
The Antifungal Mechanism of Isoxanthohumol from Humulus lupulus Linn.

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
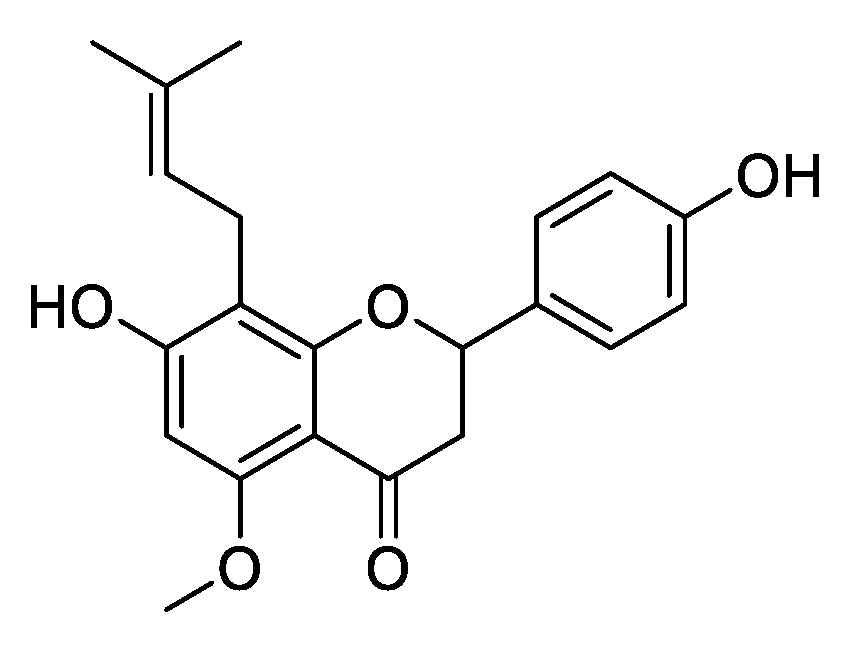
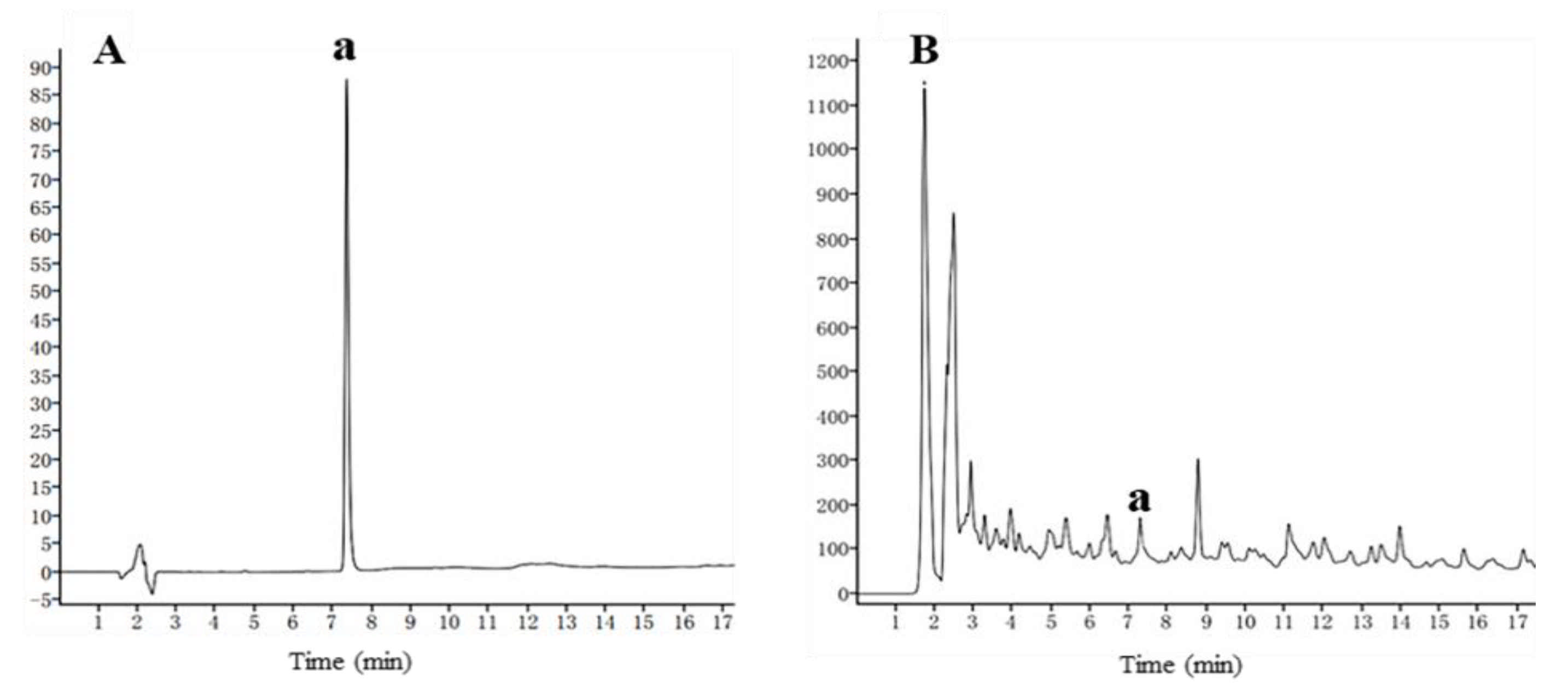
2.1. Antifungal Activity of H. lupulus Extract and Its Isoxanthohumol Content
2.2. Antifungal Activity of Isoxanthohumol
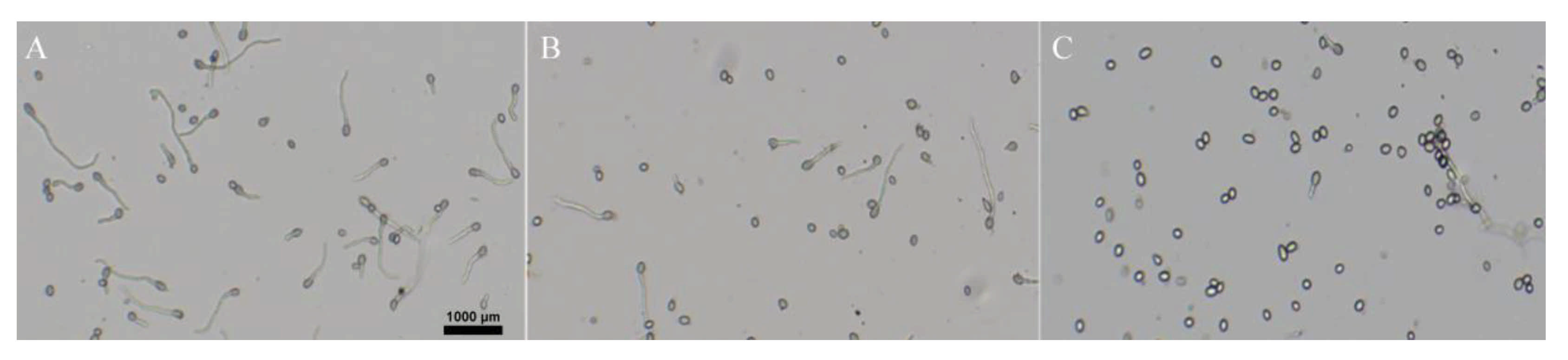
2.3. Effect of Isoxanthohumol on B. cinerea Spore Germination
2.4. Effect of Isoxanthohumol on B. cinerea Morphology
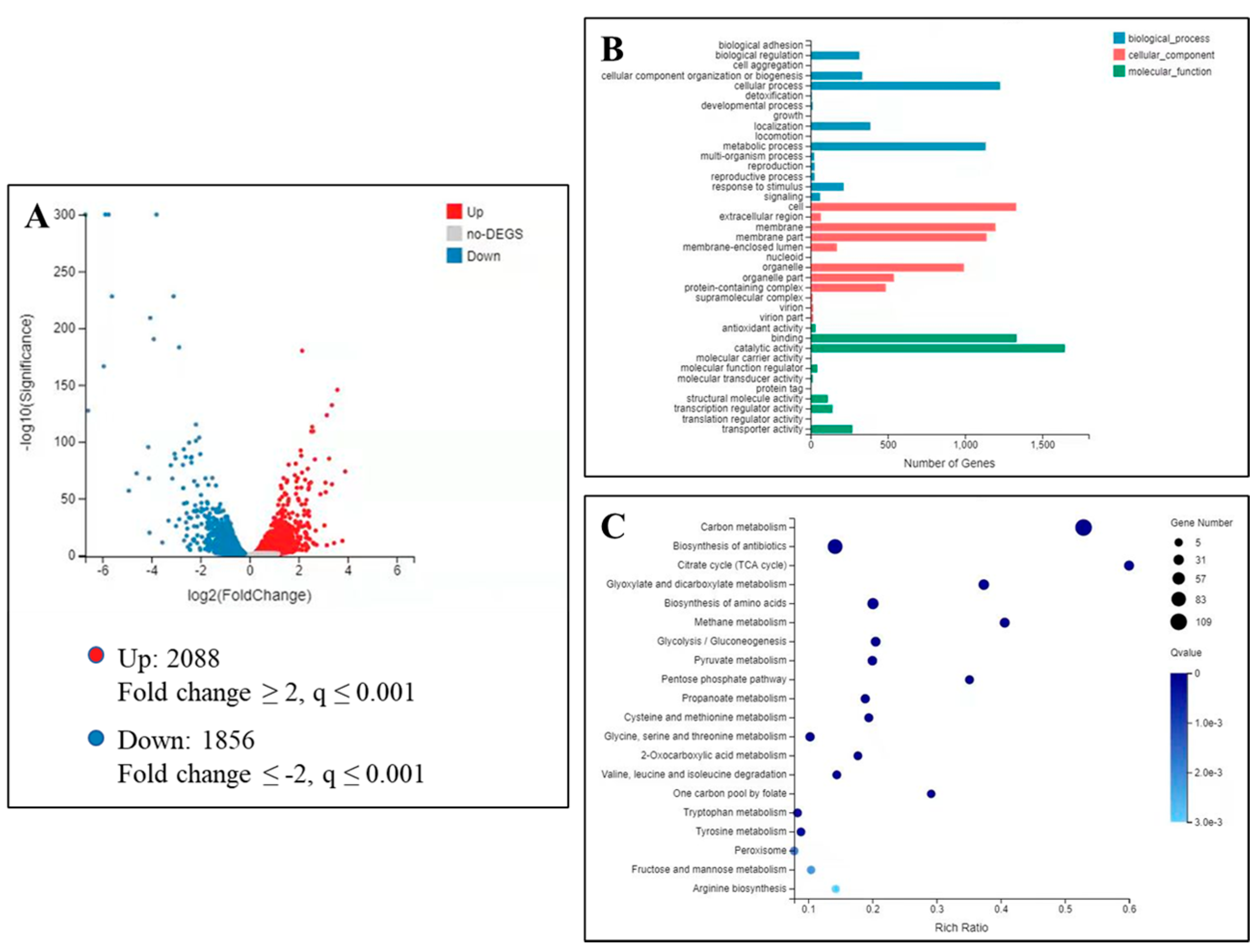
2.5. Transcriptome Analysis of B. cinerea under Isoxanthohumol Treatment
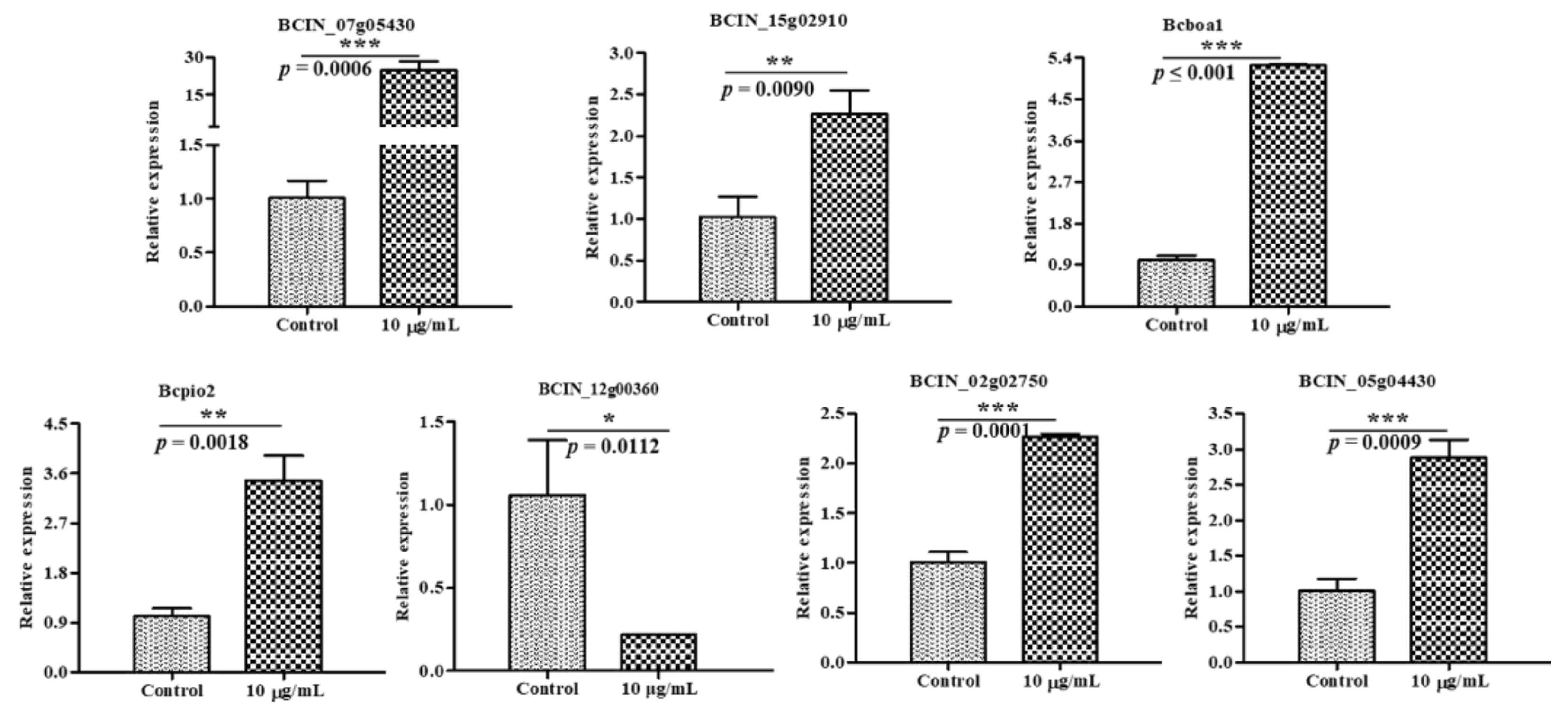
2.6. Verification of DEGs by RT-qPCR
2.7. Effects of Isoxanthohumol on B. cinerea Total Carbohydrate Content and TCA Cycle
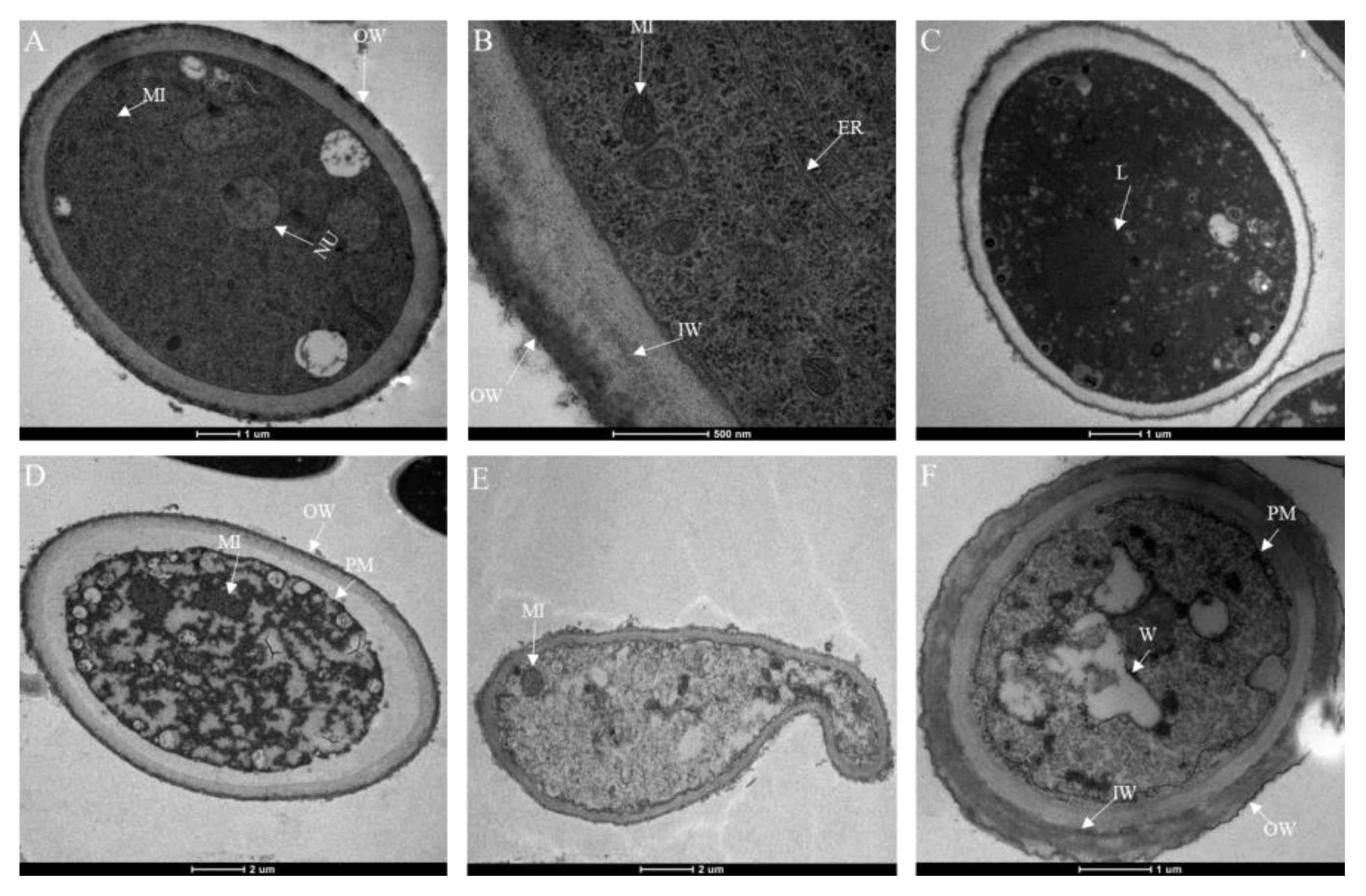
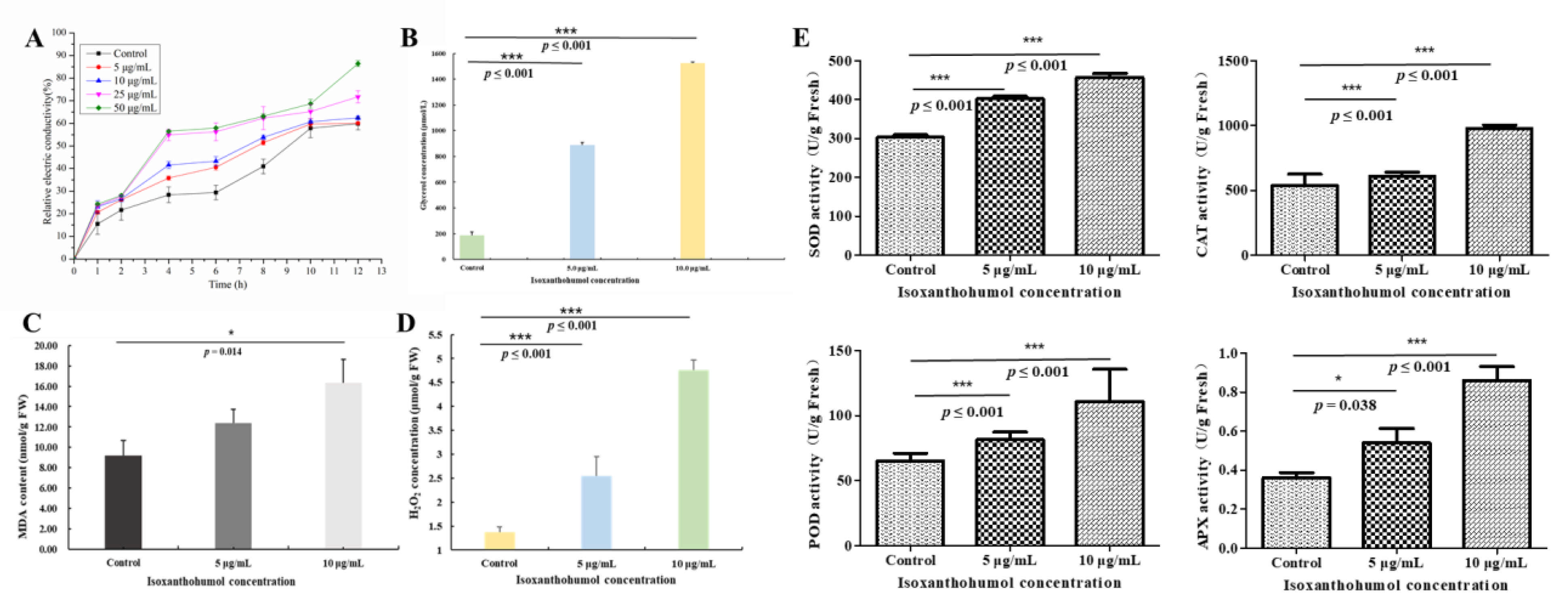
2.8. Changes Due to Isoxanthohumol in B. cinerea Cell Membrane
2.9. Effect of Isoxanthohumol on Membrane Lipid Peroxidation of B. cinerea
3. Materials and Methods
3.1. Pathogenic Fungi and Reagents
3.2. H. lupulus Extracts and HPLC Analysis
3.3. In Vitro Antifungal Activity
3.4. In Vivo Antifungal Activity
3.5. Effect of Isoxanthohumol on B. cinerea Spore Germination
3.6. Scanning Electron Microscopy (SEM) Observations
3.7. Transmission Electron Microscopy (TEM) Observations
3.8. Transcriptome Analysis
3.9. Verification of DEGs by RT-qPCR
3.10. Determination of Total Carbohydrate Content
3.11. Determination of Dehydrogenase Activities and CA Content
3.12. Determination of ATP Content and ATPase Activity
3.13. Determination of Cell Respiration
