
Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
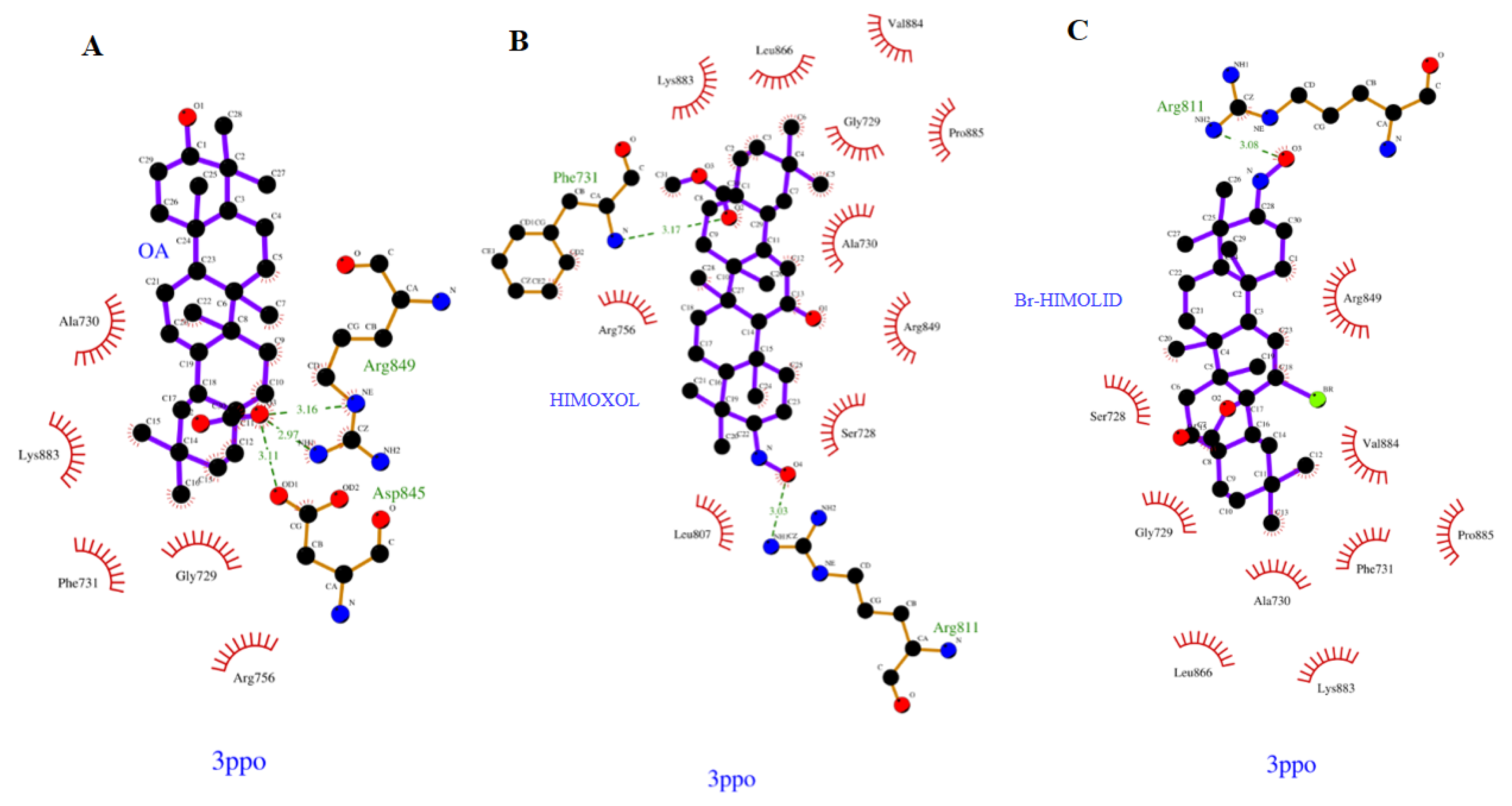
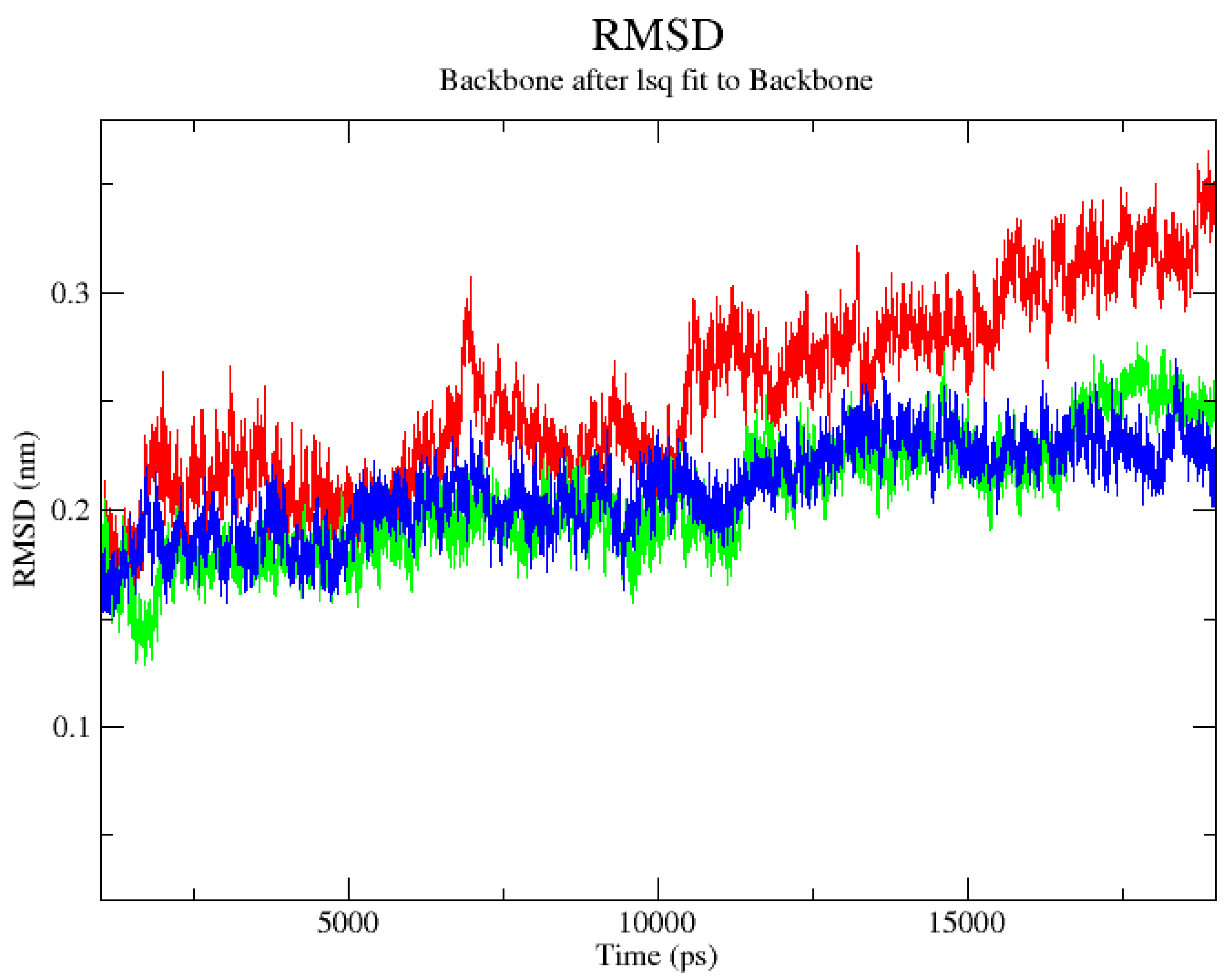
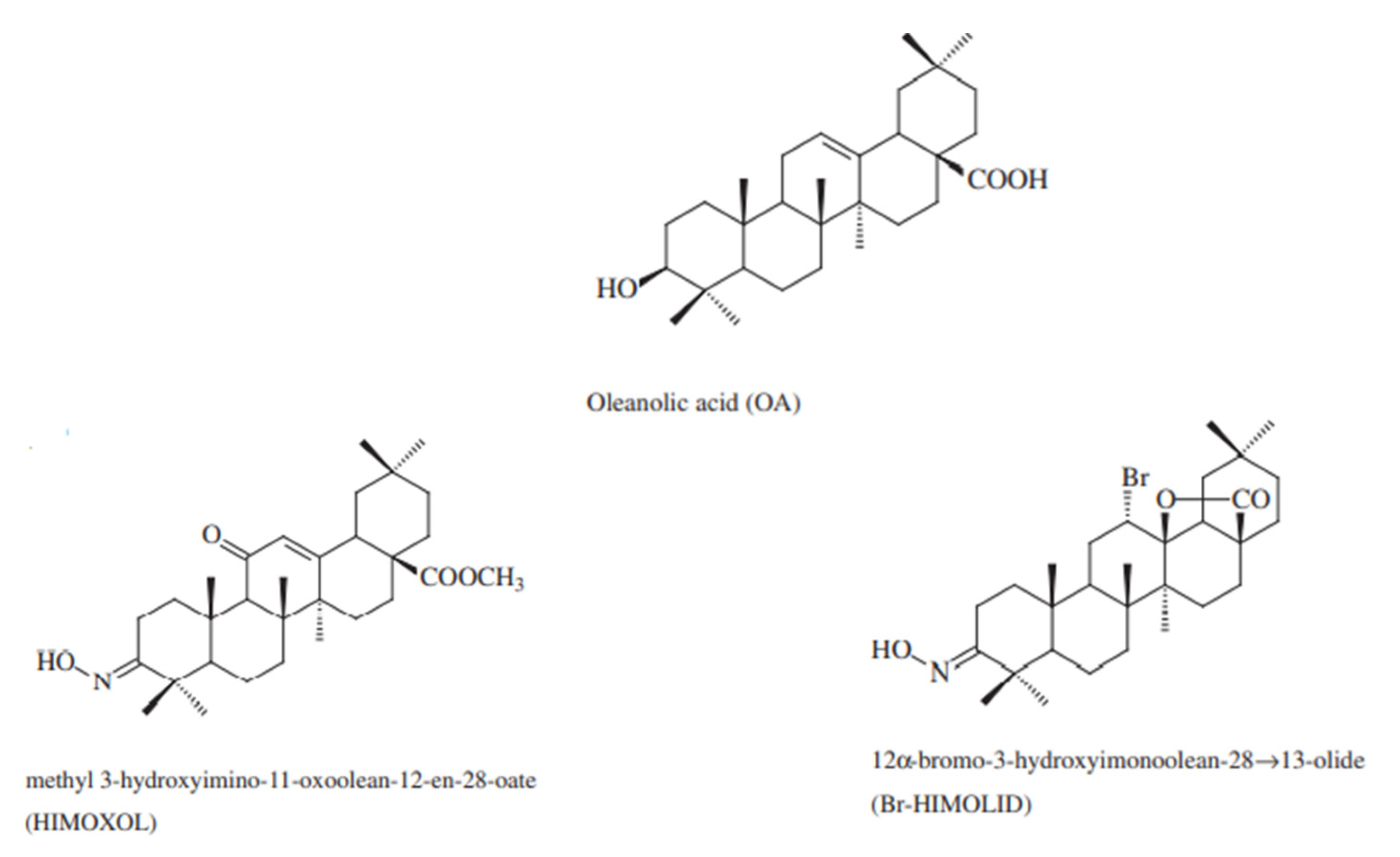
2.1. Computational Analysis
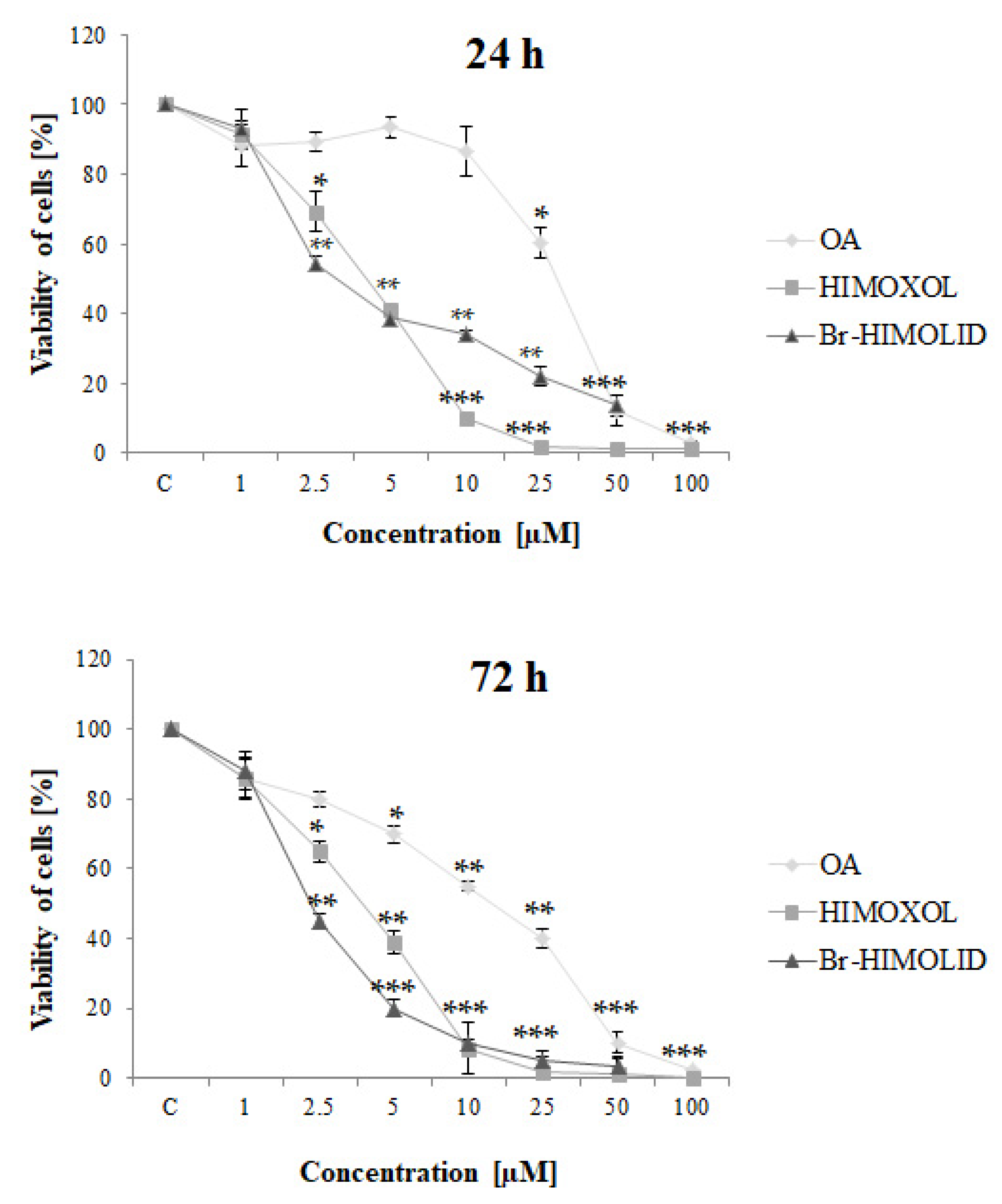
2.2. Effects of OA and Its Derivatives on Breast Cancer Cells Viability—MTT Assay
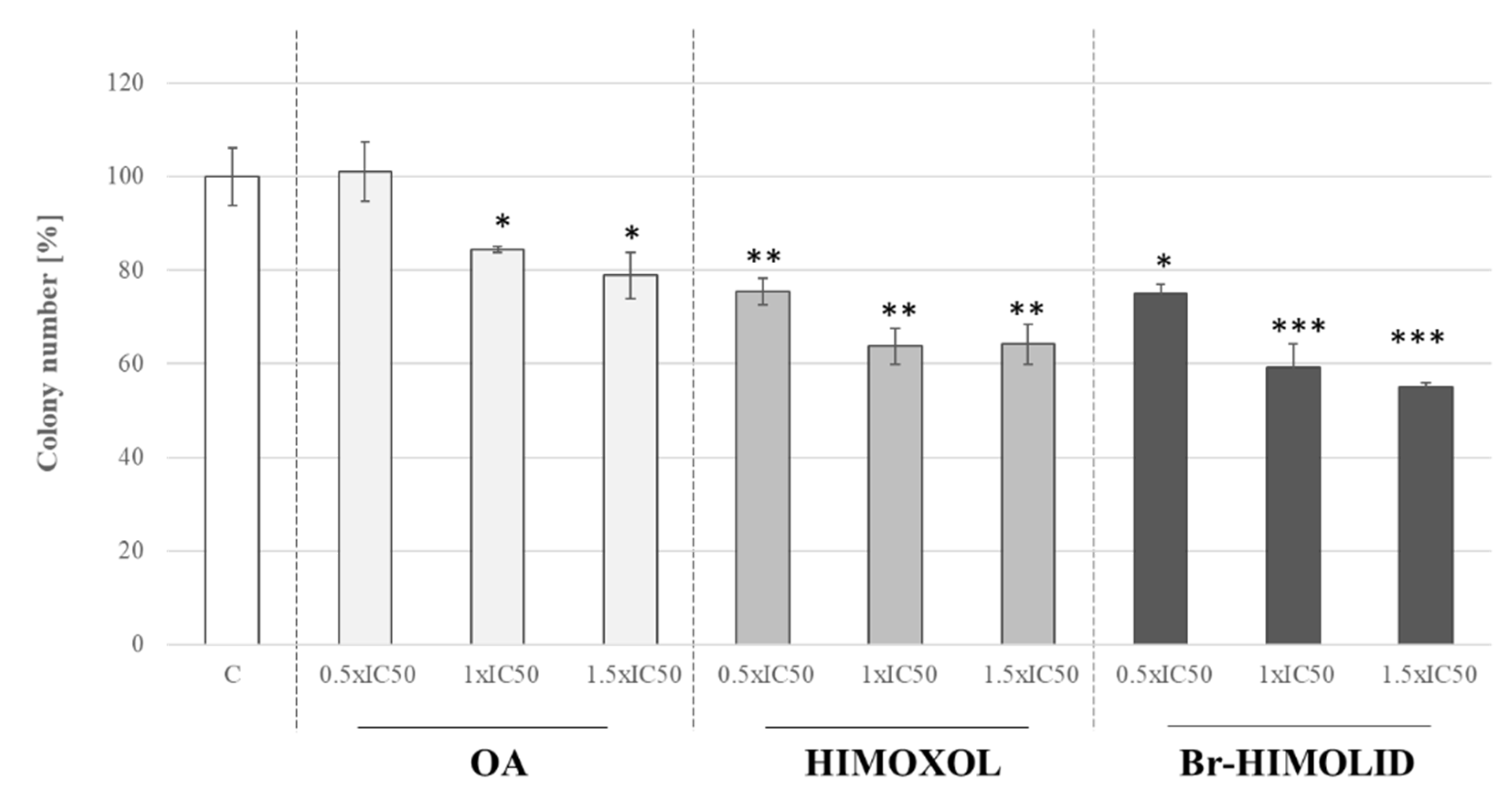
2.3. Influence of OA and Its Derivatives on the Colony Formation of SK-BR-3 Cells
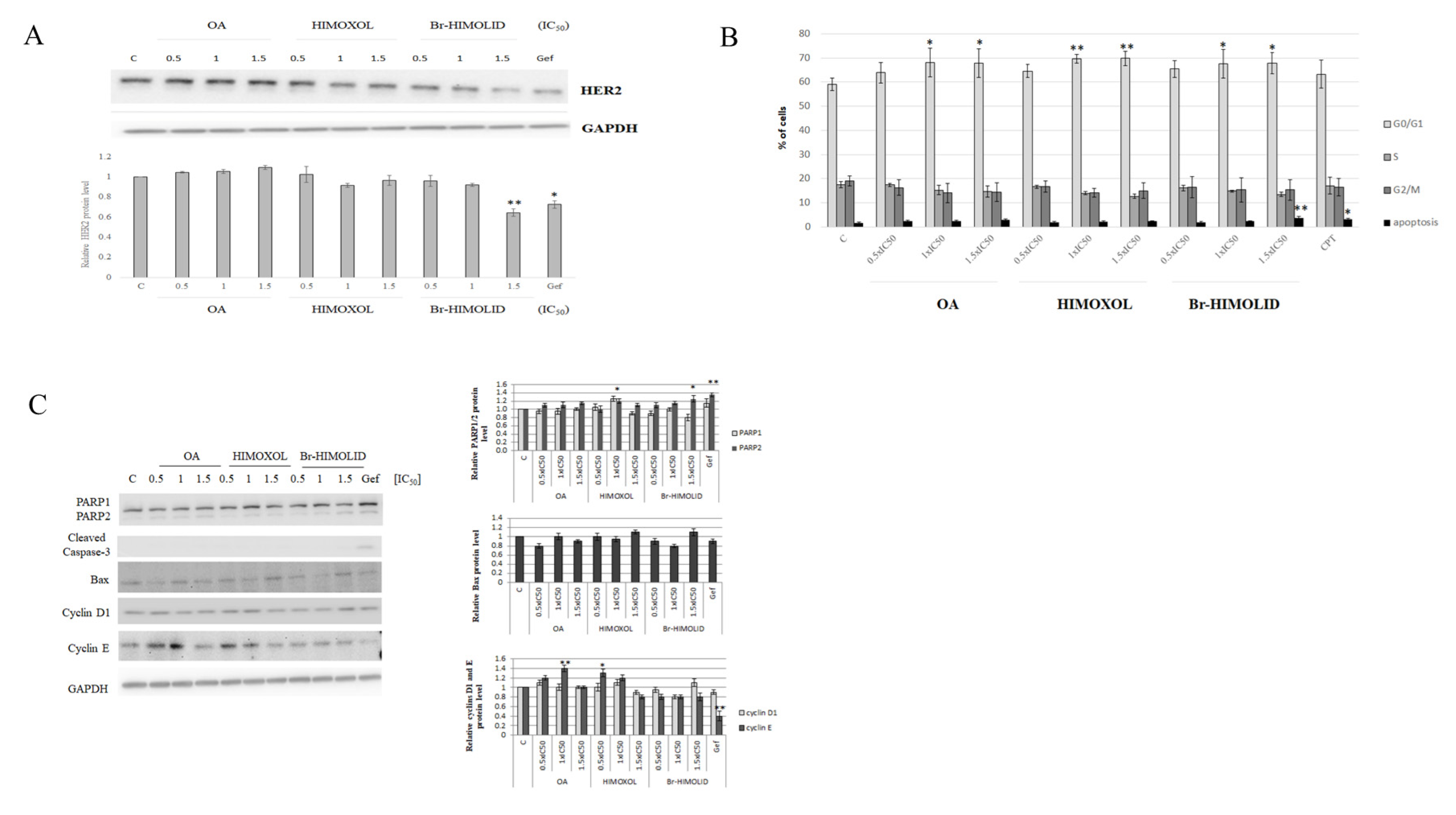
2.4. The Effect of OA and OA Derivatives on the Basal Level of HER2 and Cell Cycle in SK-BR-3 Breast Cancer Cells
2.5. Verification of Cell Cycle Modulation and Apoptosis Induction by Br-HIMOLID
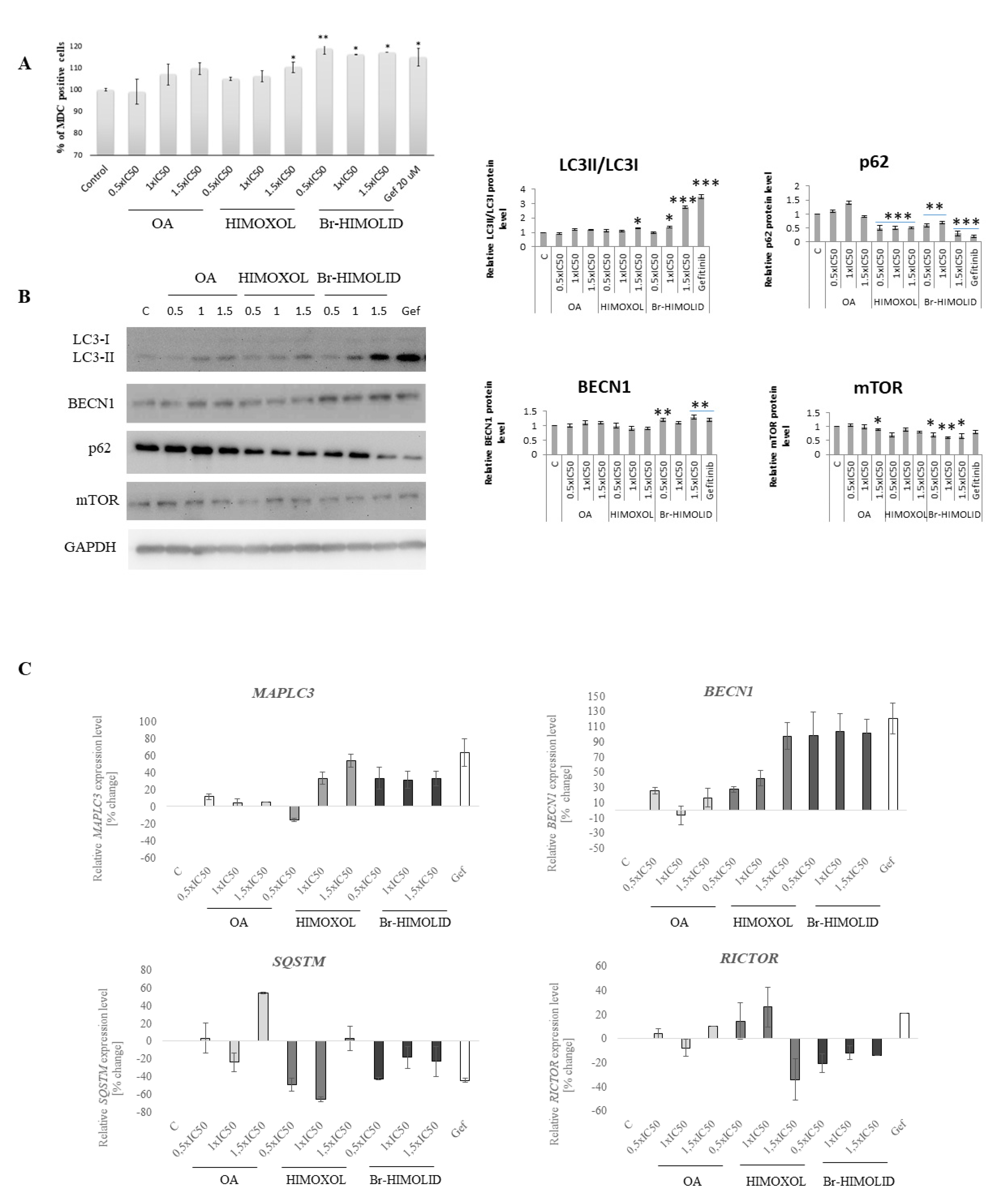
2.6. Evaluation of Autophagosomes Formation and Verification of Autophagy Pathways
2.7. Ability of Studied Semi-Synthetic Derivatives for Induction of Autophagy
2.8. OA and Its Derivatives Influence Migration of SK-BR-3—Wound Healing Assay
2.9. Verification of Pathway Engaged in Anti-Migratory Potential
3. Discussion
3.1. OA Derivatives and HER2-Positive Cancer Cells Survival
3.2. Balancing on the Edge
3.3. The Link between Adhesion and HER2 Pathway
3.4. Summary and Perspectives
4. Materials and Methods
4.1. Compounds and Reagents
4.2. Computational Details
4.3. Cell Line and Cell Culture
4.4. Viability Assay
4.5. Colony-Forming Assay
4.6. Cell Cycle Analysis by Flow Cytometry
4.7. Detection of Autophagy by Flow Cytometry
4.8. Wound Healing Assay
4.9. Quantitative Gene Expression Assessment
4.10. Immunodetection
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer Targets Ther. 2019, 11, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef]
- Howe, L.R.; Brown, P.H. Targeting the HER/EGFR/ErbB family to prevent breast cancer. Cancer Prev. Res. 2011, 4, 1149–1157. [Google Scholar] [CrossRef] [Green Version]
- Miligy, I.M.; Toss, M.S.; Gorringe, K.L.; Lee, A.H.S.; Ellis, I.O.; Green, A.R.; Rakha, E.A. The clinical and biological significance of HER2 overexpression in breast ductal carcinoma in situ: A large study from a single institution. Br. J. Cancer 2019, 120, 1075–1082. [Google Scholar] [CrossRef]
