
Macrophage-Dependent Interleukin-6-Production and Inhibition of IK Contributes to Acquired QT Prolongation in Lipotoxic Guinea Pig Heart

 ,
,
Abstract
:1. Introduction
2. Results
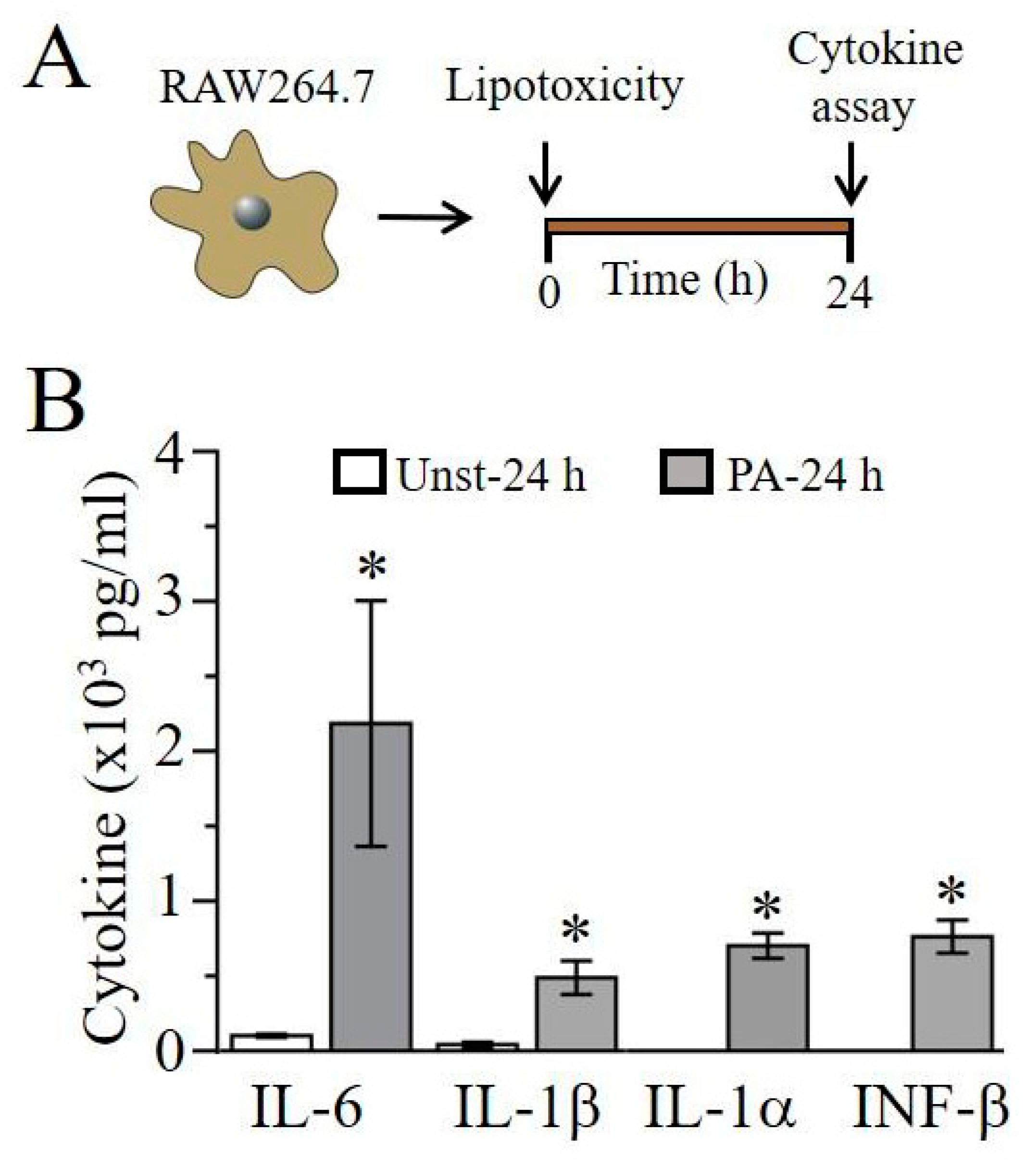
2.1. Lipotoxicity Induces Pro-Inflammatory Cytokine Release in RAW264.7 Cells
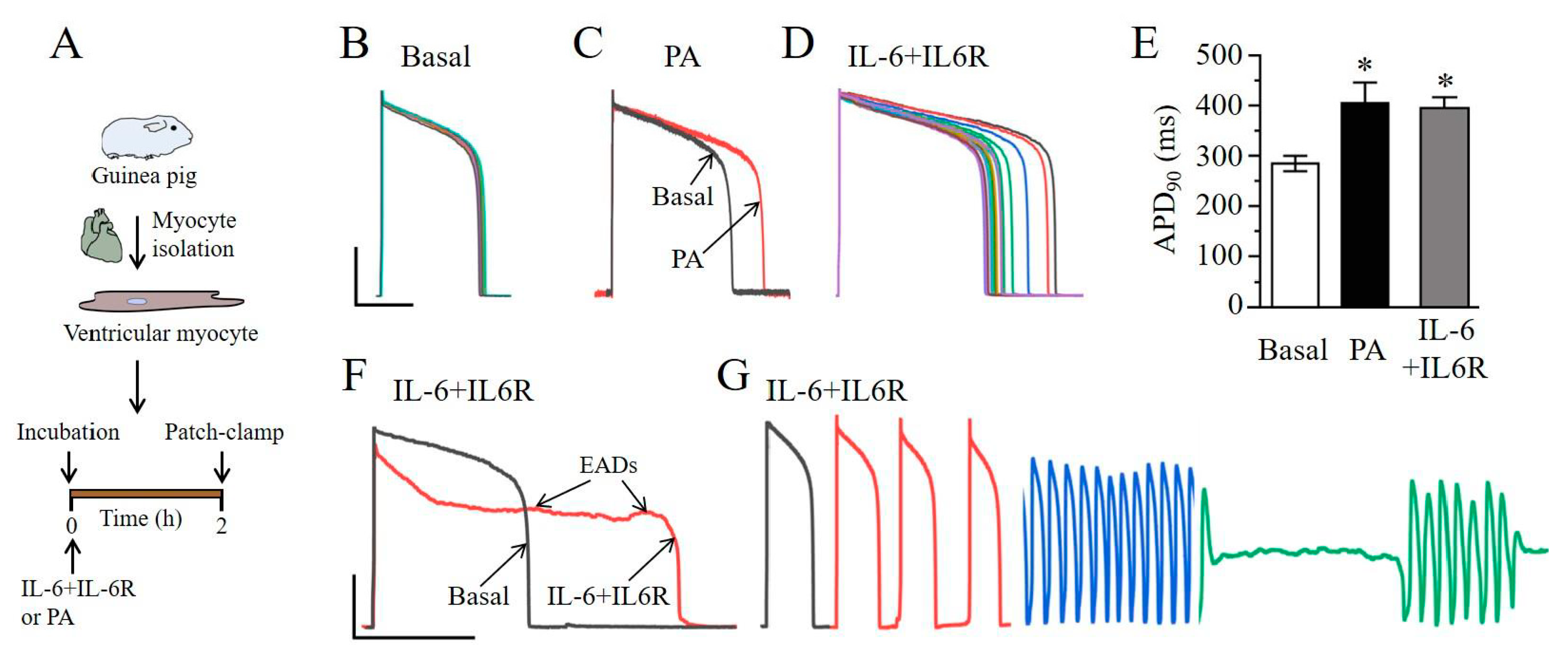
2.2. IL-6 Induces Ventricular Myocyte Electrical Vulnerability
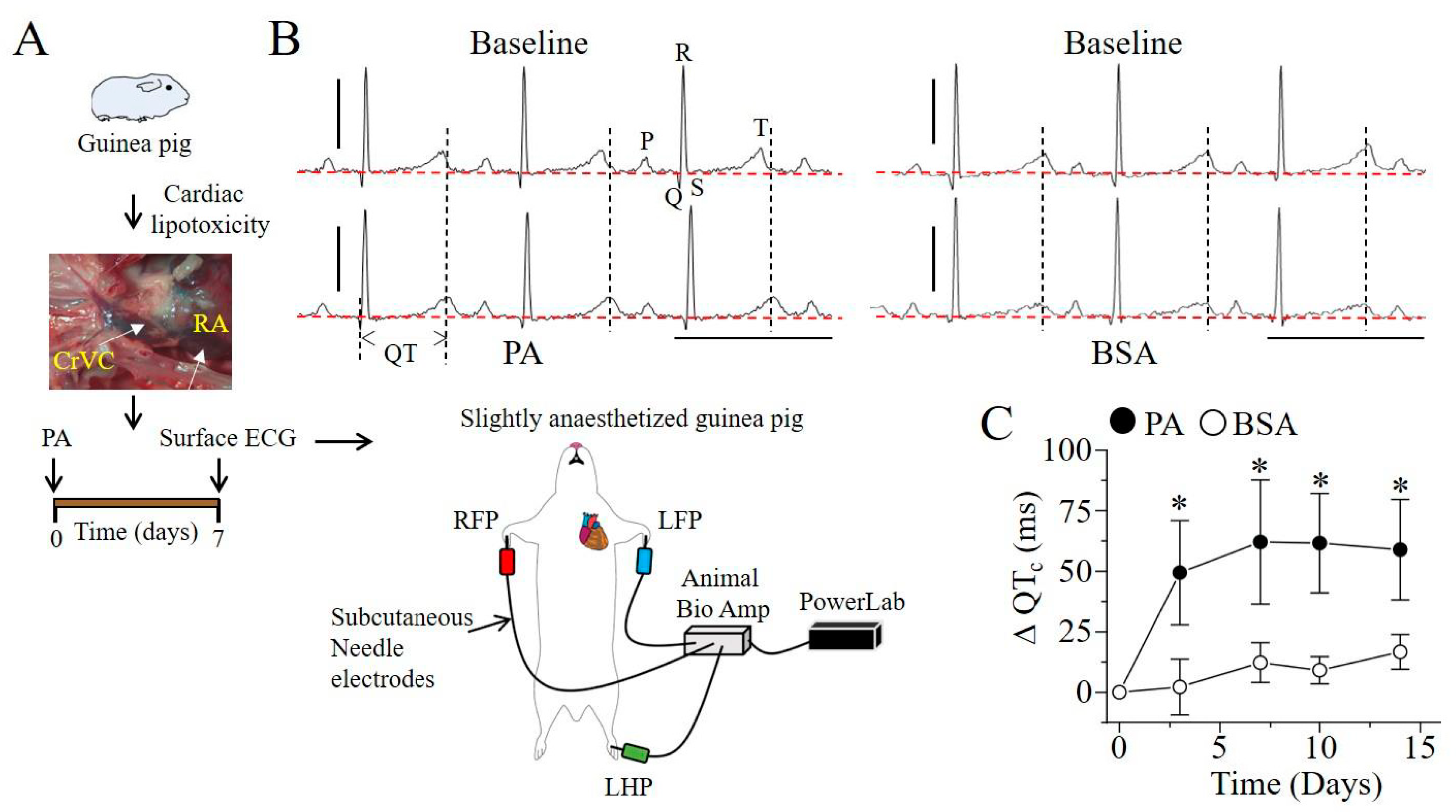
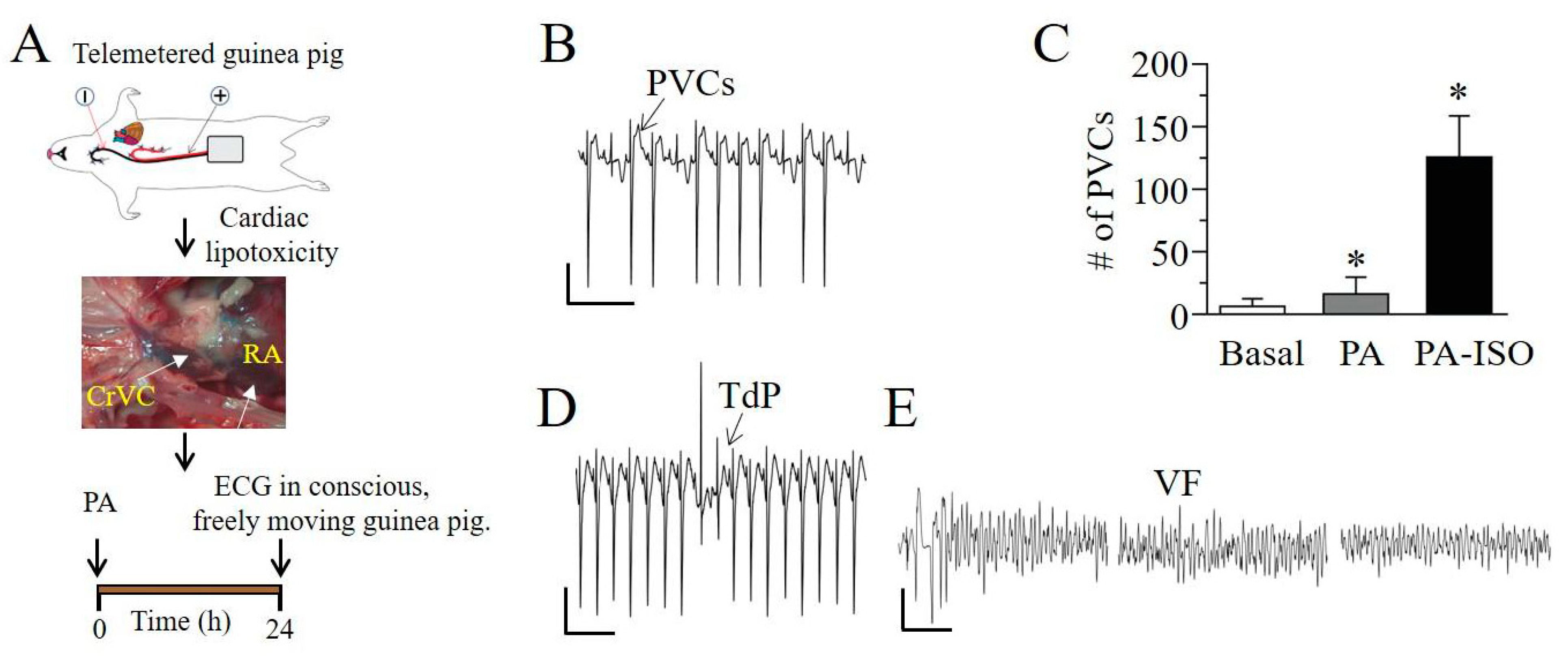
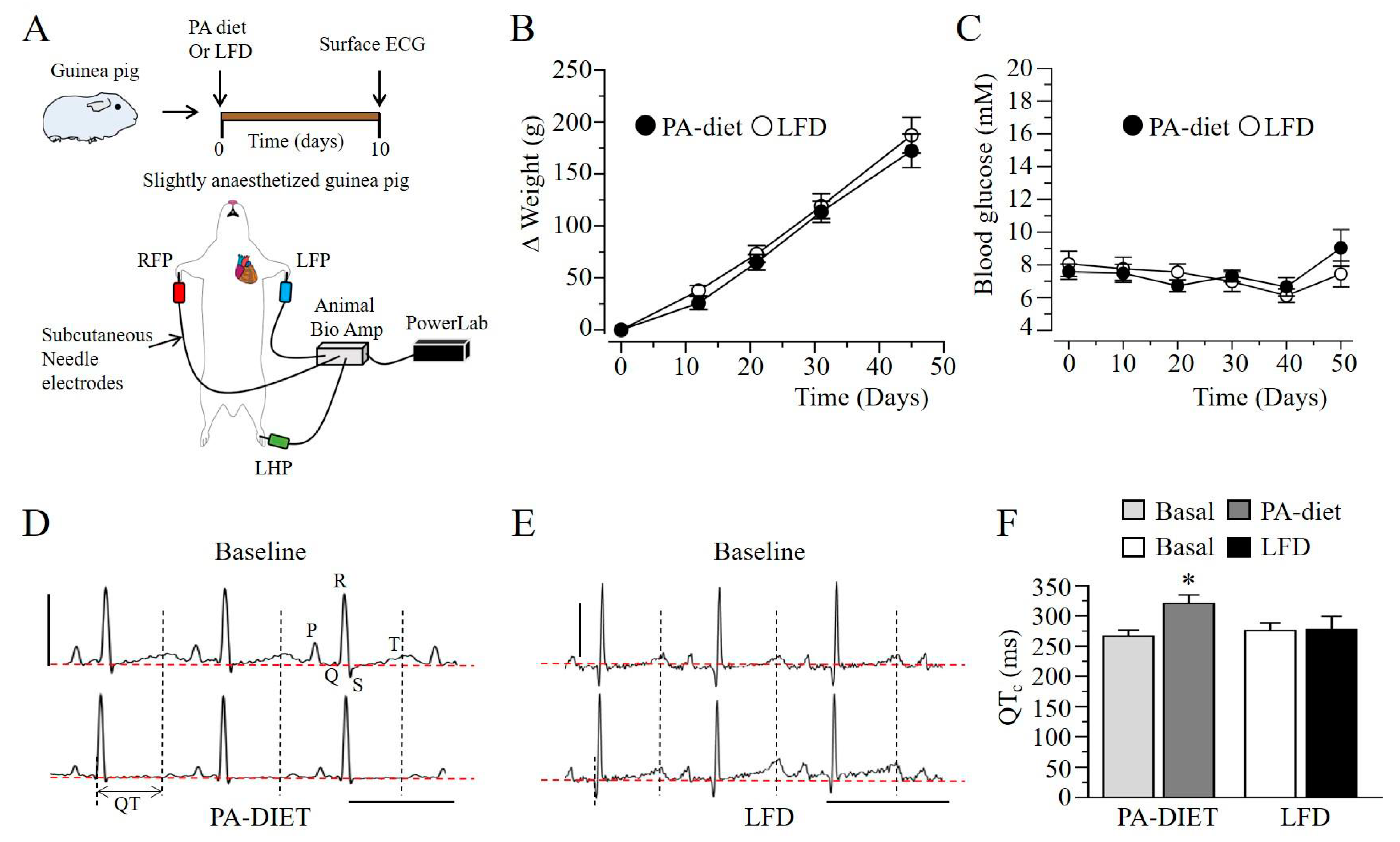
2.3. Q-T Interval Is Prolonged in Lipotoxic Hearts
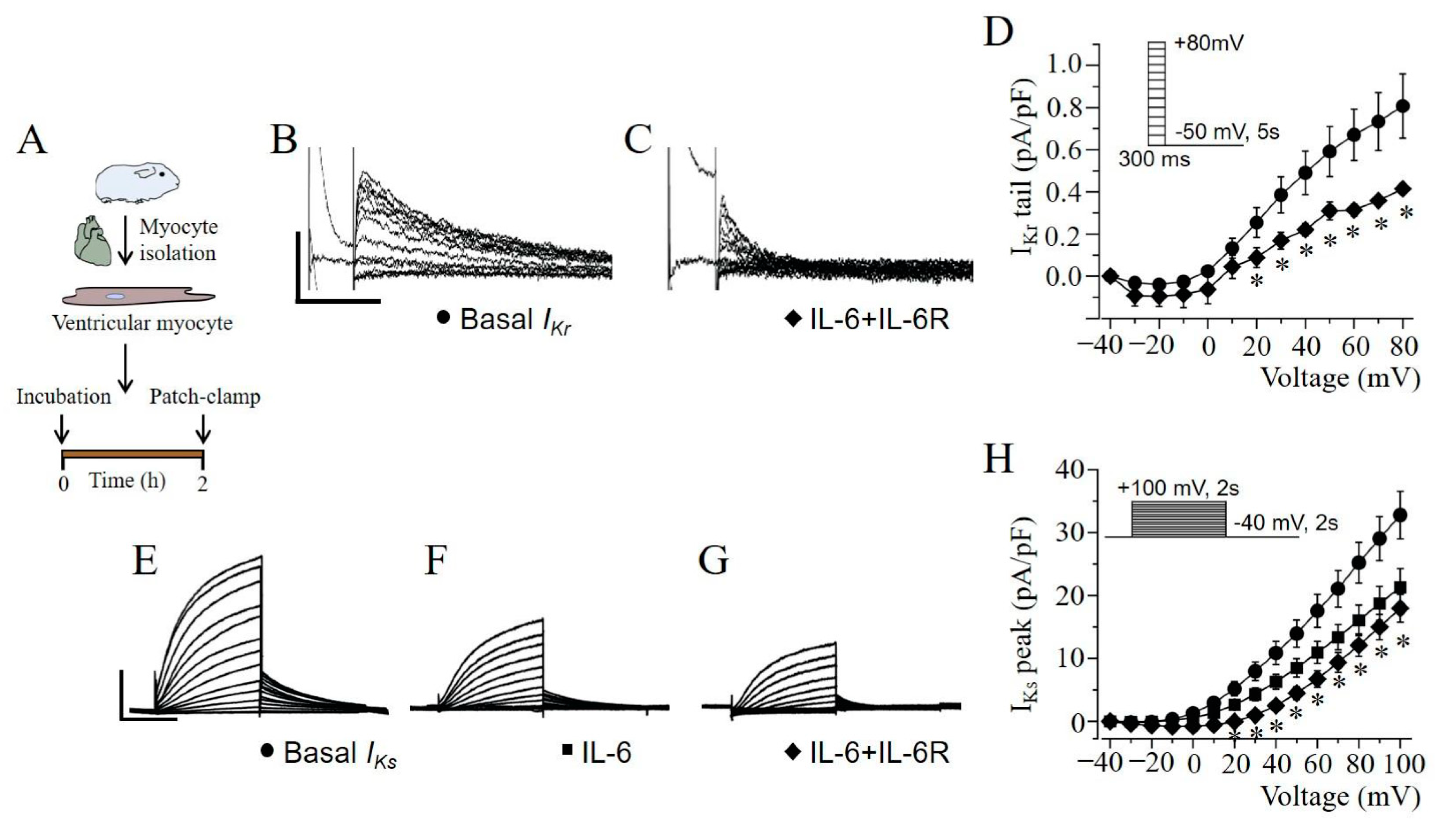
2.4. IKr and IKs Are Severely Depressed by IL-6 + IL-6R in Guinea Pig Ventricular Myocytes
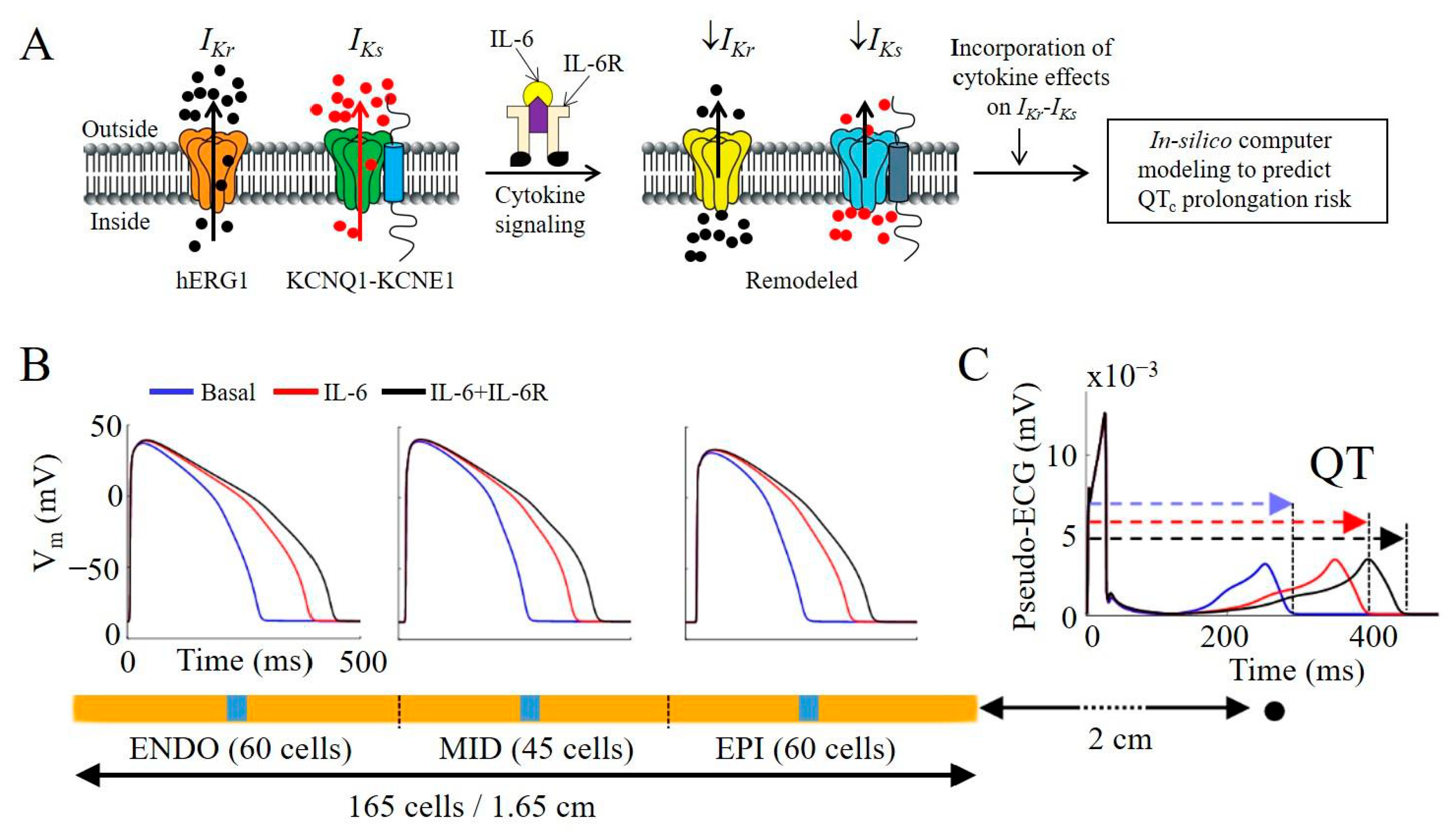
2.5. IL-6 + IL-6R Inhibition of Native IKr and IKs in Guinea Pig Ventricular Myocytes Is Proarrhythmic in a Human Ventricular Cardiomyocyte In-Silico Model
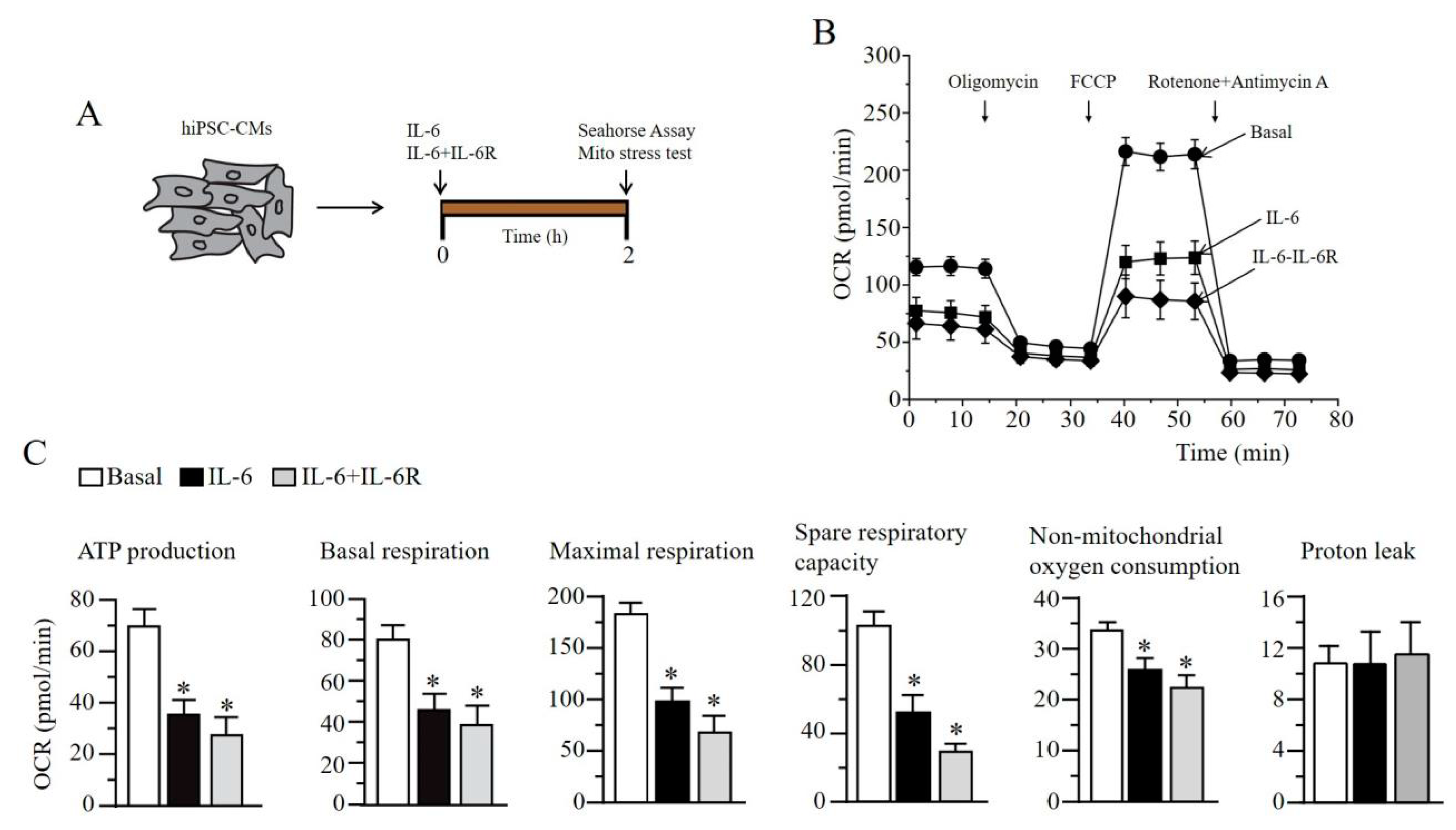
2.6. IL-6 Induces Impaired Mitochondrial Metabolism in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes
3. Discussion
4. Materials and Methods
4.1. RAW264.7 Cell Culture
4.2. Guinea Pig Ventricular Myocytes
4.3. Preparation of Bovine Serum Albumin Conjugated FFA Solutions
4.4. Low-Fat Diet and Palmitic-Acid Diet Feeding in Guinea Pig
4.5. Preparation of hiPSC-CMs
4.6. Electrophysiology
4.7. Electrocardiogram (ECG)
4.8. Telemetry
4.9. Computer Simulation
4.10. Data and Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability Statement
Acknowledgments
Conflicts of Interest
References
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arter. Thromb. Vasc. Biol. 2006, 26, 968–976. [Google Scholar] [CrossRef]
- Sonnenberg, G.E.; Krakower, G.R.; Kissebah, A.H. A novel pathway to the manifestations of metabolic syndrome. Obes. Res. 2004, 12, 180–186. [Google Scholar] [CrossRef]