3.14. Cell Membrane Permeability
3.15. Determination of Glycerol Content
3.16. Determination of MDA Content
3.17. Determination of H2O2 Content
3.18. Activity of Antioxidant-Related Enzymes
3.19. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, C.; Yang, L.; Shao, Y.; Zhu, X.Y.; Zhao, H.L.; Chen, B.W.; Song, W.; Song, X.M.; Ding, X.F.; Sun, R.F. Broad-spectrum antifungal activity of dichloromethane extract of Waltheria indica stems and isolated compounds. Ind. Crop. Prod. 2019, 142, 111855. [Google Scholar] [CrossRef]
- Liu, T.T.; Wu, H.B.; Wu, H.B.; Zhang, J. Wormwood (Artemisia absinthium L.) as a promising nematicidal and antifungal agent: Chemical composition, comparison of extraction techniques and bioassay-guided isolation. Ind. Crop. Prod. 2019, 133, 295–303. [Google Scholar] [CrossRef]
- Bi, Y.H.; Yu, Z.G. Diterpenoids from Streptomyces sp. SN194 and their antifungal activity against Botrytis cinerea. J. Agric. Food Chem. 2016, 64, 8525–8529. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, J.; Zhang, P.; Xie, S.N.; Yuan, X.L.; Hou, X.D.; Yan, N.; Fang, Y.D.; Du, Y.M. In vitro and in vivo antifungal activity and preliminary mechanism of cembratrien-diols against Botrytis cinerea. Ind. Crop. Prod. 2020, 154, 112745. [Google Scholar] [CrossRef]
- Kumar, A.; Kujur, A.; Yadav, A.; Pratap, S.; Prakash, B. Optimization and mechanistic investigations on antifungal and aflatoxin B1 inhibitory potential of nanoencapsulated plant-based bioactive compounds. Ind. Crop. Prod. 2019, 131, 213–223. [Google Scholar] [CrossRef]