- Ishii, K.; Morii, N.; Yamashiro, H. Pertuzumab in the treatment of HER2-positive breast cancer: An evidence-based review of its safety, efficacy, and place in therapy. Core Evid. 2019, 14, 51–70. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Liu, X.; Lee, E.S.; Sun, J.; Feng, Z.; Zhao, L.; Zhao, C. Synthesis of novel oleanolic acid and ursolic acid in C-28 position derivatives as potential anticancer agents. Arch. Pharm. Res. 2017, 40, 458–468. [Google Scholar] [CrossRef]
- Bishayee, A.; Ahmed, S.; Brankov, N.; Perloff, M. Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Front. Biosci. 2011, 1, 980–996. [Google Scholar] [CrossRef] [Green Version]
- Lisiak, N.; Paszel-Jaworska, A.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Kaczmarek, M.; Rybczyńska, M. Methyl 3-hydroxyimino-11-oxoolean-12-en-28-oate (HIMOXOL), a synthetic oleanolic acid derivative, induces both apoptosis and autophagy in MDA-MB-231 breast cancer cells. Chem. Biol. Interact. 2014, 208, 47–57. [Google Scholar] [CrossRef]
- Lisiak, N.; Toton, E.; Rubis, B.; Majer, B.; Rybczynska, M. The Synthetic Oleanane Triterpenoid HIMOXOL Induces Autophagy in Breast Cancer Cells via ERK1/2 MAPK Pathway and Beclin-1 Up-regulation. Anticancer Agents Med. Chem. 2016, 16, 1066–1076. [Google Scholar] [CrossRef]
- Konopleva, M.; Zhang, W.; Shi, Y.X.; McQueen, T.; Tsao, T.; Abdelrahim, M.; Munsell, M.F.; Johansen, M.; Yu, D.; Madden, T.; et al. Synthetic triterpenoid 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid induces growth arrest in HER2-overexpressing breast cancer cells. Mol. Cancer Ther. 2006, 5, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Ma, R.; Boyle, P.J.; Mikulla, B.; Bradley, M.; Smith, B.; Basu, M.; Banerjee, S.; Glycoconj, J. Apoptosis of human carcinoma cells in the presence of potential anti-cancer drugs: III. Treatment of Colo-205 and SKBR3 cells with: Cisplatin, Tamoxifen, Melphalan, Betulinic acid, L-PDMP, L-PPMP, and GD3 ganglioside. Glycoconj. J. 2004, 20, 563–577. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, M.S. Autophagy- a key player in cellular and body metabolism. Nat. Rev. Endocrinol. 2014, 10, 322–337. [Google Scholar] [CrossRef]
- Patra, S.; Mishra, S.R.; Behera, B.P.; Mahapatra, K.K.; Panigrahi, D.P.; Bhol, C.S.; Praharaj, P.P.; Sethi, G.; Patra, S.K.; Bhutia, S.K. Autophagy-modulating phytochemicals in cancer therapeutics: Current evidences and future perspectives. Semin. Cancer Biol. 2020. (In press) [Google Scholar] [CrossRef] [PubMed]
