
Genome-Wide Identification of CYP72A Gene Family and Expression Patterns Related to Jasmonic Acid Treatment and Steroidal Saponin Accumulation in Dioscorea zingiberensis


Abstract
:1. Introduction
2. Results
2.1. Identification of the CYP72A Proteins in D. zingiberensis
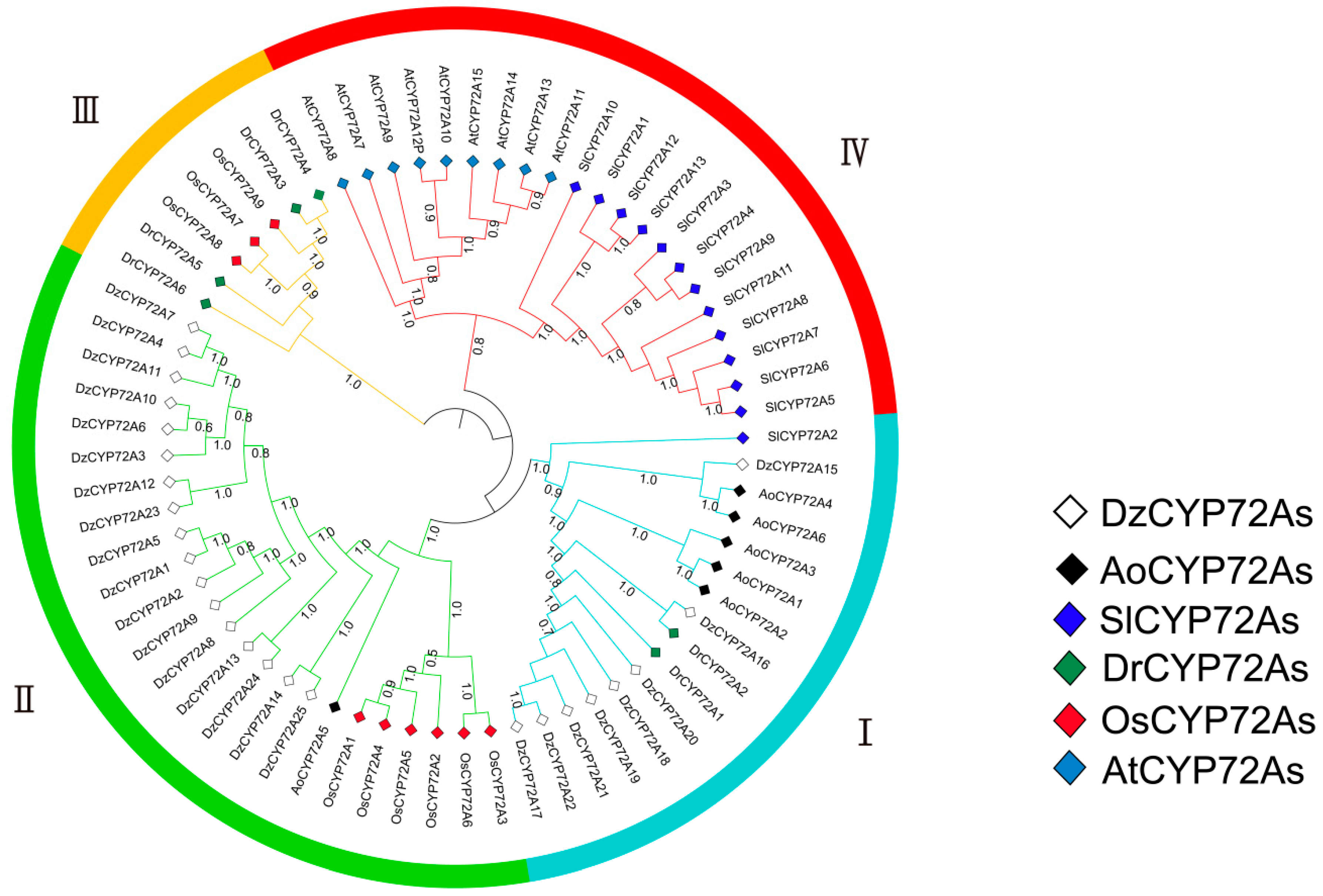
2.2. Phylogenetic Analysis and Multiple Sequence Alignment of DzCYP72A Genes
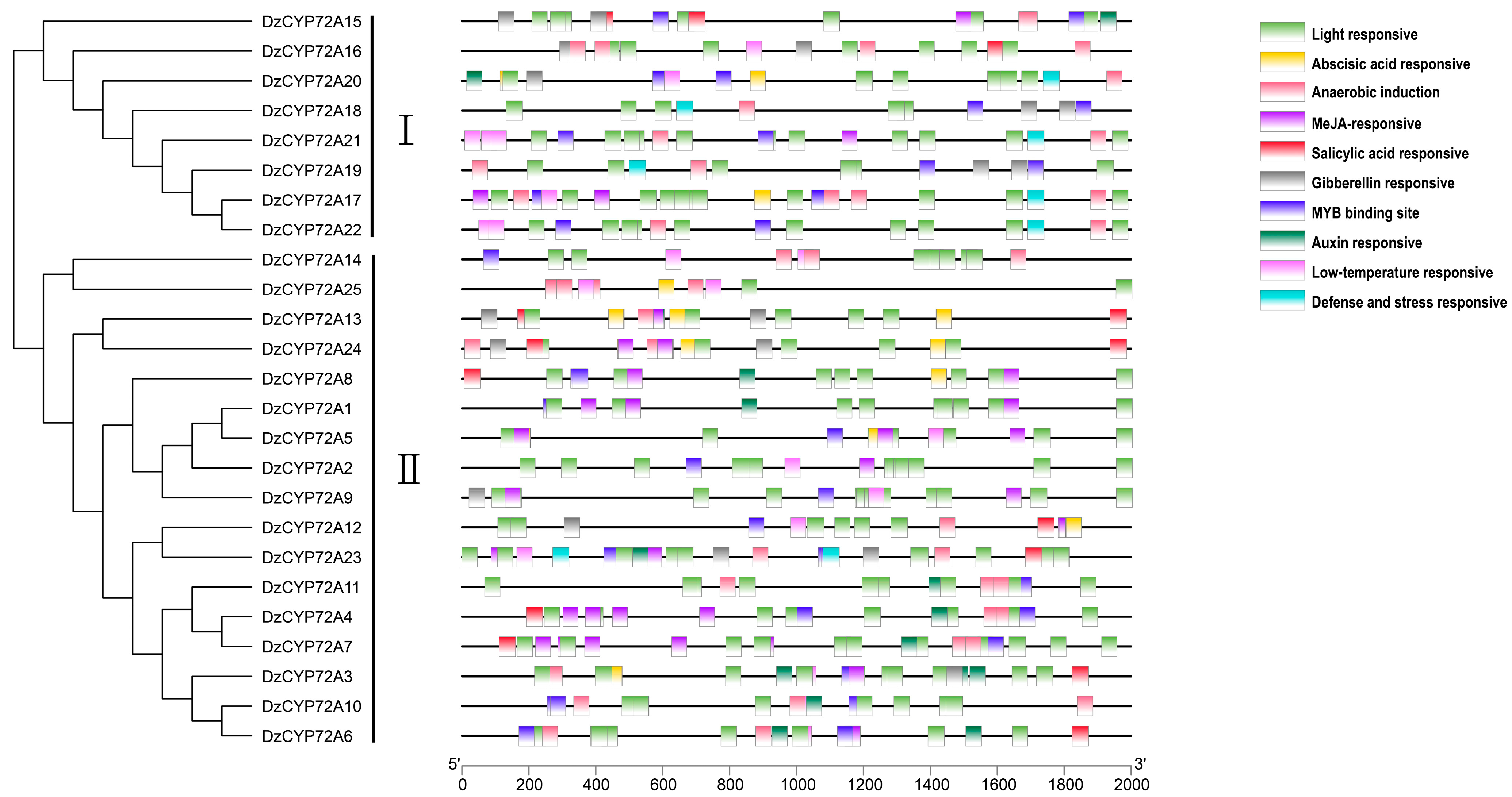
2.3. Gene Structure and Motif Analysis of DzCYP72As
2.4. Analysis of Cis-Regulatory Elements in the Promoters of DzCYP72As
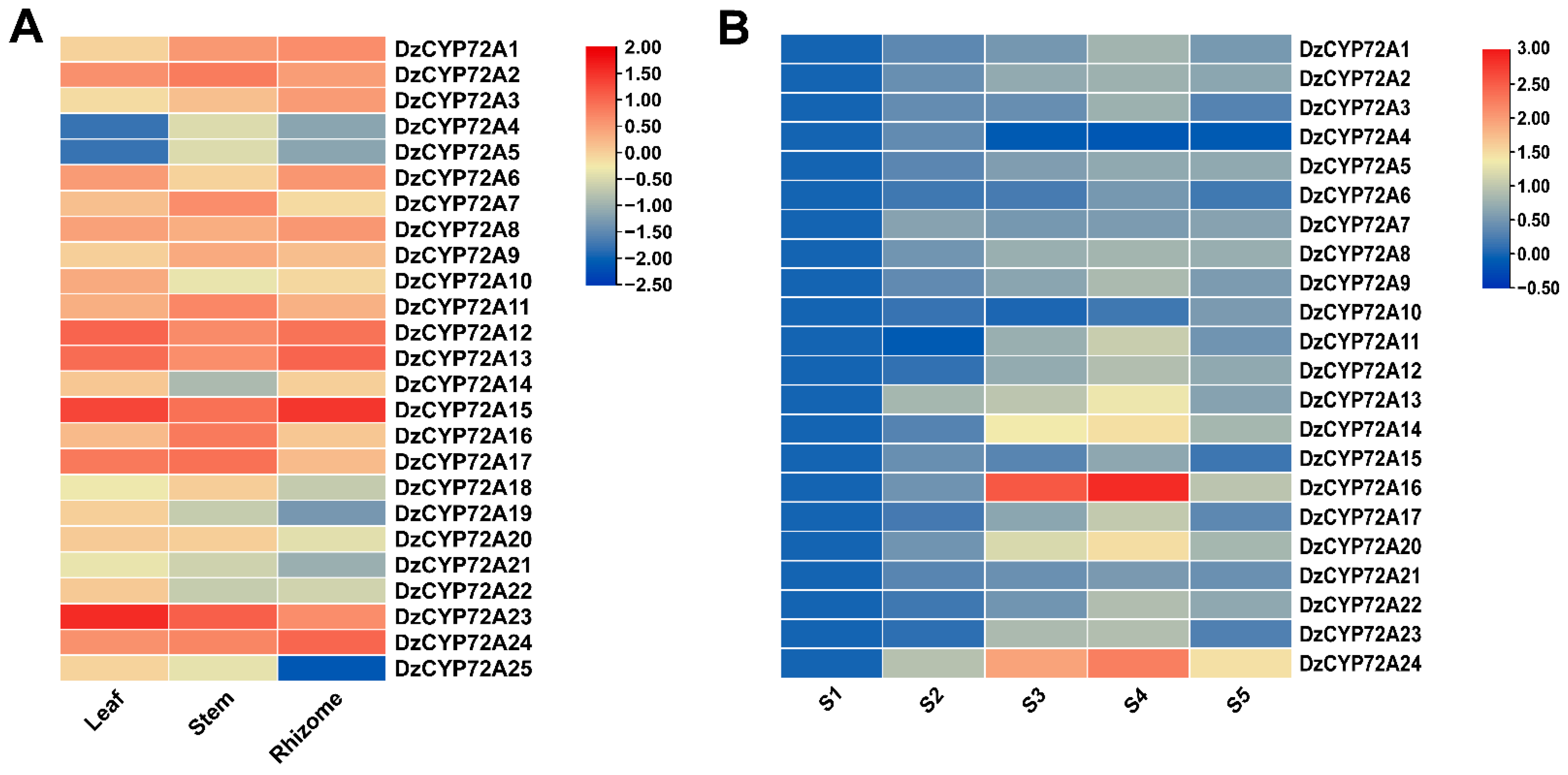
2.5. Expression Profiles of CYP72As in D. zingiberensis
2.6. Effects of JA Concentration on the Specialized Metabolites in D. zingiberensis
2.7. Correlations among DzCYP72As, Phytosterols, Diosgenin and Steroidal Saponins
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Standards and Chemical Reagents
4.3. RNA Extraction and Gene Expression Analysis
4.4. Identification and Screening of CYP72A Family Genes
4.5. Bioinformatics Analysis
4.6. Analysis of Specialized Metabolites
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, K.A. Cytochrome P450s as genes for crop improvement. Curr. Opin. Plant Biol. 2001, 4, 162–167. [Google Scholar] [CrossRef]
- Hansen, C.C.; Nelson, D.R.; Moller, B.L.; Werck-Reichhart, D. Plant cytochrome P450 plasticity and evolution. Mol. Plant 2021, 14, 1244–1265. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, Q.; Xin, P.; Yuan, J.; Ma, Y.; Wang, X.; Xu, M.; Chu, J.; Peters, R.J.; Wang, G. CYP72A enzymes catalyse 13-hydrolyzation of gibberellins. Nat. Plants 2019, 5, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Biazzi, E.; Carelli, M.; Tava, A.; Abbruscato, P.; Losini, I.; Avato, P.; Scotti, C.; Calderini, O. CYP72A67 Catalyzes a Key Oxidative Step in Medicago truncatula Hemolytic Saponin Biosynthesis. Mol. Plant 2015, 8, 1493–1506. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Nakajima, D.; Shinpo, S.; Nakamura, M.; Kawano, N.; Kawahara, N.; Yamazaki, M.; Saito, K.; Suzuki, H.; Hirakawa, H. Identification of potential genes involved in triterpenoid saponins biosynthesis in Gleditsia sinensis by transcriptome and metabolome analyses. J. Nat. Med. 2018, 73, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Mao, P.; Tao, X.; Wang, Y.; Wei, C.-M.; Ma, X.-R. Flexible shift on gene body methylation and transcription of LpCYP72A161 exposed to temperature stress in perennial ryegrass. Environ. Exp. Bot. 2017, 143, 29–37. [Google Scholar] [CrossRef]
