Witte, L.; Hartmann, T. Pyrrolizidine alkaloid patterns in populations of Senecio vulgaris, S. vernalis and their hybrids. Phytochemistry 1989, 28, 1635–1638. [Google Scholar] [CrossRef]
- Witte, L.; Ernst, L.; Adam, H.; Hartmannt, T. Chemotypes of two pyrrolizidine alkaloid-containing Senecio species. Phytochemistry 1992, 31, 559–565. [Google Scholar] [CrossRef]
- Schaneberg, B.T.; Molyneux, R.J.; Khan, I.A. Evaporative light scattering detection of pyrrolizidine alkaloids. Phytochem. Anal. 2004, 15, 36–39. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, N.; Choi, F.F.K.; Qiao, C.F.; Song, J.Z.; Li, S.L.; Liu, X.; Cai, Z.W.; Fu, P.P.; Lin, G.; et al. A new approach for simultaneous screening and quantification of toxic pyrrolizidine alkaloids in some potential pyrrolizidine alkaloid-containing plants by using ultra performance liquid chromatography-tandem quadrupole mass spectrometry. Anal. Chim. Acta 2010, 681, 33–40. [Google Scholar] [CrossRef]
- Cheng, D.; Nguyen, V.-T.; Ndihokubwayo, N.; Ge, J.; Mulder, P.P.J. Pyrrolizidine alkaloid variation in Senecio vulgaris populations from native and invasive ranges. PeerJ 2017, 5, e3686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelser, P.B.; De Vos, H.; Theuring, C.; Beuerle, T.; Vrieling, K.; Hartmann, T. Frequent gain and loss of pyrrolizidine alkaloids in the evolution of Senecio section Jacobaea (Asteraceae). Phytochemistry 2005, 66, 1285–1295. [Google Scholar] [CrossRef]
- Bicchi, C.; Rubiolo, P.; Frattini, C.; Sandra, P.; David, F. Off-Line supercritical fluid extraction and capillary gas chromatography of pyrrolidine alkaloids in Senecio species. J. Nat. Prod. 1991, 54, 941–945. [Google Scholar] [CrossRef]
- Christov, V.; Evstatieva, L. Alkaloid profile of Bulgarian species from genus Senecio L. Z. Nat. Sect. C J. Biosci. 2003, 58, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Steixjes, M.E.; Kelley, R.B.; Molyneux, R.J.; Seiber, J.N. Gc-Ms determination of pyrrolizidine alkaloids in four senecio species. J. Nat. Prod. 1991, 54, 759–773. [Google Scholar]
- Mandic, B.; Godjevac, D.; Beskoski, V.; Simic, M.; Trifunovic, S.; Tesevic, V.; Vajs, V.; Milosavljevic, S. Pyrrolizidine alkaloids from seven wild-growing Senecio species in Serbia and Montenegro. J. Serb. Chem. Soc. 2009, 74, 27–34. [Google Scholar] [CrossRef]
- Suau, R.; Cabezudo, B.; Rico, R.; Nájera, F.; López-Romero, J.M.; García, A.I. Pyrrolizidine alkaloids from three Spanish Senecio species. Biochem. Syst. Ecol. 2002, 30, 981–984. [Google Scholar] [CrossRef]
- Pestchanker, M.J.; Ascheri, M.S.; Giordano, O.S. Uspallatine, a pyrrolizidine alkaloid from senecio uspallatensis. Phytochemistry 1985, 24, 1622–1624. [Google Scholar] [CrossRef]
- Were, O.; Benn, M.; Munavu, R.M. The pyrrolizidine alkaloids of Senecio syringifolius and S. hadiensis from Kenya. Phytochemistry 1993, 32, 1595–1602. [Google Scholar] [CrossRef]
- Porter, L.A.; Geissman, T.A. Angularine, a New Pyrrolizidine alkaloid from Senecio angulatus L. J. Org. Chem. 1962, 27, 4132–4134. [Google Scholar] [CrossRef]
- Benn, M.; Were, O. Ruwenine and ruzorine: Pyrrolizidine alkaloids of Senecio ruwenzoriensis. Phytochemistry 1992, 31, 3295–3296. [Google Scholar] [CrossRef]
- Röder, E.; Wiedenfeld, H.; Pfitzer, A. Doriasenine, a pyrrolizidine alkaloid from Senecio doria. Phytochemistry 1988, 27, 4000–4001. [Google Scholar] [CrossRef]
- Bredenkamp, M.W.; Wiechers, A.; van Rooyen, P.H. A new pyrrolizidine alkaloid N-oxide and the revised structure of sceleratine. Tetrahedron Lett. 1985, 26, 5721–5724. [Google Scholar] [CrossRef]
- Furuya, T.; Hikichi, M. Alkaloids and triterpenoids of Symphytum officinale. Phytochemistry 1971, 10, 2217–2220. [Google Scholar] [CrossRef]
- Kim, N.C.; Oberlies, N.H.; Brine, D.R.; Handy, R.W.; Wani, M.C.; Wall, M.E. Isolation of symlandine from the roots of common comfrey (Symphytum officinale) using countercurrent chromatography. J. Nat. Prod. 2001, 64, 251–253. [Google Scholar] [CrossRef]
- Couet, C.E.; Crews, C.; Hanley, A.B. Analysis, separation, and bioassay of pyrrolizidine alkaloids from comfrey (Symphytum officinale). Nat. Toxins 1996, 4, 163–167. [Google Scholar] [CrossRef]
- Mossoba, M.M.; Lin, H.S.; Andrzejewski, D.; Sphon, J.A.; Betz, J.M.; Miller, L.J.; Eppley, R.M.; Trucksess, M.W.; Page, S.W. Application of Gas-Chromatography Matrix-Isolation Fourier-Transform Infrared-Spectroscopy to the Identification of Pyrrolizidine Alkaloids from Comfrey Root (Symphytum-officinale L.). J. Aoac Int. 1994, 60, 945–948. [Google Scholar] [CrossRef]
- Adamczak, A.; Opala, B.; Gryszczyńska, A.; Buchwald, W. Content of pyrrolizidine alkaloids in the leaves of coltsfoot (Tussilago farfara L.) in Poland. Acta Soc. Bot. Pol. 2013, 82, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Nedelcheva, A.; Kostova, N.; Sidjimov, A. Pyrrolizidine alkaloids in Tussilago farfara from Bulgaria. Biotechnol. Biotechnol. Equip. 2015, 29, S1–S7. [Google Scholar] [CrossRef] [Green Version]
- Kopp, T.; Salzer, L.; Abdel-Tawab, M.; Mizaikoff, B. Efficient extraction of pyrrolizidine alkaloids from plants by pressurised liquid extraction—A preliminary study. Planta Med. 2019, 86, 85–90. [Google Scholar] [CrossRef]
- Jeong, W.T.; Lim, H.B. Determination and chemical profiling of toxic pyrrolizidine alkaloids in botanical samples with UPLC–Q-TOFMS. Chromatographia 2019, 82, 1653–1664. [Google Scholar] [CrossRef]
- Kaltner, F.; Stiglbauer, B.; Rychlik, M.; Gareis, M.; Gottschalk, C. Development of a sensitive analytical method for determining 44 pyrrolizidine alkaloids in teas and herbal teas via LC-ESI-MS/MS. Anal. Bioanal. Chem. 2019, 411, 7233–7249. [Google Scholar] [CrossRef]
- Dzuman, Z.; Jonatova, P.; Stranska-Zachariasova, M.; Prusova, N.; Brabenec, O.; Novakova, A.; Fenclova, M.; Hajslova, J. Development of a new LC-MS method for accurate and sensitive determination of 33 pyrrolizidine and 21 tropane alkaloids in plant-based food matrices. Anal. Bioanal. Chem. 2020, 412, 7155–7167. [Google Scholar] [CrossRef] [PubMed]
- Bessaire, T.; Ernest, M.; Christinat, N.; Carrères, B.; Panchaud, A.; Badoud, F. High resolution mass spectrometry workflow for the analysis of food contaminants: Application to plant toxins, mycotoxins and phytoestrogens in plant-based ingredients. Food Addit. Contam. Part A 2021, 38, 978–996. [Google Scholar] [CrossRef]