- Lisiak, N.; Paszel-Jaworska, A.; Totoń, E.; Rubiś, B.; Pakuła, M.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rybczyńska, M. Semisynthetic oleanane triterpenoids inhibit migration and invasion of human breast cancer cells through downregulated expression of the ITGB1/PTK2/PXN pathway. Chem. Biol. Interact. 2017, 268, 136–147. [Google Scholar] [CrossRef]
- Guan, X. Cancer metastases: Challenges and opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žiberna, L.; Šamec, D.; Mocan, A.; Nabavi, S.F.; Bishayee, A.; Farooqi, A.A.; Sureda, A.; Nabavi, S.M. Oleanolic Acid Alters Multiple Cell Signaling Pathways: Implication in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2017, 18, 643. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, M.K.; Xiaoyun Dai, X.; Kumar, A.P.; Tan, B.K.H.; Sethi, G.; Bishayee, A. Oleanolic acid and its synthetic derivatives for the prevention and therapy of cancer: Preclinical and clinical evidence. Cancer Lett. 2014, 346, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Aertgeerts, K.; Skene, R.; Yano, J.; Sang, B.C.; Zou, H.; Snell, G.; Jennings, A.; Iwamoto, K.; Habuka, N.; Hirokawa, A.; et al. Structural Analysis of the Mechanism of Inhibition and Allosteric Activation of the Kinase Domain of HER2 Protein. J. Biol. Chem. 2011, 286, 18756–18765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RCSB PDB—3PPO: Crystal Structure of the Kinase Domain of Human HER2 (erbB2). Available online: https://www.rcsb.org/structure/43PPO (accessed on 16 April 2021).
- Gaussian 16 Rev. C.01 Release Notes. Available online: https://gaussian.com/relnotes/ (accessed on 15 April 2021).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. Ligplot: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Arianingrum, R.; Hermawan, A.; Purnomo, H.; Dewi, D.; Meiy, E. Molecular Docking Studies of a Chalcone Derivative Compound p-hydroxy-m-methoxychalcone with Tyrosine Kinase Receptors. Indian J. Public Health Res. Dev. 2019, 10, 1219–1224. [Google Scholar] [CrossRef]
- Parrish, R.M.; Burns, L.A.; Smith, D.G.A.; Simmonett, A.C.; DePrince, A.E.; Hohenstein, E.G.; Bozkaya, U.; Sokolov, A.Y.; Di Remigio, R.; Richard, R.M.; et al. Psi4 1.1: An Open-Source Electronic Structure Program Emphasizing Automation, Advanced Libraries, and Interoperability. J. Chem. Theory Comput. 2017, 13, 3185–3197. [Google Scholar] [CrossRef]