- Chang, H.-Y.; Kao, M.-C.; Way, T.-D.; Ho, C.-T.; Fu, E. Diosgenin Suppresses Hepatocyte Growth Factor (HGF)-Induced Epithelial–Mesenchymal Transition by Down-regulation of Mdm2 and Vimentin. J. Agric. Food Chem. 2011, 59, 5357–5363. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, J.-J.; Zhou, S.-J.; Chen, J.; Hu, Q.; Pu, J.-X.; Lu, J.-L. Diosgenin inhibits the expression of NEDD4 in prostate cancer cells. Am. J. Transl. Res 2019, 11, 3461–3471. [Google Scholar]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-X.; Ito, Y.; Liang, J.-R.; Liu, J.-L.; He, J.; Sun, W.-J. Therapeutic effects of total steroid saponin extracts from the rhizome of Dioscorea zingiberensis C.H.Wright in Freund’s complete adjuvant induced arthritis in rats. Int. Immunopharmacol. 2014, 23, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashidume, T.; Sasaki, K.; Hirata, J.; Kato, M.; Yoshikawa, Y.; Iwasaki, Y.; Arai, H.; Miura, S.; Miyoshi, N. Effects of sanyaku and its constituent diosgenin on the fasted and postprandial hpertriacylglycerolemia in high-fat-diet-fed KK-Ay Mice. J. Agric. Food Chem. 2018, 66, 9968–9975. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-N.; He, X.-C.; Ye, M.; Huang, H.; Chen, H.-L.; Peng, W.-L.; Zhao, Z.-Z.; Yi, T.; Chen, H.-B. Cardioprotective effect of total saponins from three medicinal species of Dioscorea against isoprenaline-induced myocardial ischemia. J. Ethnopharmacol. 2015, 175, 451–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciura, J.; Szeliga, M.; Grzesik, M.; Tyrka, M. Changes in fenugreek transcriptome induced by methyl jasmonate and steroid precursors revealed by RNA-Seq. Genomics 2018, 110, 267–276. [Google Scholar] [CrossRef]