- Wiedenfeld, H. Plants containing pyrrolizidine alkaloids—Toxicity and problems. Food Addit. Contamin. Part A 2011, 28, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Mesfin, G.; Ali, A.; Seboxa, T.; Bane, A.; Tensae, M.W.; Gebressilassie, S.; Tsegaye, Y.; Haile, T. An epidemiological study into the investigation of liver disease of unknown origin in a rural community of northern Ethiopia. Ethiop. Med. J. 2012, 50 (Suppl. 2), 1–8. [Google Scholar] [PubMed]
- Bane, A.; Seboxa, T.; Mesfin, G.; Ali, A.; Tsegaye, Y.; W/Tensae, M.; G/Selassie, S.; Haile, T. An outbreak of veno-occlusive liver disease in northern Ethiopia, clinical findings. Ethiop. Med. J. 2012, 50 (Suppl. 2), 9–16. [Google Scholar]
- Abebe, D.; Debella, A.; Tekabe, F.; Mekonnen, Y.; Degefa, A.; Mekonnen, A.; Tefera, A.; Mamo, H.; Tsegaye, B. An outbreak of liver disease in Tahtay Koraro Woreda, Tigray region of Ethiopia: A case-control study for the identification of the etiologic agent. Ethiop. Med. J. 2012, 50 (Suppl. 2), 17–25. [Google Scholar] [PubMed]
- Debella, A.; Abebe, D.; Tekabe, F.; Mamo, H.; Abebe, A.; Tsegaye, B.; Ayana, G.; Degefa, A.; Negussie, P.; Yimer, E.; et al. Toxicity study and evaluation of biochemical markers towards the identification of the causative agent for an outbreak of liver disease in Tahtay Koraro Woreda, Tigray. Ethiop. Med. J. 2012, 50 (Suppl. 2), 27–35. [Google Scholar]
- Debella, A.; Abebe, D.; Tekabe, F.; Degefa, A.; Desta, A.; Tefera, A.; Geyid, A.; Mudie, K.; Tadele, A.; Getahun, D.; et al. Physico-chemical investigation of consumables and environmental samples to determine the causative agent of liver disease outbreak in Tahitay Koraro Woreda, Tiray. Ethiop. Med. J. 2012, 50 (Suppl. 2), 37–45. [Google Scholar] [PubMed]
- Schneider, J.; Tsegaye, Y.; W/Tensae, M.; G/Selassie, S.; Haile, T.; Bane, A.; Ali, A.; Mesfin, G.; Seboxa, T. Veno-occlusive liver disease: A case report. Ethiop Med. J. 2012, 50 (Suppl. 2), 47. [Google Scholar]
- Robinson, O.; Want, E.; Coen, M.; Kennedy, R.; van den Bosch, C.; Gebrehawaria, Y.; Kudo, H.; Sadiq, F.; Goldin, R.D.; Hauser, M.L.; et al. Hirmi Valley liver disease: A disease associated with exposure to pyrrolizidine alkaloids and DDT. J. Hepatol. 2014, 60, 96–102. [Google Scholar] [CrossRef]
- Chiu, C.; Martin, C.; Woldemichael, D.; Selasie, G.W.; Tareke, I.; Luce, R.; G./Libanos, G.; Hunt, D.; Bayleyegn, T.; Addissie, A.; et al. Surveillance of a chronic liver disease of unidentified cause in a rural setting of Ethiopia: A case study. Ethiop. Med. J. 2016, 54, 27–32. [Google Scholar]
- Kisielius, V.; Hama, J.R.; Skrbic, N.; Bruun Hansen, H.C.; Strobel, B.W.; Rasmussen, L.H. The invasive butterbur contaminates stream and seepage water in groundwater wells with toxic pyrrolizidine alkaloids. Sci. Rep. 2020, 10, 19784. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Radwan, A.; Nowak, M. Horizontal natural product transfer: A so far unconsidered source of contamination of plant-derived commodities. J. Environ. Anal. Toxicol. 2015, 5, 215–225. [Google Scholar]
- Nowak, M.; Wittke, C.; Lederer, I.; Klier, B.; Kleinwächter, M.; Selmar, D. Interspecific transfer of pyrrolizidine alkaloids: An unconsidered source of contaminations of phytopharmaceuticals and plant derived commodities. Food Chem. 2016, 213, 163–168. [Google Scholar] [CrossRef]
- Selmar, D.; Abouzeid, S.; Radwan, A.; Hijazin, T.; Yahyazadeh, M.; Lewerenz, L.; Nowak, M.; Kleinwächter, M. Horizontal natural product transfer: A novel attribution in allelopathy. In Co-Evolution of Secondary Metabolites; Merillon, J.M., Ramawat, K., Eds.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Selmar, D.; Wittke, C.; Beck-von Wolffersdorff, I.; Klier, B.; Lewerenz, L.; Kleinwächter, M.; Nowak, M. Transfer of pyrrolizidine alkaloids between living plants: A disregarded source of contaminations. Environ. Pollut. 2019, 248, 456–461. [Google Scholar] [CrossRef]
- Selmar, D.; Radwan, A.; Hijazin, T.; Abouzeid, S.; Yahyazadeh, M.; Lewerenz, L.; Kleinwächter, M.; Nowak, M. Horizontal natural product transfer: Intriguing insights into a newly discovered phenomenon. J. Agric. Food Chem. 2019, 67, 8740–8745. [Google Scholar] [CrossRef]
- Hijazin, T.; Radwan, A.; Lewerenz, L.; Abouzeid, S.; Selmar, D. The uptake of alkaloids by plants from the soil is determined by rhizosphere pH. Rhizophere 2020, 15, 100234. [Google Scholar] [CrossRef]
- Chmit, M.S.; Horn, G.; Dübecke, A.; Beuerle, T. Pyrrolizidine alkaloids in the food chain: Is horizontal transfer of relevance? Foods 2021, 10, 1827. [Google Scholar] [CrossRef] [PubMed]
- Vongdala, N.; Tran, H.D.; Xuan, T.D.; Teschke, R.; Khanh, T.D. Heavy metal accumulation in water, soil, and plants of municipal solid waste landfill in Vientiane, Laos. Int. J. Environ. Res. Public Health 2019, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R.; Eickhoff, A.; Wolff, A.; Xuan, T.D. Liver injury from herbs and “dietary supplements”: Highlights of a literature review from 2015 to Curr. Pharmacol. Rep. 2018, 4, 120–131. [Google Scholar] [CrossRef]
- Philips, C.A.; Paramaguru, R.; Joy, A.K.; Anthony, K.L.; Augistine, P. Clinical outcomes, histopathological patterns, and chemical analysis of Ayurveda and herbal medicine associated with severe liver injury—A single-center experience from southern India. Indian J. Gastroenterol. 2018, 37, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Hamre, H.J.; Glockmann, A.; Heckenbach, K.; Matthes, H. Use and safety of anthroposophic medicinal products: An analysis of 44,662 patients from the EvaMed pharmacovigilance network. Drugs Real World Outcomes 2017, 4, 199–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jong, M.C.; van Wietmarschen, H.; Glockmann, A.; Baars, E.W.; Hamre, H.J. Safety of anthroposophic medicinal products: An analysis of adverse drug reactions from German pharmacovigilance databases. Drugs Real World Outcomes 2021, in press. [Google Scholar] [CrossRef]
- United States Pharmacopeia (USP). USP: 2021 USP Open Forum: Pyrrolizidine Alkaloid Contaminants. Last updated on 9 January 2021. Available online: https://www.usp.org/get-involved/provide-input/stakeholder-forums/2021-pyrrolizidine-alkaloid-contaminants (accessed on 15 June 2021).