- Boden, G.; She, P.; Mozzoli, M.; Cheung, P.; Gumireddy, K.; Reddy, P.; Xiang, X.; Luo, Z.; Ruderman, N. Free fatty acids produce insulin resistance and activate the proinflammatory nuclear factor-kappaB pathway in rat liver. Diabetes 2005, 54, 3458–3465. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Garg, S.; Mehndiratta, M.; Madhu, S.V.; Puri, D. Relationship of Serum Apolipoprotein A-V Levels, Oxidative Stress and Inflammatory Biomarkers with Hypertriglyceridemia in Type 2 Diabetes Mellitus. Int. J. Endocrinol. Metab. 2017, 15, e44805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matafome, P.; Seica, R. Function and Dysfunction of Adipose Tissue. Adv. Neurobiol. 2017, 19, 3–31. [Google Scholar]
- Djousse, L.; Benkeser, D.; Arnold, A.; Kizer, J.R.; Zieman, S.J.; Lemaitre, R.N.; Tracy, R.P.; Gottdiener, J.S.; Mozaffarian, D.; Siscovick, D.S.; et al. Plasma free fatty acids and risk of heart failure: The Cardiovascular Health Study. Circ. Heart Fail. 2013, 6, 964–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Boutjdir, M.; Aromolaran, A.S. Cardiolipotoxicity, Inflammation, and Arrhythmias: Role for Interleukin-6 Molecular Mechanisms. Front. Physiol. 2018, 9, 1866. [Google Scholar] [CrossRef] [PubMed]
- Aromolaran, A.S.; Srivastava, U.; Ali, A.; Chahine, M.; Lazaro, D.; El-Sherif, N.; Capecchi, P.L.; Laghi-Pasini, F.; Lazzerini, P.E.; Boutjdir, M. Interleukin-6 inhibition of hERG underlies risk for acquired long QT in cardiac and systemic inflammation. PLoS ONE 2018, 13, e0208321. [Google Scholar] [CrossRef] [Green Version]
- Monnerat, G.; Alarcon, M.L.; Vasconcellos, L.R.; Hochman-Mendez, C.; Brasil, G.; Bassani, R.A.; Casis, O.; Malan, D.; Travassos, L.H.; Sepulveda, M.; et al. Macrophage-dependent IL-1beta production induces cardiac arrhythmias in diabetic mice. Nat. Commun. 2016, 7, 13344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.H.; Rozanski, G.J. Effects of human recombinant interleukin-1 on electrical properties of guinea pig ventricular cells. Cardiovasc. Res. 1993, 27, 525–530. [Google Scholar] [CrossRef]