- Danielli, L.J.; Pippi, B.; Soares, K.D.; Duarte, J.A.; Maciel, A.J.; Machado, M.M.; Oliveira, L.F.S.; Bordignon, S.A.L.; Fuentefria, A.M.; Apel, M.A. Chemosensitization of filamentous fungi to antifungal agents using Nectandra Rol. ex Rottb. species essential oils. Ind. Crop. Prod. 2017, 102, 7–15. [Google Scholar] [CrossRef]
- Dan, W.J.; Tuong, T.M.L.; Wang, D.C.; Li, D.; Zhang, A.L.; Gao, J.M. Natural products as sources of new fungicides (V): Design and synthesis of acetophenone derivatives against phytopathogenic fungi in vitro and in vivo. Bioorg. Med. Chem. Lett. 2018, 28, 2861–2864. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.I.; Nenaah, G.E.; Mohamed, A.H.H. Antifungal activity of prenylated flavonoids isolated from Tephrosia apollinea L. against four phytopathogenic fungi. Crop Prot. 2013, 49, 21–25. [Google Scholar] [CrossRef]
- Di Liberto, M.G.; Stegmayer, M.I.; Svetaz, L.A.; Derita, M.G. Evaluation of Argentinean medicinal plants and isolation of their bioactive compounds as an alternative for the control of postharvest fruits phytopathogenic fungi. Rev. Bras. Farmacogn. 2019, 29, 686–688. [Google Scholar] [CrossRef]
- Wang, S.; Dunlap, T.L.; Howell, C.E.; Mbachu, O.C.; Rue, E.A.; Phansalkar, R.; Chen, S.N.; Pauli, G.F.; Dietz, B.M.; Bolton, J.L. Hop (Humulus lupulus L.) extract and 6-prenylnaringenin induce P450 1A1 catalyzed estrogen 2-hydroxylation. Chem. Res. Toxicol. 2016, 29, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, L.; Sahpaz, S.; Hilbert, J.L.; Rambaud, C.; Rivière, C. Humulus lupulus L., a very popular beer ingredient and medicinal plant: Overview of its phytochemistry, its bioactivity, and its biotechnology. Phytochem. Rev. 2018, 17, 1047–1090. [Google Scholar] [CrossRef]