- Akhkubekova, A.A.; Tamakhina, A.Y. Accumulation of alkaloids in plants of the family Boraginaceae depending on environmental conditions places of growth. E3S Web Conf. 2021, 254, 02014. [Google Scholar] [CrossRef]
- Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.D.; Khanh, T.D.; Teschke, R. Cordycepin Isolated from Cordyceps militaris: Its newly discovered herbicidal property and potential plant-based novel alternative to Glyphosate. Molecules 2019, 24, 2901. [Google Scholar] [CrossRef] [Green Version]
- Moreira, R.; Pereira, D.M.; Valentão, P.; Andrade, P.B. Pyrrolizidine alkaloids: Chemistry, pharmacology, toxicology and food safety. Int. J. Mol.Sci. 2018, 19, 1668. [Google Scholar] [CrossRef] [Green Version]
- Wink, M. Quinolizidine and pyrrolizidine alkaloid chemical ecology—A mini-review on their similarities and differences. J. Chem. Ecol. 2019, 45, 109–115. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.; González-Agüero, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding stress response. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef] [Green Version]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Mulder, P.P.J.; van der Meijden, E.; Klinkhamer, P.G.L.; Vrieling, K. The correlation between leaf-surface and leaf-tissue secondary metabolites: A case study with pyrrolizidine alkaloids in Jacobaea hybrid plants. Metabolomics 2017, 13, 47. [Google Scholar] [CrossRef]
- Kirk, H.; Vrieling, K.; Van Der Meijden, E.; Klinkhamer, P.G.L. Species by environment interactions affect pyrrolizidine alkaloid expression in Senecio jacobaea, Senecio aquaticus, and their hybrids. J. Chem. Ecol. 2010, 36, 378–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, L.H.; Stegemann, T.; Sievert, C.; Ober, D. Identification of a second site of pyrrolizidine alkaloid biosynthesis in Comfrey to boost plant defense in floral stage. Plant Physiol. 2017, 174, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Vrieling, K.; Klinkhamer, P.G.L. Phytochemical background mediates effects of pyrrolizidine alkaloids on western flower thrips. J. Chem. Ecol. 2019, 45, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Akhi, M.Z.; Haque, M.; Biswas, S. Role of secondary metabolites to attenuate stress damages in plants. In Antioxidants; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar] [CrossRef]
- Teschke, R.; Melchart, D.; Xuan, T.D. Editorial: Hormesis and dose-responses in herbal traditional Chinese medicine (TCM) alone are insufficient solving real clinical TCM challenges and associated herbal quality issues. Longhua Chin. Med. 2018, 19, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Nuringtyas, T.R.; Verpoorte, R.; Klinkhamer, P.G.L.; Hae Choi, Y.; Leiss, K.A. Metabolomic study of diurnal variation on pyrrolizidine alkaloid from Jacobaea sp. hybrids. Metabolomics 2015, 4, 3. [Google Scholar]
- Eller, A.; Chizzola, R. Seasonal variability in pyrrolizidine alkaloids in Senecio inaequidens from the Val Venosta (Northern Italy). Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2016, 150, 1212–1306. [Google Scholar] [CrossRef]
- Hama, J.R.; Bjarne, W.; Strobel, B.J. Occurrence of pyrrolizidine alkaloids in ragwort plants, soils and surface waters at the field scale in grassland. Sci. Total Environ. 2021, 755, 142822. [Google Scholar] [CrossRef] [PubMed]
- Al-Gabbiesh, A.; Kleinwächter, M.; Selmar, D. Influencing the contents of secondary metabolites in spice and medicinal plants by deliberately applying drought stress during their cultivation. Jordan J. Biol. Sci. 2015, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Skoneczny, D.; Zhu, X.; Weston, P.A.; Gurr, G.M.; Callaway, R.M.; Weston, L.A. Production of pyrrolizidine alkaloids and shikonins in Echium plantagineum L. in response to various plant stressors. Pest Manag. Sci. 2019, 75, 530–2541. [Google Scholar] [CrossRef] [PubMed]
- Hol, W.H.G. The effect of nutrients on pyrrolizidine alkaloids in Senecio plants and their interactions with herbivores and pathogens. Phytochem. Rev. 2011, 10, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, T.; Ehmke, A.; Eilert, U.; von Borstel, C.; Theuring, C. Sites of synthesis, translocation and accumulation of pyrrolizidine alkaloid N-oxides in Senecio vulgaris L. Planta 1989, 177, 98–107. [Google Scholar] [CrossRef]
- Kruse, L.H.; Stegemann, T.; Jensen-Kroll, J.; Engelhardt, A.; Wesseling, A.M.; Lippert, A.; Ludwig-Müller, J.; Ober, D. Reduction of pyrrolizidine alkaloid levels in comfrey (Symphytum officinale) hairy roots by RNAi silencing of homospermidine synthase. Planta Med. 2019, 85, 1177–1186. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Ma, J.; Ruan, J.; Ye, Y.; Fu, P.P.C.; Lin, G. Intestinal and hepatic biotransformation of pyrrolizidine alkaloid N-oxides to toxic pyrrolizidine alkaloids. Arch. Toxicol. 2019, 93, 2197–2209. [Google Scholar] [CrossRef]
- He, Y.; Zhu, L.; Ma, J.; Lin, G. Metabolism-mediated cytotoxicity and genotoxicity of pyrrolizidine alkaloids. Arch. Toxicol. 2021, 95, 1917–1942. [Google Scholar] [CrossRef]
- Wei, X.; Ruan, W.; Vrieling, K. Current knowledge and perspectives of pyrrolizidine alkaloids in pharmacological applications: A mini-review. Molecules 2021, 26, 1970. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.P.; Xia, Q.; Lin, G.; Chou, M.W. Pyrrolizidine alkaloids—Genotoxicity, metabolism enzymes, metabolic activation, and mechanisms. Drug Metab. Rev. 2004, 36, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Abegaz, B.M.; Kinfe, H.H. Secondary metabolites, their structural diversity, bioactivity, and ecological functions: An overview. Phys. Sci. Rev. 2019, 4, 20180100. [Google Scholar] [CrossRef]
- Rösemann, G.M.; Botha, C.J.; Eloff, J.N. Distinguishing between toxic and non-toxic pyrrolizidine alkaloids and quantification by liquid chromatography–mass spectrometry. Phytochem. Lett. 2014, 8, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R.; Zhu, Y. Opinion: Intestinal microbiome, endotoxins, cytochrome P450 2E1, and the gut-liver axis in alcoholic liver disease. EC Gastroenterology Dig. Syst. 2019, 5, 11. [Google Scholar]
- Fashe, M.M.; Juvonen, R.; Petsalo, A.; Vepsäläinen, J.; Pasanen, M.; Rahnasto-Rilla, M. In silico prediction of the site of oxidation by cytochrome P450 3A4 that leads to the formation of the toxic metabolites of pyrrolizidine alkaloids. Chem. Res. Toxicol. 2015, 28, 702–710. [Google Scholar] [CrossRef]