- Hagiwara, Y.; Miyoshi, S.; Fukuda, K.; Nishiyama, N.; Ikegami, Y.; Tanimoto, K.; Murata, M.; Takahashi, E.; Shimoda, K.; Hirano, T.; et al. SHP2-mediated signaling cascade through gp130 is essential for LIF-dependent I CaL, [Ca2+]i transient, and APD increase in cardiomyocytes. J. Mol. Cell Cardiol. 2007, 43, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Haim, T.E.; Wang, W.; Flagg, T.P.; Tones, M.A.; Bahinski, A.; Numann, R.E.; Nichols, C.G.; Nerbonne, J.M. Palmitate attenuates myocardial contractility through augmentation of repolarizing Kv currents. J. Mol. Cell Cardiol. 2010, 48, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Aromolaran, A.S.; Colecraft, H.M.; Boutjdir, M. High-fat diet-dependent modulation of the delayed rectifier K(+) current in adult guinea pig atrial myocytes. Biochem. Biophys. Res. Commun. 2016, 474, 554–559. [Google Scholar] [CrossRef]
- O’Connell, R.P.; Musa, H.; Gomez, M.S.; Avula, U.M.; Herron, T.J.; Kalifa, J.; Anumonwo, J.M. Free Fatty Acid Effects on the Atrial Myocardium: Membrane Ionic Currents Are Remodeled by the Disruption of T-Tubular Architecture. PLoS ONE 2015, 10, e0133052. [Google Scholar] [CrossRef] [Green Version]
- Aromolaran, A.S.; Subramanyam, P.; Chang, D.D.; Kobertz, W.R.; Colecraft, H.M. LQT1 mutations in KCNQ1 C-terminus assembly domain suppress IKs using different mechanisms. Cardiovasc. Res. 2014, 104, 501–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puckerin, A.; Aromolaran, K.A.; Chang, D.D.; Zukin, R.S.; Colecraft, H.M.; Boutjdir, M.; Aromolaran, A.S. hERG 1a LQT2 C-terminus truncation mutants display hERG 1b-dependent dominant negative mechanisms. Heart Rhythm. 2016, 13, 1121–1130. [Google Scholar] [CrossRef]