- Karabín, M.; Hudcová, T.; Jelínek, L.; Dostálek, P. Biologically active compounds from hops and prospects for their use. Compr. Rev. Food Sci. Food Saf. 2016, 15, 542–567. [Google Scholar] [CrossRef] [Green Version]
- Kramer, B.; Mignard, C.; Warschat, D.; Gürbüz, S.; Aiglstorfer, P.; Muranyi, P. Inhibition of Listeria monocytogenes on bologna by a beta acid rich hop extract. Food Control 2021, 126, 108040. [Google Scholar] [CrossRef]
- Mizobuchi, S.; Sato, Y. A new flavanone with antifungal activity isolated from hops. Agric. Biol. Chem. 1984, 48, 2771–2775. [Google Scholar]
- Bocquet, L.; Rivière, C.; Dermont, C.; Samaillie, J.; Hilbert, J.H.; Halama, P.; Siah, A.; Sahpaz, S. Antifungal activity of hop extracts and compounds against the wheat pathogen Zymoseptoria tritici. Ind. Crop. Prod. 2018, 122, 290–297. [Google Scholar] [CrossRef]
- Bartmańska, A.; Wałecka-Zacharska, E.; Tronina, T.; Popłoński, J.; Sordon, S.; Brzezowska, E.; Bania, J.; Huszcza, E. Antimicrobial properties of spent hops extracts, flavonoids isolated therefrom, and their derivatives. Molecules 2018, 23, 2059. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.Y.; Li, J.L.; Sun, M.F.; Peng, J.N.; Li, X.H.; Qi, Z.Q. SYAUP–CN–26 applies its antifungal activity against Botrytis cinerea by disrupting mitochondrial structure and function. Biochimie 2020, 176, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, D.; Li, Y.M.; Chadwick, L.R.; Pauli, G.F.; van Breemen, R.B. Metabolism of xanthohumol and isoxanthohumol, prenylated flavonoids from hops (Humulus lupulus L.), by human liver microsomes. J. Mass Spectrom. 