- Guengerich, F.P. Mechanisms of cytochrome P450 substrate oxidation: MiniReview. J. Biochem. Molec. Toxicol. 2007, 21, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Johansson, I.; Ingelman-Sundberg, M. Genetic polymorphism and toxicology—with emphasis on cytochrome P450. Toxicol. Sci. 2011, 120, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Uetrecht, J. Mechanistic studies of idiosyncratic DILI: Clinical implications. In Special issue: Clinical drug induced liver injury: Current diagnostic and mechanistic challenges; Rolf Teschke, Gaby Danan & James, H. Lewis. Front. Pharmacol. 2019, 10, 837. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R.; Uetrecht, J. Idiosyncratic drug induced liver injury (DILI): Unresolved basic issues. In special issue: Unresolved basic issues in hepatology, guest editors Ralf Weiskirchen & Wolfgang Stremmel. Ann. Transl. Med. 2021, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Eickhoff, A.; Brown, A.C.; Neuman, M.G.; Schulze, J. Diagnostic biomarkers in liver injury by drugs, herbs, and alcohol: Tricky dilemma after EMA correctly and officially retracted Letter of Support. Int. J. Mol. Sci. 2020, 21, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teschke, R.; Danan, G. Liver injury by drugs metabolized via cytochrome PJ. Mod. Med. Chem. 2020, 8, 93–98. [Google Scholar] [CrossRef]
- Falero-Perez, J.; Song, Y.-S.; Zhao, Y.; Teixeira, L.; Sorenson, C.M.; Sheibani, N. Cyp1b1 expression impacts the angiogenic and inflammatory properties of liver sinusoidal endothelial cells. PLoS ONE 2018, 13, e0206756. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sangwung, P.; Kondo, R.; Jung, Y.; McConnell, M.J.; Jeong, J.; Utsumi, T.; Sessa, W.C.; Iwakiri, Y. Alcohol-induced Hsp90 acetylation is a novel driver of liver sinusoidal endothelial dysfunction and alcohol-related liver disease. J. Hepatol. 2021, 75, 377–386. [Google Scholar] [CrossRef]
- Lu, Y.; Wong, K.Y.; Tan, C.; Ma, J.; Feng, B.; Lin, G. Establishment of a novel CYP3A4-transduced human hepatic sinusoidal endothelial cell model and its application in screening hepatotoxicity of pyrrolizidine alkaloids. J. Environ. Sci. Health Part C 2020, 38, 169–185. [Google Scholar] [CrossRef]
- Lu, A.Y.; Kuntzman, R.; West, S.; Jacobson, M.; Conney, A.H. Reconstituted liver microsomal enzyme system that hydroxylates drugs, other foreign compounds, and endogenous substrates: II Role of the cytochrome P-450 and P-448 fractions in drug and steroid hydroxylations. J. Biol. Chem. 1972, 247, 1727–1734. [Google Scholar] [CrossRef]
- Ohnishi, K.; Lieber, C.S. Reconstitution of the microsomal ethanol-oxidizing system. Qualitative and quantitative changes of cytochrome P-450 after chronic ethanol consumption. J. Biol. Chem. 1977, 52, 7124–7131. [Google Scholar] [CrossRef]
- Teschke, R.; Ohnishi, K.; Hasumura, Y.; Lieber, C.S. Hepatic microsomal ethanol oxidizing system: Isolation and reconstitution. In Microsomes and Drug Oxidations; Ullrich, V., Roots, I., Hildebrandt, A., Estabrook, R.W., Conney, A.H., Eds.; Pergamon Press: Oxford, UK, 1977; pp. 103–110. [Google Scholar]
- Guengerich, F.P. Cytochrome P450 research and The Journal of Biological Chemistry. J. Biol. Chem. 2019, 294, 1671–1680. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.; Stevens, K. Pyrrolizidine alkaloids: Occurence, biology, and chemical synthesis. Nat. Prod. Rep. 2017, 34, 62–89. [Google Scholar] [CrossRef]
- He, Y.; Lian, W.; Ding, L.; Fan, X.; Ma, J.; Zhang, Q.Y.; Ding, X.; Lin, G. Lung injury induced by pyrrolizidine alkaloids depends on metabolism by hepatic cytochrome P450s and blood transport of reactive metabolites. Arch. Toxicol. 2021, 95, 103–116. [Google Scholar] [CrossRef]
- Edgar, J.A.; Molyneux, R.J.; Colegate, S.M. Pyrrolizidine alkaloids: Potential role in the etiology of cancers, pulmonary hypertension, congenital anomalies, and liver disease. Chem. Res. Toxicol. 2015, 28, 4–20. [Google Scholar] [CrossRef]
- Chow, H.C.; So, T.H.; Choi, H.C.W.; Lam, K.O. Literature review of Traditional Chinese Medicine herbs-induced liver injury from an oncological perspective with RUCAM. Integr. Cancer Ther. 2019, 18, 1534735419869479. [Google Scholar] [CrossRef]
- Teschke, R.; Eickhoff, A.; Schulze, J.; Danan, G. Herb-induced liver injury (HILI) with 12,068 worldwide cases published with causality assessments by Roussel Uclaf Causality Assessment Method (RUCAM): An overview. Transl. Gastroenterol. Hepatol. 2021, 6, 51. [Google Scholar] [CrossRef]
- Teschke, R.; Eickhoff, A. Herbal hepatotoxicity in traditional and modern medicine: Actual key issues and new encouraging steps. Front. Pharmacol. 2015, 6, 72. [Google Scholar] [CrossRef] [Green Version]
- Frenzel, C.; Teschke, R. Herbal hepatotoxicity: Clinical characteristics and listing compilation. Int. J. Mol. Sci. 2016, 17, 588. [Google Scholar] [CrossRef] [PubMed]
- DeLeve, L.D.; Shulman, H.M.; Mcdonald, G.B. Toxic injury to hepatic sinusoids: Sinusoidal obstruction syndrome (veno-occlusive disease). Semin. Liver Dis. 2002, 22, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, Y.; Liu, Y.; Xie, W.; Zou, X.; Xu, J.; Wang, J. Chinese Society of Gastroenterology Committee of Hepatobiliary Disease. Expert consensus on the clinical management of pyrrolizidine alkaloid-induced hepatic sinusoidal obstruction syndrome. J. Gastroenterol. Hepatol. 2019, 34, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Danan, G.; Teschke, R. RUCAM in drug and herb induced liver injury: The update. In Special Issue “Drug, Herb, and Dietary Supplement Hepatotoxicity”, Teschke, R., Raúl, J. Andrade. Int. J. Mol. Sci. 2016, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, L.; Zhang, M.; Zhang, F.; Peng, C.; Zhang, B.; Chen, J.; Li, L.; He, J.; Xiao, J.; et al. Validation of the Nanjing criteria for diagnosing pyrrolizidine alkaloids-induced hepatic sinusoidal obstruction syndrome. J. Clin. Transl. Hepatol. 2021, 9, 345–352. [Google Scholar] [CrossRef]