- Franken, N.A.P.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Biederbick, A.; Kern, H.F.; Elsässer, H.P. Monodansylcadaverine (MDC) is a specific in vivo marker for autophagic vacuoles. Eur. J. Cell Biol. 1995, 66, 3–14. [Google Scholar] [PubMed]
- Oprean, C.; Mioc, M.; Csányi, E.; Ambrus, R.; Bojin, F.; Tatu, C.; Cristea, M.; Ivan, A.; Danciu, C.; Dehelean, C.; et al. Improvement of ursolic and oleanolic acids’ antitumor activity by complexation with hydrophilic cyclodextrins. Biomed. Pharmacother. 2016, 83, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.M.; da Silva, D.L.; Modolo, L.V.; Alves, R.B.; de Resende, M.A.; Martins, C.V.B.; de Angelo, F. Schiff bases: A short review of their antimicrobial activities. J. Adv. Res. 2011, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Malheiro, J.; Gomes, I.; Borges, A.; Bastos, M.M.; Maillard, J.Y.; Borges, F.; Simões, M. Biofouling. Phytochemical profiling as a solution to palliate disinfectant limitations. Biofouling 2016, 32, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca, M.N.; Vara, J.; Milla, L.; Rivarola, V.; Ortiz, C.S. Physicochemical properties and photodynamic activity of novel derivatives of triarylmethane and thiazine. Arch. Pharm. 2013, 346, 255–265. [Google Scholar] [CrossRef]
- Ndolo, R.A.; Jacobs, D.T.; Forrest, M.T.; Krise, J.P. Intracellular distribution-based anticancer drug targeting: Exploiting a lysosomal acidification defect associated with cancer cells. Mol. Cell Pharmacol. 2010, 2, 131–136. [Google Scholar]
- Kim, G.J.; Jo, H.J.; Lee, K.J.; Choi, J.W.; Jeung, H.A. Oleanolic acid induces p53-dependent apoptosis via the ERK/JNK/AKT pathway in cancer cell lines in prostatic cancer xenografts in mice. Oncotarget 2018, 9, 26370–26386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.R.H. HER2 expression is a strong independent predictor of nodal metastasis in breast cancer. J. Egypt Natl. Canc. Inst. 2016, 28, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, L.; Lim, M.; Ward, C.; Featherstone, H.; Murray, H.; D’Arcy, C.; Crown, J.; Gullo, G. Metastatic HER2+ Breast Cancer: A Potentially Curable