2005, 40, 289–299. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Romanazzi, G.; Smilanick, J.L.; Lichter, A. Postharvest ethanol and potassium sorbate treatments of table grapes to control gray mold. Postharvest Biol. Technol. 2005, 37, 129–134. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, B.; Chen, Y.; Liu, Z.Q.; Zheng, Y.G. Comparative transcriptome analysis of Streptomyces nodosus mutant with a high-yield amphotericin B. Front. Bioeng. Biotechnol. 2021, 8, 621431. [Google Scholar] [CrossRef]
- Lai, T.F.; Sun, Y.Y.; Liu, Y.Y.; Li, R.; Chen, Y.Z.; Zhou, T. Cinnamon oil inhibits Penicillium expansum growth by disturbing the carbohydrate metabolic process. J. Fungi 2021, 7, 123. [Google Scholar] [CrossRef]
- Lee, J.S.; Oh, S.J.; Choi, H.J.; Kang, J.H.; Lee, S.H.; Ha, J.S.; Woo, S.M.; Jang, H.; Lee, H.; Kim, S.Y. ATP production relies on fatty acid oxidation rather than glycolysis in pancreatic ductal adenocarcinoma. Cancers 2020, 12, 2477. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Shao, X.F.; Xu, J.Y.; Wei, Y.Y.; Xu, F.; Wang, H.F. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Z.; Yu, H.H.; Liu, W.X.; Qin, Y.K.; Xing, R.G.; Li, P.C. Integrated proteomics and metabolomics analysis reveals the antifungal mechanism of the C-coordinated O-carboxymethyl chitosan Cu(II) complex. Int. J. Biol. Macromol. 