- O’Hara, T.; Virag, L.; Varro, A.; Rudy, Y. Simulation of the undiseased human cardiac ventricular action potential: Model formulation and experimental validation. PLoS Comput. Biol. 2011, 7, e1002061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, M.T.; Ferrero, J.M.; Romero, L.; Trenor, B. Sensitivity analysis revealing the effect of modulating ionic mechanisms on calcium dynamics in simulated human heart failure. PLoS ONE 2017, 12, e0187739. [Google Scholar]
- Dutta, S.; Chang, K.C.; Beattie, K.A.; Sheng, J.; Tran, P.N.; Wu, W.W.; Wu, M.; Strauss, D.G.; Colatsky, T.; Li, Z. Optimization of an In silico Cardiac Cell Model for Proarrhythmia Risk Assessment. Front. Physiol. 2017, 8, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilde, A.A.; Bezzina, C.R. Genetics of cardiac arrhythmias. Heart 2005, 91, 1352–1358. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, T.; Akashi, K.; Keira, N.; Matoba, S.; Mano, A.; Shiraishi, J.; Yamanaka, S.; Kobara, M.; Hibino, N.; Hosokawa, S.; et al. Cytokine-induced nitric oxide inhibits mitochondrial energy production and induces myocardial dysfunction in endotoxin-treated rat hearts. J. Mol. Cell Cardiol. 2004, 37, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, X.; Shang, W.; Xu, J.; Wang, X.; Hu, X.; Ao, Y.; Cheng, H. Proinflammatory Cytokines Stimulate Mitochondrial Superoxide Flashes in Articular Chondrocytes In Vitro and In Situ. PLoS ONE 2013, 8, e66444. [Google Scholar]
- Wegrzyn, P.; Yarwood, S.J.; Fiegler, N.; Bzowska, M.; Koj, A.; Mizgalska, D.; Malicki, S.; Pajak, M.; Kasza, A.; Kachamakova-Trojanowska, N.; et al. Mimitin—A novel cytokine-regulated mitochondrial protein. BMC Cell. Biol. 2009, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos Muniz, M.G.; Palfreeman, M.; Setzu, N.; Sanchez, M.A.; Saenz Portillo, P.; Garza, K.M.; Gosselink, K.L.; Spencer, C.T. Obesity Exacerbates the Cytokine Storm Elicited by Francisella tularensis Infection of Females and Is Associated with Increased Mortality. Biomed. Res. Int. 2018, 2018, 3412732. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Reddy, M.A.; Senapati, P.; Stapleton, K.; Lanting, L.; Wang, M.; Amaram, V.; Ganguly, R.; Zhang, L.; Devaraj, S.; et al. Diabetes Mellitus-Induced Long Noncoding RNA Dnm3os Regulates Macrophage Functions and Inflammation via Nuclear Mechanisms. Arter. Thromb. Vasc. Biol. 2018, 38, 1806–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Shi, J.; Xiao, P. Associations between common ion channel single nucleotide polymorphisms and sudden cardiac death in adults: A MOOSE-compliant meta-analysis. Medicine 2018, 97, e12428. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Nakagawa, M.; Watanabe, N.; Nakamura, M. Ubiquitin-like protein MNSFbeta covalently binds to Bcl-G and enhances lipopolysaccharide/interferon gamma-induced apoptosis in macrophages. FEBS J. 2013, 280, 1281–1293. [Google Scholar] [CrossRef]
- Jost, N.; Virág, L.; Bitay, M.; Takács, J.; Lengyel, C.; Biliczki, P.; Nagy, Z.; Bogáts, G.; Lathrop, D.A.; Papp, J.G.; et al. Restricting excessive cardiac action potential and QT prolongation: A vital role for IKs in human ventricular muscle. Circulation 2005, 112, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- Pisoni, C.N.; Reina, S.; Arakaki, D.; Eimon, A.; Carrizo, C.; Borda, E. Elevated IL-1beta levels in anti-Ro/SSA connective tissue diseases patients with prolonged corrected QTc interval. Clin. Exp. Rheumatol. 2015, 33, 715–720. [Google Scholar] [PubMed]
- Lazzerini, P.E.; Acampa, M.; Capecchi, P.L.; Fineschi, I.; Selvi, E.; Moscadelli, V.; Zimbone, S.; Gentile, D.; Galeazzi, M.; Laghi-Pasini, F. Antiarrhythmic potential of anticytokine therapy in rheumatoid arthritis: Tocilizumab reduces corrected QT interval by controlling systemic inflammation. Arthritis Care Res. 2015, 67, 332–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzerini, P.E.; Capecchi, P.L.; Laghi-Pasini, F. Long QT Syndrome: An Emerging