- Wang, J.B.; Zhu, Y.; Bai, Z.F.; Wang, F.S.; Li, X.H.; Xiao, X.H.; on behalf of The Branch Committee for Hepatobiliary Diseases and Branch Committee for Chinese Patent Medicines, China Association of Chinese Medicine. Guidelines for the Diagnosis and Management of Herb-Induced Liver Injury. Chin. J. Integr. Med. 2018, 24, 696–706. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, Y.; Yang, X.; Xion, A.Z.; Wang, Z.T.; Yang, L. Gynura rhizoma containing pyrrolizidine alkaloids induces the hepatic sinusoidal obstruction syndrome in mice via upregulating fibrosis-related factors. Acta Pharmacol. Sin. 2019, 40, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.C.; Li, Y.; Chang, B. Research advances in the pathogenesis of pyrrolizidine alkaloid-induced hepatic sinusoidal obstruction syndrome. J. Clin. Hepatol. 2021, 37, 962–964. [Google Scholar] [CrossRef]
- Schrenk, D.; Gao, L.; Lin, G.; Mahony, C.; Mulder, P.P.J.; Peijnenburg, A.; Pfuhler, S.; Rietjens, I.M.C.M.; Rutz, L.; Steinhoff, B.; et al. Pyrrolizidine alkaloids in food and phytomedicine: Occurrence, exposure, toxicity, mechanisms, and risk assessment—A review. Food Chem. Toxicol. 2020, 136, 111107. [Google Scholar] [CrossRef] [PubMed]
- Bolt, H.M. Hepatotoxicity of pyrrolizidine alkaloids in rats in relation to human exposure. Arch. Toxicol. 2020, 94, 2885–2886. [Google Scholar] [CrossRef]
- Wu, G.L.; Yu, G.Y.; Chen, J. Clinical analysis of hepatic veno-occlusive disease induced by Sedum aizoon. Zhongguo Zhong Yao Za Zhi 2008, 33, 2402–2404. (in Chinese). [Google Scholar]
- Lin, G.; Wang, J.Y.; Li, N.; Li, M.; Gao, H.; Ji, Y.; Zhang, F.; Wang, H.; Zhou, Y.; Ye, Y.; et al. Hepatic sinusoidal obstruction syndrome associated with consumption of Gynura segetum. J. Hepatol. 2011, 54, 666–673. [Google Scholar] [CrossRef]
- Gao, X.S.; Xiao, S.S.; He, J.F. Analysis of alkaloids in Sedum aizoon and establishment of hepatic venoocclusive model in mice. Chin. J. Integr. Trad. West. Elect. Microsc. Med. Digest. 2006, 14, 311–313. [Google Scholar]
- Gao, H.; Li, N.; Wang, J.Y.; Zhang, S.C.; Lin, G. Definitive diagnosis of hepatic sinusoidal obstruction syndrome induced by pyrrolizidine alkaloids. J. Dig. Dis. 2012, 13, 33–39. [Google Scholar] [CrossRef]
- Wang, J.Y.; Gao, H. Tusanqi and hepatic sinusoidal obstruction syndrome. J. Dig. Dis. 2014, 15, 105–107. [Google Scholar] [CrossRef]
- Dai, H.F.; Gao, Y.; Yang, M.; Yu, C.H.; Gu, C.Y.; Chen, W.X. Hepatic venoocclusive disease induced by Gynura segetum: Report of two cases. Hepatobiliary Pancreat. Dis. Int. 2006, 5, 406–408. [Google Scholar] [PubMed]
- Kumana, C.R.; Ng, M.; Lin, H.J.; Ko, W.; Wu, P.C.; Todd, D. Hepatic veno-occlusive disease due to toxic alkaloid in herbal tea. Lancet 1983, 322, 1360–1361. [Google Scholar] [CrossRef]
- Kumana, C.R.; Ng, M.; Lin, H.J.; Ko, W.; Wu, P.C.; Todd, D. Herbal tea induced hepatic veno-occlusive disease: Quantification of toxic alkaloid exposure in adults. Gut 1985, 26, 101–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culvenor, C.C.J.; Edgar, J.A.; Smith, L.W.; Kumana, C.R.; Lin, H.J. Heliotropium lasiocarpum Fisch and Mey identified as cause of veno-occlusive disease due to herbal tea. Lancet 1986, 1, 978. [Google Scholar] [CrossRef]
- Gao, H.; Ruan, J.Q.; Chen, J.; Li, N.; Ke, C.Q.; Ye, Y.; Lin, G.; Wang, J.Y. Blood pyrrole-protein adducts as a diagnostic and prognostic index in pyrrolizidine alkaloid-hepatic sinusoidal obstruction syndrome. Drug Des. Dev. Ther. 2015, 9, 4861–4868. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Shi, M.; Wu, X.; Ma, J.; Ng, K.T.P.; Xia, Q.; Zhu, L.; Fu, P.P.C.; Man, K.; Tsui, S.K.W.; et al. Mutational signature analysis reveals widespread contribution of pyrrolizidine alkaloid exposure to human liver cancer. Hepatology 2021, 74, 264–280. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Gao, H.; Li, N.; Xue, J.; Chen, J.; Ke, C.; Ye, Y.; Fu, P.P.; Zheng, J.; Wang, J.; et al. Blood pyrrole-protein adductsa—Of pyrrolizidine alkaloid-induced liver injury in humans. J. Environ. Sci. Health Part C 2015, 33, 404–421. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, W.; He, Y.; Zhu, L.; Zhang, C.; Liu, J.; Ye, Y.; Zhuge, Y.; Lin, G. Clinical application of pyrrole–hemoglobin adducts as a biomarker of pyrrolizidine alkaloid exposure in humans. Arch. Toxicol. 2021, 95, 759–765. [Google Scholar] [CrossRef]
- Ma, J.; Xia, Q.; Fu, P.P.; Lin, G. Pyrrole-protein adducts—A biomarker of pyrrolizidine alkaloid-induced hepatotoxicity. J. Food Drug Anal. 2018, 26, 965–972. [Google Scholar] [CrossRef]
- Yang, M.; Ruan, J.; Gao, H.; Li, N.; Ma, J.; Xue, J.; Ye, Y.; Fu, P.P.; Wang, J.; Lin, G. First evidence of pyrrolizidine alkaloid N-oxide-induced hepatic sinusoidal obstruction syndrome in humans. Arch. Toxicol. 2017, 91, 3913–3925. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Yang, Y.; Chen, Y.; Zhuge, Y.; Xiong, A.; Yang, L.; Wang, Z. Blood microRNA signatures serve as potential diagnostic biomarkers for hepatic sinusoidal obstruction syndrome caused by Gynura japonica containing pyrrolizidine alkaloids. Front. Pharmacol. 2021, 12, 627126. [Google Scholar] [CrossRef]
- Chen, Y.; Xiong, F.; Wang, W.; Jiang, K.; Ye, X.; Deng, G.; Wang, C.; Yang, L.; Xiong, A.; Wang, Z. The long persistence of pyrrolizidine alkaloid-derived pyrrole-protein adducts in vivo: Kinetic study following multiple exposures of a pyrrolizidine alkaloid containing extract of Gynura japonica. Toxicol. Lett. 2020, 323, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Larrey, D.; Melchart, D.; Danan, G. Traditional Chinese Medicine (TCM) and herbal hepatotoxicity: RUCAM and the role of novel diagnostic biomarkers such as microRNAs. Medicines 2016, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Meunier, L.; Larrey, D. DILI: Biomarkers, requirements, candidates and validation. Front. Pharmacol. 2019, 10, 1482. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ruan, J.; Chen, X.; Li, D.; Yao, S.; Fu, P.P.; Ye, Y.; Gao, H.; Wang, J.; Lin, G. Pyrrole-hemoglobin adducts, a more feasible potential biomarker of pyrrolizidine alkaloid exposure. Chem. Res. Toxicol. 2019, 32, 1027–1039. [Google Scholar] [CrossRef]