2020, 155, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, T.J.; Xu, M.J.; Guo, J.H.; Zhang, C.M.; Feng, Z.Z.; Peng, X.; Li, Z.Y.; Xing, K.; Qin, S. Antifungal effect of volatile organic compounds produced by Pseudomonas chlororaphis subsp. aureofaciens SPS-41 on oxidative stress and mitochondrial dysfunction of Ceratocystis fimbriata. Pestic. Biochem. Physiol. 2021, 173, 104777. [Google Scholar] [CrossRef]
- Yang, R.P.; Miao, J.Y.; Shen, Y.T.; Cai, N.; Wan, C.P.; Zou, L.Q.; Chen, C.Y.; Chen, J.J. Antifungal effect of cinnamaldehyde, eugenol and carvacrol nanoemulsion against Penicillium digitatum and application in postharvest preservation of citrus fruit. LWT-Food Sci. Technol. 2021, 141, 110924. [Google Scholar] [CrossRef]
- Chu, W.C.; Bai, P.Y.; Yang, Z.Q.; Cui, D.Y.; Hua, Y.G.; Yang, Y.; Yang, Q.Q.; Zhang, E.; Qin, S.S. Synthesis and antibacterial evaluation of novel cationic chalcone derivatives possessing broad spectrum antibacterial activity. Eur. J. Med. Chem. 2018, 143, 905–921. [Google Scholar] [CrossRef]
- Wang, C.M.; Huo, X.; Chen, J.; Liu, J.W.; Yang, T.Y.; Mi, X.Q.; Meng, Y.; Zhou, L.; Lin, C.J.; Liu, J. An acute lytic cell death induced by xanthohumol obstructed ROS detecting in HL-60 cells. Toxicol. In Vitro 2019, 62, 104667. [Google Scholar] [CrossRef]
- Wang, B.; Liu, F.; Li, Q.; Xu, S.; Zhao, X.Z.; Xue, P.L.; Feng, X. Antifungal activity of zedoary turmeric oil against Phytophthora capsici through damaging cell membrane. Pestic. Biochem. Physiol. 2019, 159, 59–67. [Google Scholar] [CrossRef]