Role for Inflammation and Immunity. Front. Cardiovasc. Med. 2015, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, H.; Zhang, Y.; Gao, H.; Nattel, S.; Wang, Z. Impairment of HERG K(+) channel function by tumor necrosis factor-alpha: Role of reactive oxygen species as a mediator. J. Biol. Chem. 2004, 279, 13289–13292. [Google Scholar] [CrossRef] [Green Version]
- Kawada, H.; Niwano, S.; Niwano, H.; Yumoto, Y.; Wakisaka, Y.; Yuge, M.; Kawahara, K.; Izumi, T. Tumor necrosis factor-alpha downregulates the voltage gated outward K+ current in cultured neonatal rat cardiomyocytes: A possible cause of electrical remodeling in diseased hearts. Circ. J. 2006, 70, 605–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkova-Kirova, P.S.; Gursoy, E.; Mehdi, H.; McTiernan, C.F.; London, B.; Salama, G. Electrical remodeling of cardiac myocytes from mice with heart failure due to the overexpression of tumor necrosis factor-alpha. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H2098–H2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Velasco, M.; Ruiz-Hurtado, G.; Hurtado, O.; Moro, M.A.; Delgado, C. TNF-alpha downregulates transient outward potassium current in rat ventricular myocytes through iNOS overexpression and oxidant species generation. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H238–H245. [Google Scholar] [CrossRef] [Green Version]
- Marx, S.O.; Kurokawa, J.; Reiken, S.; Motoike, H.; D’Armiento, J.; Marks, A.R.; Kass, R.S. Requirement of a macromolecular signaling complex for beta adrenergic receptor modulation of the KCNQ1-KCNE1 potassium channel. Science 2002, 295, 496–499. [Google Scholar]