- Fukushima, E.O.; Seki, H.; Sawai, S.; Suzuki, M.; Ohyama, K.; Saito, K.; Muranaka, T. Combinatorial Biosynthesis of Legume Natural and Rare Triterpenoids in Engineered Yeast. Plant Cell Physiol. 2013, 54, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Zhang, Y.; Jia, L.; Sun, L.; Wang, S.; Xiao, J.; Zhan, Y.; Yin, J. Functional identification of five CYP450 genes from birch responding to MeJA and SA in the synthesis of betulinic acid from lupitol. Ind. Crop. Prod. 2021, 167, 113513. [Google Scholar] [CrossRef]
- Xue, Z.; Cao, Z.; Jin, M.; Zhang, X.; Wang, X.; Dou, J.; Zhu, Y.; Ito, Y.; Guo, Z. New steroid saponins from Dioscorea zingiberensis yam and their medicinal use against I/R via anti-inflammatory effect. Food Funct. 2021, 12, 8314–8325. [Google Scholar] [CrossRef] [PubMed]
- Alsoufi, A.S.M.; Pączkowski, C.; Szakiel, A.; Długosz, M. Effect of jasmonic acid and chitosan on triterpenoid production in Calendula officinalis hairy root cultures. Phytochem. Lett. 2019, 31, 5–11. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Wu, X.; Liu, D.; Li, J.; Li, J.; Liu, S.; Gao, W. Jasmonic acid and methyl dihydrojasmonate enhance saponin biosynthesis as well as expression of functional genes in adventitious roots of Panax notoginseng F.H. Chen. Biotechnol. Appl. Biochem. 2017, 64, 225–238. [Google Scholar] [CrossRef]
- Chaudhary, S.; Chikara, S.K.; Sharma, M.C.; Chaudhary, A.; Alam Syed, B.; Chaudhary, P.S.; Mehta, A.; Patel, M.; Ghosh, A.; Iriti, M. Elicitation of Diosgenin Production in Trigonella foenum-graecum (Fenugreek) Seedlings by Methyl Jasmonate. Int. J. Mol. Sci. 2015, 16, 29889–29899. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Li, S.; Tong, Z.; Yuan, X.; Xu, J.; Li, J. Geographical variations in fatty acid and steroid saponin biosynthesis in Dioscorea zingiberensis rhizomes. Ind. Crop. Prod. 2021, 170, 113779. [Google Scholar] [CrossRef]
- Hill, M.O.; Gauch, H.G. Detrended correspondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Abdollahi, F.; Alebrahim, M.T.; Ngov, C.; Lallemand, E.; Zheng, Y.; Villette, C.; Zumsteg, J.; André, F.; Navrot, N.; Werck-Reichhart, D.; et al. Innate promiscuity of the CYP706 family of P450 enzymes provides a suitable context for the evolution of dinitroaniline resistance in weed. New Phytol. 2021, 229, 3253–3268. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Khakimov, B.; Cárdenas, P.D.; Cozzi, F.; Olsen, C.E.; Jensen, K.R.; Hauser, T.P.; Bak, S. The cytochrome P450 CYP72A552 is key to production of hederagenin-based saponins that mediate plant defense against herbivores. New Phytol. 2019, 222, 1599–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination. BMC Biol. 2017, 15, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayasu, M.; Umemoto, N.; Akiyama, R.; Ohyama, K.; Lee, H.J.; Miyachi, H.; Watanabe, B.; Muranaka, T.; Saito, K.; Sugimoto, Y.; et al. Characterization of C-26 aminotransferase, indispensable for steroidal glycoalkaloid biosynthesis. Plant J. 2021. [Google Scholar] [CrossRef]
- Yang, L.; Cao, H.; Zhang, X.; Gui, L.; Chen, Q.; Qian, G.; Xiao, J.; Li, Z. Genome-Wide Identification and Expression Analysis of Tomato ADK Gene Family during Development and Stress. Int. J. Mol. Sci. 2021, 22, 7708. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-B.; Ma, Y.-N.; Qin, T.-F.; Tang, W.-L.; Qi, X.-W.; Wang, X.; Liu, R.-C.; Fang, H.-L.; Chen, Z.-Q.; Liang, C.-Y.; et al. Transcriptome-Wide Identification and Characterization of the JAZ Gene Family in Mentha canadensis L. Int. J. Mol. Sci. 2021, 22, 8859. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, K.; Li, Y. Highlights to phytosterols accumulation and equilibrium in plants: Biosynthetic pathway and feedback regulation. Plant Physiol. Biochem. 2020, 155, 637–649. [Google Scholar] [CrossRef]
- Li, J.; Zhou, H.; Pan, X.; Li, Z.; Lu, Y.; He, N.; Meng, T.; Yao, C.; Chen, C.; Ling, X. The role of fluconazole in the regulation of fatty acid and unsaponifiable matter biosynthesis in Schizochytrium sp. MYA 1381. BMC Microbiol. 2019, 19, 256. [Google Scholar] [CrossRef]
- Song, N.; Cheng, Y.; Peng, W.; Peng, E.; Zhao, Z.; Liu, T.; Yi, T.; Dai, L.; Wang, B.; Hong, Y. Genome-Wide Characterization and Expression Analysis of the SBP-Box Gene Family in Sweet Orange (Citrus sinensis). Int. J. Mol. Sci. 2021, 22, 8918. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Liu, Q.; Zhang, T.; Chong, X.; Yuan, H. Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L. Int. J. Mol. Sci. 2021, 22, 8793. [Google Scholar] [CrossRef]