Disease? Cureus 2017, 9, e1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, G.; Elbadawi, M.; Efferth, T. Multiple cell death modalities and their key features (Review). World Acad. Sci. J. 2020, 2, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Song, Q.; Hu, D.; Zhuang, X.; Yu, S.; Teng, D. Oleanolic acid induced autophagic cell death in hepatocellular carcinoma cells via PI3K/Akt/mTOR and ROS-dependent pathway. Korean J. Physiol. Pharmacol. 2016, 20, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, B. Targeted therapeutic options and future perspectives for HER2-positive breast cancer. Signal Transduct. Target. Ther. 2019, 4, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef] [PubMed]
- Maycotte, P.; Thorburn, A. Targeting autophagy in breast cancer. World J. Clin. Oncol. 2014, 5, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Lisiak, N.; Toton, E.; Rybczynska, M. Autophagy as a Potential Therapeutic Target in Breast Cancer Treatment. Curr. Cancer Drug Targets 2018, 18, 629–639. [Google Scholar] [CrossRef]
- Castrejón-Jiménez, N.S.; Leyva-Paredes, K.; Baltierra-Uribe, S.L.; Castillo-Cruz, J.; Campillo-Navarro, M.; Hernández-Pérez, A.D.; Luna-Angulo, A.B.; Chacón-Salinas, R.; Coral-Vázquez, R.M.; Estrada-García, I.; et al. Ursolic and Oleanolic Acids Induce Mitophagy in A549 Human Lung Cancer Cells. Molecules 2019, 24, 3444. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Lin, M.Z.; Cheng, D.; Braet, F.; Pollock, C.A.; Chen, X.M. Thioredoxin-interacting protein mediates dysfunction of tubular autophagy in diabetic kidneys through inhibiting autophagic flux. Lab Investig. 2014, 94, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar]
- Ortega, M.A.; Fraile-Martínez, O.; Asúnsolo, Á.; Buján, J.; García-Honduvilla, N.; Coca, S. Signal Transduction Pathways in Breast Cancer: The Important Role of PI3K/Akt/mTOR. J. Oncol. 2020, 2020, 9258396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, A.N.; Kwak, Y.; Kim, D.W.; Kang, S.B.; Choe, G.; Kim, W.H.; Lee, H.S. HER2 status in colorectal cancer: Its clinical significance and the relationship between HER2 gene amplification and expression. PLoS ONE 2014, 9, e98528. [Google Scholar] [CrossRef] [PubMed]