- Nicolas, C.S.; Park, K.-H.; El Harchi, A.; Camonis, J.; Kass, R.S.; Escande, D.; Mérot, J.; Loussouarn, G.; Le Bouffant, F.; Baró, I. IKs response to protein kinase A-dependent KCNQ1 phosphorylation requires direct interaction with microtubules. Cardiovasc. Res. 2008, 79, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alseikhan, B.A.; DeMaria, C.D.; Colecraft, H.M.; Yue, D.T. Engineered calmodulins reveal the unexpected eminence of Ca2+ channel inactivation in controlling heart excitation. Proc. Natl. Acad. Sci. USA 2002, 99, 17185–17190. [Google Scholar] [CrossRef] [Green Version]
- Subramanyam, P.; Chang, D.D.; Fang, K.; Xie, W.; Marks, A.R.; Colecraft, H.M. Manipulating L-type calcium channels in cardiomyocytes using split-intein protein transsplicing. Proc. Natl. Acad. Sci. USA 2013, 110, 15461–15466. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Mateu, L.; Saiz, J.; Aromolaran, A.S. Differential Modulation of IK and ICa,L Channels in High-Fat Diet-Induced Obese Guinea Pig Atria. Front. Physiol. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, E.A.; Ferrero, J.M.; Doblare, M.; Rodriguez, J.F. Adaptive macro finite elements for the numerical solution of monodomain equations in cardiac electrophysiology. Ann. Biomed. Eng. 2010, 38, 2331–2345. [Google Scholar] [CrossRef]
- Gomez, J.F.; Cardona, K.; Romero, L.; Ferrero, J.M., Jr.; Trenor, B. Electrophysiological and structural remodeling in heart failure modulate arrhythmogenesis. 1D simulation study. PLoS ONE 2014, 9, e106602. [Google Scholar] [CrossRef]
- Saiz, J.; Gomis-Tena, J.; Monserrat, M.; Ferrero, J.M., Jr.; Cardona, K.; Chorro, J. Effects of the antiarrhythmic drug dofetilide on transmural dispersion of repolarization in ventriculum. A computer modeling study. IEEE Trans. Biomed. Eng. 2011, 58, 43–53. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTc (ms) | Heart Rate (BPM) | PR Interval (ms) | QRS (ms) | |
|---|---|---|---|---|
| Baseline | 244 ± 22 | 280 ± 10.5 | 53.5 ± 1.68 | 16.6 ± 0.85 |
| PA- BSA | 306.7 ± 6.5 * | 279 ± 4.89 | 56.9 ± 1.78 | 17.6 ± 0.25 |
| Baseline | 271 ± 8.23 | 281.3 ± 17.6 | 54.3 ± 1.68 | 16.4 ± 0.86 |
| BSA | 283.4 ± 6.21 | 281.3 ± 6.19 | 54.7 ± 2.49 | 17.3 ± 0.50 |