- Maes, L.; Van Nieuwerburgh, F.C.W.; Zhang, Y.S.; Reed, D.W.; Pollier, J.; Casteele, S.R.F.V.; Inze, D.; Covello, P.S.; Deforce, D.L.D.; Goossens, A. Dissection of the phytohormonal regulation of trichome formation and biosynthesis of the antimalarial compound artemisinin in Artemisia annua plant. New Phytol. 2011, 189, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Stitz, M.; Gase, K.; Baldwin, I.T.; Gaquerel, E. Ectopic Expression of AtJMT in Nicotiana attenuata: Creating a Metabolic Sink Has Tissue-Specific Consequences for the Jasmonate Metabolic Network and Silences Downstream Gene Expression. Plant Physiol. 2011, 157, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jin, M.; Tadesse, N.; Dang, J.; Zhou, T.; Zhang, H.; Wang, S.; Guo, Z.; Ito, Y. Dioscorea zingiberensis C. H. Wright: An overview on its traditional use, phytochemistry, pharmacology, clinical applications, quality control, and toxicity. J. Ethnopharmacol. 2018, 220, 283–293. [Google Scholar] [CrossRef]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, J.; Liu, X.; Li, X.; Zhang, W.; Dai, Z.; Lu, L.; Zhou, X.; Cai, J.; Zhang, X.; et al. The origin and evolution of the diosgenin biosynthetic pathway in yam. Plant Commun. 2021, 2, 100079. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, K.; Ghosh, A.; Kumar, V.; Chaudhary, S.; Srivastava, N.; Katudia, K.; Tiwari, T.; Chikara, S.K. De Novo Transcriptome Sequencing in Trigonella foenum-graecum L. to Identify Genes Involved in the Biosynthesis of Diosgenin. Plant Genome 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Huang, Y.; Dong, X.; Wang, R.; Tang, M.; Cai, J.; Chen, J.; Zhang, X.; Nie, G. Exogenous Methyl Jasmonate Improves Heat Tolerance of Perennial Ryegrass Through Alteration of Osmotic Adjustment, Antioxidant Defense, and Expression of Jasmonic Acid-Responsive Genes. Front. Plant Sci. 2021, 12, 664519. [Google Scholar] [CrossRef]
- Paponov, M.; Antonyan, M.; Slimestad, R.; Paponov, I.A. Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting. Biomolecules 2021, 11, 1283. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.M.; Bak, S.; Feyereisen, R. Intron–Exon Organization and Phylogeny in a Large Superfamily, the Paralogous Cytochrome P450 Genes of Arabidopsis thaliana. DNA Cell Biol. 2000, 19, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Amino Acids | MW (kD) | pI | Instability Index | Aliphatic Index | GRAVY | Genomic Location |