- Scales, T.; Parsons, M. Spatial and temporal regulation of integrin signalling during cell migration. Curr. Opin. Cell Biol. 2011, 23, 562–568. [Google Scholar] [CrossRef]
- Esposito, M.; Kang, Y. Targeting tumor-stromal interactions in bone metastasis. Pharmacol Ther. 2014, 141, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Jenning, S.; Pham, T.; Ireland, S.Y.; Ruoslahti, E.; Biliran, H. Bit1 in anoikis resistance and tumor metastasis. Cancer Lett. 2013, 333, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Westhoff, M.A.; Serrels, B.; Fincham, V.J.; Frame, M.C. Src-Mediated Phosphorylation of Focal Adhesion Kinase Couples Actin and Adhesion Dynamics to Survival Signaling. Mol. Cell Biol. 2004, 24, 8113–8133. [Google Scholar] [CrossRef] [Green Version]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis molecular pathways and its role in cancer progression. Biochim. Biophys. Acta 2013, 1833, 3481–3498. [Google Scholar] [CrossRef] [Green Version]
- Tai, Y.L.; Chen, L.C.; Shen, T.L. Emerging roles of focal adhesion kinase in cancer. Biomed. Res. Int. 2015, 2015, 690690. [Google Scholar] [CrossRef]
- Cruz da Silva, E.; Dontenwill, M.; Choulier, L.; Lehmann, M. Role of Integrins in Resistance to Therapies Targeting Growth Factor Receptors in Cancer. Cancers 2019, 11, 692. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.O.; Shin, S.; Lipscomb, E.A. A novel mechanism for integrin-mediated ras activation in breast carcinoma cells: The alpha6beta4 integrin regulates ErbB2 translation and transactivates epidermal growth factor receptor/ErbB2 signaling. Cancer Res. 2006, 66, 2732–2739. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Zhou, Z.; Chen, X.; Zhao, R.; Yang, Z.; Wei, N.; Ni, Q.; Feng, Y.; Yu, X.; Ma, J.; et al. HER2 reduces breast cancer radiosensitivity by activating focal adhesion kinase in vitro and in vivo. Oncotarget 2016, 7, 45186–45198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Pylayeva, Y.; Pepe, A.; Yoshioka, T.; Muller, W.J.; Inghirami, G.; Giancotti, F.G. Beta 4 integrin amplifies ErbB2 signaling to promote mammary tumorigenesis. Cell 2006, 126, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.E.; Xiang, B.; Zent, R.; Quaranta, V.; Pozzi, A.; Artega, C.L. Transforming growth factor beta induces clustering of HER2 and integrins by activating Src-focal adhesion kinase and receptor association to the cytoskeleton. Cancer Res. 2009, 69, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, S.; Kersch, C.; Beeson, K.A.; Wu, Y.J.; Muldoon, L.L.; Neuwelt, E.A. Interactions between alphav-Integrin and HER2 and Their Role in the Invasive Phenotype of Breast Cancer Cells In Vitro and in Rat Brain. PLoS ONE 2015, 10, e0131842. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, F.; Han, M.K.L.; Dahmke, I.N.; Del Campo, A.; de Jonge, N. Anti-correlation of HER2 and focal adhesion complexes in the plasma membrane. PLoS ONE 2020, 15, e0234430. [Google Scholar] [CrossRef]
- Griffiths, G.S.; Grundl, M.; Leychenko, A.; Caliva, M.J.; Ramos, J.W.; Matter, M.L. Bit-1 Mediates Integrin-dependent Cell Survival through Activation of the NFκB Pathway. Cell Biol. 2011, 286, 14713–14723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachon, P.H. Integrin Signaling, Cell Survival, and Anoikis: Distinctions, Differences, and Differentiation. J. Signal Transduct. 2011, 2011, 738137. [Google Scholar] [CrossRef] [Green Version]
- Buchheit, C.L.; Angarola, B.L.; Steiner, A.; Weigel, K.J.; Schafer, Z.T. Anoikis evasion in inflammatory breast cancer cells is mediated by Bim-EL sequestration. Cell Death Differ. 2015, 22, 1275–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, V.U.; Rahman, A. Handbook of Natural Product Data, Pentacyclic Triterpenoids; Elsevier: Karachi, Pakistan, 1994. [Google Scholar]
- Chen, M.; Zhong, Z.; Tan, W.; Wang, S.; Wang, Y. Recent advances in nanoparticle formulation of oleanolic acid. Chin. Med. 2011, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K.G.; Tucker, D.J. The separation of substituted Olean-12-en-28-oic acids from the corresponding Urs-12-en-28-oic acid isomers. Aust. J. Chem. 1983, 36, 2297–2305. [Google Scholar] [CrossRef]
- van der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Czaja, K.; Kujawski, J.; Kamel, K.; Bernard, M.K. Selected arylsulphonyl pyrazole derivatives as potential Chk1 kinase ligands—computational investigations. J. Mol. Model. 2020, 26, 144. [Google Scholar] [CrossRef]
- Konieczna, N.; Romaniuk-Drapała, A.; Lisiak, N.; Totoń, E.; Paszel-Jaworska, A.; Kaczmarek, M.; Rubiś, B. Telomerase Inhibitor TMPyP4 Alters Adhesion and Migration of Breast-Cancer Cells MCF7 and MDA-MB-231. Int. J. Mol. Sci. 2019, 20, 2670. [Google Scholar] [CrossRef] [Green Version]