| Currents | Basal | +IL-6 | +IL-6-IL-6R |
|---|---|---|---|
| IKr | 1 | 0.54 | 0.42 |
| IKs | 1 | 0.65 | 0.55 |
| Basal | IL-6 IKr Block | IL-6 IKs Block | IL-6 IKr + IKs Block | |
| IKr | 1 | 0.54 | 1 | 0.54 |
| IKs | 1 | 1 | 0.65 | 0.65 |
| APD90 (ms) | 262 | 359 | 270 | 380 |
| ΔAPD90 (%) | 37% | 3% | 45% | |
| Basal | IL-6-IL-6R IKr Block | IL-6-IL-6R IKs Block | IL-6-IL-6R IKr + IKs Block | |
| IKr | 1 | 0.42 | 1 | 0.42 |
| IKs | 1 | 1 | 0.55 | 0.55 |
| APD90 (ms) | 262 | 398 | 273 | 431 |
| ΔAPD90 (%) | 52% | 4% | 64% |
| Simulations | Parameters | Basal | IL-6 | IL-6-IL-6R |
|---|---|---|---|---|
| Unicellular | APD90 (ms) | 264 | 384 | 437 |
| APD prolongation (%) | +45% | +65% | ||
| Strand | QT (ms) | 297 | 405 | 454 |
| QT prolongation (%) | +36% | +53% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chowdhury, M.K.H.; Martinez-Mateu, L.; Do, J.; Aromolaran, K.A.; Saiz, J.; Aromolaran, A.S. Macrophage-Dependent Interleukin-6-Production and Inhibition of IK Contributes to Acquired QT Prolongation in Lipotoxic Guinea Pig Heart. Int. J. Mol. Sci. 2021, 22, 11249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011249
Chowdhury MKH, Martinez-Mateu L, Do J, Aromolaran KA, Saiz J, Aromolaran AS. Macrophage-Dependent Interleukin-6-Production and Inhibition of IK Contributes to Acquired QT Prolongation in Lipotoxic Guinea Pig Heart. International Journal of Molecular Sciences. 2021; 22(20):11249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011249
Chicago/Turabian StyleChowdhury, Md. Kamrul Hasan, Laura Martinez-Mateu, Jenny Do, Kelly A. Aromolaran, Javier Saiz, and Ademuyiwa S. Aromolaran. 2021. "Macrophage-Dependent Interleukin-6-Production and Inhibition of IK Contributes to Acquired QT Prolongation in Lipotoxic Guinea Pig Heart" International Journal of Molecular Sciences 22, no. 20: 11249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011249