- Duran, R.; Cary, J.W.; Calvo, A.M. Role of the osmotic stress regulatory pathway in morphogenesis and secondary metabolism in filamentous fungi. Toxins 2010, 2, 367–381. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhao, J.C.; Famous, E.; Pan, S.Y.; Peng, X.; Tian, J. Antioxidant, hepatoprotective and antifungal activities of black pepper (Piper nigrum L.) essential oil. Food Chem. 2021, 346, 128845. [Google Scholar] [CrossRef] [PubMed]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kontek, B.; Jedrejek, D.; Oleszek, W.; Olas, B. Antiradical and antioxidant activity in vitro of hops-derived extracts rich in bitter acids and xanthohumol. Ind. Crop. Prod. 2021, 161, 113208. [Google Scholar] [CrossRef]
- Kong, J.; Xie, Y.F.; Yu, H.; Guo, Y.H.; Cheng, Y.L.; Qian, H.; Yao, W.R. Synergistic antifungal mechanism of thymol and salicylic acid on Fusarium solani. LWT-Food Sci. Technol. 2021, 140, 110787. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (μg/mL) | Inhibition Rate a (%) ± SD | ||||
|---|---|---|---|---|---|---|
| R. solani | B. cinerea | F. graminearum | S. sclerotiorum | M. oryzae | ||
| Ethanolic extract | 500 | 37.01 ± 0.43 | 44.42 ± 0.27 | 48.19 ± 0.54 | 51.52 ± 0.53 | 43.40 ± 0.23 |
| 250 | 25.66 ± 0.18 | 40.76 ± 0.17 | 39.56 ± 0.18 | 47.97 ± 0.47 | 29.34 ± 0.29 | |
| Isoxanthohumol | 50 | 31.90 ± 1.08 | 85.90 ± 0.24 | 63.88 ± 0.23 | 84.46 ± 0.30 | 33.50 ± 0.27 |
| 25 | 18.02 ± 0.31 | 78.45 ± 0.32 | 60.43 ± 0.82 | 79.49 ± 0.05 | 28.10 ± 0.22 | |
| Pathogenic Fungus | EC50 (µg/mL) | Pathogenic Fungus (y = ax + b) | Correlation Coefficient (R2) |
|---|---|---|---|
| S. sclerotiorum | 14.52 | y = 3.78x − 4.43 | 0.973 |
| B. cinerea | 4.32 | y = 2.08x − 1.38 | 0.836 |