|---|---|---|---|---|---|---|---|
| DzCYP72A1 | 518 | 59.53 | 9.42 | 50.42 | 92.08 | −0.207 | Chr1:54408546–54412582 |
| DzCYP72A2 | 520 | 59.52 | 9.09 | 46.29 | 93.19 | −0.169 | Chr1:54419523–54423635 |
| DzCYP72A3 | 516 | 58.89 | 9.01 | 45.88 | 92.60 | −0.192 | Chr1:54482572–54486042 |
| DzCYP72A4 | 519 | 59.74 | 8.98 | 42.74 | 92.02 | −0.182 | Chr1:54491399–54496848 |
| DzCYP72A5 | 519 | 59.40 | 9.23 | 49.52 | 94.68 | −0.161 | Chr1:54620742–54635835 |
| DzCYP72A6 | 499 | 56.98 | 8.77 | 48.86 | 91.44 | −0.208 | Chr1:54694341–54697800 |
| DzCYP72A7 | 486 | 55.91 | 9.06 | 47.04 | 90.86 | −0.241 | Chr1:54703122–54708457 |
| DzCYP72A8 | 518 | 59.46 | 9.16 | 47.69 | 92.82 | −0.186 | Chr1:54817699–54821732 |
| DzCYP72A9 | 493 | 56.52 | 9.03 | 44.79 | 92.35 | −0.176 | Chr1:54828984–54833020 |
| DzCYP72A10 | 518 | 59.28 | 9.04 | 46.59 | 93.92 | −0.200 | Chr1:54911925–54915459 |
| DzCYP72A11 | 517 | 59.57 | 9.10 | 43.23 | 93.52 | −0.162 | Chr1:54920693–54924967 |
| DzCYP72A12 | 518 | 59.21 | 9.27 | 47.44 | 95.98 | −0.142 | Chr1:54948190–54951851 |
| DzCYP72A13 | 518 | 59.29 | 9.19 | 46.19 | 95.98 | −0.186 | Chr1:54952667–54955365 |
| DzCYP72A14 | 525 | 60.06 | 9.44 | 46.68 | 93.79 | −0.204 | Chr1:55018259–55022509 |
| DzCYP72A15 | 519 | 59.51 | 8.23 | 37.99 | 97.86 | −0.093 | Chr4:20462820–20465388 |
| DzCYP72A16 | 515 | 59.04 | 8.36 | 45.35 | 94.45 | −0.167 | Chr9:1757458–1760130 |
| DzCYP72A17 | 486 | 55.77 | 8.12 | 50.32 | 92.49 | −0.271 | Chr9:1797953–1806784 |
| DzCYP72A18 | 451 | 51.87 | 8.40 | 46.16 | 91.02 | −0.292 | Chr9:1957935–1963395 |
| DzCYP72A19 | 173 | 19.63 | 6.29 | 42.93 | 94.68 | −0.070 | Chr9:1963533–1964154 |
| DzCYP72A20 | 503 | 57.97 | 9.13 | 47.24 | 95.37 | −0.203 | Chr9:2174147–2176010 |
| DzCYP72A21 | 487 | 56.05 | 7.27 | 51.27 | 93.31 | −0.244 | Chr9:2181213–2183031 |
| DzCYP72A22 | 486 | 55.86 | 8.58 | 50.57 | 91.30 | −0.290 | Chr9:2219226–2221041 |
| DzCYP72A23 | 518 | 59.07 | 9.22 | 46.65 | 96.74 | −0.121 | Unknown |
| DzCYP72A24 | 518 | 59.36 | 9.11 | 46.56 | 95.23 | −0.207 | Unknown |
| DzCYP72A25 | 525 | 60.09 | 9.41 | 46.32 | 93.79 | −0.205 | Unknown |