- Zhuge, Y.Z.; Wang, Y.; Zhang, F.; Zhu, C.K.; Zhang, W.; Zhang, M.; He, Q.; Yang, J.; He, J.; Chen, J.; et al. Clinical characteristics and treatment of pyrrolizidine alkaloid-related hepatic vein occlusive disease. Liver Int. 2018, 38, 1867–1874. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Rong, X.; Guo, H.; Xu, D.; Liu, C.; Meng, L.; Yang, X.; Guo, T.; Kan, X.; Song, Y. Clinical characteristics, CT signs, and pathological findings of pyrrolizidine alkaloids-induced sinusoidal obstructive syndrome: A retrospective study. BMC Gastroenterol. 2020, 20, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, X.; Xie, W.; Ren, X.; Zhang, M.; Xu, J. Quantitative analysis of CT images in patients with pyrrolizidine alkaloid-induced sinusoidal obstruction syndrome. Sci. Rep. 2019, 9, 2179. [Google Scholar] [CrossRef]
- Teschke, R.; Danan, G. Worldwide use of RUCAM for causality assessment in 81,856 DILI and 14,029 HILI cases published 1993—Mid 2020: A comprehensive analysis. Medicines 2020, 7, 62. [Google Scholar] [CrossRef]
- DeLeve, L.D.; McCuskey, R.S.; Wang, X.; Hu, L.; McCuskey, M.K.; Epstein, R.B.; Kanel, G.C. Characterization of a reproducible rat model of hepatic veno-occlusive disease. Hepatology 1999, 29, 1779–1791. [Google Scholar] [CrossRef]
- Kumar, A.; Palek, R.; Liska, V.A. Critical analysis of experimental animal models of sinusoidal obstruction syndrome. J. Clin. Exp. Hepatol. 2019, 9, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, S.E.H.; Miller, C.G.; McKenzie, K.A.W.; Audretsch, J.J.; Bras, G. Acute veno-occlusive disease of the liver: Fine structure in Jamaican children. Arch. Pathol. 1970, 89, 507–520. [Google Scholar] [PubMed]
- Danan, G.; Bénichou, C. Causality assessment of adverse reactions to drugs—I. A novel method based on the conclusions of international consensus meetings: Application to drug-induced liver injuries. J. Clin. Epidemiol. 1993, 46, 1323–1330. [Google Scholar] [CrossRef]
- Benichou, C.; Danan, G.; Flahault, A. Causality assessment of adverse reactions of drugs—II. An original model for validation of drug causality assessment methods: Case reports with positive rechallenge. J. Clin. Epidemiol. 1993, 46, 1331–1336. [Google Scholar] [CrossRef]
- Teschke, R. Editorial. DILI, HILI, RUCAM algorithm, and AI, the Artificial Intelligence: Provocative issues, progress, and proposals. Arch. Gastroenterol. Res. 2020, 1, 4–11. [Google Scholar]
- Peng, C.; Zhang, X.; Zhang, F.; Liu, L.; Shao, Y.; Xiang, X.; Zhang, W.; Xiao, J.; Wang, Y.; Zhang, B.; et al. Clinical efficacy and safety of anticoagulation therapy for Pyrrolizidine alkaloids-induced hepatic sinusoidal obstruction syndrome: A retrospective multicenter cohort study. Eur. J. Gastroenterol. Hepatol. 2020, 32, 1168–1178. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, L.; Du, X.; Hao, M.; Gao, P. Low molecular weight heparin in the treatment of pyrrolizidine alkaloid-induced hepatic sinusoidal obstruction syndrome: Five case reports. J. Int. Med. Res. 2020, 48, 300060520961916. [Google Scholar] [CrossRef]
- Shang, H.; Bai, T.; Zhu, S.; Yang, X.; Liu, C.; Xu, D.; Zhuge, Y.; Song, Y.; Xiaohua Hou, X. Prognostic factors for pyrrolizidine alkaloid-induced hepatic sinusoidal obstruction syndrome: A multicenter study in China. Ann. Transl. Med. 2021, 9, 1–11. [Google Scholar] [CrossRef]
- Habs, M.; Binder, K.; Krauss, S.; Müller, K.; Ernst, B.; Valentini, L.; Koller, M. A balanced risk–benefit analysis to determine human risks associated with pyrrolizidine alkaloids (PA)—The case of tea and herbal infusions. Nutrients 2017, 9, 717. [Google Scholar] [CrossRef] [Green Version]
- Rasenack, R.; Müller, C.; Kleinschmidt, M.; Rasenack, J.; Wiedenfeld, H. Veno-occlusive disease in a fetus caused by pyrrolizidine alkaloids of food origin. Fetal Diagn. Ther. 2003, 18, 223–225. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, L.Y.; Zhu, Z.J.; Wei, L.; Qu, W.; Zeng, Z.G. Is sinusoidal obstructive syndrome a recurrent disease after liver transplantation? A case report. World J. Clin. Cases 2021, 9, 489–495. [Google Scholar] [CrossRef]
- Barcelos, S.T.A.; Dall’Oglio, V.M.; de Araújo, A.; Cerski, C.T.S.; Álvares-da-Silva, M.R. Sinusoidal obstruction syndrome secondary the intake of Senecio brasiliensis: A case report. Ann. Hepatol. 2021, 20, 100138. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, Z.; Wang, F.; Shi, L. Letter to Editor. Hepatic sinusoidal obstruction syndrome after oral intake of Gynura segetum. Asian J. Surg. 2021, 44, 991–993. [Google Scholar] [CrossRef]
- Sun, Z.; Kang, J.; Zhang, Y. Hepatic veno-occlusive disease related to Gynura segetum. Medicine 2018, 97, e0552. [Google Scholar] [CrossRef] [PubMed]
- Neuman, M.G.; Cohen, L.B.; Steenkamp, V. Pyrrolizidine alkaloids enhance alcohol-induced hepatocytotoxicity in vitro in normal human hepatocytes. Eur. Rev. Med. Pharmacol. Sci. 2017, 21 (Suppl. 1), 53–68. [Google Scholar] [PubMed]
- Cen, P.; Ding, J.; Jin, J. Hepatic sinusoidal obstruction syndrome caused by the ingestion of Gynura segetum in a patient with alcoholic cirrhosis: A case report. J. Int. Med. Res. 2021, 49, 300060520980649. [Google Scholar] [CrossRef]
- Ya, L.; Yulin, M.; Liang, L.; Jing, W. Hepatic veno-occlusive disease caused by Gynura segetum: A case report. Cardiol. Cardiovasc. Med. 2019, 3, 9–14. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Zhang, W.; Ma, J.; Xia, Q.; Song, Z.; Zhu, L.; Zhang, C.; Liu, J.; Ye, Y.; Fu, P.P.; et al. Blood pyrrole–DNA adducts define the early tumorigenic risk in patients with pyrrolizidine alkaloid-induced liver injury. Environ. Sci. Technol. Lett. 2021, 8, 7. [Google Scholar] [CrossRef]
- Anderson, N.; Borlak, J. Hepatobiliary events in migraine therapy with herbs—The case of Petadolex, a Petasites hybridus extract. J. Clin. Med. 2019, 8, 652. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R.; Eickhoff, A.; Schulze, J.; Wolff, A.; Frenzel, C.; Melchart, D. Petadolex®, a herbal extract for migraine prophylaxis with spontaneous case reports of disputed liver injury: Robust causality evaluation by RUCAM, the Roussel Uclaf Causality Assessment Method. Eur. J. Pharmaceut. Med. Res. 2016, 3, 154–177. [Google Scholar]
- Diener, H.C.; FG Freitag, F.G.; Danesch, U. Safety profile of a special butterbur extract from Petasites hybridus in migraine prevention with emphasis on the liver. Cephalgia Rep. 2018, 1, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mohabbat, O.; Younos, M.S.; Merzad, A.A.; Srivastava, R.N.; Sediq, G.G.; Aram, G.N. An outbreak of hepatic veno-occlusive disease in north-western Afghanistan. Lancet 1976, 308, 269–271. [Google Scholar] [CrossRef]