| F. graminearum | 16.50 | y = 1.64x − 2.00 | 0.976 |
| Compound | Concentration (μg/mL) | Protective Effect | Curative Effect | ||
|---|---|---|---|---|---|
| Lesion Length a (mm ± SD) | Control Efficacy (%) | Lesion Length (mm ± SD) | Control Efficacy (%) | ||
| Isoxanthohumol | 50 | 23.07 ± 2.62 | 32.35 | 24.77 ± 1.23 | 27.39 |
| 100 | 21.04 ± 3.98 | 38.32 | 22.09 ± 0.78 | 35.23 | |
| 200 | 15.40 ± 2.96 | 54.84 | 20.72 ± 1.92 | 39.25 | |
| Boscalid | 50 | 15.16 ± 5.02 | 42.93 | 30.78 ± 2.52 | 9.77 |
| 100 | 13.53 ± 5.62 | 49.09 | 16.90 ± 3.11 | 50.47 | |
| 200 | 7.98 ± 2.40 | 69.95 | 16.71 ± 3.18 | 51.00 | |
| Control | - | 26.57 ± 5.04 | - | 34.11 ± 2.32 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.-F.; Wu, T.-L.; Du, S.-S.; Wu, Z.-R.; Hu, Y.-M.; Zhang, Z.-J.; Zhao, W.-B.; Yang, C.-J.; Liu, Y.-Q. The Antifungal Mechanism of Isoxanthohumol from Humulus lupulus Linn. Int. J. Mol. Sci. 2021, 22, 10853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910853
Yan Y-F, Wu T-L, Du S-S, Wu Z-R, Hu Y-M, Zhang Z-J, Zhao W-B, Yang C-J, Liu Y-Q. The Antifungal Mechanism of Isoxanthohumol from Humulus lupulus Linn. International Journal of Molecular Sciences. 2021; 22(19):10853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910853
Chicago/Turabian StyleYan, Yin-Fang, Tian-Lin Wu, Sha-Sha Du, Zheng-Rong Wu, Yong-Mei Hu, Zhi-Jun Zhang, Wen-Bin Zhao, Cheng-Jie Yang, and Ying-Qian Liu. 2021. "The Antifungal Mechanism of Isoxanthohumol from Humulus lupulus Linn." International Journal of Molecular Sciences 22, no. 19: 10853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910853