| Gene Name | Plant-PLoc | WoLF PSORT | CELLO | YLoc |
|---|---|---|---|---|
| CYP72A1 | chloroplast | chlo: 5 | mito(2.82) | chloroplast |
| CYP72A2 | mitochondrion | chlo: 6 | mito(2.38) | chloroplast |
| CYP72A3 | chloroplast | chlo: 6 | mito(2.07) | chloroplast |
| CYP72A4 | cytoplasm | plas: 7 | mito(2.06) | chloroplast |
| CYP72A5 | chloroplast | chlo: 6 | mito(2.76) | chloroplast |
| CYP72A6 | chloroplast | chlo: 5 | mito(1.7)/cyto(1.33) | chloroplast |
| CYP72A7 | chloroplast | cyto: 9 | mito(2.19) | nucleus |
| CYP72A8 | mitochondrion | chlo: 6 | mito(2.56) | chloroplast |
| CYP72A9 | mitochondrion | chlo: 7 | mito(2.08)/cyto(1.39) | chloroplast |
| CYP72A10 | chloroplast | chlo: 8 | mito(2.15) | chloroplast |
| CYP72A11 | cytoplasm | chlo: 8 | mito(2.07) | chloroplast |
| CYP72A12 | cytoplasm | chlo: 8 | mito(2.16) | chloroplast |
| CYP72A13 | cytoplasm | chlo: 5 | mito(2.59) | chloroplast |
| CYP72A14 | cytoplasm | E.R.: 5 | mito(2.97) | chloroplast |
| CYP72A15 | chloroplast | chlo: 9 | mito(1.68)/cyto(1.67) | chloroplast |
| CYP72A16 | chloroplast | chlo: 5 | cyto(1.71) | chloroplast |
| CYP72A17 | chloroplast | cyto: 7 | cyto(1.70)/mito(1.21)/nucl(1.14) | nucleus |
| CYP72A18 | chloroplast | cyto: 9 | cyto(3.09) | cytoplasm |
| CYP72A19 | cytoplasm | cyto: 11 | cyto(2.26) | cytoplasm |
| CYP72A20 | chloroplast | cyto:11 | mito(1.99)/cyto(1.42) | secreted pathway |
| CYP72A21 | chloroplast | cyto:7 | cyto(1.97) | chloroplast |
| CYP72A22 | chloroplast | cyto: 7 | cyto(1.40)/nucl(1.35)/mito(1.30) | nucleus |
| CYP72A23 | chloroplast | chlo: 5 | mito(2.21) | chloroplast |
| CYP72A24 | chloroplast | chlo: 5 | mito(2.63) | chloroplast |
| CYP72A25 | cytoplasm | E.R.: 5 | mito(2.83) | chloroplast |
| Compound Name | 0 μmol/L | 25 μmol/L | 50 μmol/L | 100 μmol/L | 200 μmol/L |
|---|---|---|---|---|---|
| Parvifloside (mg/g) | 34.88 ± 0.85 c | 39.26 ± 1.08 b | 24.83 ± 1.25 d | 54.89 ± 0.88 a | 34.20 ± 0.55 c |
| Protodeltonin (mg/g) | 3.81 ± 0.21 d | 10.75 ± 1.51 bc | 15.69 ± 1.68 a | 12.37 ± 0.11 b | 10.38 ± 0.25 c |
| Dioscin (µg/g) | 19.35 ± 1.00 c | 23.79 ± 0.84 b | 38.25 ± 1.07 a | 23.20 ± 1.07 b | 11.44 ± 0.23 d |