- Kakar, F.; Akbarian, Z.; Leslie, T.; Mustafa, M.L.; Watson, J.; van Egmond, H.P.; Omar, M.F.; Mofleh, J. An outbreak of hepatic veno-occlusive disease in western Afghanistan associated with exposure to wheat flour contaminated with pyrrolizidine alkaloids. J. Toxicol. 2010, 313280. [Google Scholar] [CrossRef]
- Tandon, R.K.; Tandon, B.N.; Tandon, H.D. Study of an epidemic of venoocclusive disease in India. Gut 1976, 17, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Tandon, B.N.; Tandon, H.D.; Tandon, R.K.; Narndranathan, M.; Joshi, Y.K. An epidemic of veno-occlusive disease of the liver in central India. Lancet 1976, 308, 272. [Google Scholar] [CrossRef]
- Chauvin, P.; Dillon, J.C.; Moren, A. Épidémie d’intoxication alimentaire á l’héliotrope, tadjikistan, Novembre 1992–Mars 1993. Cah. Études Rech. Francoph. Santé 1994, 4, 263–268. [Google Scholar]
- Tandon, H.D.; Tandon, B.N.; Mattocks, A.R. An epidemic of veno-occlusive disease of the liver in Afghanistan pathologic features. Am. J. Gastroenterol. 1978, 70, 607–613. [Google Scholar] [PubMed]
- Altaee, M.Y. An outbreak of veno-occlusive disease of the liver in northern Iraq. East. Mediterr. Health J. 1998, 4, 142–148. Available online: https://apps.who.int/iris/handle/10665/117894 (accessed on 8 July 2021). [CrossRef]
- Auerbach, B.J.; Reynolds, S.; Lamorde, M.; Merry, C.; Kukunda-Byobona, C.; Ocama, P.; Semeere, A.; Ndyanabo, A.; Boaz, I.; Kiggundu, V.; et al. Traditional herbal medicine use associated with liver fibrosis in Rural Rakai, Uganda. PLoS ONE 2012, 7, e41737. [Google Scholar] [CrossRef] [Green Version]
- Tobyn, G.; Denham, A.; Whitelegg, M. Tussilago farfara, coltsfoot. In Medical Herbs; Elsevier: Amsterdam, The Netherlands, 2011; pp. 317–326. [Google Scholar]
- Wang, D.; Huang, L.; Chen, S. Senecio scandens Buch.-Ham.: A review on its ethnopharmacology, phytochemistry, pharmacology, and toxicity. J. Ethnopharmacol. 2013, 149, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.H.; McSorley, R. Management of nematodes and soil fertility with Sunn hemp cover crop. EDIS 1969, 18, 2004. [Google Scholar] [CrossRef]
- Carvalho, S.; Macel, M.; Mulder, P.P.J.; Skidmore, A.; van der Putten, W.H. Chemical variation in Jacobaea vulgaris is influenced by the interaction of season and vegetation successional stage. Phytochemistry 2014, 99, 86–94. [Google Scholar] [CrossRef]
- Hoi, H.T. Some kinds of medicinal herbs for the treatment of sinusitis. Biomed. Pharm. J. 2020, 13, 927–932. [Google Scholar] [CrossRef]
- Aydın, A.A.; Zerbes, V.; Parlar, H.; Letzel, T. The medical plant butterbur (Petasites): Analytical and physiological (re)view. J. Pharm. Biomed. Anal. 2013, 75, 220–229. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feed Products | Mean PA Amounts (µg/kg) | ||
|---|---|---|---|
| Lower Bound | Middle Bound | Upper Bound | |
| Wheat | 23 | 171 | 320 |
| Maize | 0 | 151 | 302 |
| Millet | 0 | 151 | 302 |
| Oats | 0 | 151 | 302 |
| Rice, broken | 0 | 151 | 302 |
| Sorghum, Milo | 0 | 151 | 302 |
| Palm kemel expeller | 0 | 151 | 302 |
| Rape seed | 9 | 159 | 308 |
| Toasted soya beans | 3 | 153 | 303 |
| Sunflower seeds | 5 | 155 | 305 |
| Linseed | 30 | 177 | 325 |
| Peas | 16 | 166 | 315 |
| Carob, dried | 8 | 156 | 305 |
| Sweet lupines | 0 | 151 | 302 |
| Carrots | 0 | 151 | 302 |
| Citrus pulp | 12 | 161 | 311 |
| Lucerne (alfalfa) | 368 | 503 | 637 |
| Grass, field dried, hay | 174 | 322 | 470 |
| Plant Products for Human Use | Mean PA Amounts (µg/kg) | ||
|---|---|---|---|
| Lower Bound | Middle Bound | Upper Bound | |
| Herbal mix | 353 | 492 | 630 |
| Artichoke | 2252 | 2385 | 2517 |
| Camomile | 35 | 184 | 334 |
| Dandelion | 663 | 793 | 924 |
| Fennel | 1592 | 1732 | 1871 |
| Ginseng | 5 | 154 | 302 |
| Goldenrod | 18 | 165 | 312 |
| Knotweed | 97 | 241 | 385 |
| Leek | 0 | 151 | 302 |
| Mangold | 0 | 151 | 302 |
| Milk thistle | 12 | 161 | 309 |
| Mint | 0 | 151 | 302 |
| Nettle | 16 | 165 | 314 |
| Oregano | 89 | 235 | 381 |
| Parsley | 0 | 151 | 302 |
| Rose tip | 0 | 151 | 302 |
| Rosemary | 5 | 154 | 302 |
| Verbena | 18 | 164 | 310 |
| Plant (Family) | Compounds of Pyrrolizidine Alkaloids (Selected) |
|---|---|
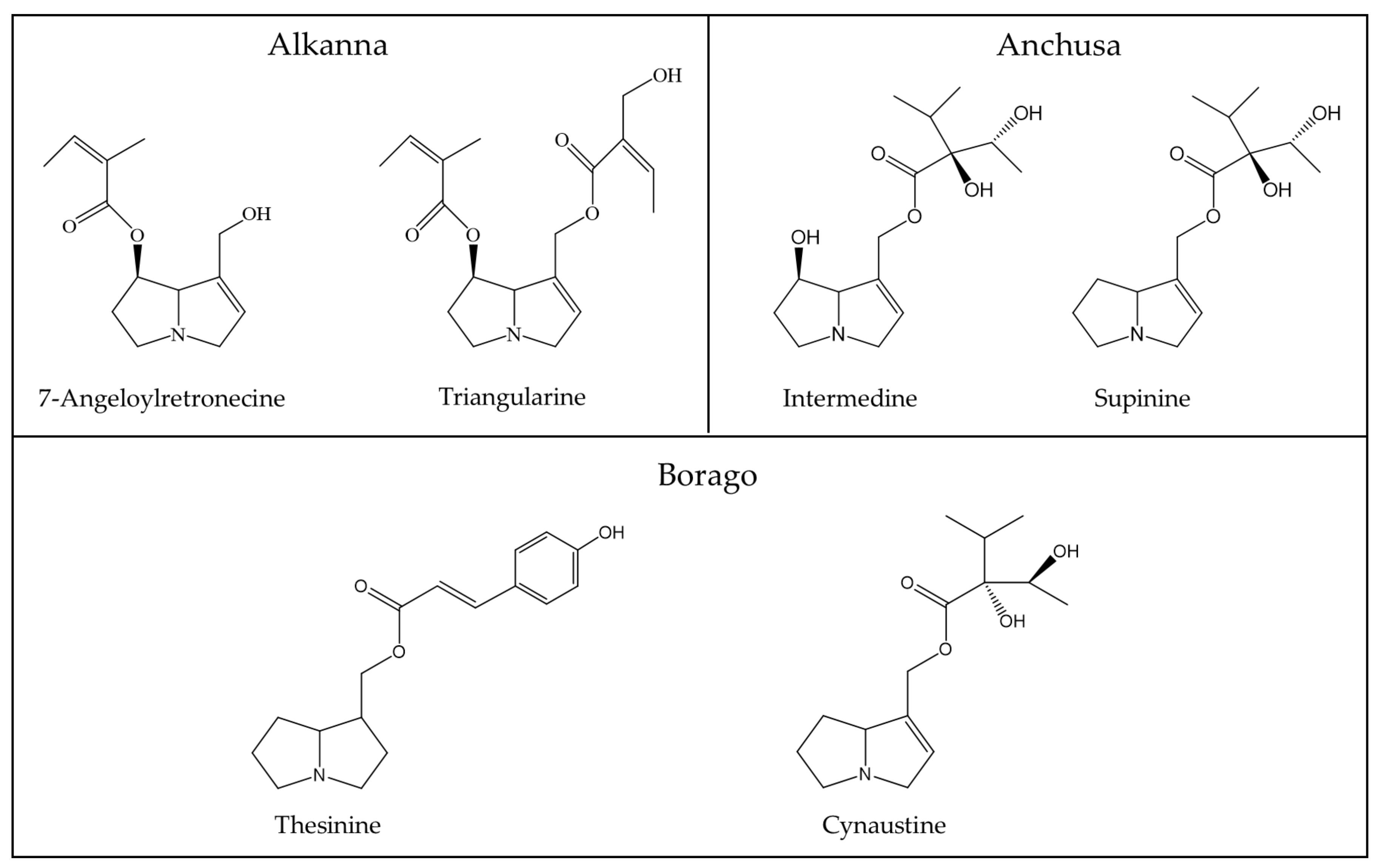
| Alkanna (Boraginaceae) | 7-Angeloylretronecine [57,58], 9-Angeloylretronecine [57], 7-Tigloylretronecine [57], 7-Senecioylretronecine [57], 7-Tigloylretronecine [57], 9-Senecioylretronecine [57], 7-Angeloyol-9-(hydroxypropenoyl) retronecine [57], 7-Tigloyl-9-(hydroxy propenoyl) retronecine [57], 7-Angeloyol-9-(2,3-dihydroxypropanoyl) retronecine [57], 7-Tigloyl-9-(2,3-dihydroxypropanoyl) retronecine [57], Triangularine [57,58], Triangularicine [57], Dihydroxytrian gularicine [57,58] |
| Anchusa (Boraginaceae) | Anthamidin [57], Supinine [57,59], Intermedin [57,59], Lycopsamine [57,59], Currassavine [59] |
| Borago (Boraginaceae) | Lycopsamine [60], Supinidine [60], Viridiflorate [60], Cynaustine [60], Amabaline [60,61], Thesinine [61,62] |
| Brachyglottis (Asteraceae) | Senecionine [63,64], Retrorsine [63,64], Integerrimine, Senkirkine [63], Hectorine [63,64], Petasinine [63,64] |
| Cineraria (Asteraceae) | Otosenine [65,66], Florosenine [65,66], Floridanine [65,66], Doronine [65], Senecionine [66,67], Integerrimine [66,67], Seneciphylline [66,67] |
| Crotalaria (Leguminosae) | Tashiromine [68], Retronecine [68], Heliotridine [68], 9-Isosenecioylretronecine [68], 9-Angeloylretronecine [68], 7-Seneciolylretronecine [68], 9-Hydroxyheptanoylretronecine [68], 9-Hydroxyisohexenoylretronecine [68], Rinderine [68], 9-Hydroxytigloylretronecine [68], Monocrotaline [68], Dihydrosenecionine isomer [68] |
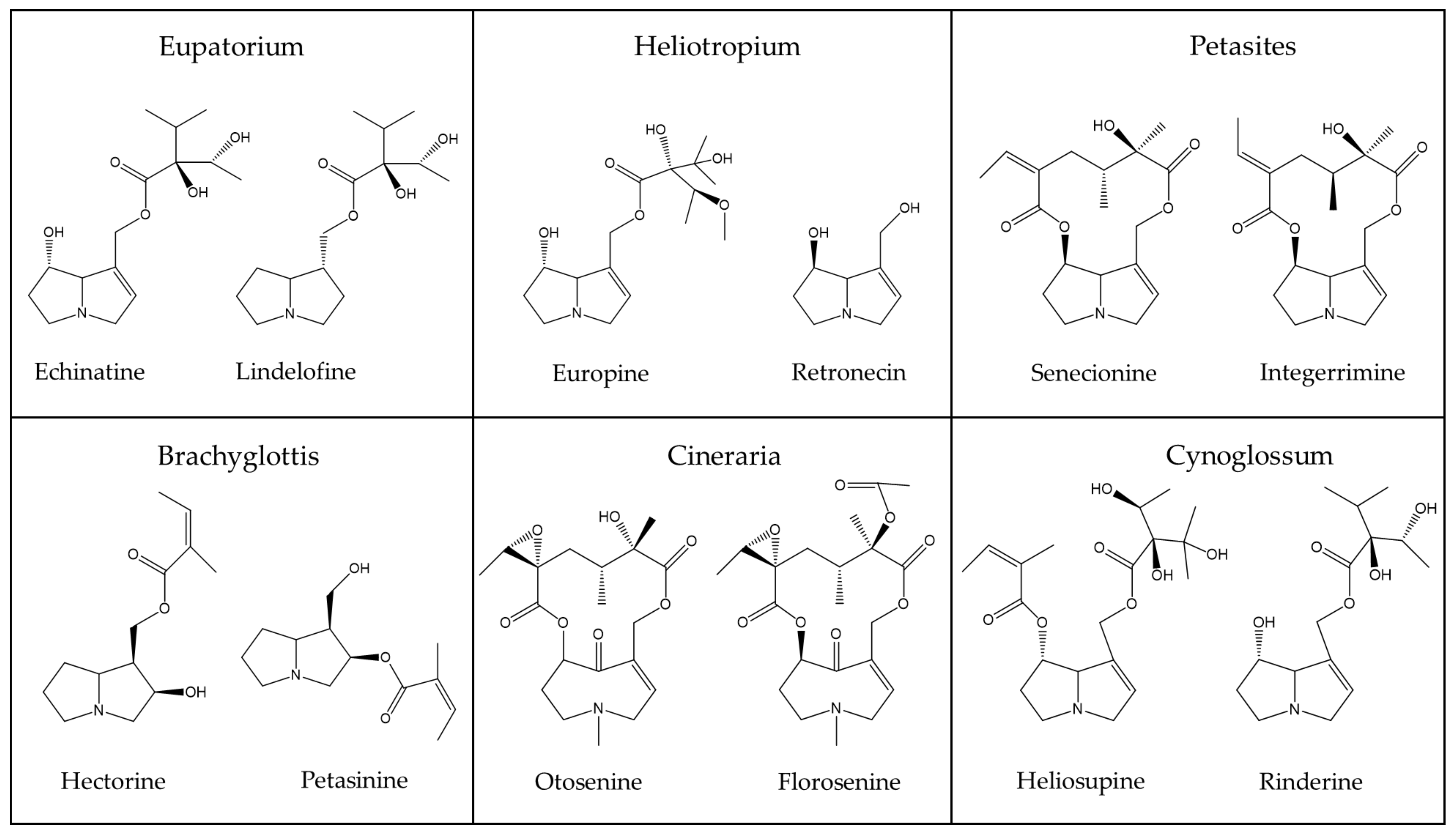
| Cynoglossum (Boraginaceae) | Heliosupine [69], Rinderine [69], Echinatine [69], Viridiflorine [69] |
| Eupatorium (Asteraceae) | Lindelofine [70], Supinine [70,71,72,73], Lycopsamine [73,74,75], Intermedin [73,74,75], Amabaline [71,76], Echinatine [71,76], Rinderine [71], Viridiflorine [71], Cynaustraline [71], Tussilagine [76] |
| Foeniculum vulgare (Apiaceae) | PA compounds not individually specified [77] |