- Rubis, B.; Holysz, H.; Gladych, M.; Toton, E.; Paszel, A.; Lisiak, N.; Kaczmarek, M.; Hofmann, J.; Rybczynska, M. Telomerase downregulation induces proapoptotic genes expression and initializes breast cancer cells apoptosis followed by DNA fragmentation in a cell type dependent manner. Mol. Biol. Rep. 2013, 40, 4995–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toton, E.; Lisiak, N.; Rubis, B.; Budzianowski, J.; Gruber, P.; Hofmann, J.; Rybczynska, M. The tetramethoxyflavone zapotin selectively activates protein kinase C epsilon, leading to its down-modulation accompanied by Bcl-2, c-Jun and c-Fos decrease. Eur. J. Pharmacol. 2012, 682, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Rubis, B.; Krozowski, Z.; Trzeciak, W.H. Arginine vasopressin stimulates 11-beta-hydroxysteroid dehydrogenase type 2 expression in the mineralocorticosteroid target cells. Mol. Cell Endocrinol. 2006, 256, 17–22. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Time | IC50 [µM] | ||
|---|---|---|---|
| OA | HIMOXOL | Br-HIMOLID | |
| 24 h | 22.5 ± 1.27 | 3.89 ± 0.28 | 3.87 ± 0.35 |
| 72 h | 11.31 ± 3.26 | 3.67 ± 0.25 | 2.56 ± 0.36 |
| IC50 | [µM]/24 h | ||
|---|---|---|---|
| OA | HIMOXOL | Br-HIMOLID | |
| 0.5× IC50 | 11.25 | 1.95 | 1.94 |
| 1× IC50 | 22.5 | 3.89 | 3.87 |
| 1.5× IC50 | 33.75 | 5.84 | 5.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisiak, N.M.; Lewicka, I.; Kaczmarek, M.; Kujawski, J.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rubis, B. Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 11273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011273
Lisiak NM, Lewicka I, Kaczmarek M, Kujawski J, Bednarczyk-Cwynar B, Zaprutko L, Rubis B. Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro. International Journal of Molecular Sciences. 2021; 22(20):11273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011273
Chicago/Turabian StyleLisiak, Natalia Magdalena, Izabela Lewicka, Mariusz Kaczmarek, Jacek Kujawski, Barbara Bednarczyk-Cwynar, Lucjusz Zaprutko, and Blazej Rubis. 2021. "Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro" International Journal of Molecular Sciences 22, no. 20: 11273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011273