| Diosgenin (µg/g) | 404.30 ± 19.97 d | 434.24 ± 6.37 d | 595.27 ± 20.86 b | 647.18 ± 24.21 a | 478.64 ± 22.59 c |
| Campesterol (µg/g) | 78.33 ± 10.92 a | 76.12 ± 2.06 ab | 80.53 ± 19.24 a | 88.13 ± 21.41 a | 51.1 ± 1.17 b |
| Stigmasterol (µg/g) | 128.91 ± 3.92 b | 117.33 ± 4.64 ab | 253.94 ± 16.34 a | 68.93 ± 6.93 c | 69.68 ± 1.66 c |
| β-Sitosterol (µg/g) | 278.62 ± 12.23 bc | 299.21 ± 16.38 ab | 281.27 ± 17.31 c | 322.62 ± 14.77 a | 226.12 ± 12.55 d |
| Statistics | RDA1 | RDA2 | F | Pr(>F) |
|---|---|---|---|---|
| DzCYP72A1 | 0.9925 | −0.1219 | 2.6747 | 0.0500 * |
| DzCYP72A3 | 0.9799 | 0.1996 | 2.6286 | 0.0417 * |
| DzCYP72A6 | 0.9486 | 0.3164 | 2.9872 | 0.0083 ** |
| DzCYP72A9 | 0.9873 | −0.1586 | 3.0657 | 0.0417 * |
| DzCYP72A11 | 0.9947 | −0.1026 | 3.7190 | 0.0417 * |
| DzCYP72A13 | 0.9960 | 0.0896 | 2.5909 | 0.0667 |
| DzCYP72A14 | 0.9838 | −0.1794 | 4.1352 | 0.0167 * |
| DzCYP72A16 | 0.9993 | −0.0386 | 4.9162 | 0.0083 ** |
| DzCYP72A17 | 0.9997 | 0.0241 | 4.3933 | 0.0083 ** |
| DzCYP72A20 | 0.9812 | −0.1931 | 3.8978 | 0.0167 * |
| DzCYP72A21 | 0.9313 | −0.3643 | 1.9759 | 0.1333 |
| DzCYP72A22 | 0.9250 | −0.3800 | 2.4202 | 0.1583 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, L.; Yuan, X.; Li, S.; Li, Y.; Li, Z.; Li, J. Genome-Wide Identification of CYP72A Gene Family and Expression Patterns Related to Jasmonic Acid Treatment and Steroidal Saponin Accumulation in Dioscorea zingiberensis. Int. J. Mol. Sci. 2021, 22, 10953. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010953
Hou L, Yuan X, Li S, Li Y, Li Z, Li J. Genome-Wide Identification of CYP72A Gene Family and Expression Patterns Related to Jasmonic Acid Treatment and Steroidal Saponin Accumulation in Dioscorea zingiberensis. International Journal of Molecular Sciences. 2021; 22(20):10953. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010953
Chicago/Turabian StyleHou, Lixiu, Xincheng Yuan, Song Li, Yi Li, Zihao Li, and Jiaru Li. 2021. "Genome-Wide Identification of CYP72A Gene Family and Expression Patterns Related to Jasmonic Acid Treatment and Steroidal Saponin Accumulation in Dioscorea zingiberensis" International Journal of Molecular Sciences 22, no. 20: 10953. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010953