| Heliotropium (Boraginaceae) | Trachelanthamine [78], Floridine [78], Heliovicine [78], Lycopsamine [79,80], Amabiline [79], Curassavine [79,81], Heliospathine [79], Intermedin [80], Europine [82,83,84,85], Liamin [82], Heliotrine [84,85,86], Lasiocarpine [84], Retronecine [80,87,88,89], Helibracteatine [87], Helifoline [88], Heliscabine [89], Heliosupine [86], Echinatine [86], Supinine [85], Heleurine [85], Coromandaline [81] |
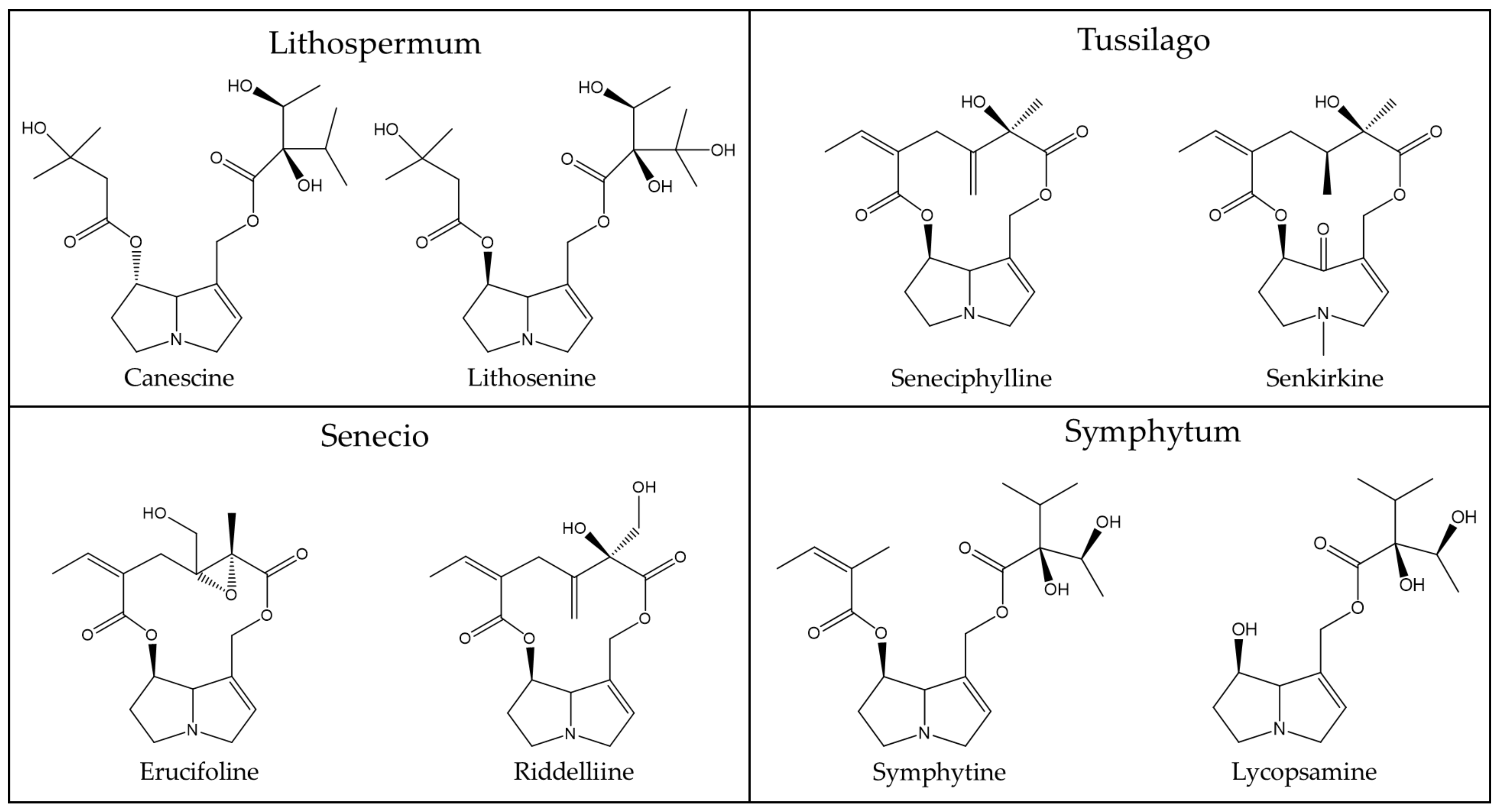
| Lithospermum (Boraginaceae) | Lithosenine [90], Lycopsamine [91], Canescine derivatives [91,92], Canescinine [91], Intermedine [91,93], Mysocorpine [93] |
| Petasites (Asteraceae) | Senkirkine [94,95], Senecionine [94,96], Integerrimine [96], Petasitenine [95], Neopetasitenine [95] |
| Senecio (Asteraceae) | Ridelline [97,98,99,100,101,102,103], Retrorsine [97,98,99,100,101,102,103], Seneciphylline [97,98,99,100,101,102,103], Senecionine [97,98,99,100,101,102,103], Senkirkine [97,98,99,102], Jacobine [104,105], Integerrimine [98,99,100,101,103], Spartiodine [98,99,100,103], Senecivernine [99,100,103,104,105], Platyphylline [97,98], Usaramine [97,99,103,104,105], Adinofoline [98,104], Florosenine [104], Erucifoline [104], Otosenine [104], Triangularine [106,107], 7-Angeloylheliotridine [108,109], Uspallatine [110], Rosmarinine [111,112], Angularine [111,112], Hadiensiene [111], Ruwenine [113], Ruzorine [113], Doriasenine [114], Sceleratine [115] |
| Symphytum (Boraginaceae) | Echimidine [116,117,118], Symphytine [116,117], Lasiocarpine [101], Intermedin [119], Lycopsamine [118,119] |
| Tussilago (Asteraceae) | Senkirkine [120,121], Senecionine [121], Intergerrimine [121], Seneciphylline [121], Senecivernine [122] |
| PA Type | Most Active CYP Isoforms |
|---|---|
| Clivorine | CYP 3A4 |
| Integerrimine | CYP 3A4, CYP 3A5 |
| Lasiocarine | CYP 3A4, CYP 3A5 |
| Monocrotaline | CYP 2A6 |
| Retrorsine | CYP 2A6, CYP 3A4, CYP 3A5 |
| Riddelline | CYP 3A4, CYP 3A5 |
| Senecionine | CYP 2A6, CYP 3A4, CYP 3A5 |
| Seneciphylline | CYP 2A6, CYP 3A4, CYP 3A5 |
| Senkirkine | CYP 3A4 |
| Parameter | Results |
|---|---|
| Cohort | n = 116 |
| Gender | Males 57 (50.4%) Females 56 (49.6%) (NA 3) |
| Age | 17–76 years |
| Ascites | 115/116 cases (99.1%) |
| Hepatomegaly | 104/113 cases (92.0%) |
| Jaundice | 95/113 cases (84%) |
| ALT elevation | 47/60 cases (78.3%) (NA 56 cases) |
| AST elevation | 50/58 cases (86.2%) (NA 58 cases) |
| Outcome | Recovery 75 cases (66.4%) Chronicity 27 cases (23.9%) Death 11 cases (9.7%) (NA 3 cases) |
| Suspected Product | Date | |
|---|---|---|
| Items for Hepatocellular Injury | Score | Result |
| 1. Time to onset from the beginning of the drug/herb consumption •5–90 days (rechallenge: 1–15 days) •<5 or >90 days (rechallenge: >15 days) Alternative: Time to onset from cessation of the drug/herb •≤15 days (except for slowly metabolized chemicals: >15 days) | +2 +1 +1 | □ □ □ |
| 2. Course of ALT after cessation of the drug/herb Percentage difference between ALT peak and ULN •Decrease ≥50% within 8 days •Decrease ≥50% within 30 days •No information or continued drug use •Decrease ≥50% after the 30th day •Decrease <50% after the 30th day or recurrent increase | +3 +2 0 0 −2 | □ □ □ □ □ |
| 3. Risk factors •Alcohol use (current drinks/d: >2 for women, >3 for men) •Alcohol use (current drinks/d: ≤2 for women, ≤3 for men) •Age ≥55 years •Age <55 years | +1 0 +1 0 | □ □ □ □ |
| 4. Concomitant drug(s)/herb(s) •None or no information •Concomitant drug/herb with incompatible time to onset •Concomitant drug/herb with time to onset 5–90 days •Concomitant drug/herb known as hepatotoxin and with time to onset •5–90 days •Concomitant drug/herb with evidence for its role in this case (positive rechallenge or validated test) | 0 0 −1 −2 −3 | □ □ □ □ □ |
| 5. Search for alternative causes Group I (7 causes) •HAV: Anti-HAV-IgM •HBV: HBsAg, anti-HBc-IgM, HBV-DNA •HCV: Anti-HCV, HCV-RNA •HEV: Anti-HEV-IgM, anti-HEV-IgG, HEV-RNA •Hepatobiliary sonography/Doppler/CT/MRC •Alcoholism (AST/ALT ≥2) •Acute recent hypotension history (particularly if underlying heart disease) Group II (5 causes) •Complications of underlying disease(s), such as sepsis, metastatic malig-nancy, autoimmune hepatitis, chronic hepatitis B or C, primary biliary cholangitis or sclerosing cholangitis, genetic liver diseases •Infection suggested by PCR and titer change for CMV (anti-CMV-IgM, anti-CMV-IgG) •EBV (anti-EBV-IgM, anti-EBV-IgG) •HSV (anti-HSV-IgM, anti-HSV-IgG) •VZV (anti-VZV-IgM, anti-VZV-IgG) Evaluation of groups I and II •All causes-groups I and II—reasonably ruled out •The 7 causes of group I ruled out •6 or 5 causes of group I ruled out •Less than 5 causes of group I ruled out •Alternative cause highly probable | Tick if negative □ □ □ □ □ □ □ □ □ □ □ □ +2 +1 0 −2 −3 | Tick if not done □ □ □ □ □ □ □ □ □ □ □ □ □ □ □ □ □ |
| 6. Previous hepatotoxicity of the drug/herb •Reaction labelled in the product characteristics •Reaction published but unlabelled •Reaction unknown | +2 +1 0 | □ □ □ |
| 7. Response to unintentional reexposure •Doubling of ALT with the drug/herb alone, provided ALT below 5 × ULN before reexposure •Doubling of ALT with the drug(s)/herb(s) already given at the time of first reaction •Increase of ALT but less than ULN in the same conditions as for the first administration •Other situations | +3 +1 −2 0 | □ □ □ □ |
| Total score of the patient under consideration through combining individual scores: | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teschke, R.; Vongdala, N.; Quan, N.V.; Quy, T.N.; Xuan, T.D. Metabolic Toxification of 1,2-Unsaturated Pyrrolizidine Alkaloids Causes Human Hepatic Sinusoidal Obstruction Syndrome: The Update. Int. J. Mol. Sci. 2021, 22, 10419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910419
Teschke R, Vongdala N, Quan NV, Quy TN, Xuan TD. Metabolic Toxification of 1,2-Unsaturated Pyrrolizidine Alkaloids Causes Human Hepatic Sinusoidal Obstruction Syndrome: The Update. International Journal of Molecular Sciences. 2021; 22(19):10419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910419
Chicago/Turabian StyleTeschke, Rolf, Noudeng Vongdala, Nguyen Van Quan, Tran Ngoc Quy, and Tran Dang Xuan. 2021. "Metabolic Toxification of 1,2-Unsaturated Pyrrolizidine Alkaloids Causes Human Hepatic Sinusoidal Obstruction Syndrome: The Update" International Journal of Molecular Sciences 22, no. 19: 10419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910419